
SLAM Family Receptors in B Cell Chronic Lymphoproliferative Disorders
 ,
,
Abstract
:1. Introduction
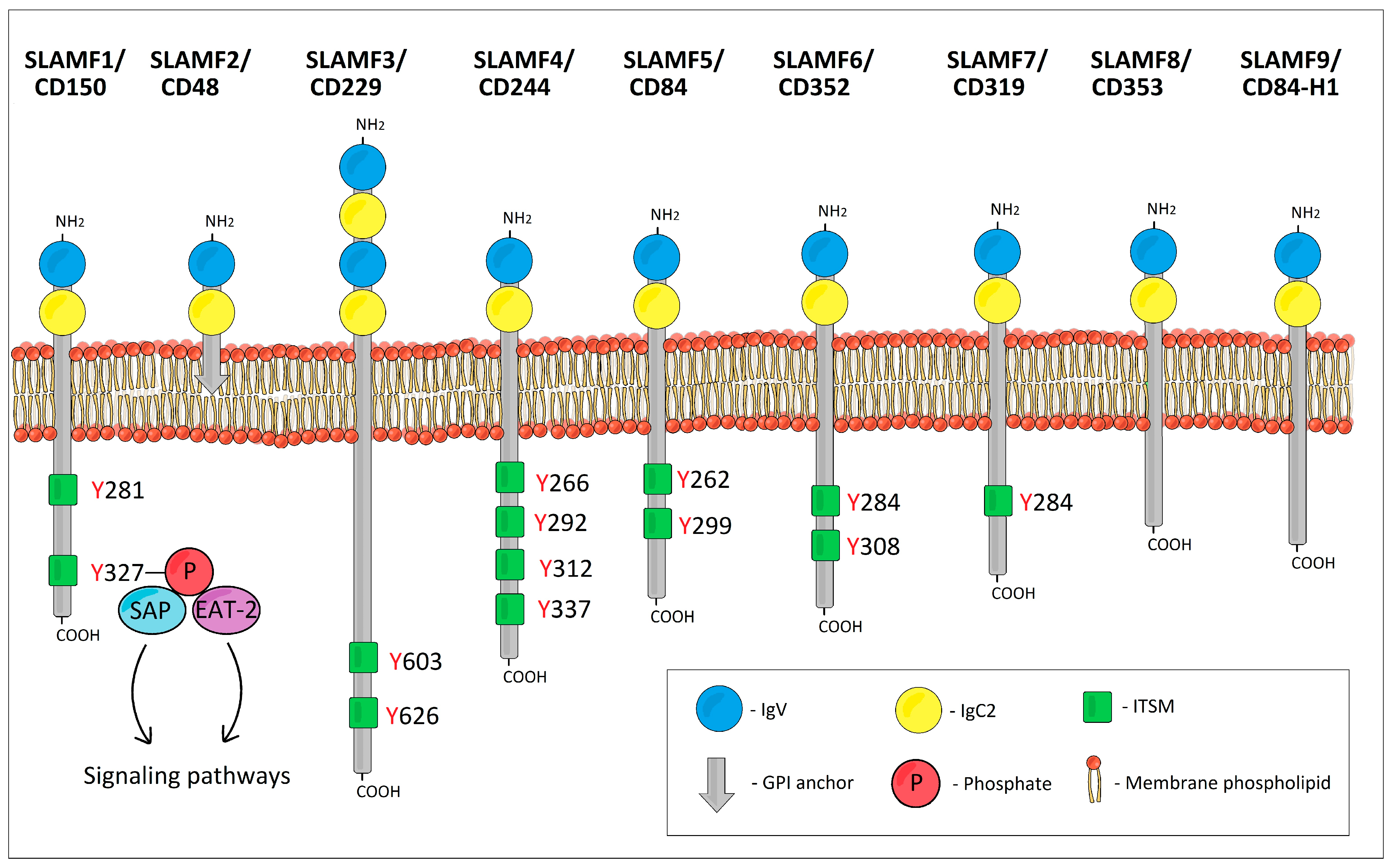
2. SLAMF1/CD150 (SLAM, IPO-3)
3. SLAMF2/CD48 (BCM1, Blast-1, HuLy-m3)
4. SLAMF3/CD229 (Ly-9)
5. SLAMF4/CD244 (2B4, NAIL)
6. SLAMF5/CD84 (Ly9b, GR6)
7. SLAMF6/CD352 (NTB-A, Ly108, SF2000, TCOM)
8. SLAMF7/CD319 (CS1, CRACC, 19A24)
9. SLAMF8/CD353 (BLAME, SBBI42)
10. SLAMF9/CD84-H1 (SF2001, CD2F10)
11. SAP–SLAM-Associated Protein (SH2D1A)
12. EAT-2—Ewing’s Sarcoma Transcript-2 (EWS/FLI1-Activated Transcript 2, SH2D1B)
13. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cannons, J.L.; Tangye, S.G.; Schwartzberg, P.L. SLAM Family Receptors and SAP Adaptors in Immunity. Annu. Rev. Immunol. 2011, 29, 665–705. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.J.; Van Der Merwe, P.A. The Structure and Ligand Interactions of CD2: Implications for T-Cell Function. Immunol. Today 1996, 17, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Sandrin, M.S.; Gumley, T.P.; Henning, M.M.; Vaughan, H.A.; Gonez, L.J.; Trapani, J.A.; McKenzie, I.F. Isolation and Characterization of CDNA Clones for Mouse Ly-9. J. Immunol. 1992, 149, 1636–1641. [Google Scholar] [CrossRef] [PubMed]
- Sandrin, M.S.; Henning, M.M.; Lo, M.F.; Baker, E.; Sutherland, G.R.; McKenzie, I.F.C. Isolation and Characterization of CDNA Clones for Humly9: The Human Homologue of Mouse Ly9. Immunogenetics 1995, 43, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Veillette, A. SLAM-Family Receptors: Immune Regulators with or without SAP-Family Adaptors. Cold Spring Harb. Perspect. Biol. 2010, 2, a002469. [Google Scholar] [CrossRef] [PubMed]
- Sayos, J.; Wu, C.; Morra, M.; Wang, N.; Zhang, X.; Allen, D.; Van Schaik, S.; Notarangelo, L.; Geha, R.; Roncarolo, M.G.; et al. The X-Linked Lymphoproliferative-Disease Gene Product SAP Regulates Signals Induced through the Co-Receptor SLAM. Nature 1998, 395, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Morra, M.; Lu, J.; Poy, F.; Martin, M.; Sayos, J.; Calpe, S.; Gullo, C.; Howie, D.; Rietdijk, S.; Thompson, A.; et al. Structural Basis for the Interaction of the Free SH2 Domain EAT-2 with SLAM Receptors in Hematopoietic Cells. EMBO J. 2001, 20, 5840–5852. [Google Scholar] [CrossRef]
- Sidorenko, S.P.; Clark, E.A. The Dual-Function CD150 Receptor Subfamily: The Viral Attraction. Nat. Immunol. 2003, 4, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Elishmereni, M.; Levi-Schaffer, F. CD48: A Co-Stimulatory Receptor of Immunity. Int. J. Biochem. Cell Biol. 2011, 43, 25–28. [Google Scholar] [CrossRef]
- Staunton, D.E.; Fisher, R.C.; LeBeau, M.M.; Lawrence, J.B.; Barton, D.E.; Francke, U.; Dustin, M.; Thorley-Lawson, D.A. Blast-1 Possesses a Glycosyl-Phosphatidylinositol (GPI) Membrane Anchor, Is Related to LFA-3 and OX-45, and Maps to Chromosome 1q21-23. J. Exp. Med. 1989, 169, 1087–1099. [Google Scholar] [CrossRef]
- Kingsbury, G.A.; Feeney, L.A.; Nong, Y.; Calandra, S.A.; Murphy, C.J.; Corcoran, J.M.; Wang, Y.; Prabhu Das, M.R.; Busfield, S.J.; Fraser, C.C.; et al. Cloning, Expression, and Function of BLAME, a Novel Member of the CD2 Family. J. Immunol. 2001, 166, 5675–5680. [Google Scholar] [CrossRef] [PubMed]
- Fraser, C.C.; Howie, D.; Morra, M.; Qiu, Y.; Murphy, C.; Shen, Q.; Gutierrez-Ramos, J.C.; Coyle, A.; Kingsbury, G.A.; Terhorst, C. Identification and Characterization of SF2000 and SF2001, Two New Members of the Immune Receptor SLAM/CD2 Family. Immunogenetics 2002, 53, 843–850. [Google Scholar] [PubMed]
- Mavaddat, N.; Mason, D.W.; Atkinson, P.D.; Evans, E.J.; Gilbert, R.J.C.; Stuart, D.I.; Fennelly, J.A.; Barclay, A.N.; Davis, S.J.; Brown, M.H. Signaling Lymphocytic Activation Molecule (CDw150) Is Homophilic but Self-Associates with Very Low Affinity. J. Biol. Chem. 2000, 275, 28100–28109. [Google Scholar] [CrossRef] [PubMed]
- Romero, X.; Zapater, N.; Calvo, M.; Kalko, S.G.; de la Fuente, M.A.; Tovar, V.; Ockeloen, C.; Pizcueta, P.; Engel, P. CD229 (Ly9) Lymphocyte Cell Surface Receptor Interacts Homophilically through Its N-Terminal Domain and Relocalizes to the Immunological Synapse. J. Immunol. 2005, 174, 7033–7042. [Google Scholar] [CrossRef] [PubMed]
- Cao, E.; Ramagopal, U.A.; Fedorov, A.; Fedorov, E.; Yan, Q.; Lary, J.W.; Cole, J.L.; Nathenson, S.G.; Almo, S.C. NTB-A Receptor Crystal Structure: Insights into Homophilic Interactions in the Signaling Lymphocytic Activation Molecule Receptor Family. Immunity 2006, 25, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Velikovsky, C.A.; Deng, L.; Chlewicki, L.K.; Fernández, M.M.; Kumar, V.; Mariuzza, R.A. Structure of Natural Killer Receptor 2B4 Bound to CD48 Reveals Basis for Heterophilic Recognition in Signaling Lymphocyte Activation Molecule Family. Immunity 2007, 27, 572–584. [Google Scholar] [CrossRef]
- Yan, Q.; Malashkevich, V.N.; Fedorov, A.; Fedorov, E.; Cao, E.; Lary, J.W.; Cole, J.L.; Nathenson, S.G.; Almo, S.C. Structure of CD84 Provides Insight into SLAM Family Function. Proc. Natl. Acad. Sci. USA 2007, 104, 10583–10588. [Google Scholar] [CrossRef]
- Dong, Z.; Davidson, D.; Pérez-Quintero, L.A.; Kurosaki, T.; Swat, W.; Veillette, A. The Adaptor SAP Controls NK Cell Activation by Regulating the Enzymes Vav-1 and SHIP-1 and by Enhancing Conjugates with Target Cells. Immunity 2012, 36, 974–985. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.J.; Garner, L.I.; Metcalfe, C.; King, E.; Margraf, S.; Brown, M.H. Fine Specificity and Molecular Competition in SLAM Family Receptor Signalling. PLoS ONE 2014, 9, e92184. [Google Scholar] [CrossRef]
- Veillette, A. Immune Regulation by SLAM Family Receptors and SAP-Related Adaptors. Nat. Rev. Immunol. 2006, 6, 56–66. [Google Scholar] [CrossRef]
- Wu, N.; Veillette, A. SLAM Family Receptors in Normal Immunity and Immune Pathologies. Curr. Opin. Immunol. 2016, 38, 45–51. [Google Scholar] [CrossRef]
- Mikhalap, S.V.; Shlapatska, L.M.; Yurchenko, O.V.; Yurchenko, M.Y.; Berdova, G.G.; Nichols, K.E.; Clark, E.A.; Sidorenko, S.P. The Adaptor Protein SH2D1A Regulates Signaling through CD150 (SLAM) in B Cells. Blood 2004, 104, 4063–4070. [Google Scholar] [CrossRef] [PubMed]
- Romero, X.; Benítez, D.; March, S.; Vilella, R.; Miralpeix, M.; Engel, P. Differential Expression of SAP and EAT-2-Binding Leukocyte Cell-Surface Molecules CD84, CD150 (SLAM), CD229 (Ly9) and CD244 (2B4). Tissue Antigens 2004, 64, 132–144. [Google Scholar] [CrossRef]
- De Salort, J.; Sintes, J.; Llinàs, L.; Matesanz-Isabel, J.; Engel, P. Expression of SLAM (CD150) Cell-Surface Receptors on Human B-Cell Subsets: From pro-B to Plasma Cells. Immunol. Lett. 2011, 134, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Llinàs, L.; Lázaro, A.; de Salort, J.; Matesanz-Isabel, J.; Sintes, J.; Engel, P. Expression Profiles of Novel Cell Surface Molecules on B-Cell Subsets and Plasma Cells as Analyzed by Flow Cytometry. Immunol. Lett. 2011, 134, 113–121. [Google Scholar] [CrossRef]
- Rodríguez-Bayona, B.; Ramos-Amaya, A.; Brieva, J.A. Differential Expression of SLAMS and Other Modulatory Molecules by Human Plasma Cells during Normal Maturation. Immunol. Lett. 2011, 134, 122–128. [Google Scholar] [CrossRef]
- Matesanz-Isabel, J.; Sintes, J.; Llinàs, L.; de Salort, J.; Lázaro, A.; Engel, P. New B-Cell CD Molecules. Immunol. Lett. 2011, 134, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.J.; Clare, S.; Mikulin, J.; Johnson, C.M.; Harcourt, K.; Lyons, P.A.; Dougan, G.; Smith, K.G.C. Signalling Lymphocyte Activation Molecule Family Member 9 Is Found on Select Subsets of Antigen-Presenting Cells and Promotes Resistance to Salmonella Infection. Immunology 2020, 159, 393–403. [Google Scholar] [CrossRef]
- Coffey, A.J.; Brooksbank, R.A.; Brandau, O.; Oohashi, T.; Howell, G.R.; Bye, J.M.; Cahn, A.P.; Durham, J.; Heath, P.; Wray, P.; et al. Host Response to EBV Infection in X-Linked Lymphoproliferative Disease Results from Mutations in an SH2-Domain Encoding Gene. Nat. Genet. 1998, 20, 129–135. [Google Scholar] [CrossRef]
- Nichols, K.E.; Harkin, D.P.; Levitz, S.; Krainer, M.; Kolquist, K.A.; Genovese, C.; Bernard, A.; Ferguson, M.; Zuo, L.; Snyder, E.; et al. Inactivating Mutations in an SH2 Domain-Encoding Gene in X-Linked Lymphoproliferative Syndrome. Proc. Natl. Acad. Sci. USA 1998, 95, 13765–13770. [Google Scholar] [CrossRef]
- Huniadi, M.; Salanci, S.; Sykora, J.; Kurhajec, S.; Sarissky, M. Optimisation of Flow Cytometric Detection of Intracellular SLAMF Receptor-Associated Adaptor Proteins SAP and EAT-2. Eur. Pharm. J. 2022, 69, 87–89. [Google Scholar] [CrossRef]
- Shlapatska, L.M.; Mikhalap, S.V.; Berdova, A.G.; Zelensky, O.M.; Yun, T.J.; Nichols, K.E.; Clark, E.A.; Sidorenko, S.P. CD150 Association with Either the SH2-Containing Inositol Phosphatase or the SH2-Containing Protein Tyrosine Phosphatase Is Regulated by the Adaptor Protein SH2D1A. J. Immunol. 2001, 166, 5480–5487. [Google Scholar] [CrossRef]
- Nagy, N.; Cerboni, C.; Mattsson, K.; Maeda, A.; Gogolák, P.; Sümegi, J.; Lányi, Á.; Székely, L.; Carbone, E.; Klein, G.; et al. SH2D1A and slam protein expression in human lymphocytes and derived cell lines. J. Cancer 2000, 88, 439–447. [Google Scholar] [CrossRef]
- Yigit, B.; Halibozek, P.J.; Chen, S.S.; O’Keeffe, M.S.; Arnason, J.; Avigan, D.; Gattei, V.; Bhan, A.; Cen, O.; Longnecker, R.; et al. A Combination of an Anti-SLAMF6 Antibody and Ibrutinib Efficiently Abrogates Expansion of Chronic Lymphocytic Leukemia Cells. Oncotarget 2016, 7, 26346–26360. [Google Scholar] [CrossRef]
- Bologna, C.; Buonincontri, R.; Serra, S.; Vaisitti, T.; Audrito, V.; Brusa, D.; Pagnani, A.; Coscia, M.; D’Arena, G.; Mereu, E.; et al. SLAMF1 Regulation of Chemotaxis and Autophagy Determines CLL Patient Response. J. Clin. Investig. 2016, 126, 181–194. [Google Scholar] [CrossRef]
- Coma, M.; Tothova, E.; Guman, T.; Hajikova, M.; Giertlova, M.; Sarissky, M. Altered Expression Pattern of SLAM Family Receptors on Pathological B Cells of Patients with Chronic Lymphocytic Leukemia. Leuk. Lymphoma 2017, 58, 1726–1729. [Google Scholar] [CrossRef] [PubMed]
- Rigolin, G.M.; Saccenti, E.; Melandri, A.; Cavallari, M.; Urso, A.; Rotondo, F.; Betulla, A.; Tognolo, L.; Bardi, M.A.; Rossini, M.; et al. In Chronic Lymphocytic Leukaemia, SLAMF1 Deregulation Is Associated with Genomic Complexity and Independently Predicts a Worse Outcome. Br. J. Haematol. 2021, 192, 1068–1072. [Google Scholar] [CrossRef]
- von Wenserski, L.; Schultheiß, C.; Bolz, S.; Schliffke, S.; Simnica, D.; Willscher, E.; Gerull, H.; Wolters-Eisfeld, G.; Riecken, K.; Fehse, B.; et al. SLAMF Receptors Negatively Regulate B Cell Receptor Signaling in Chronic Lymphocytic Leukemia via Recruitment of Prohibitin-2. Leukemia 2020, 35, 1073–1086. [Google Scholar] [CrossRef] [PubMed]
- Gordiienko, I.; Scherbina, V.; Shlapatska, L. Soluble CD150 Isoform Level in Plasma of Chronic Lymphocytic Leukemia Patients. Exp. Oncol. 2024, 45, 457–462. [Google Scholar] [CrossRef]
- Sidorenko, S.P.; Větrova, E.P.; Yurchenko, O.V.; Berdova, A.G.; Shlapatskaya, L.N.; Gluzman, D.F.; Kavetsky, R.E. Monoclonal Antibodies of IPO Series against B Cell Differentiation Antigens in Leukemia and Lymphoma Immunophenotyping. Neoplasma 1992, 39, 3–9. [Google Scholar]
- Takeda, S.; Kanbayashi, D.; Kurata, T.; Yoshiyama, H.; Komano, J. Enhanced Susceptibility of B Lymphoma Cells to Measles Virus by Epstein-Barr Virus Type III Latency That Upregulates CD150/Signaling Lymphocytic Activation Molecule. Cancer Sci. 2014, 105, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Ko, Y.H. LMP1+SLAMF1high Cells Are Associated with Drug Resistance in Epstein-Barr Virus-Positive Farage Cells. Oncotarget 2017, 8, 24621. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Kim, E.K.; Ko, Y.H. SLAMF1 Contributes to Cell Survival through the AKT Signaling Pathway in Farage Cells. PLoS ONE 2020, 15, e0238791. [Google Scholar] [CrossRef] [PubMed]
- Fanoni, D.; Tavecchio, S.; Recalcati, S.; Balice, Y.; Venegoni, L.; Fiorani, R.; Crosti, C.; Berti, E. New Monoclonal Antibodies against B-Cell Antigens: Possible New Strategies for Diagnosis of Primary Cutaneous B-Cell Lymphomas. Immunol. Lett. 2011, 134, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Thorley-Lawson, D.A.; Schooley, R.T.; Bhan, A.K.; Nadler, L.M. Epstein-Barr Virus Superinduces a New Human B Cell Differentiation Antigen (B-LAST 1) Expressed on Transformed Lymphoblasts. Cell 1982, 30, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Guan, Q.; Han, X.; Liu, X.; Li, L.; Qiu, L.; Qian, Z.; Zhou, S.; Wang, X.; Zhang, H. A Novel Prognostic Signature Based on Immune-Related Genes of Diffuse Large B-Cell Lymphoma. Aging 2021, 13, 22947–22962. [Google Scholar] [CrossRef]
- Nakajima, H.; Cella, M.; Langen, H.; Friedlein, A.; Colonna, M. Activating Interactions in Human NK Cell Recognition: The Role of 2B4-CD48. Eur. J. Immunol. 1999, 29, 1676–1683. [Google Scholar] [CrossRef]
- Mou, J.; Xie, L.; Xu, Y.; Zhou, T.; Liu, Y.; Huang, Q.; Tang, K.; Tian, Z.; Xing, H.; Qiu, S.; et al. 2B4 Inhibits the Apoptosis of Natural Killer Cells through Phosphorylated Extracellular Signal-Related Kinase/B-Cell Lymphoma 2 Signal Pathway. Cytotherapy 2023, 25, 1080–1090. [Google Scholar] [CrossRef] [PubMed]
- Greenaway, S.; Henniker, A.J.; Walsh, M.; Bradstock, K.F. A Pilot Clinical Trial of Two Murine Monoclonal Antibodies Fixing Human Complement in Patients with Chronic Lymphatic Leukaemia. Leuk. Lymphoma 1994, 13, 323–331. [Google Scholar] [CrossRef]
- Sun, H.; Biggs, J.C.; Smith, G.M. Antitumour Activity of a Chimeric Antibody against the Leucocyte Antigen CD48. Cancer Immunol. Immunother. 2000, 48, 595–602. [Google Scholar] [CrossRef]
- Hosen, N.; Ichihara, H.; Mugitani, A.; Aoyama, Y.; Fukuda, Y.; Kishida, S.; Matsuoka, Y.; Nakajima, H.; Kawakami, M.; Yamagami, T.; et al. CD48 as a Novel Molecular Target for Antibody Therapy in Multiple Myeloma. Br. J. Haematol. 2012, 156, 213–224. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente, M.A.; Tovar, V.; Villamor, N.; Zapater, N.; Pizcueta, P.; Campo, E.; Bosch, J.; Engel, P. Molecular Characterization and Expression of a Novel Human Leukocyte Cell-Surface Marker Homologous to Mouse Ly-9. Blood 2001, 97, 3513–3520. [Google Scholar] [CrossRef] [PubMed]
- Bund, D.; Mayr, C.; Kofler, D.M.; Hallek, M.; Wendtner, C.M. Human Ly9 (CD229) as Novel Tumor-Associated Antigen (TAA) in Chronic Lymphocytic Leukemia (B-CLL) Recognized by Autologous CD8+ T Cells. Exp. Hematol. 2006, 34, 860–869. [Google Scholar] [CrossRef]
- Saberi Hosnijeh, F.; van der Straten, L.; Kater, A.P.; van Oers, M.H.J.; Posthuma, W.F.M.; Chamuleau, M.E.D.; Bellido, M.; Doorduijn, J.K.; van Gelder, M.; Hoogendoorn, M.; et al. Proteomic Markers with Prognostic Impact on Outcome of Chronic Lymphocytic Leukemia Patients under Chemo-Immunotherapy: Results from the HOVON 109 Study. Exp. Hematol. 2020, 89, 55–60.e6. [Google Scholar] [CrossRef] [PubMed]
- Roncador, G.; Puñet-Ortiz, J.; Maestre, L.; Rodríguez-Lobato, L.G.; Jiménez, S.; Reyes-García, A.I.; García-González, Á.; García, J.F.; Piris, M.Á.; Montes-Moreno, S.; et al. CD229 (Ly9) a Novel Biomarker for B-Cell Malignancies and Multiple Myeloma. Cancers 2022, 14, 2154. [Google Scholar] [CrossRef]
- Li, D.; Xiong, W.; Wang, Y.; Feng, J.; He, Y.; Du, J.; Wang, J.; Yang, M.; Zeng, H.; Yang, Y.G.; et al. SLAMF3 and SLAMF4 Are Immune Checkpoints That Constrain Macrophage Phagocytosis of Hematopoietic Tumors. Sci. Immunol. 2022, 7, eabj5501. [Google Scholar]
- Binsky-Ehrenreich, I.; Marom, A.; Sobotta, M.C.; Shvidel, L.; Berrebi, A.; Hazan-Halevy, I.; Kay, S.; Aloshin, A.; Sagi, I.; Goldenberg, D.M.; et al. CD84 Is a Survival Receptor for CLL Cells. Oncogene 2014, 33, 1006–1016. [Google Scholar] [CrossRef]
- Marom, A.; Barak, A.F.; Kramer, M.P.; Lewinsky, H.; Binsky-Ehrenreich, I.; Cohen, S.; Tsitsou-Kampeli, A.; Kalchenko, V.; Kuznetsov, Y.; Mirkin, V.; et al. CD84 Mediates CLL-Microenvironment Interactions. Oncogene 2017, 36, 628–638. [Google Scholar] [CrossRef]
- Lewinsky, H.; Barak, A.F.; Huber, V.; Kramer, M.P.; Radomir, L.; Sever, L.; Orr, I.; Mirkin, V.; Dezorella, N.; Shapiro, M.; et al. CD84 Regulates PD-1/PD-L1 Expression and Function in Chronic Lymphocytic Leukemia. J. Clin. Investig. 2018, 128, 5479–5488. [Google Scholar] [CrossRef]
- Shi, Y.; Ding, W.; Gu, W.; Shen, Y.; Li, H.; Zheng, Z.; Zheng, X.; Liu, Y.; Ling, Y. Single-Cell Phenotypic Profiling to Identify a Set of Immune Cell Protein Biomarkers for Relapsed and Refractory Diffuse Large B Cell Lymphoma: A Single-Center Study. J. Leukoc. Biol. 2022, 112, 1633–1648. [Google Scholar] [CrossRef]
- Korver, W.; Singh, S.; Liu, S.; Zhao, X.; Yonkovich, S.; Sweeney, A.; Anton, K.; Lomas, W.E.; Greenwood, R.; Smith, A.; et al. The Lymphoid Cell Surface Receptor NTB-A: A Novel Monoclonal Antibody Target for Leukaemia and Lymphoma Therapeutics. Br. J. Haematol. 2007, 137, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Yigit, B.; Wang, N.; Hacken, E.T.; Chen, S.S.; Bhan, A.K.; Suarez-Fueyo, A.; Katsuyama, E.; Tsokos, G.C.; Chiorazzi, N.; Wu, C.J.; et al. SLAMF6 as a Regulator of Exhausted CD8+ T Cells in Cancer. Cancer Immunol. Res. 2019, 7, 1485–1496. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Mathew, S.O.; Vaidya, S.V.; Kumaresan, P.R.; Mathew, P.A. CS1 (CRACC, CD319) Induces Proliferation and Autocrine Cytokine Expression on Human B Lymphocytes. J. Immunol. 2007, 179, 4672–4678. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Bodo, J.; Zhao, X.; Durkin, L.; Goyal, T.; Meyerson, H.; Hsi, E.D. SLAMF7 (CD319/CS1) Is Expressed in Plasmablastic Lymphoma and Is a Potential Diagnostic Marker and Therapeutic Target. Br. J. Haematol. 2019, 185, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Panaampon, J.; Kariya, R.; Okada, S. Elotuzumab, a Potential Therapeutic Humanized Anti-SLAMF7 Monoclonal Antibody, Enhances Natural Killer Cell-Mediated Killing of Primary Effusion Lymphoma Cells. Cancer Immunol. Immunother. 2022, 71, 2497–2509. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wan, T.; Li, N.; Yuan, Z.; He, L.; Zhu, X.; Yu, M.; Cao, X. Genetic Approach to Insight into the Immunobiology of Human Dendritic Cells and Identification of CD84-H1, a Novel CD84 Homologue. Clin. Cancer Res. 2001, 7, 822s–829s. [Google Scholar]
- Roncador, G.; Verdes-Montenegro, J.F.G.; Tedoldi, S.; Paterson, J.C.; Klapper, W.; Ballabio, E.; Maestre, L.; Pileri, S.; Hansmann, M.L.; Piris, M.A.; et al. Expression of Two Markers of Germinal Center T Cells (SAP and PD-1) in Angioimmunoblastic T-Cell Lymphoma. Haematologica 2007, 92, 1059–1066. [Google Scholar] [CrossRef]


{kind=link}
| B-CLPD Type: | CLL | SLL | DLBCL | FL | MCL | MZL | LPL | HCL | PBL/ PEL | BL |
|---|---|---|---|---|---|---|---|---|---|---|
| SLAMF1/CD150 | + down | − | + (ABC) − (GC) − (PC, LT) | + (PC) | + | − (PC) | − | + | nr | − or + |
| SLAMF2/CD48 | + down | nr | + + var (PC) | − (PC) | nr | + (PC) | nr | nr | nr | + |
| SLAMF3/CD229 | + up | nr | + | + | + | + | nr | − or + | nr | + |
| SLAMF4/CD244 | − | nr | nr | nr | nr | nr | nr | nr | nr | − |
| SLAMF5/CD84 | + up | nr | + up | nr | nr | nr | nr | nr | nr | + |
| SLAMF6/CD352 | + up | + | + | + | + | nr | nr | nr | nr | + |
| SLAMF7/CD319 | + down | nr | nr | nr | - | nr | nr | nr | + up | + |
| SLAMF8/CD353 | +? | nr | nr | nr | nr | nr | nr | nr | nr | nr |
| SLAMF9/CD84-H1 | nr | nr | nr | nr | nr | nr | nr | nr | nr | + |
| SAP | − | − | + (ABC) + (GC) | − | − | − | − | − | nr | − or + |
| EAT-2 | + | nr | nr | nr | nr | nr | nr | nr | nr | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kľoc, D.; Kurhajec, S.; Huniadi, M.; Sýkora, J.; Guman, T.; Šarišský, M. SLAM Family Receptors in B Cell Chronic Lymphoproliferative Disorders. Int. J. Mol. Sci. 2024, 25, 4014. https://doi.org/10.3390/ijms25074014
Kľoc D, Kurhajec S, Huniadi M, Sýkora J, Guman T, Šarišský M. SLAM Family Receptors in B Cell Chronic Lymphoproliferative Disorders. International Journal of Molecular Sciences. 2024; 25(7):4014. https://doi.org/10.3390/ijms25074014
Chicago/Turabian StyleKľoc, Dominik, Slavomír Kurhajec, Mykhailo Huniadi, Ján Sýkora, Tomáš Guman, and Marek Šarišský. 2024. "SLAM Family Receptors in B Cell Chronic Lymphoproliferative Disorders" International Journal of Molecular Sciences 25, no. 7: 4014. https://doi.org/10.3390/ijms25074014
APA StyleKľoc, D., Kurhajec, S., Huniadi, M., Sýkora, J., Guman, T., & Šarišský, M. (2024). SLAM Family Receptors in B Cell Chronic Lymphoproliferative Disorders. International Journal of Molecular Sciences, 25(7), 4014. https://doi.org/10.3390/ijms25074014