
The Oligomeric State of Vasorin in the Plasma Membrane Measured Non-Invasively by Quantitative Fluorescence Fluctuation Spectroscopy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
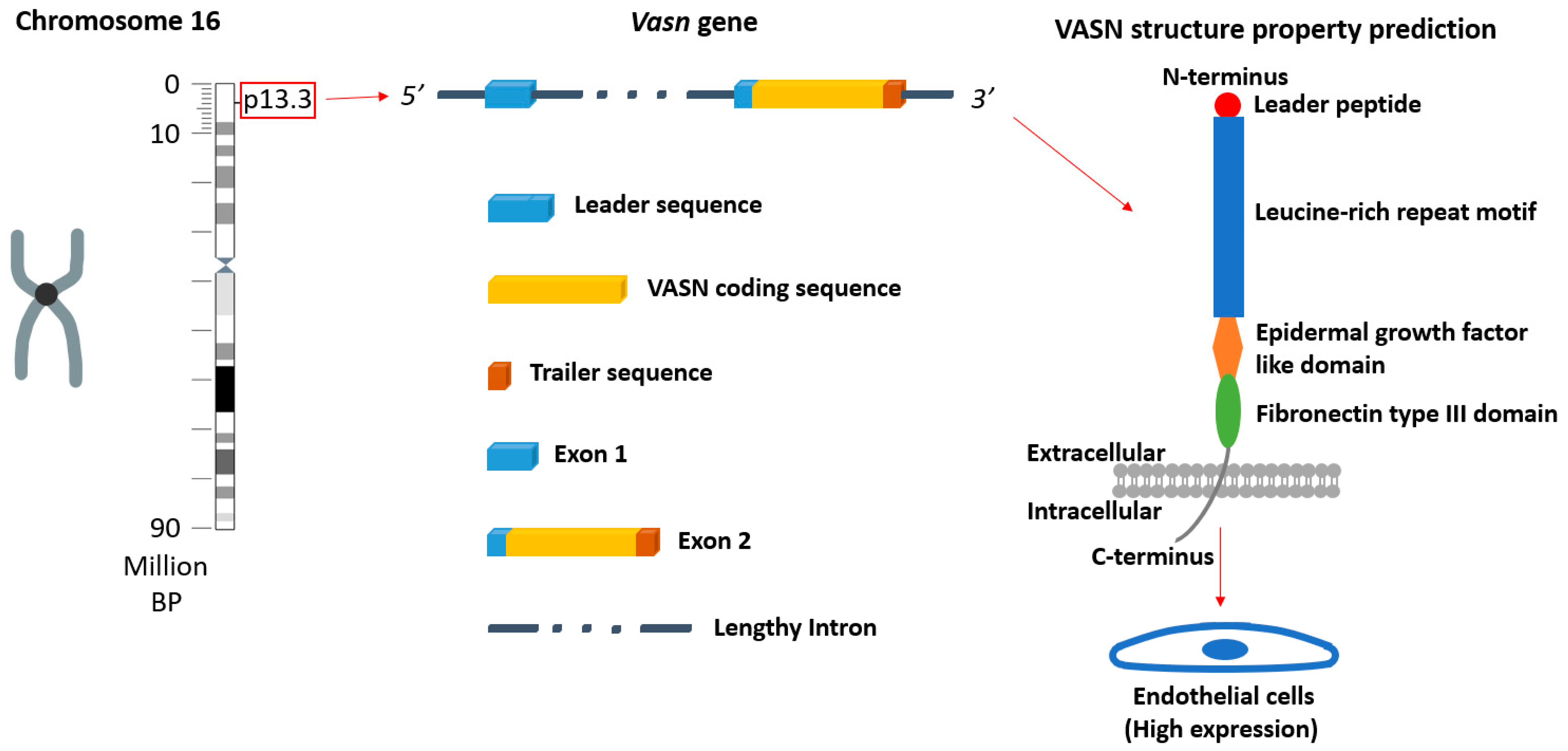
:1. Introduction
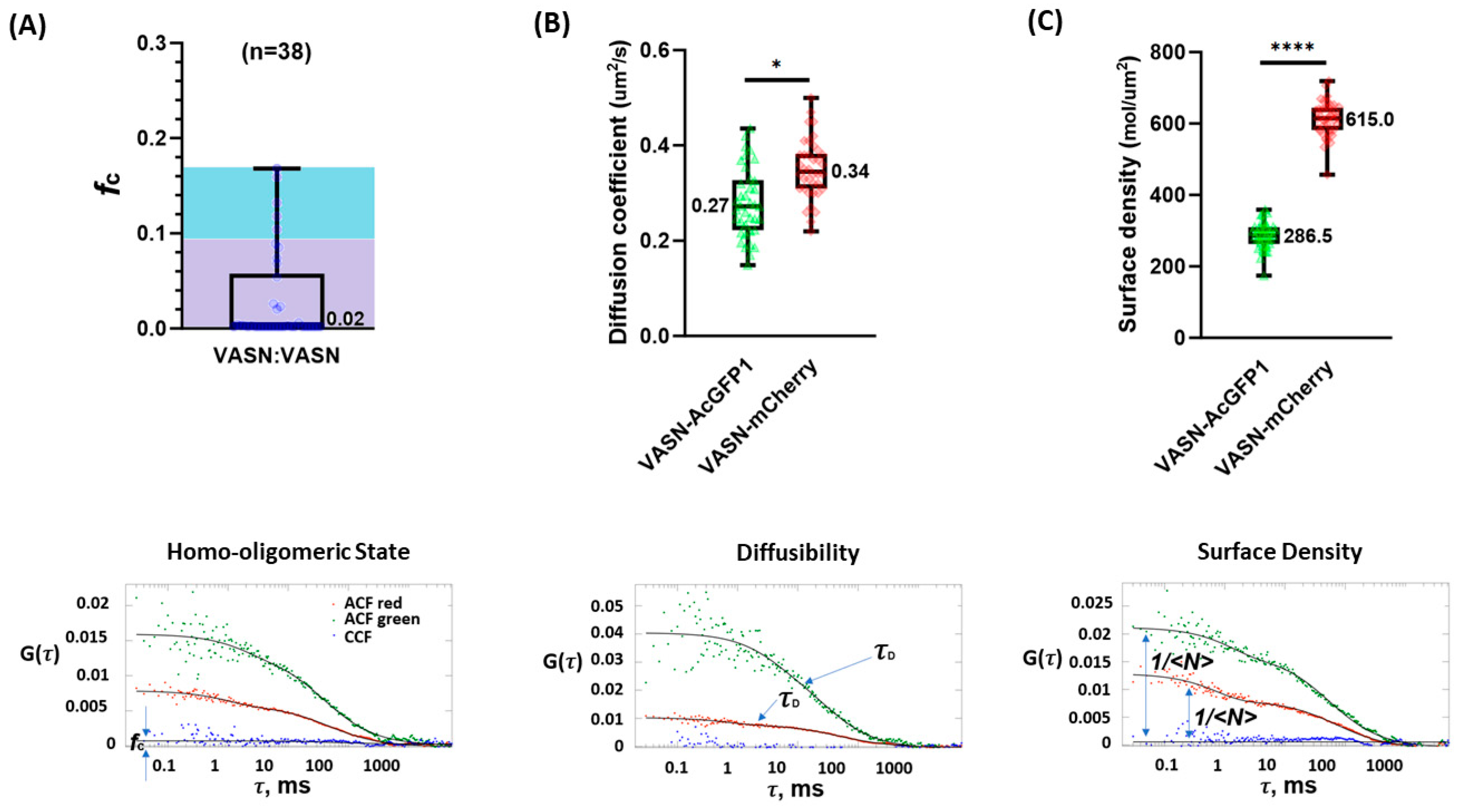
2. Results
3. Discussion
4. Methods and Materials
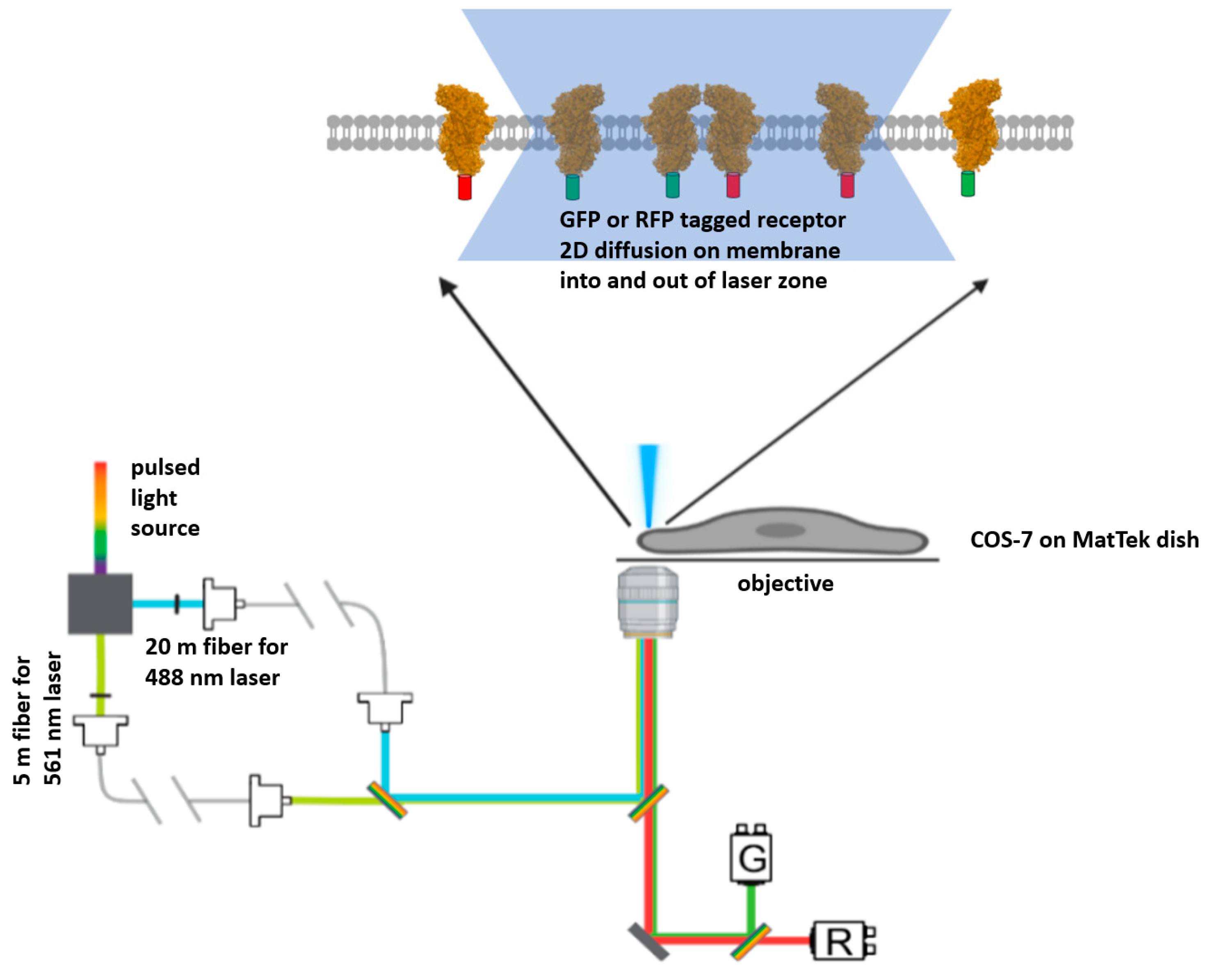
4.1. Pulsed Interleaved Excitation Fluorescence Cross-Correlation Spectroscopy (PIE-FCCS) and Experimental Description
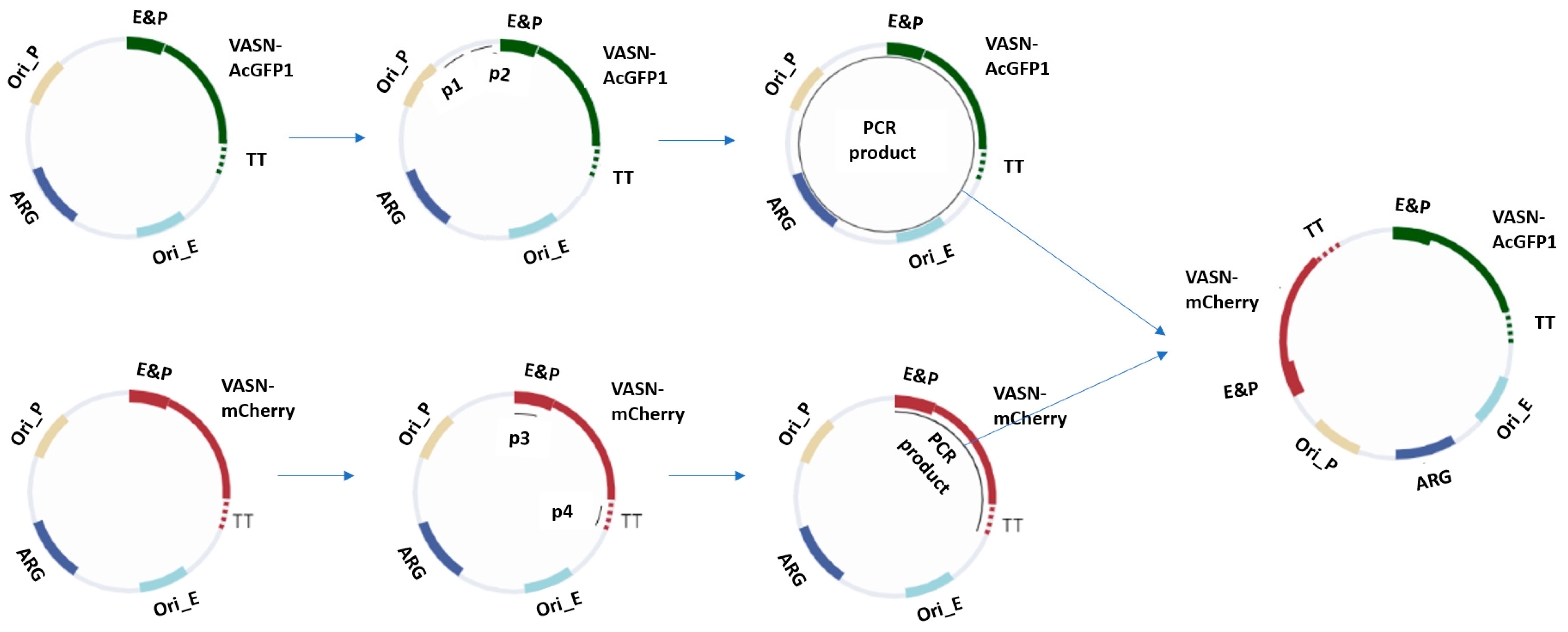
4.2. Construct Architecture
4.3. COS-7 Culturing and Transfection
4.4. FCCS Data Gathering and Processing
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ikeda, Y.; Imai, Y.; Kumagai, H.; Nosaka, T.; Morikawa, Y.; Hisaoka, T.; Manabe, I.; Maemura, K.; Nakaoka, T.; Imamura, T.; et al. Vasorin, a transforming growth factor β-binding protein expressed in vascular smooth muscle cells, modulates the arterial response to injury in vivo. Proc. Natl. Acad. Sci. USA 2004, 101, 10732–10737. [Google Scholar] [CrossRef] [PubMed]
- Louvet, L.; Lenglet, G.; Krautzberger, A.M.; Mentaverri, R.; Hague, F.; Kowalewski, C.; Mahtal, N.; Lesieur, J.; Bonnet, A.; Andrique, C.; et al. Vasorin plays a critical role in vascular smooth muscle cells and arterial functions. J. Cell. Physiol. 2022, 237, 3845–3859. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, A.-L.; Chaussain, C.; Broutin, I.; Rochefort, G.Y.; Schrewe, H.; Gaucher, C. From Vascular Smooth Muscle Cells to Folliculogenesis: What About Vasorin? Front. Med. 2018, 5, 335. [Google Scholar] [CrossRef] [PubMed]
- Luong, T.T.D.; Estepa, M.; Boehme, B.; Pieske, B.; Lang, F.; Eckardt, K.-U.; Voelkl, J.; Alesutan, I. Inhibition of vascular smooth muscle cell calcification by vasorin through interference with TGFβ1 signaling. Cell. Signal. 2019, 64, 109414. [Google Scholar] [CrossRef] [PubMed]
- Korte, L.; Widmer-Teske, R.; Donde, K.; Dutzmann, J.; Daniel, J.M.; Bauersachs, J.; Sedding, D.G. 13Vasorin controls smooth muscle cell proliferation by regulating EGFR activation. Cardiovasc. Res. 2018, 114 (Suppl. S1), S3. [Google Scholar] [CrossRef]
- Liu, J.; Tang, H.; Xu, C.; Zhou, S.; Zhu, X.; Li, Y.; Prézeau, L.; Xu, T.; Pin, J.-P.; Rondard, P.; et al. Biased signaling due to oligomerization of the G protein-coupled platelet-activating factor receptor. Nat. Commun. 2022, 13, 6365. [Google Scholar] [CrossRef] [PubMed]
- Breitwieser, G.E. G Protein–Coupled Receptor Oligomerization: Implications for G Protein Activation and Cell Signaling. Circ. Res. 2004, 94, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Naval, J.; De Miguel, D.; Gallego-Lleyda, A.; Anel, A.; Martinez-Lostao, L. Importance of TRAIL Molecular Anatomy in Receptor Oligomerization and Signaling. Implications for Cancer Therapy. Cancers 2019, 11, 444. [Google Scholar] [CrossRef] [PubMed]
- Caré, B.R.; Soula, H.A. Impact of receptor clustering on ligand binding. BMC Syst. Biol. 2011, 5, 48. [Google Scholar] [CrossRef]
- Zhang, Y.; DeVries, M.E.; Skolnick, J. Structure Modeling of All Identified G Protein–Coupled Receptors in the Human Genome. PLoS Comput. Biol. 2006, 2, e13. [Google Scholar] [CrossRef]
- Ellaithy, A.; Gonzalez-Maeso, J.; Logothetis, D.A.; Levitz, J. Structural and Biophysical Mechanisms of Class C G Protein-Coupled Receptor Function. Trends Biochem. Sci. 2020, 45, 1049–1064. [Google Scholar] [CrossRef]
- Chun, L.; Zhang, W.; Liu, J. Structure and ligand recognition of class C GPCRs. Acta Pharmacol. Sin. 2012, 33, 312–323. [Google Scholar] [CrossRef]
- Livnah, O.; Stura, E.A.; Middleton, S.A.; Johnson, D.L.; Jolliffe, L.K.; Wilson, I.A. Crystallographic Evidence for Preformed Dimers of Erythropoietin Receptor Before Ligand Activation. Science 1999, 283, 987–990. [Google Scholar] [CrossRef]
- Pang, X.; Zhou, H.-X. Activation of signaling receptors: Do ligands bind to receptor monomer, dimer, or both? BMC Biophys. 2013, 6, 7. [Google Scholar] [CrossRef]
- Peng, J.; Xu, J. Raptorx: Exploiting structure information for protein alignment by statistical inference. Proteins Struct. Funct. Bioinform. 2011, 79, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Källberg, M.; Wang, H.; Wang, S.; Peng, J.; Wang, Z.; Lu, H.; Xu, J. Template-based protein structure modeling using the RaptorX web server. Nat. Protoc. 2012, 7, 1511–1522. [Google Scholar] [CrossRef]
- Liang, J.; Qin, M.; Xu, R.; Gao, X.; Shen, Y.; Xu, Q.; Cao, Y.; Wang, W. A genetically encoded copper(i) sensor based on engineered structural distortion of EGFP. Chem. Commun. 2012, 48, 3890. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Guo, L.; Ding, Y.; Xia, L.; Shen, Y.; Qin, M.; Xu, Q.; Cao, Y.; Wang, W. Genetically encoded red fluorescent copper(I) sensors for cellular copper(I) imaging. Biochem. Biophys. Res. Commun. 2014, 443, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Tsien, R.Y. The green fluorescent protein. Annu. Rev. Biochem. 1998, 67, 509–544. [Google Scholar] [CrossRef]
- Hack, N.J.; Billups, B.; Guthrie, P.B.; Rogers, J.H.; Muir, E.M.; Parks, T.N.; Kater, S.B. Green fluorescent protein as a quantitative tool. J. Neurosci. Methods 2000, 95, 177–184. [Google Scholar] [CrossRef]
- Hu, C.-D.; Kerppola, T.K. Simultaneous visualization of multiple protein interactions in living cells using multicolor fluorescence complementation analysis. Nat. Biotechnol. 2003, 21, 539–545. [Google Scholar] [CrossRef]
- Piston, D.W.; Kremers, G.-J. Fluorescent protein FRET: The good, the bad and the ugly. Trends Biochem. Sci. 2007, 32, 407–414. [Google Scholar] [CrossRef]
- Mueller, J.D. Fluorescence Fluctuation Spectroscopy. In Encyclopedia of Biophysics; Roberts, G.C.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 800–803. Available online: http://link.springer.com/10.1007/978-3-642-16712-6_504 (accessed on 4 March 2024).
- Jameson, D.M.; Ross, J.A.; Albanesi, J.P. Fluorescence fluctuation spectroscopy: Ushering in a new age of enlightenment for cellular dynamics. Biophys. Rev. 2009, 1, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Elson, E.L. Fluorescence Correlation Spectroscopy: Past, Present, Future. Biophys. J. 2011, 101, 2855–2870. [Google Scholar] [CrossRef] [PubMed]
- Ehrenberg, M.; Rigler, R. Rotational brownian motion and fluorescence intensify fluctuations. Chem. Phys. 1974, 4, 390–401. [Google Scholar] [CrossRef]
- Elson, E.L.; Magde, D. Fluorescence correlation spectroscopy. I. Conceptual basis and theory. Biopolymers 1974, 13, 1–27. [Google Scholar] [CrossRef]
- Magde, D.; Elson, E.L.; Webb, W.W. Fluorescence correlation spectroscopy. II. An experimental realization. Biopolymers 1974, 13, 29–61. [Google Scholar] [CrossRef] [PubMed]
- Kaliszewski, M.J.; Shi, X.; Hou, Y.; Lingerak, R.; Kim, S.; Mallory, P.; Smith, A.W. Quantifying membrane protein oligomerization with fluorescence cross-correlation spectroscopy. Methods 2018, 140–141, 40–51. [Google Scholar] [CrossRef]
- Liang, W.; Guo, B.; Ye, J.; Liu, H.; Deng, W.; Lin, C.; Zhong, X.; Wang, L. Vasorin stimulates malignant progression and angiogenesis in glioma. Cancer Sci. 2019, 110, 2558–2572. [Google Scholar] [CrossRef]
- Singh, D.R.; Pandey, K.; Mishra, A.K.; Pandey, P.; Vivcharuk, V. Glutamate binding triggers monomerization of unliganded mGluR2 dimers. Arch. Biochem. Biophys. 2021, 697, 108632. [Google Scholar] [CrossRef]
- Asher, W.B.; Geggier, P.; Holsey, M.D.; Gilmore, G.T.; Pati, A.K.; Meszaros, J.; Terry, D.S.; Mathiasen, S.; Kaliszewski, M.J.; McCauley, M.D.; et al. Single-molecule FRET imaging of GPCR dimers in living cells. Nat. Methods 2021, 18, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.J. Minireview: Receptor Dimerization in GH and Erythropoietin Action—It Takes Two to Tango, But How? Endocrinology 2002, 143, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Maruyama, I.N. Brain-derived neurotrophic factor receptor TrkB exists as a preformed dimer in living cells. J. Mol. Signal. 2012, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Comar, W.D.; Schubert, S.M.; Jastrzebska, B.; Palczewski, K.; Smith, A.W. Time-Resolved Fluorescence Spectroscopy Measures Clustering and Mobility of a G Protein-Coupled Receptor Opsin in Live Cell Membranes. J. Am. Chem. Soc. 2014, 136, 8342–8349. [Google Scholar] [CrossRef]
- Liang, J.; Seghiri, M.; Singh, P.K.; Seo, H.G.; Lee, J.Y.; Jo, Y.; Song, Y.B.; Park, C.; Zalicki, P.; Jeong, J.-Y.; et al. The β2-adrenergic receptor associates with CXCR4 multimers in human cancer cells. Proc. Natl. Acad. Sci. USA 2024, 121, e2304897121. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, J.; Smith, A.W. The Oligomeric State of Vasorin in the Plasma Membrane Measured Non-Invasively by Quantitative Fluorescence Fluctuation Spectroscopy. Int. J. Mol. Sci. 2024, 25, 4115. https://doi.org/10.3390/ijms25074115
Liang J, Smith AW. The Oligomeric State of Vasorin in the Plasma Membrane Measured Non-Invasively by Quantitative Fluorescence Fluctuation Spectroscopy. International Journal of Molecular Sciences. 2024; 25(7):4115. https://doi.org/10.3390/ijms25074115
Chicago/Turabian StyleLiang, Junyi, and Adam W. Smith. 2024. "The Oligomeric State of Vasorin in the Plasma Membrane Measured Non-Invasively by Quantitative Fluorescence Fluctuation Spectroscopy" International Journal of Molecular Sciences 25, no. 7: 4115. https://doi.org/10.3390/ijms25074115
APA StyleLiang, J., & Smith, A. W. (2024). The Oligomeric State of Vasorin in the Plasma Membrane Measured Non-Invasively by Quantitative Fluorescence Fluctuation Spectroscopy. International Journal of Molecular Sciences, 25(7), 4115. https://doi.org/10.3390/ijms25074115