
Ganglioside GD3 Regulates Inflammation and Epithelial-to-Mesenchymal Transition in Human Nasal Epithelial Cells


 and
and
Abstract
:1. Introduction
2. Results
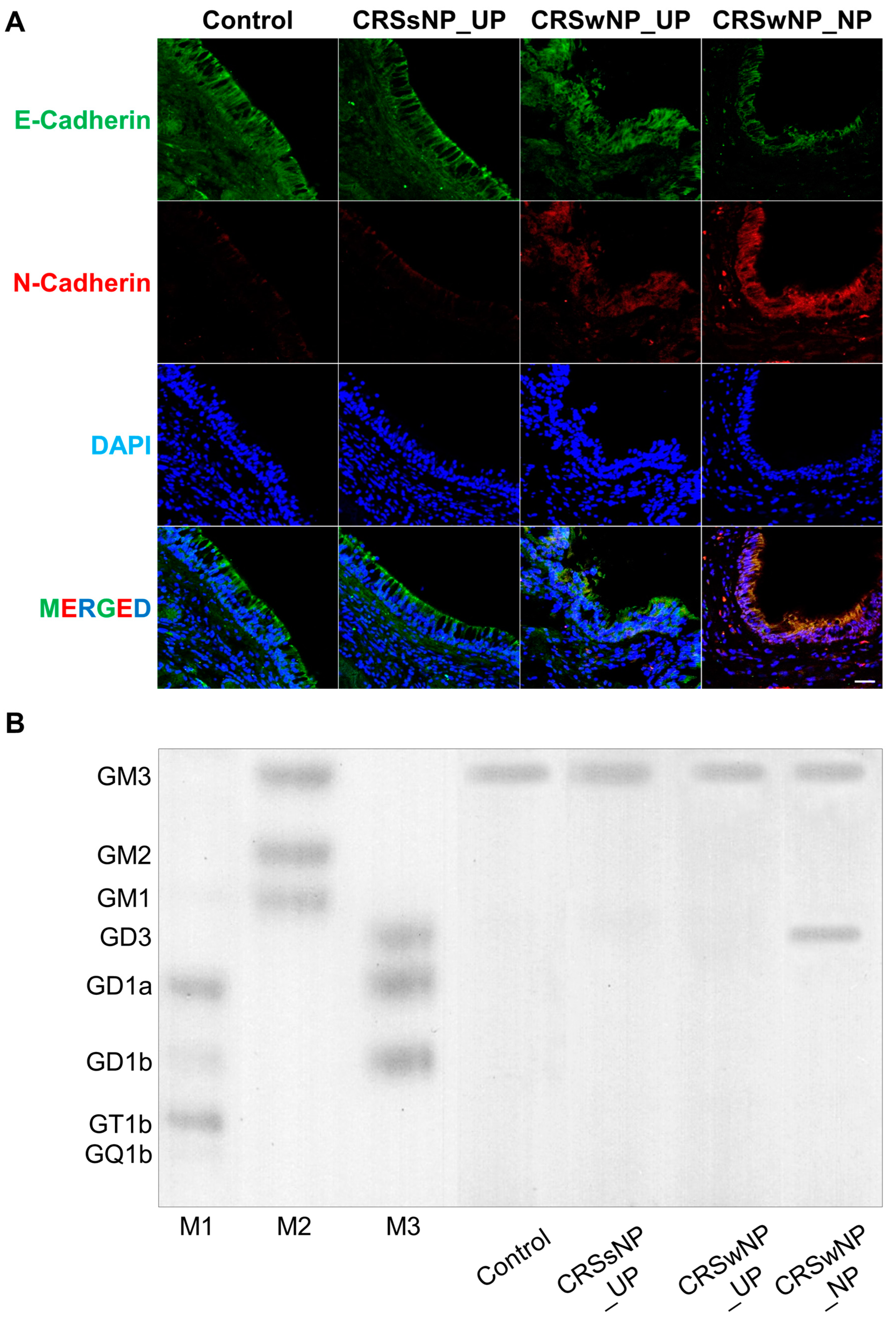
2.1. Analysis of EMT Markers and Ganglioside Expression in Patients with CRS
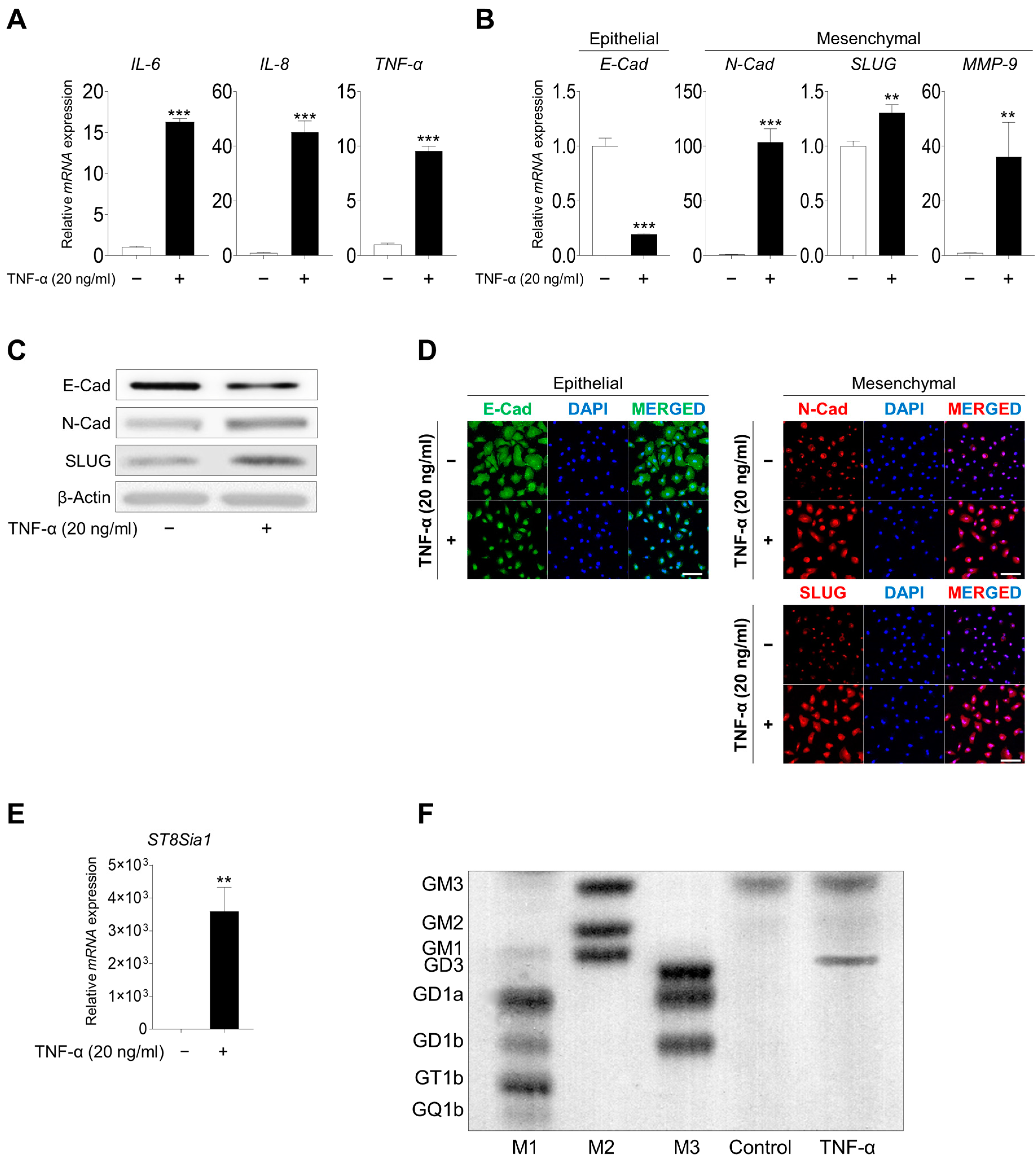
2.2. Inflammation Induced the EMT Process and GD3 Expression upon TNF-α Stimulation
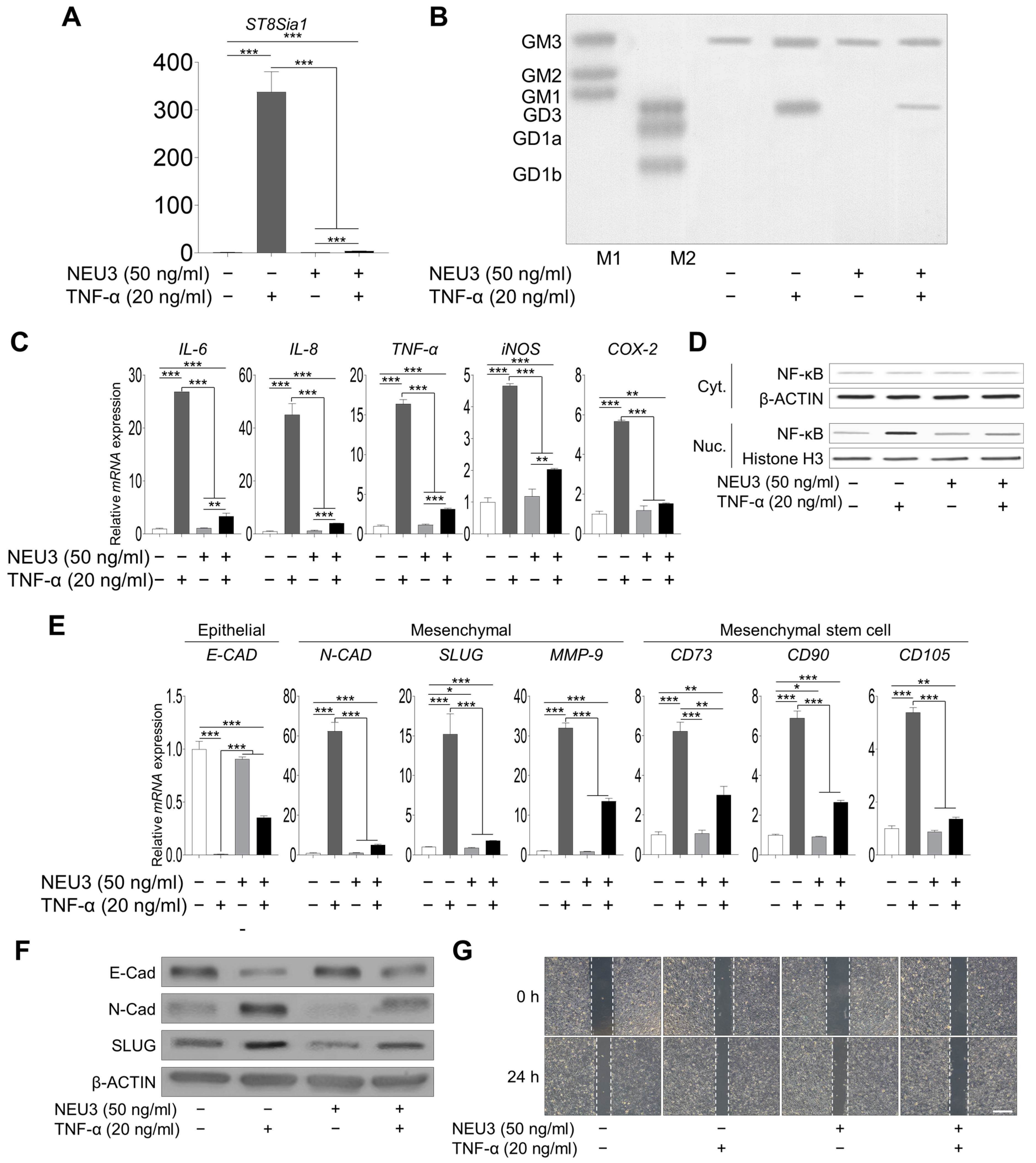
2.3. Direct Correlation between EMT Process and Ganglioside GD3 Expression upon TNF-α Treatment
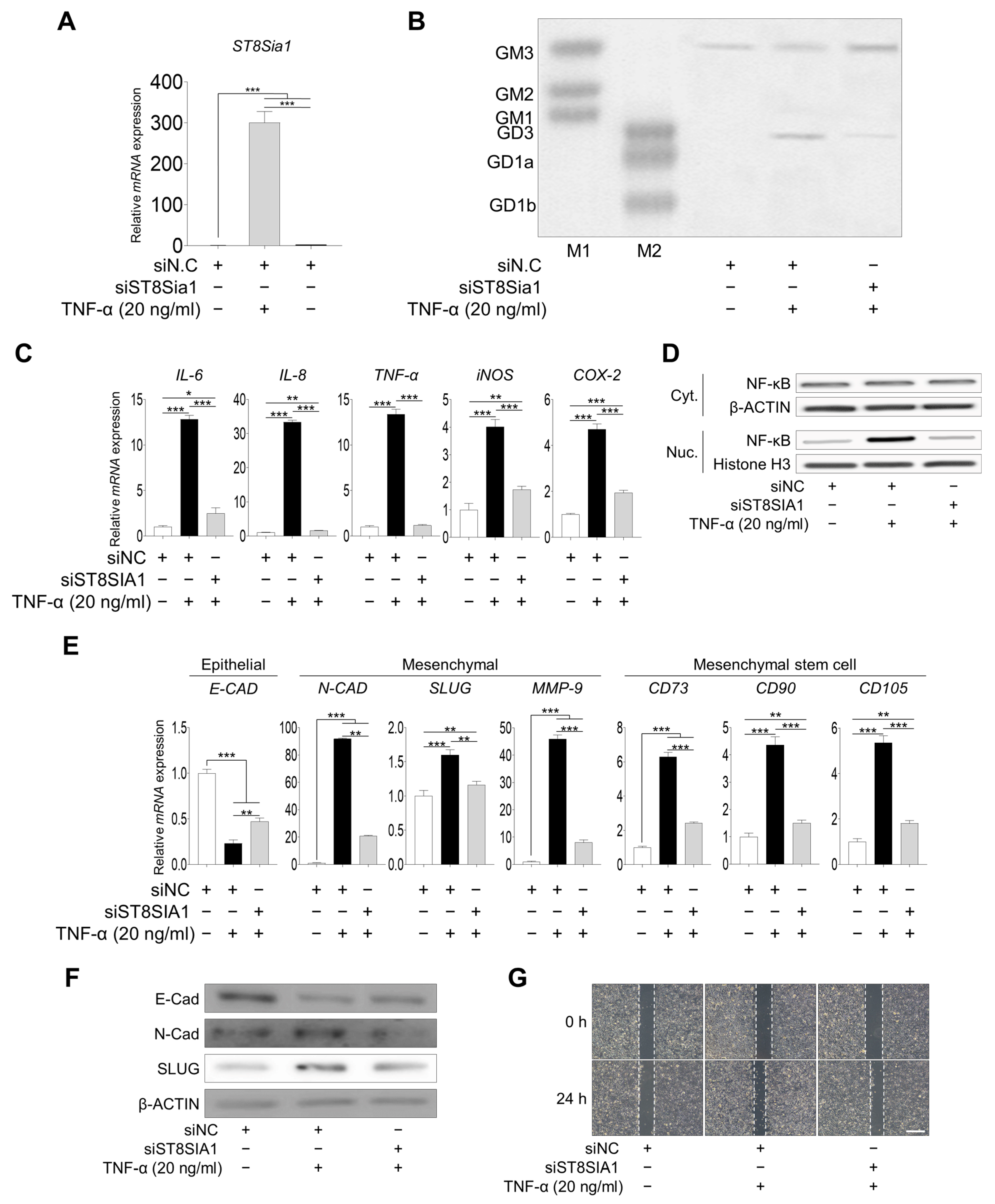
2.4. Inhibition of Ganglioside GD3 Expression Suppressed Inflammation and the EMT Process
3. Discussion
4. Materials and Methods
4.1. Patients and Tissue Samples
4.2. Immunohistochemistry and Immunocytochemistry Analysis
4.3. Extraction and Purification of Gangliosides
4.4. High-Performance Thin-Layer Chromatography
4.5. Culture of Human Nasal Epithelial Cells
4.6. Transfection of Short Interfering RNA
4.7. Total RNA Extraction and Quantitative Real-Time PCR
4.8. Western Blot Analysis
4.9. Wound Healing Assay
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stevens, W.W.; Lee, R.J.; Schleimer, R.P.; Cohen, N.A. Chronic rhinosinusitis pathogenesis. J. Allergy Clin. Immunol. 2015, 136, 1442–1453. [Google Scholar] [CrossRef] [PubMed]
- Hellings, P.W.; Verhoeven, E.; Fokkens, W.J. State-of-the-art overview on biological treatment for CRSwNP. Rhinology 2021, 59, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Fokkens, W.J.; Lund, V.J.; Hopkins, C.; Hellings, P.W.; Kern, R.; Reitsma, S.; Toppila-Salmi, S.; Bernal-Sprekelsen, M.; Mullol, J.; Alobid, I.; et al. European Position Paper on Rhinosinusitis and Nasal Polyps 2020. Rhinology 2020, 58 (Suppl. S29), 1–464. [Google Scholar] [CrossRef] [PubMed]
- Eggesbo, H.B. Imaging of sinonasal tumours. Cancer Imaging 2012, 12, 136–152. [Google Scholar] [CrossRef] [PubMed]
- Ijaz, T.; Pazdrak, K.; Kalita, M.; Konig, R.; Choudhary, S.; Tian, B.; Boldogh, I.; Brasier, A.R. Systems biology approaches to understanding Epithelial Mesenchymal Transition (EMT) in mucosal remodeling and signaling in asthma. World Allergy Organ. J. 2014, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- Konnecke, M.; Burmeister, M.; Pries, R.; Boscke, R.; Bruchhage, K.L.; Ungefroren, H.; Klimek, L.; Wollenberg, B. Epithelial-Mesenchymal Transition in Chronic Rhinosinusitis: Differences Revealed Between Epithelial Cells from Nasal Polyps and Inferior Turbinates. Arch. Immunol. Ther. Exp. 2017, 65, 157–173. [Google Scholar] [CrossRef]
- Hupin, C.; Gohy, S.; Bouzin, C.; Lecocq, M.; Polette, M.; Pilette, C. Features of mesenchymal transition in the airway epithelium from chronic rhinosinusitis. Allergy 2014, 69, 1540–1549. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.P.; Sleeman, J.P. Complex networks orchestrate epithelial-mesenchymal transitions. Nat. Rev. Mol. Cell Biol. 2006, 7, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Ong, Y.K.; Wang, Y. Novel findings in immunopathophysiology of chronic rhinosinusitis and their role in a model of precision medicine. Allergy 2020, 75, 769–780. [Google Scholar] [CrossRef]
- Zavadil, J.; Bottinger, E.P. TGF-beta and epithelial-to-mesenchymal transitions. Oncogene 2005, 24, 5764–5774. [Google Scholar] [CrossRef] [PubMed]
- Ryu, G.; Mo, J.H.; Shin, H.W. Epithelial-to-mesenchymal transition in neutrophilic chronic rhinosinusitis. Curr. Opin. Allergy Clin. Immunol. 2021, 21, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.W.; Cho, K.; Kim, D.W.; Han, D.H.; Khalmuratova, R.; Kim, S.W.; Jeon, S.Y.; Min, Y.G.; Lee, C.H.; Rhee, C.S.; et al. Hypoxia-inducible factor 1 mediates nasal polypogenesis by inducing epithelial-to-mesenchymal transition. Am. J. Respir. Crit. Care Med. 2012, 185, 944–954. [Google Scholar] [CrossRef] [PubMed]
- Huvenne, W.; van Bruaene, N.; Zhang, N.; van Zele, T.; Patou, J.; Gevaert, P.; Claeys, S.; Van Cauwenberge, P.; Bachert, C. Chronic rhinosinusitis with and without nasal polyps: What is the difference? Curr. Allergy Asthma Rep. 2009, 9, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Wang, H.; Yin, J. The Role of Epithelial-Mesenchymal Transition in Chronic Rhinosinusitis. Int. Arch. Allergy Immunol. 2022, 183, 1029–1039. [Google Scholar] [CrossRef] [PubMed]
- Hakomori, S. Bifunctional role of glycosphingolipids. Modulators for transmembrane signaling and mediators for cellular interactions. J. Biol. Chem. 1990, 265, 18713–18716. [Google Scholar] [CrossRef] [PubMed]
- Hakomori, S.; Yamamura, S.; Handa, A.K. Signal transduction through glyco(sphingo)lipids. Introduction and recent studies on glyco(sphingo)lipid-enriched microdomains. Ann. N. Y. Acad. Sci. 1998, 845, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hakomori, S. Glycosphingolipids in cellular interaction, differentiation, and oncogenesis. Annu. Rev. Biochem. 1981, 50, 733–764. [Google Scholar] [CrossRef] [PubMed]
- Hakomori, S. Glycosylation defining cancer malignancy: New wine in an old bottle. Proc. Natl. Acad. Sci. USA 2002, 99, 10231–10233. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.W.; Lee, S.A.; Shin, J.M.; Park, I.H.; Lee, H.M. Glucocorticoids ameliorate TGF-beta1-mediated epithelial-to-mesenchymal transition of airway epithelium through MAPK and Snail/Slug signaling pathways. Sci. Rep. 2017, 7, 3486. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.K.; Macala, L.J.; Taki, T.; Weinfield, H.M.; Yu, F.S. Developmental changes in ganglioside composition and synthesis in embryonic rat brain. J. Neurochem. 1988, 50, 1825–1829. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.K. Development regulation of ganglioside metabolism. Prog. Brain Res. 1994, 101, 31–44. [Google Scholar] [PubMed]
- Kim, S.J.; Chung, T.W.; Choi, H.J.; Kwak, C.H.; Song, K.H.; Suh, S.J.; Kwon, K.M.; Chang, Y.C.; Park, Y.G.; Chang, H.W.; et al. Ganglioside GM3 participates in the TGF-beta1-induced epithelial-mesenchymal transition of human lens epithelial cells. Biochem. J. 2013, 449, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Julien, S.; Bobowski, M.; Steenackers, A.; Le Bourhis, X.; Delannoy, P. How Do Gangliosides Regulate RTKs Signaling? Cells 2013, 2, 751–767. [Google Scholar] [CrossRef] [PubMed]
- Mathow, D.; Chessa, F.; Rabionet, M.; Kaden, S.; Jennemann, R.; Sandhoff, R.; Grone, H.J.; Feuerborn, A. Zeb1 affects epithelial cell adhesion by diverting glycosphingolipid metabolism. EMBO Rep. 2015, 16, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Levade, T.; Andrieu-Abadie, N.; Micheau, O.; Legembre, P.; Segui, B. Sphingolipids modulate the epithelial-mesenchymal transition in cancer. Cell Death Discov. 2015, 1, 15001. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, T.R.; Battula, V.L.; Werden, S.J.; Vijay, G.V.; Ramirez-Pena, E.Q.; Taube, J.H.; Chang, J.T.; Miura, N.; Porter, W.; Sphyris, N.; et al. GD3 synthase regulates epithelial-mesenchymal transition and metastasis in breast cancer. Oncogene 2015, 34, 2958–2967. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, Y.; Izumoto, S.; Suzuki, T.; Kinoshita, M.; Kagawa, N.; Wada, K.; Hashimoto, N.; Maruno, M.; Nakatsuji, Y.; Yoshimine, T. Ganglioside GM3 inhibits proliferation and invasion of glioma. J. Neurooncol 2005, 71, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Dewald, J.H.; Colomb, F.; Bobowski-Gerard, M.; Groux-Degroote, S.; Delannoy, P. Role of Cytokine-Induced Glycosylation Changes in Regulating Cell Interactions and Cell Signaling in Inflammatory Diseases and Cancer. Cells 2016, 5, 43. [Google Scholar] [CrossRef] [PubMed]
- Fokkens, W.J.; Lund, V.J.; Mullol, J.; Bachert, C.; Alobid, I.; Baroody, F.; Cohen, N.; Cervin, A.; Douglas, R.; Gevaert, P.; et al. EPOS 2012: European position paper on rhinosinusitis and nasal polyps 2012. A summary for otorhinolaryngologists. Rhinology 2012, 50, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Schleimer, R.P. Immunopathogenesis of Chronic Rhinosinusitis and Nasal Polyposis. Annu. Rev. Pathol. 2017, 12, 331–357. [Google Scholar] [CrossRef] [PubMed]
- Seyrantepe, V.; Landry, K.; Trudel, S.; Hassan, J.A.; Morales, C.R.; Pshezhetsky, A.V. Neu4, a novel human lysosomal lumen sialidase, confers normal phenotype to sialidosis and galactosialidosis cells. J. Biol. Chem. 2004, 279, 37021–37029. [Google Scholar] [CrossRef] [PubMed]
- Smutova, V.; Albohy, A.; Pan, X.; Korchagina, E.; Miyagi, T.; Bovin, N.; Cairo, C.W.; Pshezhetsky, A.V. Structural basis for substrate specificity of mammalian neuraminidases. PLoS ONE 2014, 9, e106320. [Google Scholar] [CrossRef] [PubMed]
- Kao, S.S.; Bassiouni, A.; Ramezanpour, M.; Finnie, J.; Chegeni, N.; Colella, A.D.; Chataway, T.K.; Wormald, P.J.; Vreugde, S.; Psaltis, A.J. Proteomic analysis of nasal mucus samples of healthy patients and patients with chronic rhinosinusitis. J. Allergy Clin. Immunol. 2021, 147, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef]
- Lopez-Novoa, J.M.; Nieto, M.A. Inflammation and EMT: An alliance towards organ fibrosis and cancer progression. EMBO Mol. Med. 2009, 1, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Zhou, C.; Ma, C.; Cao, Y.; Hu, G.; Li, H. TGF-beta1 induces epithelial-to-mesenchymal transition in chronic rhinosinusitis with nasal polyps through microRNA-182. Asian Pac. J. Allergy Immunol. 2024, 42, 61–73. [Google Scholar] [PubMed]
- You, B.; Zhang, T.; Zhang, W.; Pei, Y.; Huang, D.; Lei, Y.; Zhang, S.; Qiu, C.; Zhang, J.; Gu, Z.; et al. IGFBP2 derived from PO-MSCs promote epithelial barrier destruction by activating FAK signaling in nasal polyps. iScience 2023, 26, 106151. [Google Scholar] [CrossRef] [PubMed]
- Mesuraca, M.; Nistico, C.; Lombardo, N.; Piazzetta, G.L.; Lobello, N.; Chiarella, E. Cellular and Biochemical Characterization of Mesenchymal Stem Cells from Killian Nasal Polyp. Int. J. Mol. Sci. 2022, 23, 13214. [Google Scholar] [CrossRef] [PubMed]
- Guan, F.; Handa, K.; Hakomori, S.I. Specific glycosphingolipids mediate epithelial-to-mesenchymal transition of human and mouse epithelial cell lines. Proc. Natl. Acad. Sci. USA 2009, 106, 7461–7466. [Google Scholar] [CrossRef] [PubMed]
- Dewald, J.H.; Cavdarli, S.; Steenackers, A.; Delannoy, C.P.; Mortuaire, M.; Spriet, C.; Noel, M.; Groux-Degroote, S.; Delannoy, P. TNF differentially regulates ganglioside biosynthesis and expression in breast cancer cell lines. PLoS ONE 2018, 13, e0196369. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, H.S.; Zhou, B.H.; Li, C.L.; Zhang, F.; Wang, X.F.; Zhang, G.; Bu, X.Z.; Cai, S.H.; Du, J. Epithelial-mesenchymal transition (EMT) induced by TNF-alpha requires AKT/GSK-3beta-mediated stabilization of snail in colorectal cancer. PLoS ONE 2013, 8, e56664. [Google Scholar]
- Bradley, J.R. TNF-mediated inflammatory disease. J. Pathol. 2008, 214, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Song, K.S.; Lee, W.J.; Chung, K.C.; Koo, J.S.; Yang, E.J.; Choi, J.Y.; Yoon, J.H. Interleukin-1 beta and tumor necrosis factor-alpha induce MUC5AC overexpression through a mechanism involving ERK/p38 mitogen-activated protein kinases-MSK1-CREB activation in human airway epithelial cells. J. Biol. Chem. 2003, 278, 23243–23250. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.T.; Zhang, Y.; Zhang, P.; Wu, H.; Sun, X.Q.; Shen, S.Y.; Dou, D.B. Tumor necrosis factor-alpha promotes airway mucus hypersecretion by repressing miR-146a-5p and miR-134-5p levels in human airway epithelial cells. Transl. Cancer Res. 2021, 10, 4047–4056. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Kim, K.S.; Kim, H.U.; Linton, J.A.; Lee, J.G. Effects of TNF-alpha and IL-1 beta on mucin, lysozyme, IL-6 and IL-8 in passage-2 normal human nasal epithelial cells. Acta Otolaryngol. 1999, 119, 905–910. [Google Scholar]
- Jiang, Y.; Liu, B.; Bao, X.; Zhou, P.; Li, J. TNF-alpha Regulates the Glucocorticoid Receptor Alpha Expression in Human Nasal Epithelial Cells Via p65-NF-kappab and p38-MAPK Signaling Pathways. Iran. J. Biotechnol. 2023, 21, e3117. [Google Scholar]
- Ryu, J.S.; Ko, K.; Lee, J.W.; Park, S.B.; Byun, S.J.; Jeong, E.J.; Ko, K.; Choo, Y.K. Gangliosides are involved in neural differentiation of human dental pulp-derived stem cells. Biochem. Biophys. Res. Commun. 2009, 387, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Morales, A.; Colell, A.; Mari, M.; Garcia-Ruiz, C.; Fernandez-Checa, J.C. Glycosphingolipids and mitochondria: Role in apoptosis and disease. Glycoconj. J. 2004, 20, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Huang, Z.M.; Wu, J.S.; Chen, X.R.; Jin, S.Q.; Huang, Q.K.; Wang, D. Ganglioside GD3 enhances the anti-tumor effect of doxorubicin on hepatoma cells. Zhonghua Gan Zang Bing Za Zhi 2006, 14, 710–711. [Google Scholar] [PubMed]
- Malisan, F.; Testi, R. The ganglioside GD3 as the Greek goddess Hecate: Several faces turned towards as many directions. IUBMB Life 2005, 57, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Suh, M.; Thomson, A.B.; Ramanujam, K.S.; Clandinin, M.T. Dietary gangliosides increase the content and molecular percentage of ether phospholipids containing 20:4n-6 and 22:6n-3 in weanling rat intestine. J. Nutr. Biochem. 2006, 17, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Rueda, R.; Maldonado, J.; Narbona, E.; Gil, A. Neonatal dietary gangliosides. Early Hum. Dev. 1998, 53 (Suppl. S1), S135–S147. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Arita, Y.; Satomi, N.; Eisinger, M.; Lloyd, K.O. Tumor necrosis factor enhances GD3 ganglioside expression in cultured human melanocytes. Arch. Biochem. Biophys. 1990, 281, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Miyata, M.; Ichihara, M.; Tajima, O.; Sobue, S.; Kambe, M.; Sugiura, K.; Furukawa, K.; Furukawa, K. UVB-irradiated keratinocytes induce melanoma-associated ganglioside GD3 synthase gene in melanocytes via secretion of tumor necrosis factor alpha and interleukin 6. Biochem. Biophys. Res. Commun. 2014, 445, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.Y.; Kim, C.H.; Kim, K.S.; Ko, J.H.; Lee, J.H.; Jeong, Y.K.; Lee, Y.C. Expression of the human CMP-NeuAc:GM3 alpha2,8-sialyltransferase (GD3 synthase) gene through the NF-kappaB activation in human melanoma SK-MEL-2 cells. Biochim. Biophys. Acta 2007, 1769, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.Y.; Dae, H.M.; Song, N.R.; Kim, K.S.; Kim, C.H.; Lee, Y.C. Valproic acid induces transcriptional activation of human GD3 synthase (hST8Sia I) in SK-N-BE(2)-C human neuroblastoma cells. Mol. Cells 2009, 27, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Bobowski, M.; Vincent, A.; Steenackers, A.; Colomb, F.; Van Seuningen, I.; Julien, S.; Delannoy, P. Estradiol represses the G(D3) synthase gene ST8SIA1 expression in human breast cancer cells by preventing NFkappaB binding to ST8SIA1 promoter. PLoS ONE 2013, 8, e62559. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, R.; Kambe, M.; Miyata, M.; Jeyadevan, U.; Tajima, O.; Furukawa, K.; Furukawa, K. TNFalpha-signal and cAMP-mediated signals oppositely regulate melanoma- associated ganglioside GD3 synthase gene in human melanocytes. Sci. Rep. 2019, 9, 14740. [Google Scholar] [CrossRef]
- Monti, E.; Bonten, E.; D’Azzo, A.; Bresciani, R.; Venerando, B.; Borsani, G.; Schauer, R.; Tettamanti, G. Sialidases in vertebrates: A family of enzymes tailored for several cell functions. Adv. Carbohydr. Chem. Biochem. 2010, 64, 403–479. [Google Scholar] [PubMed]
- Vitner, E.B.; Platt, F.M.; Futerman, A.H. Common and uncommon pathogenic cascades in lysosomal storage diseases. J. Biol. Chem. 2010, 285, 20423–20427. [Google Scholar] [CrossRef] [PubMed]
- Bosch, M.E.; Kielian, T. Neuroinflammatory paradigms in lysosomal storage diseases. Front. Neurosci. 2015, 9, 417. [Google Scholar] [CrossRef] [PubMed]
- Lillehoj, E.P.; Hyun, S.W.; Feng, C.; Zhang, L.; Liu, A.; Guang, W.; Nguyen, C.; Sun, W.; Luzina, I.G.; Webb, T.J.; et al. Human airway epithelia express catalytically active NEU3 sialidase. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 306, L876–L886. [Google Scholar] [CrossRef] [PubMed]
- Abdel Hadi, L.; Di Vito, C.; Riboni, L. Fostering Inflammatory Bowel Disease: Sphingolipid Strategies to Join Forces. Mediators Inflamm. 2016, 2016, 3827684. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.K.; Cho, S.H.; Kim, K.W.; Jeon, J.H.; Ko, J.H.; Kim, B.Y.; Kim, C.H. Overexpression of membrane sialic acid-specific sialidase Neu3 inhibits matrix metalloproteinase-9 expression in vascular smooth muscle cells. Biochem. Biophys. Res. Commun. 2007, 356, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Ghiroldi, A.; Piccoli, M.; Creo, P.; Cirillo, F.; Rota, P.; D’Imperio, S.; Ciconte, G.; Monasky, M.M.; Micaglio, E.; Garatti, A.; et al. Role of sialidase Neu3 and ganglioside GM3 in cardiac fibroblasts activation. Biochem. J. 2020, 477, 3401–3415. [Google Scholar] [CrossRef] [PubMed]
- Cumin, C.; Huang, Y.L.; Rossdam, C.; Ruoff, F.; Cespedes, S.P.; Liang, C.Y.; Lombardo, F.C.; Coelho, R.; Rimmer, N.; Konantz, M.; et al. Glycosphingolipids are mediators of cancer plasticity through independent signaling pathways. Cell Rep. 2022, 40, 111181. [Google Scholar] [CrossRef] [PubMed]
- Ladisch, S.; Gillard, B. A solvent partition method for microscale ganglioside purification. Anal. Biochem. 1985, 146, 220–231. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Control Patients | Patients with CRSsNP | Patients with CRSwNP | |
|---|---|---|---|---|
| Subjects (n) | 8 | 8 | 8 | 8 |
| Tissue used | UP | UP | UP | NP |
| Age (y) | 47.5 ± 15.35 | 43.06 ± 16.0 | 46.9 ± 12.52 | |
| Male/Female, n/n | 3/5 | 5/3 | 4/4 | |
| Allergic rhinitis (n) | 0 | 0 | 0 | |
| Aspirin sensitivity (n) | 0 | 0 | 0 | |
| Smoking, n (%) | 1 (12.5%) | 2 (25%) | 2 (25%) | |
| Methodologies used | ||||
| HPTLC | 5 | 5 | 5 | 5 |
| Tissue IHC | 3 | 3 | 3 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, J.H.; Ryu, J.-S.; Yu, J.O.; Choo, Y.-K.; Kang, J.; Kim, J.-Y. Ganglioside GD3 Regulates Inflammation and Epithelial-to-Mesenchymal Transition in Human Nasal Epithelial Cells. Int. J. Mol. Sci. 2024, 25, 4054. https://doi.org/10.3390/ijms25074054
Hwang JH, Ryu J-S, Yu JO, Choo Y-K, Kang J, Kim J-Y. Ganglioside GD3 Regulates Inflammation and Epithelial-to-Mesenchymal Transition in Human Nasal Epithelial Cells. International Journal of Molecular Sciences. 2024; 25(7):4054. https://doi.org/10.3390/ijms25074054
Chicago/Turabian StyleHwang, Ji Hyeon, Jae-Sung Ryu, Jin Ok Yu, Young-Kug Choo, Jaeku Kang, and Jong-Yeup Kim. 2024. "Ganglioside GD3 Regulates Inflammation and Epithelial-to-Mesenchymal Transition in Human Nasal Epithelial Cells" International Journal of Molecular Sciences 25, no. 7: 4054. https://doi.org/10.3390/ijms25074054
APA StyleHwang, J. H., Ryu, J. -S., Yu, J. O., Choo, Y. -K., Kang, J., & Kim, J. -Y. (2024). Ganglioside GD3 Regulates Inflammation and Epithelial-to-Mesenchymal Transition in Human Nasal Epithelial Cells. International Journal of Molecular Sciences, 25(7), 4054. https://doi.org/10.3390/ijms25074054