

Understanding the Impact of Polyunsaturated Fatty Acids on Age-Related Macular Degeneration: A Review

 , ,
, ,  and
and
Abstract
:1. Introduction
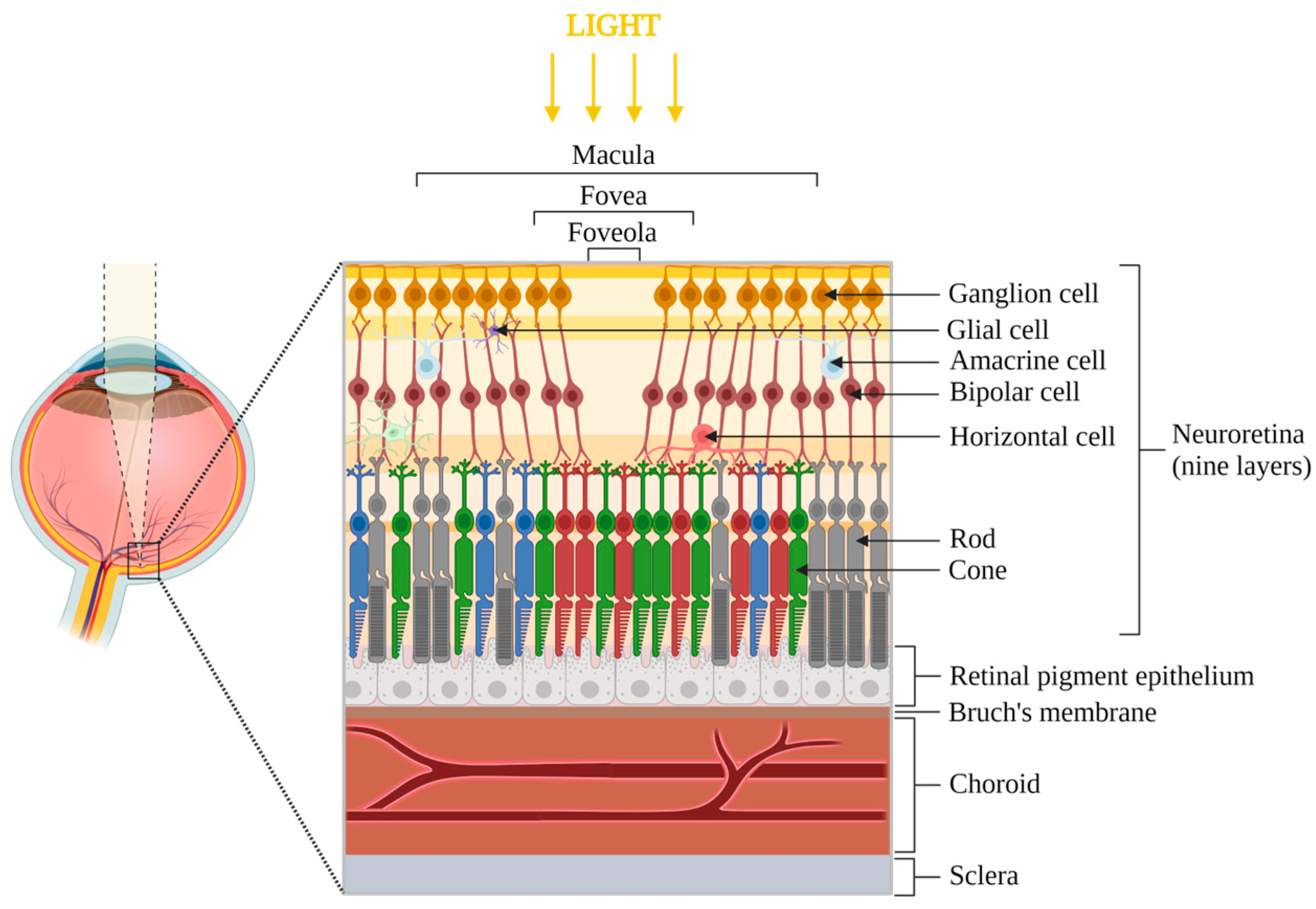
2. Retina and Vision
2.1. Eye Anatomy and Principal Functionalities
2.2. Physiological Aging
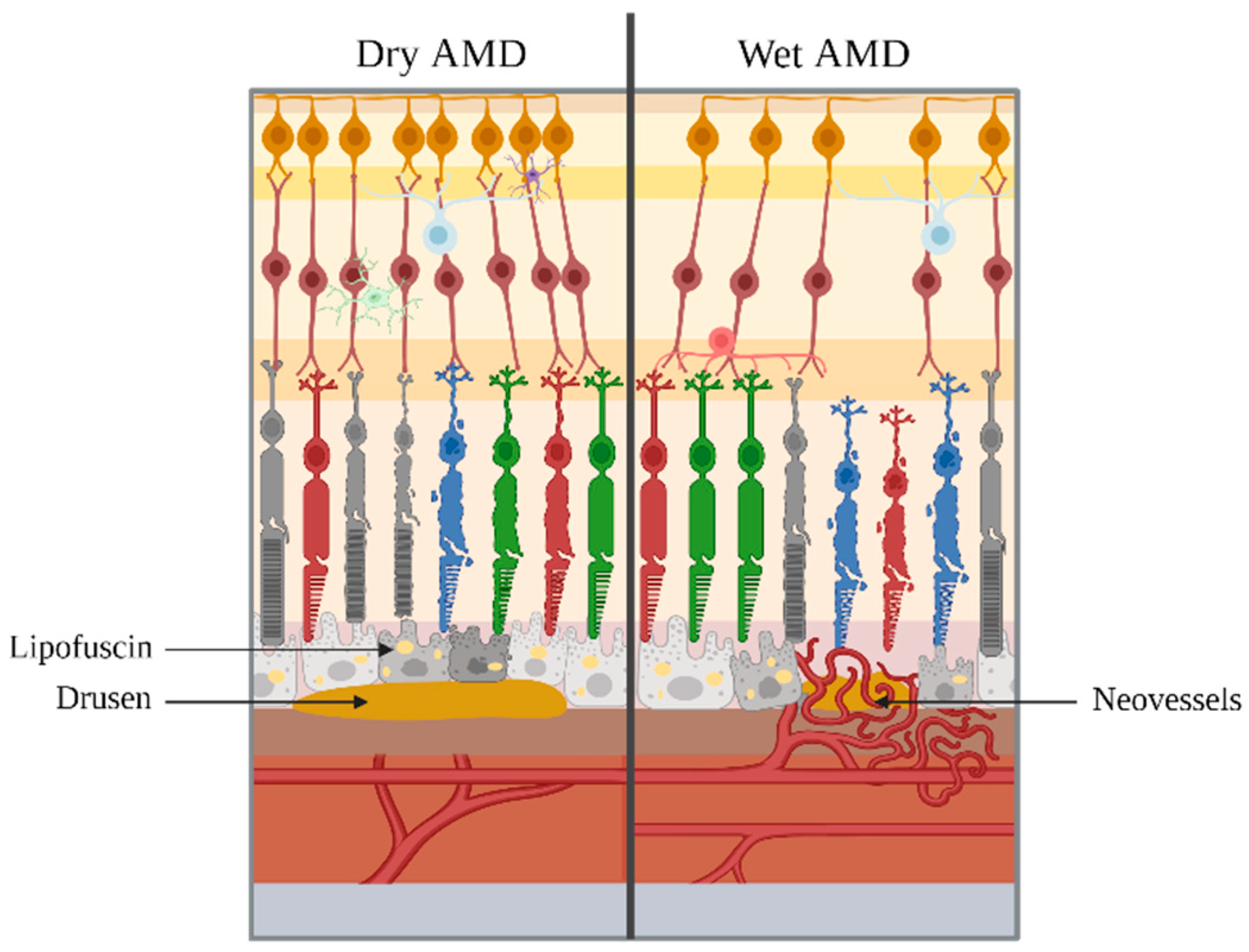
3. AMD: Clinical Aspects
3.1. Risk Factors
3.2. Evolution of AMD
4. AMD: Pathophysiology
4.1. Oxidative Stress
4.2. Inflammation
4.3. Choroidal Neovascularization
4.4. RPE and Bruch’s Membrane Degradation
5. Dry and Wet AMD Treatments
6. Representative Models of AMD
6.1. In Vitro and Ex Vivo Models
6.2. In Vivo Models
7. The Specific Role of Fatty Acids in AMD
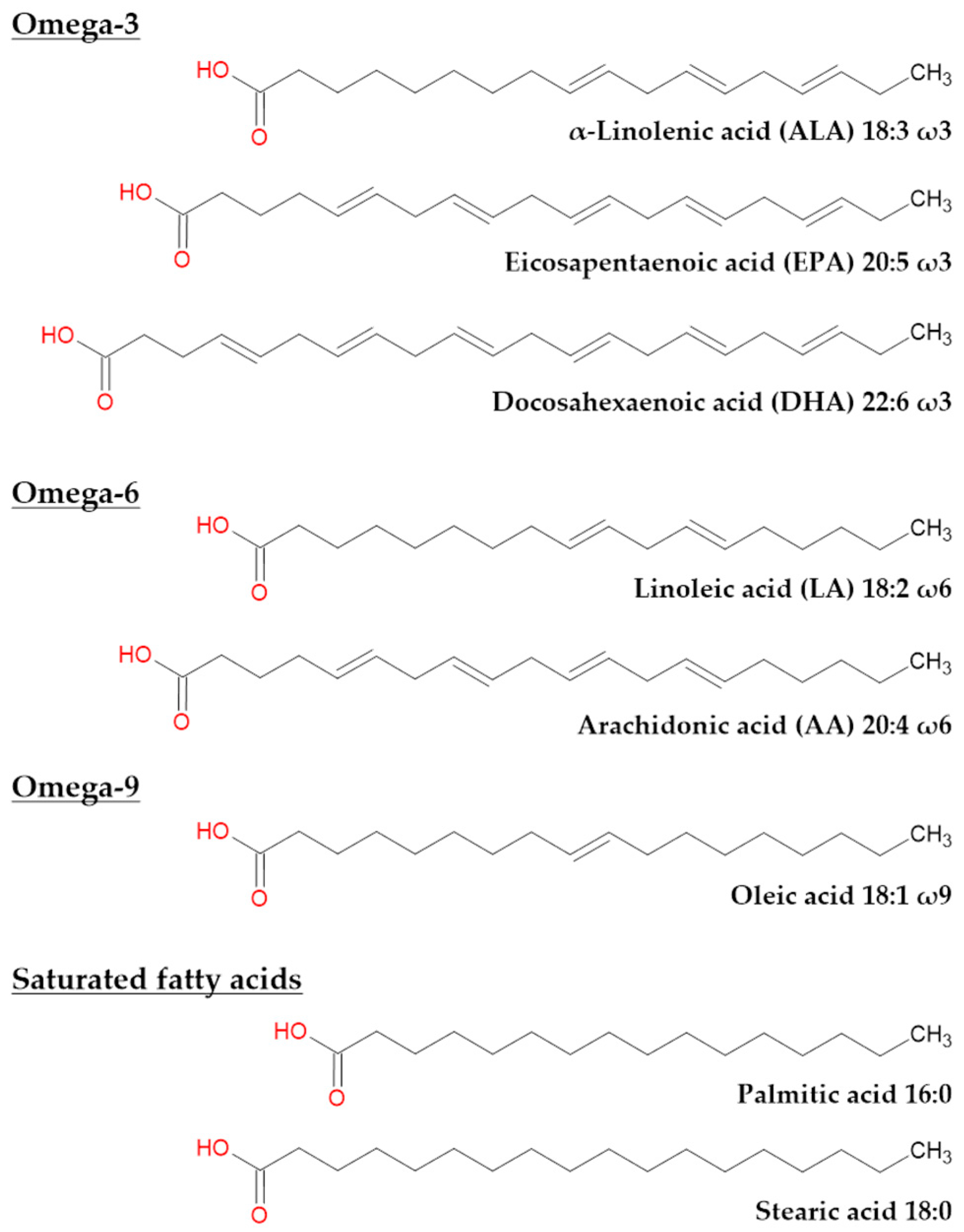
7.1. Fatty Acids in Humans
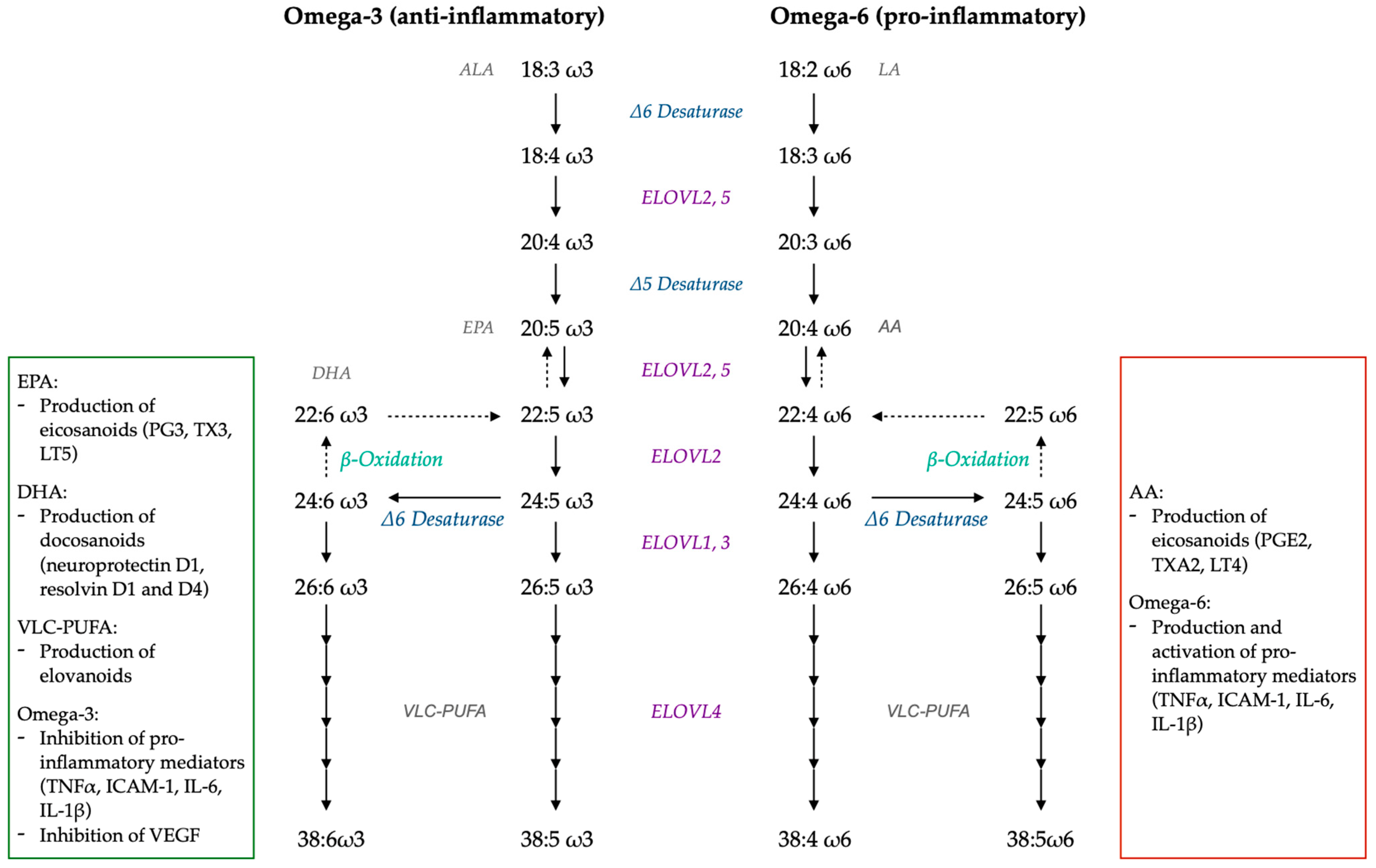
7.2. LC-PUFA and VLC-PUFA Metabolism

7.3. Retinal LC-PUFA and VLC-PUFA: Physiological Role
7.4. Retinal LC-PUFA and VLC-PUFA: Role in AMD
7.4.1. PUFA and Oxidative Stress
7.4.2. PUFA and Inflammation
7.5. Interest of Omega-3 LC-PUFA and VLC-PUFA in the Prevention and the Treatment of Dry and Wet AMD: Clinical Studies
8. Conclusions
9. Future Directions and Challenges
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Fernández-Robredo, P.; Sancho, A.; Johnen, S.; Recalde, S.; Gama, N.; Thumann, G.; Groll, J.; García-Layana, A. Current Treatment Limitations in Age-Related Macular Degeneration and Future Approaches Based on Cell Therapy and Tissue Engineering. J. Ophthalmol. 2014, 2014, 510285. [Google Scholar] [CrossRef]
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.G.; Klein, R.; Cheng, C.-Y.; Wong, T.Y. Global Prevalence of Age-Related Macular Degeneration and Disease Burden Projection for 2020 and 2040: A Systematic Review and Meta-Analysis. Lancet Glob. Health 2014, 2, e106–e116. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Chang, J.; Lin, Y.; Shen, Z.; Bernstein, P. Long-Chain and Very Long-Chain Polyunsaturated Fatty Acids in Ocular Aging and Age-Related Macular Degeneration. J. Lipid Res. 2010, 51, 3217–3229. [Google Scholar] [CrossRef] [PubMed]
- Watson, S.; Lowe, G. Ocular Anatomy and Physiology Relevant to Anaesthesia. Anaesth. Intensive Care Med. 2019, 20, 710–715. [Google Scholar] [CrossRef]
- Behar-Cohen, F.; Gelizé, E.; Jonet, L.; Lassiaz, P. Anatomy of the retina. Med. Sci. 2020, 36, 594–599. [Google Scholar]
- Lecerf, J.-M. Micronutriments: L’exemple de la dégénérescence maculaire liée à l’âge (DMLA): Micronutrients and Age-Related Macular Degeneration. Méd. Mal. Métab. 2009, 3, 496–501. [Google Scholar]
- Jacobs, G.H. Photopigments and the Dimensionality of Animal Color Vision. Neurosci. Biobehav. Rev. 2018, 86, 108–130. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, B.G.; Weisinger, H.S.; Neuringer, M.; Mitchell, D.C. The Role of Docosahexaenoic Acid in Retinal Function. Lipids 2001, 36, 859–871. [Google Scholar] [CrossRef] [PubMed]
- Gregg, R.; McCall, M.; Massey, S. Function and Anatomy of the Mammalian Retina. Retina 2012, 1, 360–400. [Google Scholar]
- Ratnayake, K.; Payton, J.L.; Lakmal, O.H.; Karunarathne, A. Blue Light Excited Retinal Intercepts Cellular Signaling. Sci. Rep. 2018, 8, 10207. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.-C.; Zhou, Y.; Tan, G.; Li, J. Research Progress about the Effect and Prevention of Blue Light on Eyes. Int. J. Ophthalmol. 2018, 11, 1999–2003. [Google Scholar] [PubMed]
- Bonnel, S.; Mohand-Said, S.; Sahel, J.-A. The Aging of the Retina. Exp. Gerontol. 2003, 38, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Ao, J.; Wood, J.P.; Chidlow, G.; Gillies, M.C.; Casson, R.J. Retinal Pigment Epithelium in the Pathogenesis of Age-Related Macular Degeneration and Photobiomodulation as a Potential Therapy? Clin. Experiment. Ophthalmol. 2018, 46, 670–686. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-D.; Presley, J.B.; Chimento, M.F.; Curcio, C.A.; Johnson, M. Age-Related Changes in Human Macular Bruch’s Membrane as Seen by Quick-Freeze/Deep-Etch. Exp. Eye Res. 2007, 85, 202–218. [Google Scholar] [CrossRef] [PubMed]
- Nickla, D.L.; Wallman, J. The Multifunctional Choroid. Prog. Retin. Eye Res. 2010, 29, 144–168. [Google Scholar] [CrossRef] [PubMed]
- Lejoyeux, R.; Benillouche, J.; Ong, J.; Errera, M.-H.; Rossi, E.A.; Singh, S.R.; Dansingani, K.K.; da Silva, S.; Sinha, D.; Sahel, J.-A.; et al. Choriocapillaris: Fundamentals and Advancements. Prog. Retin. Eye Res. 2022, 87, 100997. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Luo, C.; Zhao, J.; Devarajan, G.; Xu, H. Immune Regulation in the Aging Retina. Prog. Retin. Eye Res. 2019, 69, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Campello, L.; Singh, N.; Advani, J.; Mondal, A.K.; Corso-Díaz, X.; Swaroop, A. Aging of the Retina: Molecular and Metabolic Turbulences and Potential Interventions. Annu. Rev. Vis. Sci. 2021, 7, 633–664. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Ami, T.B.; Hong, S.; Heintzmann, R.; Gerig, G.; Ablonczy, Z.; Curcio, C.A.; Ach, T.; Smith, R.T. Hyperspectral Autofluorescence Imaging of Drusen and Retinal Pigment Epithelium in Donor Eyes with Age-Related Macular Degeneration. Retina 2016, 36, S127–S136. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.H.; Talaga, K.C.; Rivest, A.J.; Barron, E.; Hageman, G.S.; Johnson, L.V. Characterization of Beta Amyloid Assemblies in Drusen: The Deposits Associated with Aging and Age-Related Macular Degeneration. Exp. Eye Res. 2004, 78, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.L.; Quinn, J.; Xue, K. Interactions between Apolipoprotein E Metabolism and Retinal Inflammation in Age-Related Macular Degeneration. Life 2021, 11, 635. [Google Scholar] [CrossRef]
- Russell, S.R.; Mullins, R.F.; Schneider, B.L.; Hageman, G.S. Location, Substructure, and Composition of Basal Laminar Drusen Compared with Drusen Associated with Aging and Age-Related Macular Degeneration. Am. J. Ophthalmol. 2000, 129, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Ferris, F.L.; Wilkinson, C.P.; Bird, A.; Chakravarthy, U.; Chew, E.; Csaky, K.; Sadda, S.R. Clinical Classification of Age-Related Macular Degeneration. Ophthalmology 2013, 120, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Jabbehdari, S.; Handa, J.T. Oxidative Stress as a Therapeutic Target for the Prevention and Treatment of Early Age-Related Macular Degeneration. Surv. Ophthalmol. 2021, 66, 423–440. [Google Scholar] [CrossRef]
- Crouch, R.K.; Koutalos, Y.; Kono, M.; Schey, K.; Ablonczy, Z. A2E and Lipofuscin. Prog. Mol. Biol. Transl. Sci. 2015, 134, 449–463. [Google Scholar] [PubMed]
- Ardeljan, D.; Chan, C.-C. Aging Is Not a Disease: Distinguishing Age-Related Macular Degeneration from Aging. Prog. Retin. Eye Res. 2013, 37, 68–89. [Google Scholar] [CrossRef] [PubMed]
- Rozing, M.P.; Durhuus, J.A.; Krogh Nielsen, M.; Subhi, Y.; Kirkwood, T.B.; Westendorp, R.G.; Sørensen, T.L. Age-Related Macular Degeneration: A Two-Level Model Hypothesis. Prog. Retin. Eye Res. 2020, 76, 100825. [Google Scholar] [CrossRef] [PubMed]
- Chauvaud, D. La dégénérescence maculaire liée à l’âge en 2020. Bull. Acad. Natl. Méd. 2021, 205, 161–165. [Google Scholar] [CrossRef]
- Tzoumas, N.; Hallam, D.; Harris, C.L.; Lako, M.; Kavanagh, D.; Steel, D.H.W. Revisiting the Role of Factor H in Age-Related Macular Degeneration: Insights from Complement-Mediated Renal Disease and Rare Genetic Variants. Surv. Ophthalmol. 2021, 66, 378–401. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Liu, S.; Hao, Q.; Liu, L.; Zhang, J.; Chen, X.; Hu, W.; Huang, P. Association Between Complement Factor C2/C3/CFB/CFH Polymorphisms and Age-Related Macular Degeneration: A Meta-Analysis. Genet. Test. Mol. Biomark. 2018, 22, 526–540. [Google Scholar] [CrossRef] [PubMed]
- Armento, A.; Ueffing, M.; Clark, S.J. The Complement System in Age-Related Macular Degeneration. Cell. Mol. Life Sci. 2021, 78, 4487–4505. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.-G.; May, A.; Dinh, B.; Lin, V.; Su, F.; Tran, C.; Adivikolanu, H.; Ehlen, R.; Che, B.; Wang, Z.-H.; et al. The Interplay of Oxidative Stress and ARMS2-HTRA1 Genetic Risk in Neovascular AMD. Vessel. Plus 2021, 5, 4. [Google Scholar] [CrossRef]
- Marais, A.D. Apolipoprotein E in Lipoprotein Metabolism, Health and Cardiovascular Disease. Pathology 2019, 51, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Song, R.; Ai, Y.; Zhu, J.; He, J.; Dang, M.; Li, H. APOE2 Promotes the Development and Progression of Subretinal Neovascularization in Age-Related Macular Degeneration via MAPKs Signaling Pathway. Saudi J. Biol. Sci. 2020, 27, 2770–2777. [Google Scholar] [CrossRef] [PubMed]
- McKay, G.J.; Patterson, C.C.; Chakravarthy, U.; Dasari, S.; Klaver, C.C.; Vingerling, J.R.; Ho, L.; de Jong, P.T.; Fletcher, A.E.; Young, I.S.; et al. Evidence of Association of APOE with Age-Related Macular Degeneration—A Pooled Analysis of 15 Studies. Hum. Mutat. 2011, 32, 1407–1416. [Google Scholar] [CrossRef] [PubMed]
- Liutkeviciene, R.; Vilkeviciute, A.; Smalinskiene, A.; Tamosiunas, A.; Petkeviciene, J.; Zaliuniene, D.; Lesauskaite, V. The Role of Apolipoprotein E (Rs7412 and Rs429358) in Age-Related Macular Degeneration. Ophthalmic Genet. 2018, 39, 457–462. [Google Scholar] [CrossRef]
- Viturino, M.G.; Neto, J.M.; Bajano, F.F.; Costa, S.M.; Roque, A.B.; Borges, G.F.; Ananina, G.; Rim, P.H.; Medina, F.M.; Costa, F.F.; et al. Evaluation of APOE Polymorphisms and the Risk for Age-Related Macular Degeneration in a Southeastern Brazilian Population. Exp. Biol. Med. 2021, 246, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Vega, B.; García, M.; Olivares, L.; Álvarez, L.; González-Fernández, A.; Artime, E.; Fernández-Vega Cueto, A.; Cobo, T.; Coca-Prados, M.; Vega, J.A.; et al. The Association Study of Lipid Metabolism Gene Polymorphisms with AMD Identifies a Protective Role for APOE-E2 Allele in the Wet Form in a Northern Spanish Population. Acta Ophthalmol. 2020, 98, e282–e291. [Google Scholar] [CrossRef]
- Velilla, S.; García-Medina, J.J.; García-Layana, A.; Dolz-Marco, R.; Pons-Vázquez, S.; Pinazo-Durán, M.D.; Gómez-Ulla, F.; Arévalo, J.F.; Díaz-Llopis, M.; Gallego-Pinazo, R. Smoking and Age-Related Macular Degeneration: Review and Update. J. Ophthalmol. 2013, 2013, 895147. [Google Scholar] [CrossRef]
- Saunier, V.; Merle, B.M.J.; Delyfer, M.-N.; Cougnard-Grégoire, A.; Rougier, M.-B.; Amouyel, P.; Lambert, J.-C.; Dartigues, J.-F.; Korobelnik, J.-F.; Delcourt, C. Incidence of and Risk Factors Associated with Age-Related Macular Degeneration. JAMA Ophthalmol. 2018, 136, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Keenan, T.D.; Agrón, E.; Mares, J.; Clemons, T.E.; van Asten, F.; Swaroop, A.; Chew, E.Y. Adherence to the Mediterranean Diet and Progression to Late Age-Related Macular Degeneration in the Age-Related Eye Disease Studies 1 and 2. Ophthalmology 2020, 127, 1515–1528. [Google Scholar] [CrossRef] [PubMed]
- Merle, B.M.J.; Silver, R.E.; Rosner, B.; Seddon, J.M. Adherence to a Mediterranean Diet, Genetic Susceptibility, and Progression to Advanced Macular Degeneration: A Prospective Cohort Study. Am. J. Clin. Nutr. 2015, 102, 1196–1206. [Google Scholar] [CrossRef] [PubMed]
- Abdouh, M.; Lu, M.; Chen, Y.; Goyeneche, A.; Burnier, J.V.; Burnier, M.N. Filtering Blue Light Mitigates the Deleterious Effects Induced by the Oxidative Stress in Human Retinal Pigment Epithelial Cells. Exp. Eye Res. 2022, 217, 108978. [Google Scholar] [CrossRef] [PubMed]
- Delcourt, C.; Cougnard-Grégoire, A.; Boniol, M.; Carrière, I.; Doré, J.-F.; Delyfer, M.-N.; Rougier, M.-B.; Le Goff, M.; Dartigues, J.-F.; Barberger-Gateau, P.; et al. Lifetime Exposure to Ambient Ultraviolet Radiation and the Risk for Cataract Extraction and Age-Related Macular Degeneration: The Alienor Study. Investig. Ophthalmol. Vis. Sci. 2014, 55, 7619–7627. [Google Scholar] [CrossRef] [PubMed]
- Mainster, M.A.; Findl, O.; Dick, H.B.; Desmettre, T.; Ledesma-Gil, G.; Curcio, C.A.; Turner, P.L. The Blue-Light-Hazard vs. Blue-Light-Hype. Am. J. Ophthalmol. 2022, 240, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Margrain, T.H.; Boulton, M.; Marshall, J.; Sliney, D.H. Do Blue Light Filters Confer Protection against Age-Related Macular Degeneration? Prog. Retin. Eye Res. 2004, 23, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Desmettre, T. Épigénétique et dégénérescence maculaire liée à l’âge (DMLA). J. Fr. Ophtalmol. 2018, 41, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Van Lookeren Campagne, M.; LeCouter, J.; Yaspan, B.L.; Ye, W. Mechanisms of Age-Related Macular Degeneration and Therapeutic Opportunities. J. Pathol. 2014, 232, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Schultz, N.M.; Bhardwaj, S.; Barclay, C.; Gaspar, L.; Schwartz, J. Global Burden of Dry Age-Related Macular Degeneration: A Targeted Literature Review. Clin. Ther. 2021, 43, 1792–1818. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Zimbrón, L.F.; Zamora-Alvarado, R.; Ochoa-De la Paz, L.; Velez-Montoya, R.; Zenteno, E.; Gulias-Cañizo, R.; Quiroz-Mercado, H.; Gonzalez-Salinas, R. Age-Related Macular Degeneration: New Paradigms for Treatment and Management of AMD. Oxid. Med. Cell. Longev. 2018, 2018, 8374647. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.; Sennlaub, F.; Souied, E.; Cohen, S.-Y.; Béhar-Cohen, F.; Milano, G.; Tadayoni, R. Expertises croisées dans la dégénérescence maculaire liée à l’âge. Focus sur la physiopathologie, l’angiogenèse, les données pharmacologiques et cliniques. J. Fr. Ophtalmol. 2014, 37, 566–579. [Google Scholar] [CrossRef] [PubMed]
- Ajana, S. Prédiction Du Risque de DMLA: Identification de Nouveaux Biomarqueurs et Modélisation Du Risque. Ph.D. Thesis, Université de Bordeaux, Bordeaux, France, 2019. [Google Scholar]
- Khanani, A.M.; Skelly, A.; Bezlyak, V.; Griner, R.; Torres, L.R.; Sagkriotis, A. SIERRA-AMD: A Retrospective, Real-World Evidence Study of Patients with Neovascular Age-Related Macular Degeneration in the United States. Ophthalmol. Retina 2020, 4, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, K.; Kannan, R.; Hinton, D.R. Molecular Mechanisms of Subretinal Fibrosis in Age-Related Macular Degeneration. Exp. Eye Res. 2016, 142, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Xu, H. Parainflammation, Chronic Inflammation, and Age-Related Macular Degeneration. J. Leukoc. Biol. 2015, 98, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The Impact of Oxidative Stress and Inflammation on RPE Degeneration in Non-Neovascular AMD. Prog. Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Fujihara, M.; Nagai, N.; Sussan, T.E.; Biswal, S.; Handa, J.T. Chronic Cigarette Smoke Causes Oxidative Damage and Apoptosis to Retinal Pigmented Epithelial Cells in Mice. PLoS ONE 2008, 3, e3119. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, M.; Milliner, C.; Bell, B.A.; Bonilha, V.L. Oxidative Stress in the Retina and Retinal Pigment Epithelium (RPE): Role of Aging, and DJ-1. Redox Biol. 2020, 37, 101623. [Google Scholar] [CrossRef] [PubMed]
- Nowak, J.Z. Age-Related Macular Degeneration (AMD): Pathogenesis and Therapy. Pharmacol. Rep. 2006, 58, 353–363. [Google Scholar] [PubMed]
- Barker, F.M., II; Snodderly, D.M.; Johnson, E.J.; Schalch, W.; Koepcke, W.; Gerss, J.; Neuringer, M. Nutritional Manipulation of Primate Retinas, V: Effects of Lutein, Zeaxanthin, and n–3 Fatty Acids on Retinal Sensitivity to Blue-Light–Induced Damage. Investig. Ophthalmol. Vis. Sci. 2011, 52, 3934–3942. [Google Scholar] [CrossRef] [PubMed]
- Sparrow, J.R.; Nakanishi, K.; Parish, C.A. The Lipofuscin Fluorophore A2E Mediates Blue Light-Induced Damage to Retinal Pigmented Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1981–1989. [Google Scholar]
- Wakx, A.; Dutot, M.; Massicot, F.; Mascarelli, F.; Limb, G.; Rat, P. Amyloid β Peptide Induces Apoptosis Through P2X7 Cell Death Receptor in Retinal Cells: Modulation by Marine Omega-3 Fatty Acid DHA and EPA. Appl. Biochem. Biotechnol. 2016, 178, 368–381. [Google Scholar] [CrossRef] [PubMed]
- Mathis, T.; Housset, M.; Eandi, C.; Beguier, F.; Touhami, S.; Reichman, S.; Augustin, S.; Gondouin, P.; Sahel, J.-A.; Kodjikian, L.; et al. Activated Monocytes Resist Elimination by Retinal Pigment Epithelium and Downregulate Their OTX2 Expression via TNF-α. Aging Cell 2017, 16, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Kauppinen, A.; Paterno, J.J.; Blasiak, J.; Salminen, A.; Kaarniranta, K. Inflammation and Its Role in Age-Related Macular Degeneration. Cell. Mol. Life Sci. 2016, 73, 1765–1786. [Google Scholar] [CrossRef] [PubMed]
- Mankan, A.K.; Dau, T.; Jenne, D.; Hornung, V. The NLRP3/ASC/Caspase-1 Axis Regulates IL-1β Processing in Neutrophils. Eur. J. Immunol. 2012, 42, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Hsu, L.-C.; Ali, S.R.; McGillivray, S.; Tseng, P.-H.; Mariathasan, S.; Humke, E.W.; Eckmann, L.; Powell, J.J.; Nizet, V.; Dixit, V.M.; et al. A NOD2-NALP1 Complex Mediates Caspase-1-Dependent IL-1beta Secretion in Response to Bacillus Anthracis Infection and Muramyl Dipeptide. Proc. Natl. Acad. Sci. USA 2008, 105, 7803–7808. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Cheng, R.; Benyajati, S.; Ma, J. PEDF and Its Roles in Physiological and Pathological Conditions: Implication in Diabetic and Hypoxia-Induced Angiogenic Diseases. Clin. Sci. 2015, 128, 805–823. [Google Scholar] [CrossRef]
- Witmer, A.N.; Vrensen, G.F.J.M.; Van Noorden, C.J.F.; Schlingemann, R.O. Vascular Endothelial Growth Factors and Angiogenesis in Eye Disease. Prog. Retin. Eye Res. 2003, 22, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; McGuire, P.G. Retinal and Choroidal Angiogenesis: Pathophysiology and Strategies for Inhibition. Prog. Retin. Eye Res. 2003, 22, 721–748. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Zou, J.; Yoshida, S.; Jiang, B.; Zhou, Y. The Role of Inflammation in Age-Related Macular Degeneration. Int. J. Biol. Sci. 2020, 16, 2989–3001. [Google Scholar] [CrossRef]
- Murali, A.; Krishnakumar, S.; Subramanian, A.; Parameswaran, S. Bruch’s Membrane Pathology: A Mechanistic Perspective. Eur. J. Ophthalmol. 2020, 30, 1195–1206. [Google Scholar] [CrossRef]
- Murphy, R.P. Age-Related Macular Degeneration. Ophthalmology 1986, 93, 969–971. [Google Scholar] [CrossRef] [PubMed]
- Avery, R.L.; Castellarin, A.A.; Steinle, N.C.; Dhoot, D.S.; Pieramici, D.J.; See, R.; Couvillion, S.; Nasir, M.A.; Rabena, M.D.; Maia, M.; et al. Systemic Pharmacokinetics and Pharmacodynamics of Intravitreal Aflibercept, Bevacizumab, and Ranibizumab. Retina 2017, 37, 1847–1858. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.D.; Das, A.; Do, D.V.; Dugel, P.U.; Gomes, A.; Holz, F.G.; Koh, A.; Pan, C.K.; Sepah, Y.J.; Patel, N.; et al. Brolucizumab: Evolution through Preclinical and Clinical Studies and the Implications for the Management of Neovascular Age-Related Macular Degeneration. Ophthalmology 2020, 127, 963–976. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Ramón, P.V.; Hernández Martínez, P.; Muñoz-Negrete, F.J. New Therapeutic Targets in the Treatment of Age-Related Macular Degeneration. Arch. Soc. Esp. Oftalmol. 2020, 95, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Nicolò, M.; Ferro Desideri, L.; Vagge, A.; Traverso, C.E. Faricimab: An Investigational Agent Targeting the Tie-2/Angiopoietin Pathway and VEGF-A for the Treatment of Retinal Diseases. Expert Opin. Investig. Drugs 2021, 30, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, P.R.; Madanagopalan, V.G. KSI-301: Antibody Biopolymer Conjugate in Retinal Disorders. Ther. Adv. Ophthalmol. 2021, 13, 25158414211027708. [Google Scholar] [CrossRef] [PubMed]
- Kodiak Sciences Inc. A Prospective, Randomized, Double-Masked, Active Comparator-Controlled, Multi-Center, Two-Arm, Phase 3 Study to Evaluate the Efficacy and Safety of Intravitreal KSI-301 Compared with Intravitreal Aflibercept in Participants with Neovascular (Wet) Age-Related Macular Degeneration (wAMD). Available online: https://clinicaltrials.gov/study/NCT04964089 (accessed on 19 February 2024).
- EyePoint Pharmaceuticals, Inc. A Phase 2, Multicenter, Prospective, Randomized, Double-Masked, Parallel Study of EYP-1901, a Tyrosine Kinase Inhibitor (TKI), Compared to Aflibercept in Subjects with Wet AMD. Available online: https://clinicaltrials.gov/study/NCT05381948 (accessed on 19 February 2024).
- Clearside Biomedical, Inc. ODYSSEY: A Phase 2b Study of Suprachoroidally Administered CLS-AX in Participants with Neovascular Age-Related Macular Degeneration. Available online: https://clinicaltrials.gov/study/NCT05891548 (accessed on 19 February 2024).
- Ocular Therapeutix, Inc. A Phase 3, Multicenter, Double-Masked, Randomized, Parallel-Group Study to Evaluate the Efficacy and Safety of Intravitreal OTX-TKI (Axitinib Implant) in Subjects with Neovascular Age-Related Macular Degeneration (nAMD). Available online: https://clinicaltrials.gov/study/NCT06223958 (accessed on 19 February 2024).
- Novartis Pharmaceuticals. A Randomized, Participant and Investigator Masked, Placebo-Controlled, Multicenter, Proof-of-Concept Study to Assess the Safety and Efficacy of LNP023 (Iptacopan) in Patients With Early and Intermediate Age-Related Macular Degeneration. Available online: https://clinicaltrials.gov/study/NCT05230537 (accessed on 20 February 2024).
- Innovent Biologics (Suzhou) Co., Ltd. A Multi-Center, Randomized, Double-Blind, Active-Controlled Phase II Study to Evaluate the Efficacy and Safety of IBI302 in Subjects with Neovascular Age-Related Macular Degeneration. Available online: https://clinicaltrials.gov/study/NCT04820452 (accessed on 21 February 2024).
- AbbVie. A Phase 2, Open-Label Study to Explore the Pharmacodynamics of Two Doses in Two Formulations of RGX-314 Gene Therapy Administered Via Subretinal Delivery in Participants with Neovascular Age-Related Macular Degeneration. Available online: https://clinicaltrials.gov/study/NCT04832724 (accessed on 21 February 2024).
- Adverum Biotechnologies, Inc. A Multi-Center, Randomized, Double-Masked Phase 2 Study to Assess Safety and Efficacy of ADVM-022 (AAV.7m8-Aflibercept) in Anti-VEGF Treatment-Experienced Patients with Neovascular (Wet) Age-Related Macular Degeneration (nAMD) [LUNA]. Available online: https://clinicaltrials.gov/study/NCT05536973 (accessed on 20 February 2024).
- Oxford BioMedica. A Long Term Follow-up Study to Evaluate the Safety of RetinoStat® in Patients With Age-Related Macular Degeneration. Available online: https://clinicaltrials.gov/study/NCT01678872 (accessed on 20 February 2024).
- Age-Related Eye Disease Study Research Group. A Randomized, Placebo-Controlled, Clinical Trial of High-Dose Supplementation with Vitamins C and E, Beta Carotene, and Zinc for Age-Related Macular Degeneration and Vision Loss: AREDS Report No. 8. Arch Ophthalmol. 2001, 119, 1417–1436. [Google Scholar] [CrossRef] [PubMed]
- Sun, X. Effect of Omega-3 Fatty Acid (Krill Oil, 500 Mg, Including EPA 60 Mg and DHA 27.5 Mg, Total Phospholipid 200 Mg, Astaxanthin 50 Μg) Supplementation on Dry-AMD Progression. Available online: https://clinicaltrials.gov/ct2/show/NCT05465252 (accessed on 20 February 2024).
- Syfovre. European Medicines Agency. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/syfovre (accessed on 19 February 2024).
- Kang, C. Avacincaptad Pegol: First Approval. Drugs 2023, 83, 1447–1453. [Google Scholar] [CrossRef] [PubMed]
- Alkeus Pharmaceuticals, Inc. A Phase 2/3 Multicenter, Randomized, Double-Masked, Parallel-Group, Placebo-Controlled Study to Investigate the Safety, Pharmacokinetics, Tolerability, and Efficacy of ALK-001 in Geographic Atrophy Secondary to Age-Related Macular Degeneration. Available online: https://clinicaltrials.gov/study/NCT03845582 (accessed on 19 February 2024).
- ONL Therapeutics. A Phase 1b Multicenter, Randomized, Controlled, Single-Dose Study of the Safety and Tolerability of ONL1204 Ophthalmic Solution in Patients with Geographic Atrophy (GA) Associated with Age-Related Macular Degeneration (AMD). Available online: https://clinicaltrials.gov/study/NCT04744662 (accessed on 19 February 2024).
- Ionis Pharmaceuticals, Inc. A Phase 2, Randomized Placebo-Controlled, Double-Masked Study to Assess Safety and Efficacy of Multiple Doses of IONIS-FB-LRx, an Antisense Inhibitor of Complement Factor B, in Patients with Geographic Atrophy Secondary to Age-Related Macular Degeneration (AMD). Available online: https://clinicaltrials.gov/study/NCT03815825 (accessed on 20 February 2024).
- Alexion Pharmaceuticals, Inc. A Phase 2, Double-Masked, Placebo-Controlled, Dose Range Finding Study of Danicopan (ALXN2040) in Patients with Geographic Atrophy (GA) Secondary to Age-Related Macular Degeneration (AMD). Available online: https://clinicaltrials.gov/study/NCT05019521 (accessed on 19 February 2024).
- Gyroscope Therapeutics Limited. HORIZON: A Phase II, Open-Label, Outcomes-Assessor Masked, Multicentre, Randomised, Controlled Study to Evaluate the Safety and Efficacy of Two Doses of GT005 Administered as a Single Subretinal Injection in Subjects with Geographic Atrophy Secondary to Dry Age-Related Macular Degeneration. Available online: https://clinicaltrials.gov/study/NCT04566445 (accessed on 21 February 2024).
- Hoffmann-La Roche. Phase I/IIa Dose Escalation Safety and Efficacy Study of Human Embryonic Stem Cell-Derived Retinal Pigment Epithelium Cells Transplanted Subretinally in Patients with Advanced Dry-Form Age-Related Macular Degeneration (Geographic Atrophy). Available online: https://clinicaltrials.gov/study/NCT02286089 (accessed on 21 February 2024).
- Pixium Vision SA. Feasibility Study of Compensation for Blindness with the PRIMA System in Patients with Dry Age Related Macular Degeneration. Available online: https://clinicaltrials.gov/study/NCT03333954 (accessed on 19 February 2024).
- Pixium Vision SA. Feasibility Study of Compensation for Blindness with the PRIMA System in Patients with Atrophic Dry Age Related Macular Degeneration. Available online: https://clinicaltrials.gov/study/NCT03392324 (accessed on 21 February 2024).
- Pixium Vision SA. Restoration of Central Vision with the PRIMA System in Patients with Atrophic Age-Related Macular Degeneration. Available online: https://clinicaltrials.gov/ct2/show/NCT04676854 (accessed on 21 February 2024).
- Chen, D.; Chao, D.L.; Rocha, L.; Kolar, M.; Nguyen Huu, V.A.; Krawczyk, M.; Dasyani, M.; Wang, T.; Jafari, M.; Jabari, M.; et al. The Lipid Elongation Enzyme ELOVL2 Is a Molecular Regulator of Aging in the Retina. Aging Cell 2020, 19, e13100. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhang, T.; Yao, Y.; Zeng, S.; Li, M.; Xiang, H.; Zhao, C.; Cao, G.; Li, M.; Wan, R.; et al. Efficacy of Lenvatinib, a Multitargeted Tyrosine Kinase Inhibitor, on Laser-Induced CNV Mouse Model of Neovascular AMD. Exp. Eye Res. 2018, 168, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Voisin, A.; Plancheron, A.; Monville, C.; Balbous, A.; Gaillard, A.; Leveziel, N. iPSC-RPE Cells Derived from Atrophic Age-Related Macular Degeneration Patients Have a Typical Disease Phenotype. Investig. Ophthalmol. Vis. Sci. 2019, 60, 1224. [Google Scholar]
- Johnson, L.V.; Forest, D.L.; Banna, C.D.; Radeke, C.M.; Maloney, M.A.; Hu, J.; Spencer, C.N.; Walker, A.M.; Tsie, M.S.; Bok, D.; et al. Cell Culture Model That Mimics Drusen Formation and Triggers Complement Activation Associated with Age-Related Macular Degeneration. Proc. Natl. Acad. Sci. USA 2011, 108, 18277–18282. [Google Scholar] [CrossRef] [PubMed]
- Gorham, R.D.; Forest, D.L.; Tamamis, P.; López de Victoria, A.; Kraszni, M.; Kieslich, C.A.; Banna, C.D.; Bellows-Peterson, M.L.; Larive, C.K.; Floudas, C.A.; et al. Novel Compstatin Family Peptides Inhibit Complement Activation by Drusen-like Deposits in Human Retinal Pigmented Epithelial Cell Cultures. Exp. Eye Res. 2013, 116, 96–108. [Google Scholar] [CrossRef] [PubMed]
- De Cillà, S.; Farruggio, S.; Cocomazzi, G.; Mary, D.; Alkabes, M.; Rossetti, L.; Vujosevic, S.; Grossini, E. Aflibercept and Ranibizumab Modulate Retinal Pigment Epithelial Cells Function by Acting on Their Cross Talk with Vascular Endothelial Cells. Cell. Physiol. Biochem. 2020, 54, 161–179. [Google Scholar] [PubMed]
- Palanisamy, K.; Karunakaran, C.; Raman, R.; Chidambaram, S. Optimization of an in Vitro Bilayer Model for Studying the Functional Interplay between Human Primary Retinal Pigment Epithelial and Choroidal Endothelial Cells Isolated from Donor Eyes. BMC Res. Notes 2019, 12, 307. [Google Scholar] [CrossRef] [PubMed]
- Arık, Y.B.; Buijsman, W.; Loessberg-Zahl, J.; Cuartas-Vélez, C.; Veenstra, C.; Logtenberg, S.; Grobbink, A.M.; Bergveld, P.; Gagliardi, G.; den Hollander, A.I.; et al. Microfluidic Organ-on-a-Chip Model of the Outer Blood-Retinal Barrier with Clinically Relevant Read-Outs for Tissue Permeability and Vascular Structure. Lab A Chip 2021, 21, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.; Lee, S.; Lee, B.J.; Son, K.; Jeon, N.L.; Kim, J.H. Wet-AMD on a Chip: Modeling Outer Blood-Retinal Barrier In Vitro. Adv. Healthc. Mater. 2018, 7, 1700028. [Google Scholar] [CrossRef] [PubMed]
- Yeste Lozano, J.; Garcia-Ramirez, M.; Brunet, A.; Hernández, C.; Illa, X.; Canonge, R.; Villa, R. A Compartmentalized Microfluidic Chip with Crisscross Microgrooves and Electrophysiological Electrodes for Modeling the Blood-Retinal Barrier. Lab A Chip 2017, 18, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-J.; Ito, S.; Kai, H.; Nagamine, K.; Nagai, N.; Nishizawa, M.; Abe, T.; Kaji, H. Microfluidic Co-Cultures of Retinal Pigment Epithelial Cells and Vascular Endothelial Cells to Investigate Choroidal Angiogenesis. Sci. Rep. 2017, 7, 3538. [Google Scholar] [CrossRef] [PubMed]
- Buskin, A.; Zhu, L.; Chichagova, V.; Basu, B.; Mozaffari-Jovin, S.; Dolan, D.; Droop, A.; Collin, J.; Bronstein, R.; Mehrotra, S.; et al. Disrupted Alternative Splicing for Genes Implicated in Splicing and Ciliogenesis Causes PRPF31 Retinitis Pigmentosa. Nat. Commun. 2018, 9, 4234. [Google Scholar] [CrossRef]
- Chichagova, V.; Hilgen, G.; Ghareeb, A.; Georgiou, M.; Carter, M.; Sernagor, E.; Lako, M.; Armstrong, L. Human iPSC Differentiation to Retinal Organoids in Response to IGF1 and BMP4 Activation Is Line- and Method-dependent. Stem Cells 2020, 38, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Achberger, K.; Probst, C.; Haderspeck, J.; Bolz, S.; Rogal, J.; Chuchuy, J.; Nikolova, M.; Cora, V.; Antkowiak, L.; Haq, W.; et al. Merging Organoid and Organ-on-a-Chip Technology to Generate Complex Multi-Layer Tissue Models in a Human Retina-on-a-Chip Platform. eLife 2019, 8, e46188. [Google Scholar] [CrossRef] [PubMed]
- Fietz, A.; Schnichels, S.; Hurst, J. Co-Cultivation of Primary Porcine RPE Cells and Neuroretina Induces Inflammation: A Potential Inflammatory AMD-Model. Sci. Rep. 2023, 13, 19345. [Google Scholar] [CrossRef] [PubMed]
- Labrador-Velandia, S.; Alonso-Alonso, M.L.; Di Lauro, S.; García-Gutierrez, M.T.; Srivastava, G.K.; Pastor, J.C.; Fernandez-Bueno, I. Mesenchymal Stem Cells Provide Paracrine Neuroprotective Resources That Delay Degeneration of Co-Cultured Organotypic Neuroretinal Cultures. Exp. Eye Res. 2019, 185, 107671. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, G.A.; Lutz, D.; Shen, J.; Yuan, X.; Shen, H.; Cunningham, J.; Rivers, H.M. Topical Drug Delivery to the Posterior Segment of the Eye: Addressing the Challenge of Preclinical to Clinical Translation. Pharm. Res. 2018, 35, 245. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wu, Q. Normal Corneal Thickness Measurements in Pigmented Rabbits Using Spectral-Domain Anterior Segment Optical Coherence Tomography. Vet. Ophthalmol. 2013, 16, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Doughty, M.J.; Zaman, M.L. Human Corneal Thickness and Its Impact on Intraocular Pressure Measures: A Review and Meta-Analysis Approach. Surv. Ophthalmol. 2000, 44, 367–408. [Google Scholar] [CrossRef] [PubMed]
- Thomasy, S.M.; Eaton, J.S.; Timberlake, M.J.; Miller, P.E.; Matsumoto, S.; Murphy, C.J. Species Differences in the Geometry of the Anterior Segment Differentially Affect Anterior Chamber Cell Scoring Systems in Laboratory Animals. J. Ocul. Pharmacol. Ther. 2016, 32, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Qi, M.; He, M.; Wu, L.; Lin, S. Ethnic Difference of the Anterior Chamber Area and Volume and Its Association with Angle Width. Investig. Ophthalmol. Vis. Sci. 2012, 53, 3139–3144. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.W.; Sun, Y.; Warther, D.; Huffman, K.; Freeman, W.R.; Sailor, M.J.; Cheng, L. New Model of Proliferative Vitreoretinopathy in Rabbit for Drug Delivery and Pharmacodynamic Studies. Drug Deliv. 2018, 25, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Azhdam, A.M.; Goldberg, R.A.; Ugradar, S. In Vivo Measurement of the Human Vitreous Chamber Volume Using Computed Tomography Imaging of 100 Eyes. Transl. Vis. Sci. Technol. 2020, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Penn, J.; Mihai, D.M.; Washington, I. Morphological and Physiological Retinal Degeneration Induced by Intravenous Delivery of Vitamin A Dimers in Rabbits. Dis. Model. Mech. 2015, 8, 131–138. [Google Scholar] [PubMed]
- Wang, Q.; Wei, W.B.; Wang, Y.X.; Yan, Y.N.; Yang, J.Y.; Zhou, W.J.; Chan, S.Y.; Xu, L.; Jonas, J.B. Thickness of Individual Layers at the Macula and Associated Factors: The Beijing Eye Study 2011. BMC Ophthalmol. 2020, 20, 49. [Google Scholar] [CrossRef] [PubMed]
- Zernii, E.Y.; Baksheeva, V.E.; Iomdina, E.N.; Averina, O.A.; Permyakov, S.E.; Philippov, P.P.; Zamyatnin, A.A.; Senin, I.I. Rabbit Models of Ocular Diseases: New Relevance for Classical Approaches. CNS Neurol. Disord. Drug Targets 2016, 15, 267–291. [Google Scholar] [CrossRef] [PubMed]
- Coffey, P.J.; Gias, C.; McDermott, C.J.; Lundh, P.; Pickering, M.C.; Sethi, C.; Bird, A.; Fitzke, F.W.; Maass, A.; Chen, L.L.; et al. Complement Factor H Deficiency in Aged Mice Causes Retinal Abnormalities and Visual Dysfunction. Proc. Natl. Acad. Sci. USA 2007, 104, 16651–16656. [Google Scholar] [CrossRef] [PubMed]
- Cashman, S.M.; Desai, A.; Ramo, K.; Kumar-Singh, R. Expression of Complement Component 3 (C3) from an Adenovirus Leads to Pathology in the Murine Retina. Investig. Ophthalmol. Vis. Sci. 2011, 52, 3436–3445. [Google Scholar] [CrossRef] [PubMed]
- Ufret-Vincenty, R.L.; Aredo, B.; Liu, X.; McMahon, A.; Chen, P.W.; Sun, H.; Niederkorn, J.Y.; Kedzierski, W. Transgenic Mice Expressing Variants of Complement Factor H Develop AMD-like Retinal Findings. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5878–5887. [Google Scholar] [CrossRef] [PubMed]
- Tobalem, S.J.; Kecik, M.M.; Midroit, M.; Scaldino, D.; Tardy, S.; Bascuas Castillo, T.; Harmening, N.; Scapozza, L.; Thumann, G.; Kropp, M. Functional and Anatomical Characterization of Atrophic Age-Related Macular Degeneration in an Aged Mouse Model. J. Ophthalmol. Res. 2021, 4, 128. [Google Scholar] [CrossRef]
- Zhao, Z.; Chen, Y.; Wang, J.; Sternberg, P.; Freeman, M.L.; Grossniklaus, H.E.; Cai, J. Age-Related Retinopathy in NRF2-Deficient Mice. PLoS ONE 2011, 6, e19456. [Google Scholar] [CrossRef] [PubMed]
- Malek, G.; Johnson, L.V.; Mace, B.E.; Saloupis, P.; Schmechel, D.E.; Rickman, D.W.; Toth, C.A.; Sullivan, P.M.; Rickman, C.B. Apolipoprotein E Allele-Dependent Pathogenesis: A Model for Age-Related Retinal Degeneration. Proc. Natl. Acad. Sci. USA 2005, 102, 11900–11905. [Google Scholar] [CrossRef]
- Dasari, B.; Prasanthi, J.R.; Marwarha, G.; Singh, B.B.; Ghribi, O. Cholesterol-Enriched Diet Causes Age-Related Macular Degeneration-like Pathology in Rabbit Retina. BMC Ophthalmol. 2011, 11, 22. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.-C.; Ross, R.J.; Shen, D.; Ding, X.; Majumdar, Z.; Bojanowski, C.M.; Zhou, M.; Salem, N.; Bonner, R.; Tuo, J. Ccl2/Cx3cr1-Deficient Mice: An Animal Model for Age-Related Macular Degeneration. Ophthalmic Res. 2008, 40, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Ambati, J.; Anand, A.; Fernandez, S.; Sakurai, E.; Lynn, B.C.; Kuziel, W.A.; Rollins, B.J.; Ambati, B.K. An Animal Model of Age-Related Macular Degeneration in Senescent Ccl-2- or Ccr-2-Deficient Mice. Nat. Med. 2003, 9, 1390–1397. [Google Scholar] [CrossRef] [PubMed]
- Luhmann, U.F.O.; Robbie, S.; Munro, P.M.G.; Barker, S.E.; Duran, Y.; Luong, V.; Fitzke, F.W.; Bainbridge, J.W.B.; Ali, R.R.; MacLaren, R.E. The Drusenlike Phenotype in Aging Ccl2-Knockout Mice Is Caused by an Accelerated Accumulation of Swollen Autofluorescent Subretinal Macrophages. Investig. Ophthalmol. Vis. Sci. 2009, 50, 5934–5943. [Google Scholar] [CrossRef] [PubMed]
- Yasukawa, T.; Wiedemann, P.; Hoffmann, S.; Kacza, J.; Eichler, W.; Wang, Y.-S.; Nishiwaki, A.; Seeger, J.; Ogura, Y. Glycoxidized Particles Mimic Lipofuscin Accumulation in Aging Eyes: A New Age-Related Macular Degeneration Model in Rabbits. Graefes Arch. Clin. Exp. Ophthalmol. 2007, 245, 1475–1485. [Google Scholar] [CrossRef] [PubMed]
- Pennesi, M.E.; Neuringer, M.; Courtney, R.J. Animal Models of Age Related Macular Degeneration. Mol. Aspects Med. 2012, 33, 487–509. [Google Scholar] [CrossRef] [PubMed]
- Doukas, J.; Mahesh, S.; Umeda, N.; Kachi, S.; Akiyama, H.; Yokoi, K.; Cao, J.; Chen, Z.; Dellamary, L.; Tam, B.; et al. Topical Administration of a Multi-Targeted Kinase Inhibitor Suppresses Choroidal Neovascularization and Retinal Edema. J. Cell. Physiol. 2008, 216, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Yafai, Y.; Yang, X.M.; Niemeyer, M.; Nishiwaki, A.; Lange, J.; Wiedemann, P.; King, A.G.; Yasukawa, T.; Eichler, W. Anti-Angiogenic Effects of the Receptor Tyrosine Kinase Inhibitor, Pazopanib, on Choroidal Neovascularization in Rats. Eur. J. Pharmacol. 2011, 666, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Xie, W.; Hein, T.W.; Kuo, L.; Rosa, R.H. Laser-Induced Choroidal Neovascularization in Rats. Methods Mol. Biol. 2021, 2319, 77–85. [Google Scholar] [PubMed]
- Nguyen, V.P.; Henry, J.; Zhe, J.; Hu, J.; Wang, X.; Paulus, Y.M. Multimodal Imaging of Laser-Induced Choroidal Neovascularization in Pigmented Rabbits. Sci. Rep. 2023, 13, 8396. [Google Scholar] [CrossRef] [PubMed]
- Rotov, A.Y.; Astakhova, L.A.; Sitnikova, V.S.; Evdokimov, A.A.; Boitsov, V.M.; Dubina, M.V.; Ryazantsev, M.N.; Firsov, M.L. New Experimental Models of Retinal Degeneration for Screening Molecular Photochromic Ion Channel Blockers. Acta Naturae 2018, 10, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, E.M.; Emri, E.; Merle, B.M.J.; Colijn, J.M.; Kersten, E.; Cougnard-Gregoire, A.; Dammeier, S.; Meester-Smoor, M.; Pool, F.M.; de Jong, E.K.; et al. A New Perspective on Lipid Research in Age-Related Macular Degeneration. Prog. Retin. Eye Res. 2018, 67, 56–86. [Google Scholar] [CrossRef] [PubMed]
- Fattahi, N.; Shahbazi, M.-A.; Maleki, A.; Hamidi, M.; Ramazani, A.; Santos, H.A. Emerging Insights on Drug Delivery by Fatty Acid Mediated Synthesis of Lipophilic Prodrugs as Novel Nanomedicines. J. Control. Release 2020, 326, 556–598. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, J.P.; Hahn, A. Bioavailability of Long-Chain Omega-3 Fatty Acids. Prostaglandins Leukot. Essent. Fat. Acids 2013, 89, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Glick, N.R.; Fischer, M.H. The Role of Essential Fatty Acids in Human Health. J. Evid.-Based Complement. Altern. Med. 2013, 18, 268–289. [Google Scholar] [CrossRef]
- Whelan, J.; Fritsche, K. Linoleic Acid. Adv. Nutr. 2013, 4, 311–312. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Keum, Y.-S. Omega-3 and Omega-6 Polyunsaturated Fatty Acids: Dietary Sources, Metabolism, and Significance—A Review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Burns-Whitmore, B.; Froyen, E.; Heskey, C.; Parker, T.; San Pablo, G. Alpha-Linolenic and Linoleic Fatty Acids in the Vegan Diet: Do They Require Dietary Reference Intake/Adequate Intake Special Consideration? Nutrients 2019, 11, 2365. [Google Scholar] [CrossRef] [PubMed]
- Kishan, A.U.; Modjtahedi, B.S.; Martins, E.N.; Modjtahedi, S.P.; Morse, L.S. Lipids and Age-Related Macular Degeneration. Surv. Ophthalmol. 2011, 56, 195–213. [Google Scholar] [CrossRef] [PubMed]
- Hopiavuori, B.R.; Bennett, L.D.; Brush, R.S.; Hook, M.J.V.; Thoreson, W.B.; Anderson, R.E. Very Long-Chain Fatty Acids Support Synaptic Structure and Function in the Mammalian Retina. OCL 2016, 23, D113. [Google Scholar] [CrossRef]
- SanGiovanni, J.P.; Chew, E.Y. The Role of Omega-3 Long-Chain Polyunsaturated Fatty Acids in Health and Disease of the Retina. Prog. Retin. Eye Res. 2005, 24, 87–138. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, M.G.; Li, J.; Alvarez, R.A.; Anderson, R.E.; O’Brien, P.J. Metabolism of Linolenic Acid and Docosahexaenoic Acid in Rat Retinas and Rod Outer Segments. Exp. Eye Res. 1991, 53, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Agbaga, M.-P.; Mandal, M.N.A.; Anderson, R.E. Retinal Very Long-Chain PUFAs: New Insights from Studies on ELOVL4 Protein. J. Lipid Res. 2010, 51, 1624–1642. [Google Scholar] [CrossRef]
- Hopiavuori, B.R.; Anderson, R.E.; Agbaga, M.-P. ELOVL4: Very Long-Chain Fatty Acids Serve an Eclectic Role in Mammalian Health and Function. Prog. Retin. Eye Res. 2019, 69, 137–158. [Google Scholar] [CrossRef] [PubMed]
- Sassa, T.; Kihara, A. Metabolism of Very Long-Chain Fatty Acids: Genes and Pathophysiology. Biomol. Ther. 2014, 22, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Benham, A.; Logan, S.; Brush, R.S.; Mandal, M.N.A.; Anderson, R.E.; Agbaga, M.-P. ELOVL4 Protein Preferentially Elongates 20:5n3 to Very Long Chain PUFAs over 20:4n6 and 22:6n3. J. Lipid Res. 2012, 53, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Garagnani, P.; Bacalini, M.G.; Pirazzini, C.; Gori, D.; Giuliani, C.; Mari, D.; Di Blasio, A.M.; Gentilini, D.; Vitale, G.; Collino, S.; et al. Methylation of ELOVL2 Gene as a New Epigenetic Marker of Age. Aging Cell 2012, 11, 1132–1134. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.-A.; Hammer, R.E.; Horton, J.D. Deletion of ELOVL5 Leads to Fatty Liver through Activation of SREBP-1c in Mice. J. Lipid Res. 2009, 50, 412–423. [Google Scholar] [CrossRef] [PubMed]
- Logan, S.; Anderson, R.E. Dominant Stargardt Macular Dystrophy (STGD3) and ELOVL4. Adv. Exp. Med. Biol. 2014, 801, 447–453. [Google Scholar] [PubMed]
- Deák, F.; Anderson, R.E.; Fessler, J.L.; Sherry, D.M. Novel Cellular Functions of Very Long Chain-Fatty Acids: Insight From ELOVL4 Mutations. Front. Cell. Neurosci. 2019, 13, 428. [Google Scholar] [CrossRef] [PubMed]
- Swinkels, D.; Baes, M. The Essential Role of Docosahexaenoic Acid and Its Derivatives for Retinal Integrity. Pharmacol. Ther. 2023, 247, 108440. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Song, J.-T. Potential Mechanisms of Macular Degeneration Protection by Fatty Fish Consumption. Curr. Opin. Pharmacol. 2022, 63, 102186. [Google Scholar] [CrossRef] [PubMed]
- Agbaga, M.-P.; Merriman, D.K.; Brush, R.S.; Lydic, T.A.; Conley, S.M.; Naash, M.I.; Jackson, S.; Woods, A.S.; Reid, G.E.; Busik, J.V.; et al. Differential Composition of DHA and Very-Long-Chain PUFAs in Rod and Cone Photoreceptors. J. Lipid Res. 2018, 59, 1586–1596. [Google Scholar] [CrossRef] [PubMed]
- Rotstein, N.P.; Politi, L.E.; German, O.L.; Girotti, R. Protective Effect of Docosahexaenoic Acid on Oxidative Stress-Induced Apoptosis of Retina Photoreceptors. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2252–2259. [Google Scholar] [CrossRef] [PubMed]
- Van Kuijk, F.J.; Buck, P. Fatty Acid Composition of the Human Macula and Peripheral Retina. Investig. Ophthalmol. Vis. Sci. 1992, 33, 3493–3496. [Google Scholar]
- Liu, A.; Lin, Y.; Terry, R.; Nelson, K.; Bernstein, P.S. Role of Long-Chain and Very-Long-Chain Polyunsaturated Fatty Acids in Macular Degenerations and Dystrophies. Clin. Lipidol. 2011, 6, 593–613. [Google Scholar] [CrossRef] [PubMed]
- Shindou, H.; Koso, H.; Sasaki, J.; Nakanishi, H.; Sagara, H.; Nakagawa, K.M.; Takahashi, Y.; Hishikawa, D.; Iizuka-Hishikawa, Y.; Tokumasu, F.; et al. Docosahexaenoic Acid Preserves Visual Function by Maintaining Correct Disc Morphology in Retinal Photoreceptor Cells. J. Biol. Chem. 2017, 292, 12054–12064. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Marcheselli, V.L.; Serhan, C.N.; Bazan, N.G. Neuroprotectin D1: A Docosahexaenoic Acid-Derived Docosatriene Protects Human Retinal Pigment Epithelial Cells from Oxidative Stress. Proc. Natl. Acad. Sci. USA 2004, 101, 8491–8496. [Google Scholar] [CrossRef] [PubMed]
- Connor, K.M.; SanGiovanni, J.P.; Lofqvist, C.; Aderman, C.M.; Chen, J.; Higuchi, A.; Hong, S.; Pravda, E.A.; Majchrzak, S.; Carper, D.; et al. Increased Dietary Intake of Omega-3-Polyunsaturated Fatty Acids Reduces Pathological Retinal Angiogenesis. Nat. Med. 2007, 13, 868–873. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.; Oliveira, L.T.; Oger, C.; Galano, J.-M.; Bultel-Poncé, V.; Richard, S.; Guimaraes, A.G.; Vilela, J.M.C.; Andrade, M.S.; Durand, T.; et al. Polymeric Nanocapsules Prevent Oxidation of Core-Loaded Molecules: Evidence Based on the Effects of Docosahexaenoic Acid and Neuroprostane on Breast Cancer Cells Proliferation. J. Exp. Clin. Cancer Res. 2015, 34, 155. [Google Scholar] [CrossRef] [PubMed]
- Merle, B.; Buaud, B.; Korobelnik, J.-F.; Bron, A.; Delyfer, M.-N.; Rougier, M.-B.; Savel, H.; Vaysse, C.; Creuzot-Garcher, C.; Delcourt, C. Plasma Long-Chain Omega-3 Polyunsaturated Fatty Acids and Macular Pigment in Subjects with Family History of Age-Related Macular Degeneration: The Limpia Study. Acta Ophthalmol. 2017, 95, e763–e769. [Google Scholar] [CrossRef] [PubMed]
- Bennett, L.D.; Hopiavuori, B.R.; Brush, R.S.; Chan, M.; Van Hook, M.J.; Thoreson, W.B.; Anderson, R.E. Examination of VLC-PUFA-Deficient Photoreceptor Terminals. Investig. Ophthalmol. Vis. Sci. 2014, 55, 4063–4072. [Google Scholar] [CrossRef] [PubMed]
- Catalá, A. An Overview of Lipid Peroxidation with Emphasis in Outer Segments of Photoreceptors and the Chemiluminescence Assay. Int. J. Biochem. Cell Biol. 2006, 38, 1482–1495. [Google Scholar] [CrossRef] [PubMed]
- Yakubenko, V.P.; Byzova, T.V. Biological and Pathophysiological Roles of End-Products of DHA Oxidation. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Romero-Vazquez, S.; Llorens, V.; Soler-Boronat, A.; Figueras-Roca, M.; Adan, A.; Molins, B. Interlink between Inflammation and Oxidative Stress in Age-Related Macular Degeneration: Role of Complement Factor H. Biomedicines 2021, 9, 763. [Google Scholar] [CrossRef]
- Kaarniranta, K.; Uusitalo, H.; Blasiak, J.; Felszeghy, S.; Kannan, R.; Kauppinen, A.; Salminen, A.; Sinha, D.; Ferrington, D. Mechanisms of Mitochondrial Dysfunction and Their Impact on Age-Related Macular Degeneration. Prog. Retin. Eye Res. 2020, 79, 100858. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Eicosanoids. Essays Biochem. 2020, 64, 423–441. [Google Scholar] [PubMed]
- Thierry, M. Rôle des AGPI Alimentaires dans L’homéostasie Lipidique de la Rétine en Conditions Physiologiques et Pathologiques Liées au Vieillissement. Ph.D. Thesis, Université de Bourgogne, Dijon, France, 2014. [Google Scholar]
- Skowronska-Krawczyk, D.; Chao, D.L. Long-Chain Polyunsaturated Fatty Acids and Age-Related Macular Degeneration. Adv. Exp. Med. Biol. 2019, 1185, 39–43. [Google Scholar] [PubMed]
- Phillis, J.W.; Horrocks, L.A.; Farooqui, A.A. Cyclooxygenases, Lipoxygenases, and Epoxygenases in CNS: Their Role and Involvement in Neurological Disorders. Brain Res. Rev. 2006, 52, 201–243. [Google Scholar] [CrossRef] [PubMed]
- Koto, T.; Nagai, N.; Kurihara, T.; Izumi-Nagai, K.; Satofuka, S.; Shinoda, H.; Noda, K.; Ozawa, Y.; Inoue, M.; Tsubota, K.; et al. Eicosapentaenoic Acid Is Anti-Inflammatory in Preventing Choroidal Neovascularization in Mice. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4328–4334. [Google Scholar] [CrossRef]
- Seregina, M.; Lydic, T.; Opreanu, M.; Li Calzi, S.; Bozack, S.; McSorley, K.; Sochacki, A.; Faber, M.; Hazra, S.; Duclos, S.; et al. N-3 Polyunsaturated Fatty Acids Prevent Diabetic Retinopathy by Inhibition of Retinal Vascular Damage and Enhanced Endothelial Progenitor Cell Reparative Function. PLoS ONE 2013, 8, e55177. [Google Scholar]
- So, J.; Wu, D.; Lichtenstein, A.H.; Tai, A.K.; Matthan, N.R.; Maddipati, K.R.; Lamon-Fava, S. EPA and DHA Differentially Modulate Monocyte Inflammatory Response in Subjects with Chronic Inflammation in Part via Plasma Specialized Pro-Resolving Lipid Mediators: A Randomized, Double-Blind, Crossover Study. Atherosclerosis 2021, 316, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Jun, B.; Mukherjee, P.K.; Asatryan, A.; Kautzmann, M.-A.; Heap, J.; Gordon, W.C.; Bhattacharjee, S.; Yang, R.; Petasis, N.A.; Bazan, N.G. Elovanoids Are Novel Cell-Specific Lipid Mediators Necessary for Neuroprotective Signaling for Photoreceptor Cell Integrity. Sci. Rep. 2017, 7, 5279. [Google Scholar] [CrossRef] [PubMed]
- Leung, H.H.; Ng, A.L.; Durand, T.; Kawasaki, R.; Oger, C.; Balas, L.; Galano, J.-M.; Wong, I.Y.; Chung-Yung Lee, J. Increase in Omega-6 and Decrease in Omega-3 Polyunsaturated Fatty Acid Oxidation Elevates the Risk of Exudative AMD Development in Adults with Chinese Diet. Free Radic. Biol. Med. 2019, 145, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Punia, S.; Sandhu, K.S.; Siroha, A.K.; Dhull, S.B. Omega 3-Metabolism, Absorption, Bioavailability and Health Benefits–A Review. PharmaNutrition 2019, 10, 100162. [Google Scholar] [CrossRef]
- Wu, J.; Cho, E.; Giovannucci, E.; Rosner, B.; Sastry, S.; Willett, W.; Schaumberg, D. Dietary Intakes of Eicosapentaenoic Acid and Docosahexaenoic Acid and Risk of Age-Related Macular Degeneration. Ophthalmology 2017, 124, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Ren, A.; Deng, X.; Huang, Z.; Jiang, Z.; Li, Z.; Gong, Y. Long-Chain Polyunsaturated Fatty Acids and Their Metabolites Regulate Inflammation in Age-Related Macular Degeneration. J. Inflamm. Res. 2022, 15, 865–880. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Shi, X.; Fan, Y.; Wang, D.; Li, B.; Zhou, J.; Pei, C.; Ma, L. Dietary Omega-3 Polyunsaturated Fatty Acids and Fish Intake and Risk of Age-Related Macular Degeneration. Clin. Nutr. 2021, 40, 5662–5673. [Google Scholar] [CrossRef]
- Chong, E.; Kreis, A.; Wong, T.-Y.; Simpson, J.; Guymer, R. Dietary ω-3 Fatty Acid and Fish Intake in the Primary Prevention of Age-Related Macular Degeneration: A Systematic Review and Meta-Analysis. Arch. Ophthalmol. 2008, 126, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Csader, S.; Korhonen, S.; Kaarniranta, K.; Schwab, U. The Effect of Dietary Supplementations on Delaying the Progression of Age-Related Macular Degeneration: A Systematic Review and Meta-Analysis. Nutrients 2022, 14, 4273. [Google Scholar] [CrossRef] [PubMed]
- Chong, E.W.-T.; Robman, L.D.; Simpson, J.A.; Hodge, A.M.; Aung, K.Z.; Dolphin, T.K.; English, D.R.; Giles, G.G.; Guymer, R.H. Fat Consumption and Its Association with Age-Related Macular Degeneration. Arch. Ophthalmol. 2009, 127, 674–680. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Wang, J.; Flood, V.; Mitchell, P. Dietary Fatty Acids and the 10-Year Incidence of Age-Related Macular Degeneration the Blue Mountains Eye Study. Arch. Ophthalmol. 2009, 127, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Agrón, E.; Mares, J.; Clemons, T.E.; Swaroop, A.; Chew, E.Y.; Keenan, T.D.L. Dietary Nutrient Intake and Progression to Late Age-Related Macular Degeneration in the Age-Related Eye Disease Studies 1 and 2. Ophthalmology 2021, 128, 425–442. [Google Scholar] [CrossRef] [PubMed]
- Hogg, R.E.; Woodside, J.V.; McGrath, A.; Young, I.S.; Vioque, J.L.; Chakravarthy, U.; de Jong, P.T.; Rahu, M.; Seland, J.; Soubrane, G.; et al. Mediterranean Diet Score and Its Association with Age-Related Macular Degeneration: The European Eye Study. Ophthalmology 2017, 124, 82–89. [Google Scholar] [CrossRef] [PubMed]
- SanGiovanni, J.P.; Agrón, E.; Meleth, A.; Reed, G.; Sperduto, R.; Clemons, T.; Chew, E. ω-3 Long-Chain Polyunsaturated Fatty Acid Intake and 12-y Incidence of Neovascular Age-Related Macular Degeneration and Central Geographic Atrophy: AREDS Report 30, a Prospective Cohort Study from the Age-Related Eye Disease Study. Am. J. Clin. Nutr. 2009, 90, 1601–1607. [Google Scholar] [CrossRef] [PubMed]
- Age-Related Eye Disease Study 2 Research Group. Lutein + Zeaxanthin and Omega-3 Fatty Acids for Age-Related Macular Degeneration: The Age-Related Eye Disease Study 2 (AREDS2) Randomized Clinical Trial. JAMA 2013, 309, 2005–2015. [Google Scholar] [CrossRef] [PubMed]
- Souied, E.H.; Aslam, T.; Garcia-Layana, A.; Holz, F.G.; Leys, A.; Silva, R.; Delcourt, C. Omega-3 Fatty Acids and Age-Related Macular Degeneration. Ophthalmic Res. 2015, 55, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Seddon, J.M.; George, S.; Rosner, B. Cigarette Smoking, Fish Consumption, Omega-3 Fatty Acid Intake, and Associations with Age-Related Macular Degeneration: The US Twin Study of Age-Related Macular Degeneration. Arch. Ophthalmol. 2006, 124, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Cho, E.; Giovannucci, E.; Rosner, B.; Sastry, S.; Schaumberg, D.; Willett, W. Dietary Intake of α-Linolenic Acid and Risk of Age-Related Macular Degeneration. Am. J. Clin. Nutr. 2017, 105, 1483–1492. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhong, Y.; Yang, F.; Hu, C.; Liu, X.; Zhu, Y.; Yao, K. Causal Effects of N-6 Polyunsaturated Fatty Acids on Age-Related Macular Degeneration: A Mendelian Randomization Study. J. Clin. Endocrinol. Metab. 2021, 106, e3565–e3572. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Harada, S.; Tsubota, K.; Yasukawa, T.; Takebayashi, T.; Nishiwaki, Y.; Kawasaki, R. Dietary Saturated Fatty Acid Intake and Early Age-Related Macular Degeneration in a Japanese Population. Investig. Ophthalmol. Vis. Sci. 2020, 61, 23. [Google Scholar] [CrossRef] [PubMed]
- Cougnard-Grégoire, A.; Merle, B.M.J.; Korobelnik, J.-F.; Rougier, M.-B.; Delyfer, M.-N.; Le Goff, M.; Samieri, C.; Dartigues, J.-F.; Delcourt, C. Olive Oil Consumption and Age-Related Macular Degeneration: The Alienor Study. PLoS ONE 2016, 11, e0160240. [Google Scholar] [CrossRef] [PubMed]
- Delcourt, C.; Carrière, I.; Cristol, J.-P.; Lacroux, A.; Gerber, M. Dietary Fat and the Risk of Age-Related Maculopathy: The POLANUT Study. Eur. J. Clin. Nutr. 2007, 61, 1341–1344. [Google Scholar] [CrossRef] [PubMed]
- ANSES. Actualisation des Apports Nutritionnels Conseillés pour les Acides Gras—Version Intégrant les Modifications Apportées par L’erratum du 28 Juillet 2011; ANSES: Paris, France, 2011; p. 323. [Google Scholar]
- Merle, B.; Delyfer, M.-N.; Korobelnik, J.-F.; Rougier, M.-B.; Colin, J.; Malet, F.; Féart, C.; Le Goff, M.; Dartigues, J.-F.; Barberger-Gateau, P.; et al. Dietary Omega-3 Fatty Acids and the Risk for Age-Related Maculopathy: The Alienor Study. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6004–6011. [Google Scholar] [CrossRef] [PubMed]
- Souied, E.; Delcourt, C.; Querques, G.; Merle, B.; Smith, T.; Benlian, P. NAT2 study group NAT2 study; omega-3 levels in red blood cells membranes correlates the preventive effect. Investig. Ophthalmol. Vis. Sci. 2013, 54, 3277. [Google Scholar]
- Merle, B.M.J.; Colijn, J.M.; Cougnard-Grégoire, A.; de Koning-Backus, A.P.M.; Delyfer, M.-N.; Kiefte-de Jong, J.C.; Meester-Smoor, M.; Féart, C.; Verzijden, T.; Samieri, C.; et al. Mediterranean Diet and Incidence of Advanced Age-Related Macular Degeneration: The EYE-RISK Consortium. Ophthalmology 2019, 126, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Acar, N.; Merle, B.M.J.; Ajana, S.; He, Z.; Grégoire, S.; Hejblum, B.P.; Martine, L.; Buaud, B.; Bron, A.M.; Creuzot-Garcher, C.P.; et al. Predicting the Retinal Content in Omega-3 Fatty Acids for Age-related Macular-degeneration. Clin. Transl. Med. 2021, 11, e404. [Google Scholar] [CrossRef] [PubMed]
- Abreu-González, R.; Díaz-Rodríguez, R.; Abreu-Reyes, J.A. Nutritional Supplements with Omega 3 in Retinal Disease: Relationship between Volume and Concentration in Commercial Products. Arch. Soc. Esp. Oftalmol. 2018, 93, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Othman, K.; Cercy, C.; Mohamed, A.; Doly, M.; Ranchon-Cole, I. Dietary Supplement Enriched in Antioxidants and Omega-3 Protects from Progressive Light-Induced Retinal Degeneration. PLoS ONE 2015, 10, e0128395. [Google Scholar]
- Dey, T.K.; Koley, H.; Ghosh, M.; Dey, S.; Dhar, P. Effects of Nano-Sizing on Lipid Bioaccessibility and Ex Vivo Bioavailability from EPA-DHA Rich Oil in Water Nanoemulsion. Food Chem. 2019, 275, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Ghasemifard, S.; Turchini, G.M.; Sinclair, A.J. Omega-3 Long Chain Fatty Acid “Bioavailability”: A Review of Evidence and Methodological Considerations. Prog. Lipid Res. 2014, 56, 92–108. [Google Scholar] [CrossRef] [PubMed]
- Lapointe, J.-F.; Harvey, L.; Aziz, S.; Jordan, H.; Hegele, R.A.; Lemieux, P. A Single-Dose, Comparative Bioavailability Study of a Formulation Containing OM3 as Phospholipid and Free Fatty Acid to an Ethyl Ester Formulation in the Fasting and Fed States. Clin. Ther. 2019, 41, 426–444. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Administration Mode | Mechanism of Action | Refs. | ||
|---|---|---|---|---|---|
| Wet AMD | |||||
| Commercialized (Europe) | Laser photocoagulation | Local laser | Destruction of extrafoveolar neovessels under effect of temperature (thermal argon laser). | [59,72] | |
| Verteporfin (Visudyne®) + photodynamic therapy | Oral administration + local laser | Destruction of neovessels by applying local infrared radiation to a photosensitive molecule. | [59] | ||
| Anti-VEGF | Ranibizumab (Lucentis®), Bevacizumab (Avastin®) 1, Aflibercept (Eylea®), Brolucizumab (Beovu®), Faricimab (Vabysmo®) | Intravitreal injection | Inhibition of VEGF (mainly VEGF-A) by binding to them and blocking their interactions with receptors. | [73,74,75,76] | |
| In development (worldwide) | Anti-VEGF | KSI-301 (phase III done, waiting for results) | Intravitreal injection | Inhibition of VEGF (mainly VEGF-A). | [77,78] |
| Tyrosine kinase inhibitors | Vorolanib (Eyp-1901, phase II ongoing) | Intravitreal injection | Inhibition of VEGFR tyrosine kinase. | [79] | |
| Axitinib (CLS-AX, phase II ongoing) | Suprachoroidal injection | [80] | |||
| Axitinib (OTX-TKI, phase III ongoing) | Ocular implant | [81] | |||
| Anti-complements | Iptacopan (phase II ongoing) | Oral administration | Inhibition of C5 or complement factor B. | [82] | |
| Anti-VEGF and anti-complement | IBI302 (phase II ongoing) | Intravitreal injections | Bispecific fusion protein acting as a decoy: inhibition of VEGF-mediated signaling pathway and complement factors C3b and C4b. | [83] | |
| Gene therapy | RGX-314 (phase II ongoing) | Subretinal injection | Adeno-associated virus (AAV) vector delivering a gene encoding for anti-VEGF Fab portion. | [84] | |
| ADVM-022 (phase II ongoing) | Intravitreal injection | AAV vector encoding aflibercept. | [85] | ||
| RetinoStat® (phase I ongoing) | Subretinal injection | Lentivirus delivering anti-angiogenic genes (endostatins and angiostatin). | [86] | ||
| Dry AMD | |||||
| Commercialized (Europe) | Food supplements | Macular pigments, vitamins, minerals, antioxidants, fatty acids | Oral administration | Supplementation of minerals and fatty acids necessary for the development and function of photoreceptors. | [87,88] |
| In evaluation by the European Medicines Agency | Anti-complements | Pegcetacoplan (Syfovre®) 2 | Intravitreal injection | Inhibition of C3 complement factors. | [89] |
| Avacincaptad pegol (Izervay®) | Intravitreal injection | Inhibition of C5 complement factors. | [90] | ||
| In development (worldwide) | Vitamin supplements | ALK-001 (phase III ongoing) | Oral administration | Replacement of natural vitamin A by chemically modified vitamin A to prevent formation of vitamin A dimers. | [91] |
| Anti-TNFα | ONL-1204 (phase I ongoing) | Intravitreal injection | Inhibition of Fas receptor (TNFalpha family). | [92] | |
| Anti-complements | IONIS-FB-LRx (phase II ongoing) | Subcutaneous administration | Inhibition of complement factor B. | [93] | |
| Danicopan (ALXN2040, phase II ongoing) | Oral administration | Inhibition of complement factor D. | [94] | ||
| Gene therapy | GT005 (phase II ongoing) | Subretinal injection | AAV vector encoding for complement factor I proteins. | [95] | |
| Cellular therapy | OpRegen (phase I/IIa ongoing) | Subretinal surgery | Administration of human embryonic stem cell to regenerate retinal pigment epithelial. | [96] | |
| Retinal implant + medical device (glasses): Prima technology | Subretinal implant | Implantation of a photovoltaic matrix which simulates photoreceptors. | [97,98,99] | ||
| Study | Objective | Stage of AMD | Number of Participants | Conclusion | References |
|---|---|---|---|---|---|
| Blue Mountain Eye Study | To assess the relationship between baseline dietary fatty acids and 10-year incident AMD. | Early | 2454 | Protective effect against early AMD with regular fish consumption, high consumption of omega-3 PUFA and low intake of foods rich in linoleic acid. | [194] |
| Polanut study | To assess the associations of dietary fat with the risk of ARM. | Early | 832 | Increased risk of ARM with high total, saturated and monounsaturated fat intake. Reduced risk (60%) of ARM with fatty fish intake (more than once a month). | [205] |
| Alienor study | To report the associations of ARM with past dietary intake in elderly French subjects. | Early | 1289 | Decreased risk of ARM in subjects with high intake of omega-3 LC-PUFA. | [207] |
| Melbourne Collaborative Cohort Study | To evaluate the association between fat intake and prevalence of AMD. | Early and late | 6734 | Decreased risk of AMD with a diet low in trans-unsaturated fat and rich in omega-3 fatty acids and olive oil. | [193] |
| Nurses’ health study and the health professional follow-up study | To evaluate the association between intake of EPA and DHA and the intermediate and advanced stages of AMD. | Intermediate and late | 114,850 | Prevention or delay of visually significant intermediate AMD with higher intake of EPA. No association with advanced AMD. | [188] |
| US twin study of AMD | To evaluate modifiable risk and protective factors for AMD among elderly twins. | Intermediate and late | 681 | Increased risk of AMD with cigarette smoking. Reduced risk of AMD with fish consumption and omega-3 fatty acid intake. | [200] |
| NAT2 study | To evaluate the efficacy of DHA-enriched oral supplementation in preventing exudative AMD. | Late | 263 | Reduction of CNV incidence with high EPA plus DHA index. | [208] |
| Eye-risk consortium | To investigate association of adherence to the Mediterranean diet with incidence of advanced AMD. | Late | 4996 | Risk reduction (41%) of incidence of advanced AMD with higher adherence to the Mediterranean diet. | [209] |
| European Eye study | To examine association between adherence to a Mediterranean diet and prevalence of AMD in countries ranging from Southern to Northern Europe. | Late | 5060 | Protective effect of adherence to a Mediterranean diet in late AMD patients. | [196] |
| AREDS | To investigate whether omega-3 LC-PUFA intake was associated with a reduced likelihood of developing central GA and neovascular AMD. | Late | 1837 | Lower 12-y incidence of central GA and neovascular AMD in participants at moderate-to-high risk for those reporting the highest consumption of omega-3 LC-PUFA. | [197] |
| AREDS 2 | To determine whether adding lutein + zeaxanthin, DHA + EPA or both to the AREDS formulation decreases the risk of developing advanced AMD and to evaluate the effect of eliminating beta carotene, lowering zinc doses or both in the AREDS formulation. | Late | 4203 | No risk reduction of advanced AMD with addition of lutein + zeaxanthin, DHA + EPA or both to the AREDS formulation. | [198] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brito, M.; Sorbier, C.; Mignet, N.; Boudy, V.; Borchard, G.; Vacher, G. Understanding the Impact of Polyunsaturated Fatty Acids on Age-Related Macular Degeneration: A Review. Int. J. Mol. Sci. 2024, 25, 4099. https://doi.org/10.3390/ijms25074099
Brito M, Sorbier C, Mignet N, Boudy V, Borchard G, Vacher G. Understanding the Impact of Polyunsaturated Fatty Acids on Age-Related Macular Degeneration: A Review. International Journal of Molecular Sciences. 2024; 25(7):4099. https://doi.org/10.3390/ijms25074099
Chicago/Turabian StyleBrito, Maëlis, Capucine Sorbier, Nathalie Mignet, Vincent Boudy, Gerrit Borchard, and Gaëlle Vacher. 2024. "Understanding the Impact of Polyunsaturated Fatty Acids on Age-Related Macular Degeneration: A Review" International Journal of Molecular Sciences 25, no. 7: 4099. https://doi.org/10.3390/ijms25074099
APA StyleBrito, M., Sorbier, C., Mignet, N., Boudy, V., Borchard, G., & Vacher, G. (2024). Understanding the Impact of Polyunsaturated Fatty Acids on Age-Related Macular Degeneration: A Review. International Journal of Molecular Sciences, 25(7), 4099. https://doi.org/10.3390/ijms25074099