
The Clinicopathological Significance of the Cyclin D1/E1–Cyclin-Dependent Kinase (CDK2/4/6)–Retinoblastoma (RB1/pRB1) Pathway in Epithelial Ovarian Cancers


 and
and 
Abstract
:1. Introduction
2. Results
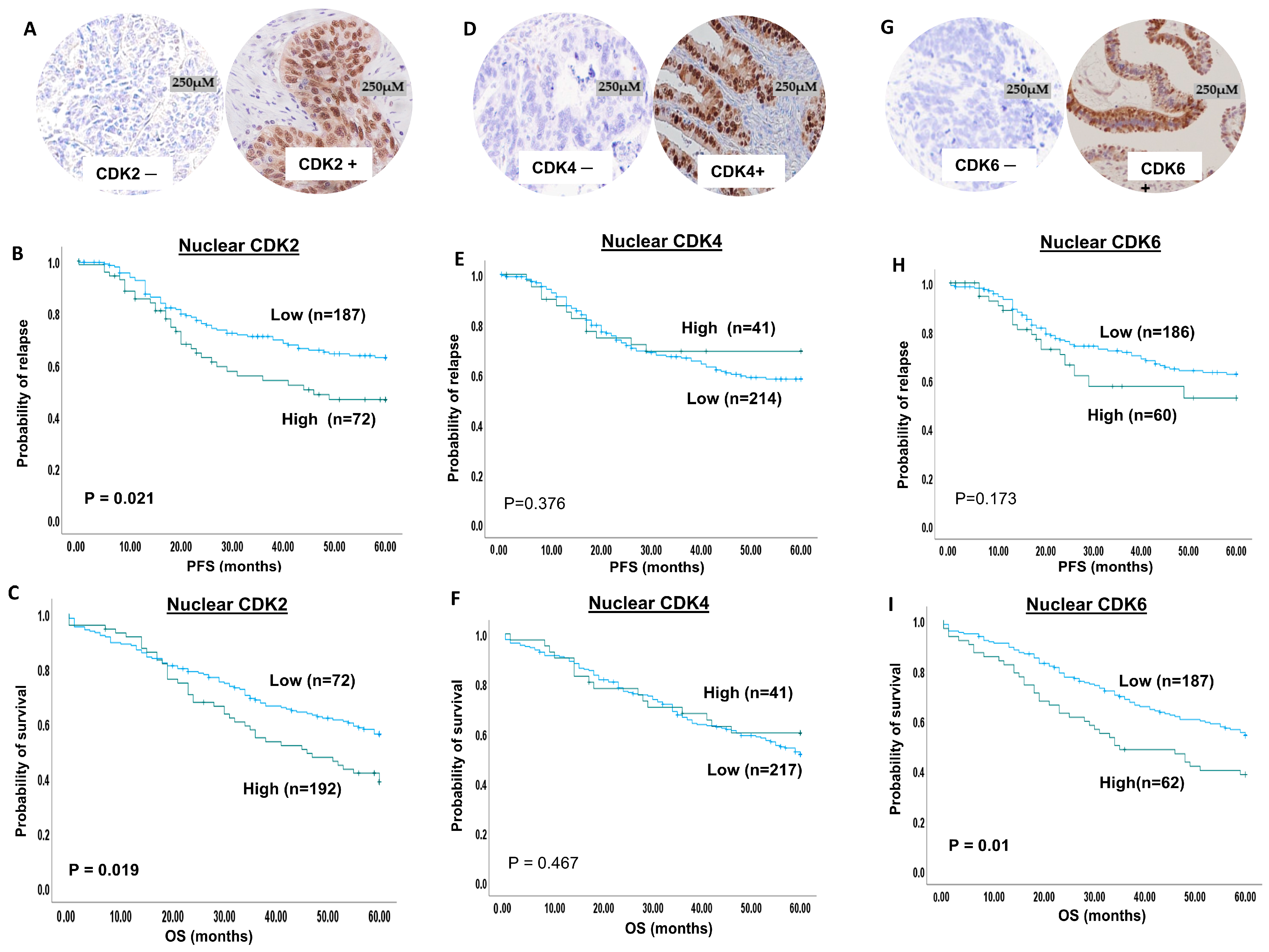
2.1. CDK2 Expression
2.2. CDK4 Expression
2.3. CDK6 Expression
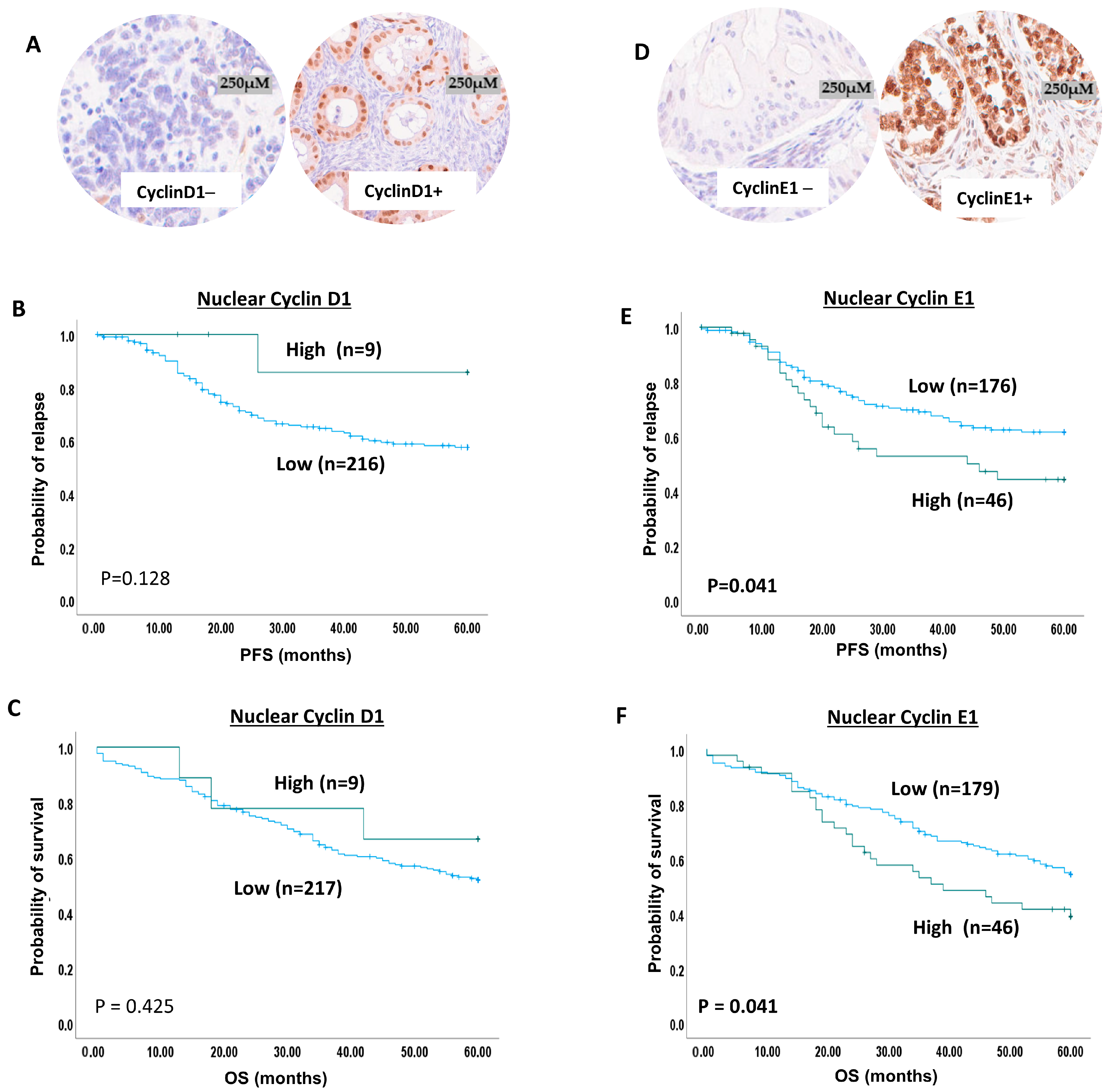
2.4. Cyclin D1 Expression in Ovarian Cancer
2.5. Cyclin E1 Protein Expression in Ovarian Cancer
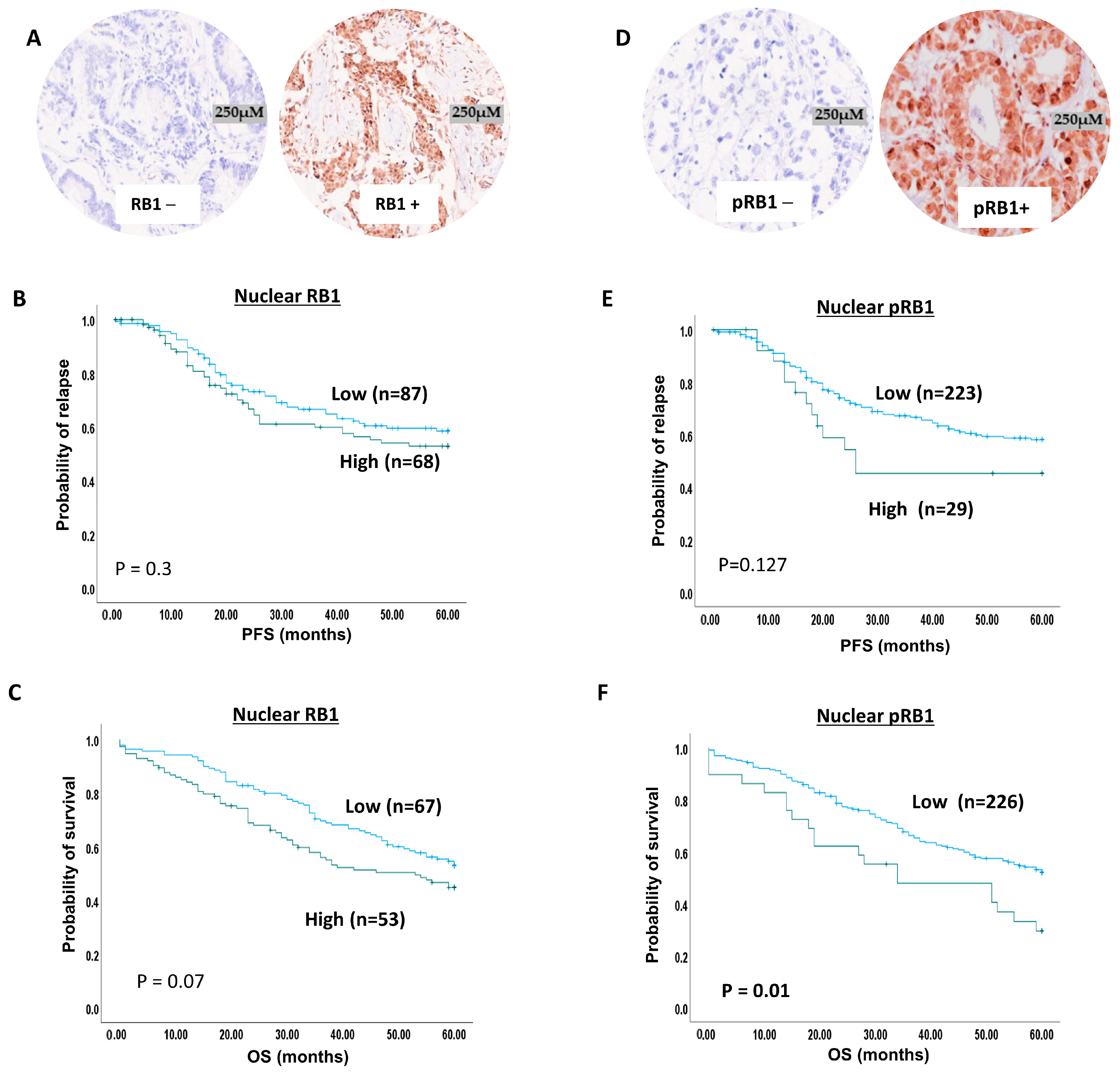
2.6. RB1 Expression and Ovarian Cancer
2.7. pRb1 Expression and Ovarian Cancer
2.8. Multivariate Analysis
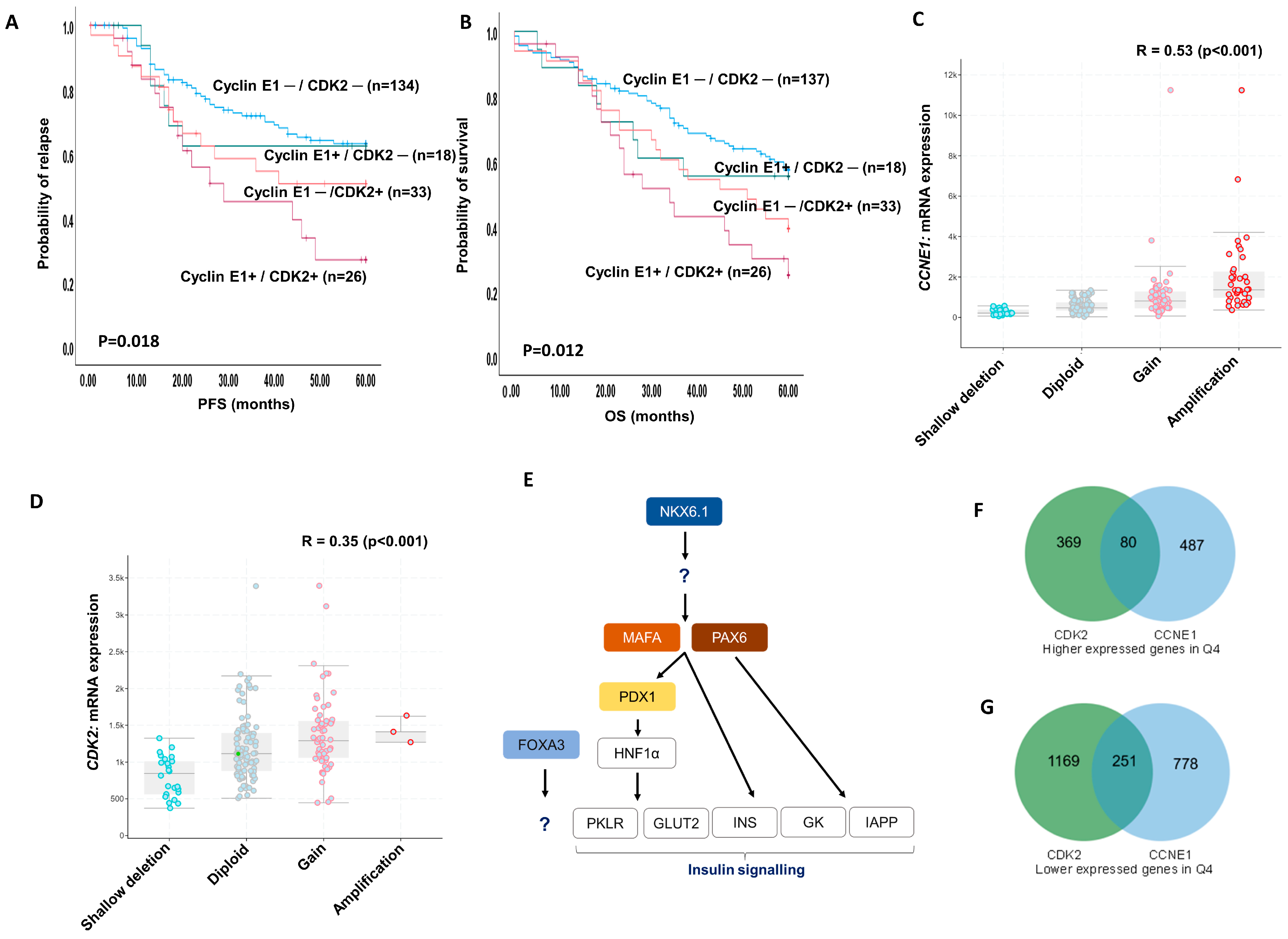
2.9. Cyclin E1 and CDK2 Protein Co-Expression
2.10. Bioinformatics in Ovarian TCGA
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Study Cohort
5.2. Antibodies and Western Blot
5.3. Tissue Microarrays (TMAs) and Immunohistochemistry (IHC)
5.4. Evaluation of the Immune Staining
5.5. CDK4, CDK6, CDK2, Cyclin D1, Cyclin E1 and RB1 Transcripts in Ovarian Cancers
5.6. Statistical Analysis
5.7. Bioinformatics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- CRUK, Ovarian Cancer Statistics. Cancer Research UK. 2017. Available online: https://www.cancerresearchuk.org/health-professional/cancer-statistics/statistics-by-cancer-type/ovarian-cancer (accessed on 20 November 2023).
- Algethami, M.; Kulkarni, S.; Sadiq, M.T.; Tang, H.K.C.; Brownlie, J.; Jeyapalan, J.N.; Mongan, N.P.; Rakha, E.A.; Madhusudan, S. Towards Personalized Management of Ovarian Cancer. Cancer Manag. Res. 2022, 14, 3469–3483. [Google Scholar] [CrossRef] [PubMed]
- Brownlie, J.; Kulkarni, S.; Algethami, M.; Jeyapalan, J.N.; Mongan, N.P.; Rakha, E.A.; Madhusudan, S. Targeting DNA damage repair precision medicine strategies in cancer. Curr. Opin. Pharmacol. 2023, 70, 102381. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.; Brownlie, J.; Jeyapalan, J.N.; Mongan, N.P.; Rakha, E.A.; Madhusudan, S. Evolving DNA repair synthetic lethality targets in cancer. Biosci. Rep. 2022, 42, 42. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Kulkarni, S.; Peters, C.; Eddison, J.; Al-Ani, M.; Madhusudan, S. The Current Status of DNA-Repair-Directed Precision Oncology Strategies in Epithelial Ovarian Cancers. Int. J. Mol. Sci. 2023, 24, 7293. [Google Scholar] [CrossRef] [PubMed]
- Hartwell, L.H.; Weinert, T.A. Checkpoints: Controls that ensure the order of cell cycle events. Science 1989, 246, 629–634. [Google Scholar] [CrossRef]
- Malumbres, M.; Barbacid, M. To cycle or not to cycle: A critical decision in cancer. Nat. Rev. Cancer 2001, 1, 222–231. [Google Scholar] [CrossRef]
- Norbury, C.; Nurse, P. Cyclins and cell cycle control. Curr. Biol. 1991, 1, 23–24. [Google Scholar] [CrossRef] [PubMed]
- Norbury, C.; Nurse, P. Animal cell cycles and their control. Annu. Rev. Biochem. 1992, 61, 441–470. [Google Scholar] [CrossRef]
- D’Andrilli, G.; Kumar, C.; Scambia, G.; Giordano, A. Cell cycle genes in ovarian cancer: Steps toward earlier diagnosis and novel therapies. Clin. Cancer Res. 2004, 10, 8132–8141. [Google Scholar] [CrossRef]
- Nam, E.J.; Kim, Y.T. Alteration of cell-cycle regulation in epithelial ovarian cancer. Int. J. Gynecol. Cancer 2008, 18, 1169–1182. [Google Scholar] [CrossRef]
- Dall’Acqua, A.; Sonego, M.; Pellizzari, I.; Pellarin, I.; Canzonieri, V.; D’Andrea, S.; Benevol, S.; Sorio, R.; Giorda, G.; Califano, D.; et al. CDK6 protects epithelial ovarian cancer from platinum-induced death via FOXO3 regulation. EMBO Mol. Med. 2017, 9, 1415–1433. [Google Scholar] [CrossRef]
- Hortobagyi, G.N. Ribociclib for HR-Positive, Advanced Breast Cancer. N. Engl. J. Med. 2017, 376, 289. [Google Scholar] [PubMed]
- Hortobagyi, G.N.; Stemmer, S.M.; Burris, H.A.; Yap, Y.S.; Sonke, G.S.; Hart, L.; Campone, M.; Petrakova, K.; Winer, E.P.; Janni, W.; et al. Overall Survival with Ribociclib plus Letrozole in Advanced Breast Cancer. N. Engl. J. Med. 2022, 386, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Hortobagyi, G.N.; Stemmer, S.M.; Burris, H.A.; Yap, Y.S.; Sonke, G.S.; Paluch-Shimon, S.; Campone, M.; Blackwell, K.L.; Andre, F.; Winer, E.P.; et al. Ribociclib as First-Line Therapy for HR-Positive, Advanced Breast Cancer. N. Engl. J. Med. 2016, 375, 1738–1748. [Google Scholar] [CrossRef] [PubMed]
- Hortobagyi, G.N.; Stemmer, S.M.; Burris, H.A.; Yap, Y.S.; Sonke, G.S.; Paluch-Shimon, S.; Campone, M.; Petrakova, K.; Blackwell, K.L.; Winer, E.P.; et al. Updated results from MONALEESA-2, a phase III trial of first-line ribociclib plus letrozole versus placebo plus letrozole in hormone receptor-positive, HER2-negative advanced breast cancer. Ann. Oncol. 2018, 29, 1541–1547. [Google Scholar] [CrossRef] [PubMed]
- Giordano, S.H.; Freedman, R.A.; Somerfield, M.R. Abemaciclib with Endocrine Therapy in the Treatment of High-Risk Early Breast Cancer: ASCO Optimal Adjuvant Chemotherapy and Targeted Therapy Guideline Rapid Recommendation Update. J. Clin. Oncol. 2022, 40, 307–309. [Google Scholar] [CrossRef] [PubMed]
- Slamon, D.J.; Stroyakovskiy, D.; Yardley, D.A.; Huang, C.-S.; Fasching, P.A.; Crown, J.; Bardia, A.; Chia, S.; Im, S.-A.; Martin, M.; et al. Ribociclib and endocrine therapy as adjuvant treatment in patients with HR+/HER2- early breast cancer: Primary results from the phase III NATALEE trial. J. Clin. Oncol. 2023, 41 (Suppl S17), LBA500. [Google Scholar] [CrossRef]
- Schmitz, N.M.; Hirt, A.; Aebi, M.; Leibundgut, K. Limited redundancy in phosphorylation of retinoblastoma tumor suppressor protein by cyclin-dependent kinases in acute lymphoblastic leukemia. Am. J. Pathol. 2006, 169, 1074–1079. [Google Scholar] [CrossRef] [PubMed]
- Mei, J.; Tian, H.; Huang, H.S.; Wu, N.; Liou, Y.L.; Chu, T.Y.; Wang, J.; Zhang, W. CCNE1 is a potential target of Metformin for tumor suppression of ovarian high-grade serous carcinoma. Cell Cycle 2023, 22, 85–89. [Google Scholar] [CrossRef]
- Wang, L.L.; Xiu, Y.L.; Chen, X.; Sun, K.X.; Chen, S.; Wu, D.D.; Liu, B.L.; Zhao, Y. The transcription factor FOXA1 induces epithelial ovarian cancer tumorigenesis and progression. Tumour Biol. 2017, 39, 1010428317706210. [Google Scholar] [CrossRef]
- Lou, T.; Liu, C.; Qu, H.; Zhang, Z.; Wang, S.; Zhuang, H. FOXA1 can be modulated by HDAC3 in the progression of epithelial ovarian carcinoma. J. Transl. Med. 2022, 20, 19. [Google Scholar] [CrossRef] [PubMed]
- Fassl, A.; Geng, Y.; Sicinski, P. CDK4 and CDK6 kinases: From basic science to cancer therapy. Science 2022, 375, eabc1495. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.J.; Poulikakos, P.I.; Irie, H.Y.; Parekh, S.; Reddy, E.P. CDK4: A master regulator of the cell cycle and its role in cancer. Genes Cancer 2022, 13, 21–45. [Google Scholar] [CrossRef] [PubMed]
- Fuste, N.P.; Fernandez-Hernandez, R.; Cemeli, T.; Mirantes, C.; Pedraza, N.; Rafel, M.; Torres-Rosell, J.; Colomina, N.; Ferrezuelo, F.; Dolcet, X.; et al. Cytoplasmic cyclin D1 regulates cell invasion and metastasis through the phosphorylation of paxillin. Nat. Commun. 2016, 7, 11581. [Google Scholar] [CrossRef] [PubMed]
- Goel, S.; Bergholz, J.S.; Zhao, J.J. Targeting CDK4 and CDK6 in cancer. Nat. Rev. Cancer 2022, 22, 356–372. [Google Scholar] [CrossRef] [PubMed]
- Nebenfuehr, S.; Kollmann, K.; Sexl, V. The role of CDK6 in cancer. Int. J. Cancer 2020, 147, 2988–2995. [Google Scholar] [CrossRef]
- Sherr, C.J.; Beach, D.; Shapiro, G.I. Targeting CDK4 and CDK6: From Discovery to Therapy. Cancer Discov. 2016, 6, 353–367. [Google Scholar] [CrossRef]
- Brown, K.A.; Roberts, R.L.; Arteaga, C.L.; Law, B.K. Transforming growth factor-beta induces Cdk2 relocalization to the cytoplasm coincident with dephosphorylation of retinoblastoma tumor suppressor protein. Breast Cancer Res. 2004, 6, R130–R139. [Google Scholar] [CrossRef]
- Tadesse, S.; Caldon, E.C.; Tilley, W.; Wang, S. Cyclin-Dependent Kinase 2 Inhibitors in Cancer Therapy: An Update. J. Med. Chem. 2019, 62, 4233–4251. [Google Scholar] [CrossRef]
- Mayr, D.; Kanitz, V.; Anderegg, B.; Luthardt, B.; Engel, J.; Löhrs, U.; Amann, G.; Diebold, J. Analysis of gene amplification and prognostic markers in ovarian cancer using comparative genomic hybridization for microarrays and immunohistochemical analysis for tissue microarrays. Am. J. Clin. Pathol. 2006, 126, 101–109. [Google Scholar] [CrossRef]
- Sui, L.; Dong, Y.; Ohno, M.; Goto, M.; Inohara, T.; Sugimoto, K.; Tai, Y.; Hando, T.; Tokuda, M. Inverse expression of Cdk4 and p16 in epithelial ovarian tumors. Gynecol. Oncol. 2000, 79, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Kommoss, S.; du Bois, A.; Ridder, R.; Trunk, M.J.; Schmidt, D.; Pfisterer, J.; Kommoss, F. Independent prognostic significance of cell cycle regulator proteins p16(INK4a) and pRb in advanced-stage ovarian carcinoma including optimally debulked patients: A translational research subprotocol of a randomised study of the Arbeitsgemeinschaft Gynaekologische Onkologie Ovarian Cancer Study Group. Br. J. Cancer 2007, 96, 306–313. [Google Scholar]
- Dhar, K.K.; Branigan, K.; Parkes, J.; Howells, R.E.; Hand, P.; Musgrove, C.; Strange, R.C.; Fryer, A.A.; Redman, C.W.; Hoban, P.R. Expression and subcellular localization of cyclin D1 protein in epithelial ovarian tumour cells. Br. J. Cancer 1999, 81, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Bali, A.; O’Brien, P.M.; Edwards, L.S.; Sutherland, R.L.; Hacker, N.F.; Henshall, S.M. Cyclin D1, p53, and p21Waf1/Cip1 expression is predictive of poor clinical outcome in serous epithelial ovarian cancer. Clin. Cancer Res. 2004, 10, 5168–5177. [Google Scholar] [CrossRef] [PubMed]
- Abdelrahman, A.E.; Fathy, A.; Elsebai, E.A.; Nawar, N.; Etman, W.M. Prognostic impact of Apaf-1, Cyclin D1, and AQP-5 in serous ovarian carcinoma treated with the first-line chemotherapy. Ann. Diagn. Pathol. 2018, 35, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Yanaihara, N.; Okamoto, A.; Nikaido, T.; Saito, M.; Takakura, S.; Yasuda, M.; Sasaki, H.; Ochiai, K.; Tanaka, T. Cyclin D1 predicts the prognosis of advanced serous ovarian cancer. Exp. Ther. Med. 2011, 2, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Quan, L.L.; Liu, J.Y.; Qu, L.X.; La, H.; Wang, H.L.; Chen, X.X.; Wang, N.; Wei, Z.Z. Expression of Cyclin D1 gene in ovarian cancer and effect of silencing its expression on ovarian cancer cells based on the Oncomine database. Bioengineered 2021, 12, 9290–9300. [Google Scholar] [CrossRef]
- Iyengar, M.; O’Hayer, P.; Cole, A.; Sebastian, T.; Yang, K.; Coffman, L.; Buckanovich, R.J. CDK4/6 inhibition as maintenance and combination therapy for high grade serous ovarian cancer. Oncotarget 2018, 9, 15658–15672. [Google Scholar] [CrossRef] [PubMed]
- Ling, C.; Liu, S.; Wang, Y.; Zhang, F.C.; Du, Y. Expression of CDK6 in early ovarian cancer and its clinical significance. Nan Fang Yi Ke Da Xue Xue Bao J. South. Med. Univ. 2016, 36, 1271–1275. [Google Scholar]
- Sui, L.; Dong, Y.; Ohno, M.; Sugimoto, K.; Tai, Y.; Hando, T.; Tokuda, M. Implication of malignancy and prognosis of p27(kip1), Cyclin E, and Cdk2 expression in epithelial ovarian tumors. Gynecol. Oncol. 2001, 83, 56–63. [Google Scholar] [CrossRef]
- Nakayama, N.; Nakayama, K.; Shamima, Y.; Ishikawa, M.; Katagiri, A.; Iida, K.; Miyazaki, K. Gene amplification CCNE1 is related to poor survival and potential therapeutic target in ovarian cancer. Cancer 2010, 116, 2621–2634. [Google Scholar] [CrossRef] [PubMed]
- Pils, D.; Bachmayr-Heyda, A.; Auer, K.; Svoboda, M.; Auner, V.; Hager, G.; Obermayr, E.; Reiner, A.; Reinthaller, A.; Speiser, P.; et al. Cyclin E1 (CCNE1) as independent positive prognostic factor in advanced stage serous ovarian cancer patients—A study of the OVCAD consortium. Eur. J. Cancer 2014, 50, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Ayhan, A.; Kuhn, E.; Wu, R.C.; Ogawa, H.; Bahadirli-Talbott, A.; Mao, T.L.; Sugimura, H.; Shih, I.M.; Wang, T.L. CCNE1 copy-number gain and overexpression identify ovarian clear cell carcinoma with a poor prognosis. Mod. Pathol. 2017, 30, 297–303. [Google Scholar] [CrossRef]
- Petersen, S.; Wilson, A.J.; Hirst, J.; Roby, K.F.; Fadare, O.; Crispens, M.A.; Beeghly-Fadiel, A.; Khabele, D. CCNE1 and BRD4 co-amplification in high-grade serous ovarian cancer is associated with poor clinical outcomes. Gynecol. Oncol. 2020, 157, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Fang, D.; Chen, H.; Lu, Y.; Dong, Z.; Ding, H.F.; Jing, Q.; Su, S.B.; Huang, S. Cyclin-dependent kinase 2 is an ideal target for ovary tumors with elevated cyclin E1 expression. Oncotarget 2015, 6, 20801–20812. [Google Scholar] [CrossRef] [PubMed]
- Talbot, T.; Lu, H.; Aboagye, E.O. Amplified therapeutic targets in high-grade serous ovarian carcinoma—A review of the literature with quantitative appraisal. Cancer Gene Ther. 2023, 30, 955–963. [Google Scholar] [CrossRef]
- McShane, L.M.; Altman, D.G.; Sauerbrei, W.; Taube, S.E.; Gion, M.; Clark, G.M. REporting recommendations for tumour MARKer prognostic studies (REMARK). Br. J. Cancer 2005, 93, 387–391. [Google Scholar] [CrossRef]
- Camp, R.L.; Dolled-Filhart, M.; Rimm, D.L. X-tile: A new bio-informatics tool for biomarker assessment and outcome-based cut-point optimization. Clin. Cancer Res. 2004, 10, 7252–7259. [Google Scholar] [CrossRef]
- Bartha, A.; Gyorffy, B. TNMplot.com: A Web Tool for the Comparison of Gene Expression in Normal, Tumor and Metastatic Tissues. Int. J. Mol. Sci. 2021, 22, 2622. [Google Scholar] [CrossRef]
- Győrffy, B. Discovery and ranking of the most robust prognostic biomarkers in serous ovarian cancer. Geroscience 2023, 45, 1889–1898. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. WebGestalt 2019: Gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Nuclear CDK2 Expression | |||
|---|---|---|---|---|
| Low (H Score ≤ 5) | High (H Score > 5) | p Value | Adjusted p Value | |
| N (%) | N (%) | |||
| Pathology Type | ||||
| Serous | 108 (51%) | 53 (65%) | 0.026 | 0.06 |
| Mucinous | 38 (18%) | 5 (6%) | ||
| Endometriod | 33 (15%) | 7 (9%) | ||
| Clear Cell | 14 (6%) | 10 (12%) | ||
| Other | 9 (4%) | 2 (3%) | ||
| Mixed | 12 (6%) | 4 (5%) | ||
| Pathology Grade | ||||
| Low | 31 (17%) | 7 (10%) | 0.001 | 0.005 |
| Med | 47 (26%) | 7 (10%) | ||
| High | 101 (57%) | 58 (80%) | ||
| Pathology Stage | ||||
| 1 | 89 (44%) | 23 (28%) | 0.031 | 0.09 |
| 2 | 32 (16%) | 11 (14%) | ||
| 3 | 74 (37%) | 45 (56%) | ||
| 4 | 7 (3%) | 2 (2%) | ||
| Residual TumourPost Surgery | ||||
| Non | 142 (73%) | 48 (65%) | 0.511 | 0.511 |
| <1 cm | 22 (11%) | 11 (15%) | ||
| 1–2 cm | 7 (4%) | 5 (7%) | ||
| >2 cm | 24 (12%) | 10 (13%) | ||
| Platinum Sensitivity | 0.312 | 0.31 | ||
| Sensitive | 168 (92%) | 60 (88%) | ||
| Resistant | 14 (8%) | 8 (12%) | ||
| Parameters | Progression Free Survival | Overall Survival | ||||
|---|---|---|---|---|---|---|
| Hazard Ratio | 95% (CI) | p-Value | Hazard Ratio | 95% (CI) | p-Value | |
| CDK4 | 0.63 | 0.29–1.3 | 0.23 | 0.78 | 0.41–1.5 | 0.46 |
| CDK6 | 0.81 | 0.32–2.1 | 0.65 | 1.4 | 0.68–2.8 | 0.38 |
| CDK2 | 0.82 | 0.35–1.9 | 0.64 | 0.99 | 0.45–2.1 | 0.98 |
| Cyclin D1 | 0.64 | 0.07–5.3 | 0.68 | 0.77 | 0.16–3.5 | 0.73 |
| Cyclin E1 | 2.11 | 1.1–4.2 | 0.03 | 1.4 | 0.73–2.8 | 0.32 |
| pRB1 | 1.69 | 0.68–4.2 | 0.26 | 1.7 | 0.73–4.5 | 0.19 |
| RB1 | 1.33 | 0.71–2.5 | 0.36 | 0.89 | 0.5–1.5 | 0.67 |
| Pathology stage | 1.9 | 1.3–2.8 | <0.0001 | 2.4 | 1.69–3.41 | <0.0001 |
| Variables | Cyclin E1 Expression | p-Value | |
|---|---|---|---|
| Low | High | ||
| CDK2 expression | |||
| Low | 159 (81%) | 38 (19%) | 34.7 |
| High | 20 (39%) | 31 (61%) | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lashen, A.; Algethami, M.; Alqahtani, S.; Shoqafi, A.; Sheha, A.; Jeyapalan, J.N.; Mongan, N.P.; Rakha, E.A.; Madhusudan, S. The Clinicopathological Significance of the Cyclin D1/E1–Cyclin-Dependent Kinase (CDK2/4/6)–Retinoblastoma (RB1/pRB1) Pathway in Epithelial Ovarian Cancers. Int. J. Mol. Sci. 2024, 25, 4060. https://doi.org/10.3390/ijms25074060
Lashen A, Algethami M, Alqahtani S, Shoqafi A, Sheha A, Jeyapalan JN, Mongan NP, Rakha EA, Madhusudan S. The Clinicopathological Significance of the Cyclin D1/E1–Cyclin-Dependent Kinase (CDK2/4/6)–Retinoblastoma (RB1/pRB1) Pathway in Epithelial Ovarian Cancers. International Journal of Molecular Sciences. 2024; 25(7):4060. https://doi.org/10.3390/ijms25074060
Chicago/Turabian StyleLashen, Ayat, Mashael Algethami, Shatha Alqahtani, Ahmed Shoqafi, Amera Sheha, Jennie N. Jeyapalan, Nigel P. Mongan, Emad A. Rakha, and Srinivasan Madhusudan. 2024. "The Clinicopathological Significance of the Cyclin D1/E1–Cyclin-Dependent Kinase (CDK2/4/6)–Retinoblastoma (RB1/pRB1) Pathway in Epithelial Ovarian Cancers" International Journal of Molecular Sciences 25, no. 7: 4060. https://doi.org/10.3390/ijms25074060
APA StyleLashen, A., Algethami, M., Alqahtani, S., Shoqafi, A., Sheha, A., Jeyapalan, J. N., Mongan, N. P., Rakha, E. A., & Madhusudan, S. (2024). The Clinicopathological Significance of the Cyclin D1/E1–Cyclin-Dependent Kinase (CDK2/4/6)–Retinoblastoma (RB1/pRB1) Pathway in Epithelial Ovarian Cancers. International Journal of Molecular Sciences, 25(7), 4060. https://doi.org/10.3390/ijms25074060