
Spin Trapping of Nitric Oxide by Hemoglobin and Ferrous Diethyldithiocarbamate in Model Tumors Differing in Vascularization

 , ,
, , 
Abstract
:1. Introduction
2. Results
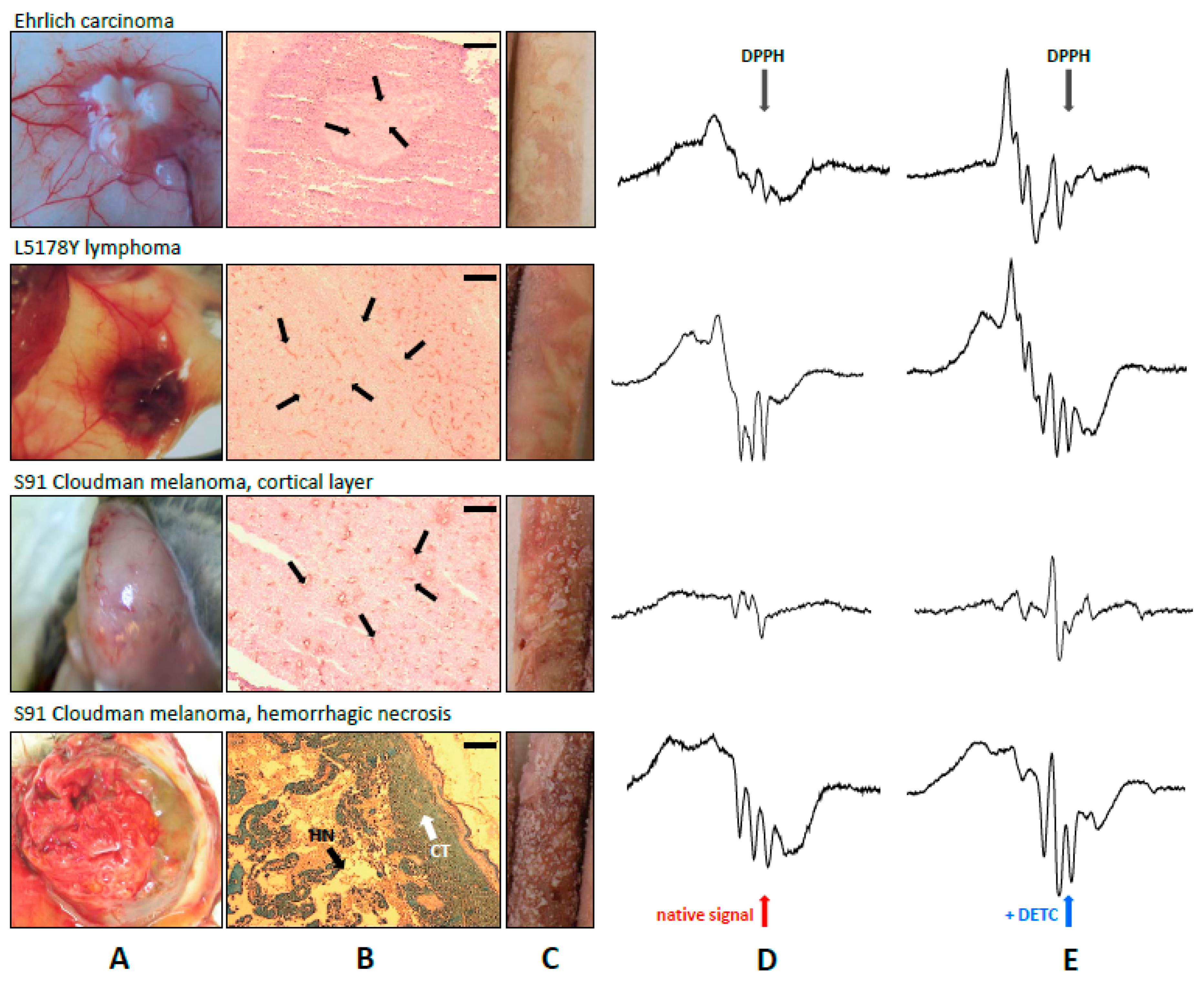
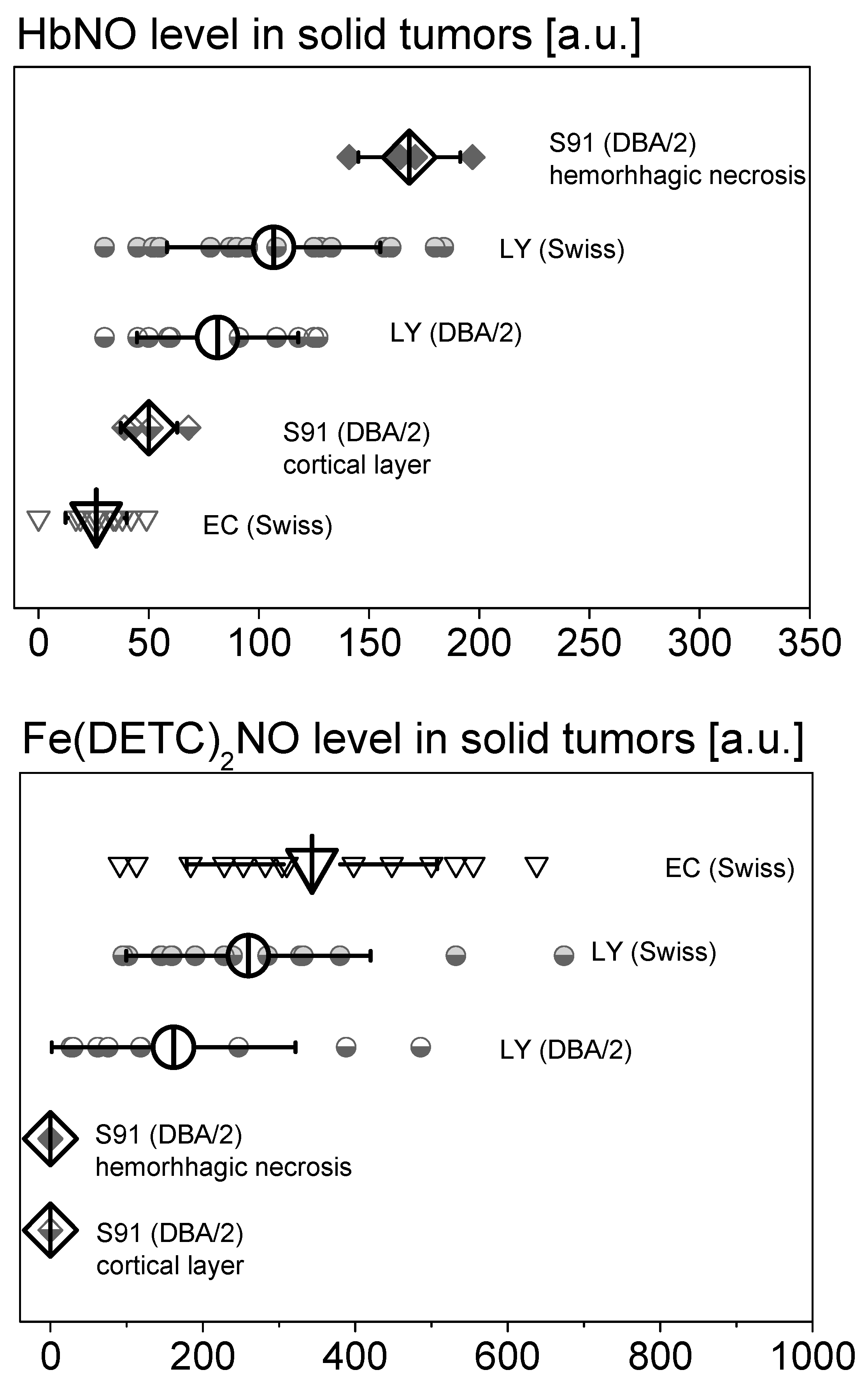
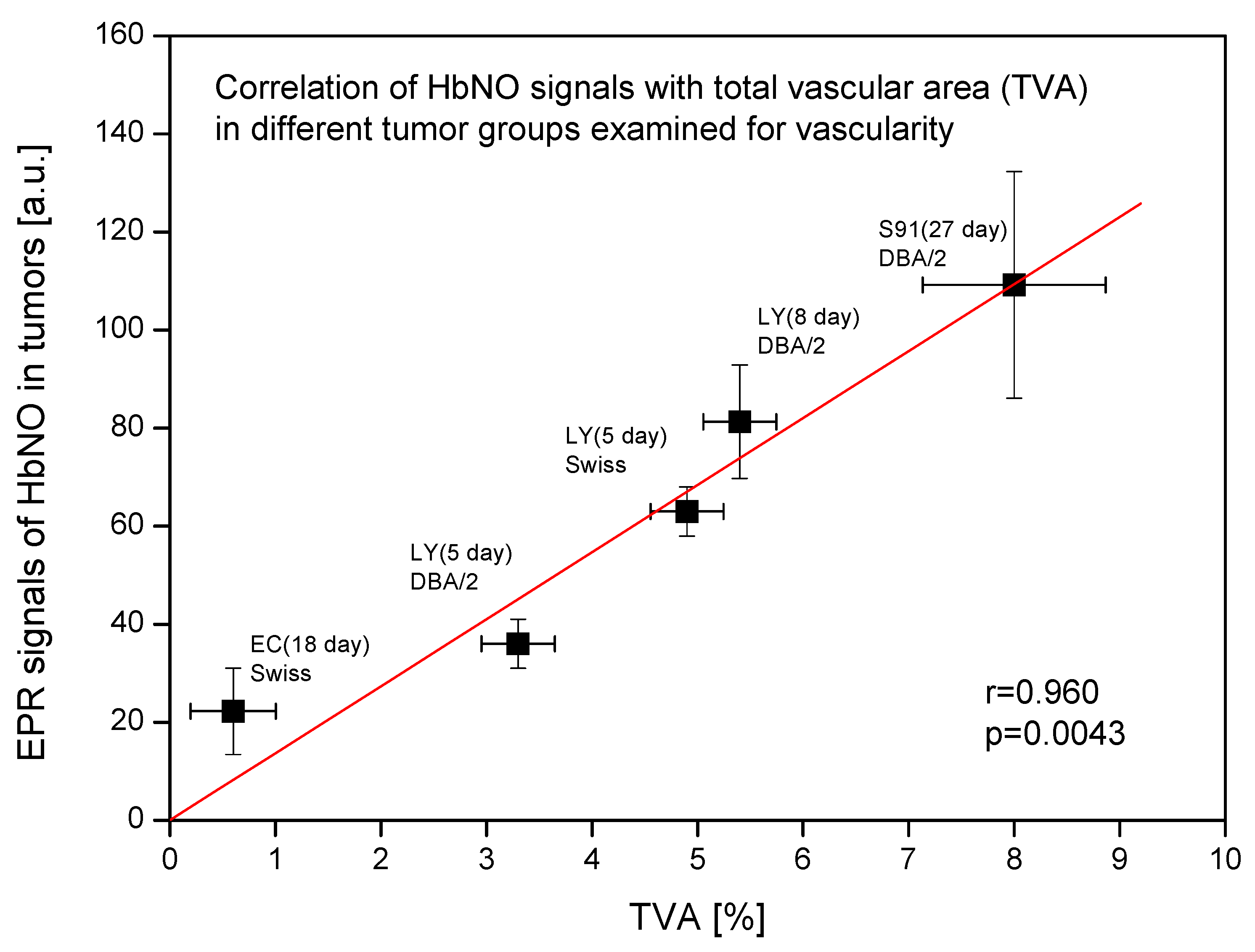
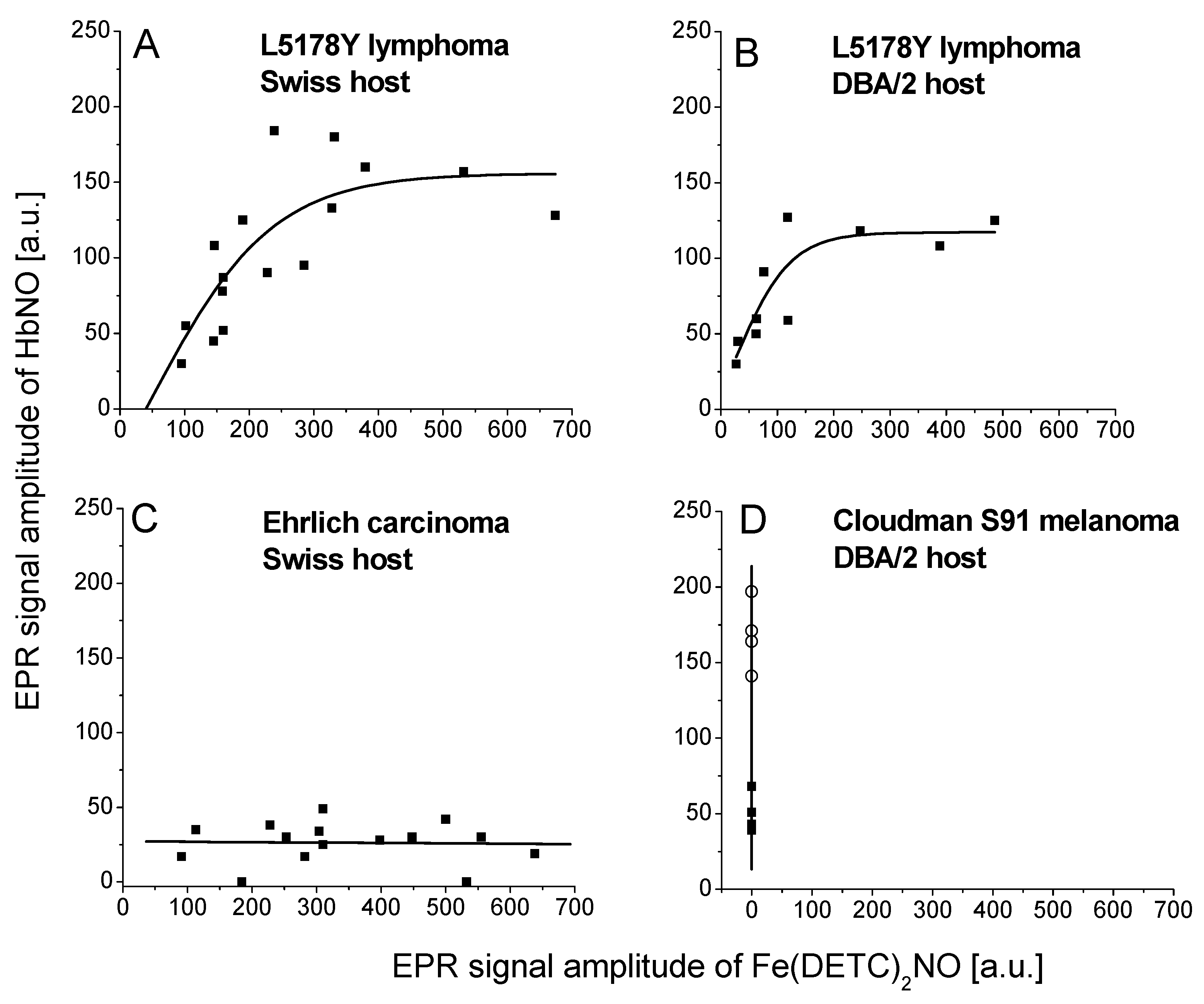
2.1. Fe–NO Complexes in Tumors Differing in Vascularization
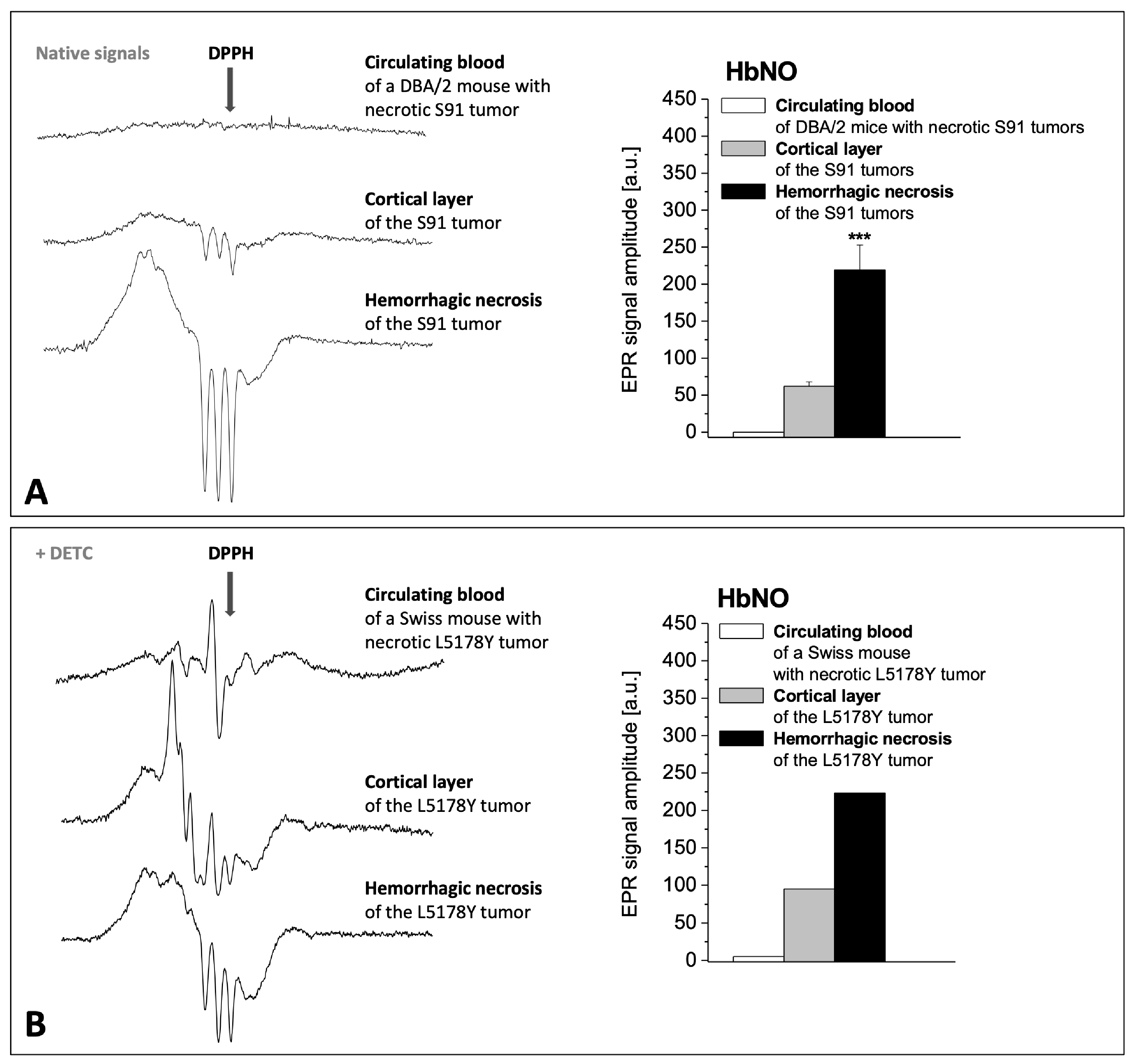
2.2. Fe–NO Complexes in the Areas of Hemorrhagic Necrosis in the Tumors
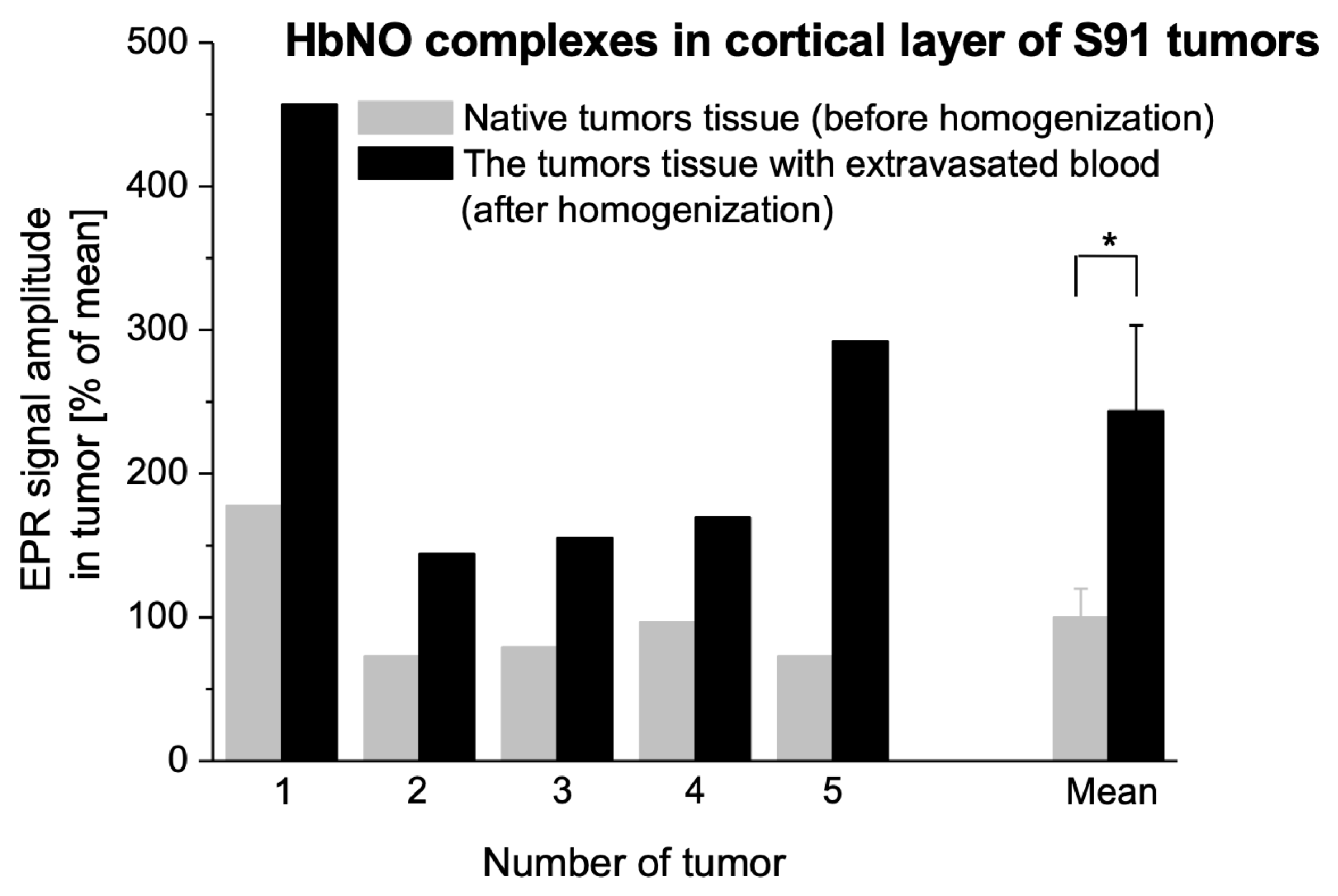
2.3. HbNO Complexes in Tumors with Blood Extravasation
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals and Tumor Cell Lines
4.3. EPR Sample Preparation
4.4. Analysis of Tumor Necrosis
4.5. Tumor Homogenization
4.6. Blood EPR Analysis
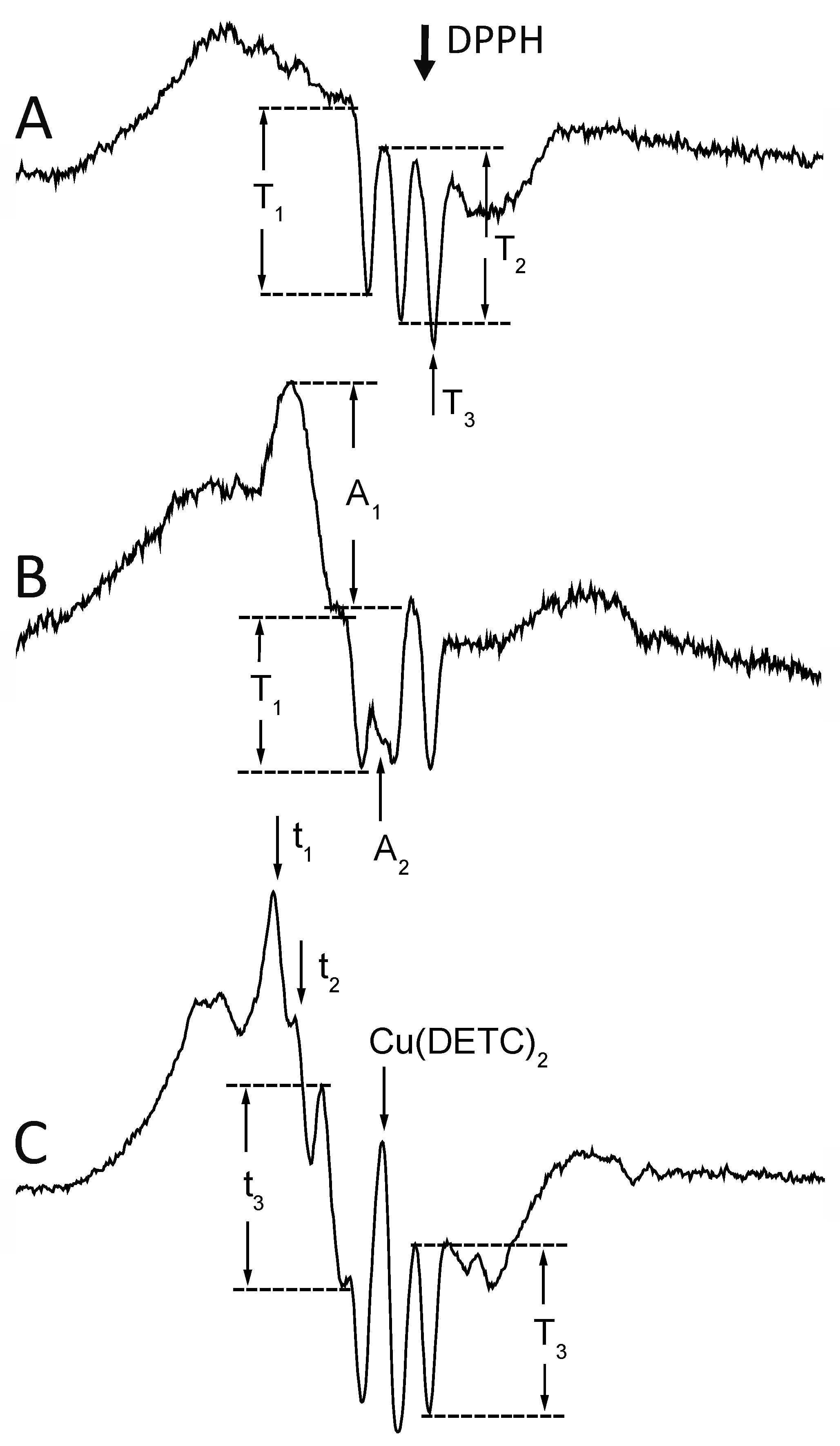
4.7. EPR Measurement
4.8. Tumor Microvascularization Analysis
4.9. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ingram, D.J.E.; Bennett, J.E. Paramagnetic resonance in phthalocyanine, haemoglobin, and other organic derivatives. Discuss. Faraday Soc. 1955, 19, 140–146. [Google Scholar] [CrossRef]
- Gordy, W.; Rexroad, H.N. Electron paramagnetic resonance studies of cytochrome and hemoglobin (irradiated and unirradiated). In Free Radicals in Biological Systems, Proceedings of the Symposium on Free Radicals in Biological Systems, Stanford University, USA, March 1960; Blois, M.S., Jr., Brown, H.W., Lemmon, R.M., Lindblom, R.O., Weissbluth, M., Eds.; Academic Press: New York, NY, USA; London, UK, 1961; pp. 263–278. [Google Scholar]
- Emanuel, N.M.; Saprin, A.N.; Shabalkin, V.A.; Kozlova, L.E.; Krugljakova, K.E. Detection and investigation of a new type of ESR signal characteristic of some tumour tissues. Nature 1969, 222, 165–167. [Google Scholar] [CrossRef]
- Brennan, M.J.; Cole, T.; Singley, J.A. A unique hyperfine ESR spectrum in mouse neoplasms analyzed by computer simulation. Proc. Soc. Exp. Biol. Med. 1966, 123, 715–718. [Google Scholar] [CrossRef]
- Symons, M.C.R.; Rowland, I.J.; Deighton, N.; Shorrock, K.; West, K.P. Electron spin resonance studies of nitrosyl haemoglobin in human liver, colon and stomach tumour tissues. Free Radic. Res. 1994, 21, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Loibl, S.; Von Minckwitz, G.; Weber, S.; Sinn, H.P.; Schini-Kerth, V.B.; Lobysheva, I.; Nepveu, F.; Wolf, G.; Strebhardt, K.; Kaufmann, M. Expression of endothelial and inducible nitric oxide synthase in benign and malignant lesions of the breast and measurement of nitric oxide using electron paramagnetic resonance spectroscopy. Cancer 2002, 95, 1191–1198. [Google Scholar] [CrossRef]
- Elas, M.; Raczek, J.; Bratasz, A.; Rudnicka, B.; Krawczyk, K.; Szypulka, R.; Hodor, K.; Lukiewicz, S.J. Paramagnetic center observed in human tumors by EPR technique. Curr. Top. Biophys. 1999, 23, 39–45. [Google Scholar]
- Maruyama, T.; Kataoka, N.; Nagase, S.; Nakada, H.; Sato, H.; Sasaki, H. Identification of Three-Line Electron Spin Resonance Signal and Its Relationship to Ascites Tumors. Cancer Res. 1971, 31, 179–184. [Google Scholar] [PubMed]
- Saprin, A.N.; Shuliakovskaia, T.S. New type of ESR signal arising in animal tissues treated with a series of chemical agents and during necroses. Dokl. Akad. Nauk. SSSR 1969, 189, 889–891. [Google Scholar] [PubMed]
- Rowlands, J.R.; Cadena, D.G., Jr.; Gross, A.L. Effects of tobacco smoke on lung tissue as measured by electron spin resonance. Nature 1967, 213, 1256–1258. [Google Scholar] [CrossRef]
- Azhipa, Y.I.; Kayushin, L.P.; Nikishkin, E.I. Electron paramagnetic resonance of tissues of animals on exposure to certain forms of tissue hypoxia. Biofizika 1966, 11, 710–713. [Google Scholar]
- Louro, S.R.W.; Ribeiro, P.C.; Bemski, G. EPR spectral changes of nitrosyl hemes and their relation to the hemoglobin T-R transition. Biochim. Biophys. Acta (BBA) Protein Struct. 1981, 670, 56–63. [Google Scholar] [CrossRef]
- Hille, R.; Olson, J.S.; Palmer, G. Spectral transitions of nitrosyl hemes during ligand binding to hemoglobin. J. Biol. Chem. 1979, 254, 12110–12120. [Google Scholar] [CrossRef] [PubMed]
- Kon, H.; Kataoka, N. Electron paramagnetic resonance of nitric oxide-protoheme complexes with some nitrogenous base. Model systems of nitric oxide hemoproteins. Biochemistry 1969, 8, 4757–4762. [Google Scholar] [CrossRef] [PubMed]
- Westenberger, U.; Thanner, S.; Ruf, H.H.; Gersonde, K.; Sutter, G.; Trentz, O. Formation of free radicals and nitric oxide derivative of hemoglobin in rats during shock syndrome. Free Radic. Res. Commun. 1990, 11, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Plonka, P.M.; Chlopicki, S.; Plonka, B.K.; Jawien, J.; Gryglewski, R.J. Endotoxaemia in rats: Detection of nitrosyl-haemoglobin in blood and lung by EPR. Curr. Top. Biophys. 1999, 23, 47–53. [Google Scholar]
- Plonka, P.M.; Plonka, B.K.; Cieszka, K.; Raczek, J.; Lukiewicz, S.J. EPR analysis of tumor-host interactions. In Nitric Oxide in Transplant Rejection and Anti-Tumor Defense; Lukiewicz, S., Zweier, J.L., Eds.; Kluwer Academic Publishers: Boston, MA, USA; Dordrecht, The Netherlands; London, UK, 1998; pp. 313–326. [Google Scholar]
- Yim, C.Y.; Bastian, N.R.; Smith, J.C.; Hibbs, J.B.; Samlowski, W.E. Macrophage nitric oxide synthesis delays progression of ultraviolet light-induced murine skin cancers. Cancer Res. 1993, 53, 5507–5511. [Google Scholar] [PubMed]
- Lukiewicz, S.J.; Plonka, P.M.; Plonka, B.K.; Raczek, J.; Pajak, S.; Cieszka, K. Animal EPR studies on allo- and xenograft rejection. In Nitric Oxide in Transplant Rejection and Anti-Tumor Defense; Lukiewicz, S., Zweier, J.L., Eds.; Kluwer Academic Publishers: Boston, MA, USA; Dordrecht, The Netherlands; London, UK, 1998; pp. 157–187. [Google Scholar]
- Lancaster, J.R., Jr.; Langrehr, J.M.; Bergonia, H.A.; Murase, N.; Simmons, R.L.; Hoffman, R.A. EPR detection of heme and nonheme iron-containing protein nitrosylation by nitric oxide during rejection of rat heart allograft. J. Biol. Chem. 1992, 267, 10994–10998. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, J.R., Jr.; Hibbs, J.B., Jr. EPR demonstration of iron-nitrosyl complex formation by cytotoxic activated macrophages. Proc. Natl. Acad. Sci. USA 1990, 87, 1223–1227. [Google Scholar] [CrossRef] [PubMed]
- Haldane, J.; Makgill, R.H.; Mavrogordato, A.E. The action as poisons of nitrites and other physiologically related substances. J. Physiol. 1897, 21, 160–189. [Google Scholar] [CrossRef]
- Gamgee, A. Researches on the blood. On the action of nitrites on blood. Philos. Trans. R. Soc. Lond. 1868, 158, 589–625. [Google Scholar]
- Jenkins, D.C.; Charles, I.G.; Thomsen, L.L.; Moss, D.W.; Holmes, L.S.; Baylis, S.A.; Rhodes, P.; Westmore, K.; Emson, P.C.; Moncada, S. Roles of nitric oxide in tumor growth. Proc. Natl. Acad. Sci. USA 1995, 92, 4392–4396. [Google Scholar] [CrossRef] [PubMed]
- Dutka, M.; Pyka, J.; Plonka, P.M. EPR studies on understanding the physical intricacy of HbNO complexes. In Electron Spin Resonance Spectroscopy in Medicine; Shukla, A.K., Ed.; Springer Nature Singapore Pte Ltd.: Singapore, 2019; pp. 23–43. [Google Scholar]
- Vanin, A.F.; Serezhenkov, V.A.; Mikoyan, V.D.; Genkin, M.V. The 2.03 signal as an indicator of dinitrosyl-iron complexes with thiol-containing ligands. Nitric Oxide 1998, 2, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Vanin, A.F.; Nalbandyan, R.M. Free Radicals of a New Type in Yeast Cells. Biofizika 1965, 10, 167–168. [Google Scholar] [PubMed]
- Bastian, N.R.; Yim, C.Y.; Hibbs, J.B., Jr.; Samlowski, W.E. Induction of iron-derived EPR signals in murine cancers by nitric oxide. Evidence for multiple intracellular targets. J. Biol. Chem. 1994, 269, 5127–5131. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.J.; Petersson, A.S.; Wennmalm, Å.; Hansson, G.K. Cytokine-induced expression of nitric oxide synthase results in nitrosylation of heme and nonheme iron proteins in vascular smooth muscle cells. Exp. Cell Res. 1994, 214, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Plonka, B.K.; Gurbiel, R.J.; Plonka, P.M. Complexity of EPR spectra of L5178Y-R lymphoma tumors growing in various murine hosts. Curr. Top. Biophys. 1999, 23, 55–62. [Google Scholar]
- Vanin, A.F.; Mordvintsev, P.I.; Kleshchov, A.L. Nitrogen oxide appearance in animal tissues in vivo. Stud. Biophys. 1984, 192, 135–143. [Google Scholar]
- Pustelny, K.; Bielanska, J.; Plonka, P.M.; Rosen, G.M.; Elas, M. In vivo spin trapping of nitric oxide from animal tumors. Nitric Oxide 2007, 16, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Goodman, B.A.; Raynor, J.B.; Symons, M.C.R. Electron spin resonance of bis(NN-diethylthiocarbamato)nitrosyliron. J. Chem. Soc. A Inorg. Phys. Theor. 1969, 2572–2575. [Google Scholar] [CrossRef]
- Vanin, A.F.; von Faassen, E. Mononitrosyl-iron complexes with dithiocarbamate ligands: Physico-chemical properties. In Radicals for Life: The Various Forms of Nitric Oxide; von Faassen, E., Vanin, A.F., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 383–405. [Google Scholar]
- Plonka, B.K.; Cieszka, K.; Chlopicki, S.; Plonka, P.M.; Pajak, S.; Raczek, J.; Lukiewicz, S.J. ESR signals detectable in tumors of murine L5178Y-R lymphoma. Curr. Top. Biophys. 1996, 20, 81–85. [Google Scholar]
- Plonka, P.M.; Wisniewska, M.; Chlopicki, S.; Elas, M.; Rosen, G.M. X-band and S-band EPR detection of nitric oxide in murine endotoxaemia using spin trapping by ferro-di(N-(dithiocarboxy)sarcosine). Acta Biochim. Pol. 2003, 50, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Pou, S.; Tsai, P.; Porasuphatana, S.; Halpern, H.J.; Chandramouli, G.V.R.; Barth, E.D.; Rosen, G.M. Spin trapping of nitric oxide by ferro-chelates: Kinetic and in vivo pharmacokinetic studies. Biochim. Biophys. Acta (BBA) Gen. Subj. 1999, 1427, 216–226. [Google Scholar] [CrossRef]
- Folkman, J. Tumor Angiogenesis: Therapeutic implications. N. Engl. J. Med. 1971, 285, 1182–1186. [Google Scholar] [PubMed]
- Quesada, A.R.; Muñoz-Chápuli, R.; Medina, M.A. Anti-Angiogenic Drugs: From Bench to Clinical Trials. Med. Res. Rev. 2006, 26, 483–530. [Google Scholar] [CrossRef] [PubMed]
- Dahl, O.; Borkamo, E.D.; Fluge, O. Current status of antivascular therapy and targeted treatment in the clinic. Int. J. Hyperth. 2008, 24, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Siemann, D.W.; Chaplin, D.J.; Horsman, M.R. Vascular-targeting therapies for treatment of malignant disease. Cancer 2004, 100, 2491–2499. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Sharma, M.C.; Sarkar, C. Morphology of Angiogenesis in Human Cancer: A Conceptual Overview, Histoprognostic Perspective and Significance of Neoangiogenesis. Histopathology 2005, 46, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Pezzella, F. Overview on the different patterns of tumor vascularization. Cells 2021, 10, 639. [Google Scholar] [CrossRef] [PubMed]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor angiogenesis: Causes, consequences, challenges and opportunities. Cell. Mol. Life Sci. 2020, 77, 1745–1770. [Google Scholar] [CrossRef]
- Burke, A.J.; Sullivan, F.J.; Giles, F.J.; Glynn, S.A. The yin and yang of nitric oxide in cancer progression. Carcinogenesis 2013, 34, 503–512. [Google Scholar] [CrossRef]
- Korde Choudhari, S.; Chaudhary, M.; Bagde, S.; Gadbail, A.R.; Joshi, V. Nitric oxide and cancer: A review. World J. Surg. Oncol. 2013, 11, 118. [Google Scholar] [CrossRef] [PubMed]
- Mintz, J.; Vedenko, A.; Rosete, O.; Shah, K.; Goldstein, G.; Hare, J.M.; Ramasamy, R.; Arora, H. Current advances of nitric oxide in cancer and anticancer therapeutics. Vaccines 2021, 9, 94. [Google Scholar] [CrossRef]
- Williamson, K.E.; Din, O.S.E.; O’Kane, H.F. Apoptosis. In The Cancer Handbook; Alison, M.R., Ed.; John Wiley & Sons Ltd.: Chichester, UK, 2007; Volume 1, pp. 47–59. [Google Scholar]
- Thomlinson, R.H.; Gray, L.H. The histological structure of some human lung cancers and the possible implications for radiotherapy. Br. J. Cancer 1955, 9, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, K.; Kanematsu, Y.; Yoshizumi, M.; Ohnishi, H.; Kirima, K.; Izawa, Y.; Shikishima, M.; Ishida, T.; Kondo, S.; Kagami, S.; et al. Nitrite is an alternative source of NO in vivo. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H2163–H2170. [Google Scholar] [CrossRef] [PubMed]
- Zweier, J.L.; Wang, P.; Samouilov, A.; Kuppusamy, P. Enzyme-independent formation of nitric oxide in biological tissues. Nat. Med. 1995, 1, 804–809. [Google Scholar] [CrossRef] [PubMed]
- Wennmalm, Å.; Lanne, B.; Petersson, A.S. Detection of endothelial-derived relaxing factor in human plasma in the basal state and following ischemia using electron paramagnetic resonance spectrometry. Anal. Biochem. 1990, 187, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.M. Tumor hypoxia in cancer therapy. Meth. Enzymol. 2007, 435, 297–321. [Google Scholar]
- Lotfi, R.; Lee, J.J.; Lotze, M.T. Eosinophilic granulocytes and damage-associated molecular pattern molecules (DAMPs): Role in the inflammatory response within tumors. J. Immunother. 2007, 30, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, L.L.; Ching, L.M.; Joseph, W.R.; Baguley, B.C.; Gavin, J.B. Nitric oxide production in endotoxin-resistant C3H/HeJ mice stimulated with flavone-8-acetic acid and xanthenone-4-acetic acid analogues. Biochem. Pharmacol. 1992, 43, 2401–2406. [Google Scholar] [CrossRef]
- Watanabe, Y.; Nagayama, M.; Okumura, A.; Amoh, Y.; Suga, T.; Terai, A.; Dodo, Y. MR imaging of testicular torsion: Features of testicular hemorrhagic necrosis and clinical outcomes. J. Magn. Reson. Imaging 2007, 26, 100–108. [Google Scholar] [CrossRef]
- Chacko, A.G.; Chacko, G.; Seshadri, M.S.; Chandy, M.J. Hemorrhagic necrosis of pituitary adenomas. Neurol. India 2002, 50, 490–493. [Google Scholar] [PubMed]
- Stetson, C.A. Studies on the mechanism of the Shwartzman phenomenon; certain factors involved in the production of the local hemorrhagic necrosis. J. Exp. Med. 1951, 93, 489–504. [Google Scholar] [CrossRef] [PubMed]
- Shwartzman, G. A new phenomenon of local skin reactivity to B. typhosus culture filtrate. Proc. Soc. Exp. Biol. Med. 1928, 25, 560–561. [Google Scholar] [CrossRef]
- Szczygiel, D.; Plonka, P.M.; Elas, M.; Szczygiel, M.; Plonka, B.K.; Lukiewicz, S.J. Appearance of iron-nitrosyl complexes in murine L5178Y lymphoma as a function of the form of growth and the type of host. Curr. Top. Biophys. 2005, 29, 65–71. [Google Scholar]
- Drapier, J.C.; Pellat, C.; Henry, Y. Generation of EPR-detectable nitrosyl-iron complexes in tumor target cells cocultured with activated macrophages. J. Biol. Chem. 1991, 266, 10162–10167. [Google Scholar] [CrossRef] [PubMed]
- Brovkovych, V.; Zhang, Y.; Brovkovych, S.; Minshall, R.D.; Skidgel, R.A. A novel pathway for receptor-mediated post-translational activation of inducible nitric oxide synthase. J. Cell. Mol. Med. 2011, 15, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Gross, S.S.; Jaffe, E.A.; Levi, R.; Kilbourn, R.G. Cytokine-activated endothelial cells express an isotype of nitric oxide synthase which is tetrahydrobiopterin-dependent, calmodulin-independent and inhibited by arginine analogs with a rank-order of potency characteristic of activated macrophages. Biochem. Biophys. Res. Commun. 1991, 178, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, L.L.; Miles, D.W.; Happerfield, L.; Bobrow, L.G.; Knowles, R.G.; Moncada, S. Nitric oxide synthase activity in human breast cancer. Br. J. Cancer 1995, 72, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.C.; Charles, I.G.; Baylis, S.A.; Lelchuk, R.; Radomski, M.W.; Moncada, S. Human colon cancer cell lines show a diverse pattern of nitric oxide synthase gene expression and nitric oxide generation. Br. J. Cancer 1994, 70, 847–849. [Google Scholar] [CrossRef]
- Thomsen, L.L.; Lawton, F.G.; Knowles, R.G.; Beesley, J.E.; Riveros-Moreno, V.; Moncada, S. Nitric oxide synthase activity in human gynecological cancer. Cancer Res. 1994, 54, 1352–1354. [Google Scholar]
- Lancaster, J.R., Jr. Simulation of the diffusion and reaction of endogenously produced nitric oxide. Proc. Natl. Acad. Sci. USA 1994, 91, 8137–8141. [Google Scholar] [CrossRef] [PubMed]
- Sinha, B.K.; Bhattacharjee, S.; Chatterjee, S.; Jiang, J.; Motten, A.G.; Kumar, A.; Espey, M.G.; Mason, R.P. Role of nitric oxide in the chemistry and anticancer activity of etoposide (VP-16,213). Chem. Res. Toxicol. 2013, 26, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Studenovsky, M.; Sivak, L.; Sedlacek, O.; Konefal, R.; Horkova, V.; Etrych, T.; Kovar, M.; Rihova, B.; Sirova, M. Polymer nitric oxide donors potentiate the treatment of experimental solid tumours by increasing drug accumulation in the tumour tissue. J. Control. Release 2018, 269, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Plonka, P.M.; Chlopicki, S.; Wisniewska, M.; Plonka, B.K. Kinetics of increased generation of NO in endotoxaemic rats as measured by EPR. Acta Biochim. Polon. 2003, 50, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Ziaja, M.; Pyka, J.; Machowska, A.; Maslanka, A.; Plonka, P.M. Nitric oxide spin-trapping and NADPH-diaphorase activity in mature rat brain after injury. J. Neurotrauma 2007, 24, 1845–1854. [Google Scholar] [CrossRef]
- Ziaja, M.; Pyka, J.; Ciombor, J.; Płonka, P.M. Kinetics of nitric oxide release in neonatal and mature rat brain during endotoxemia, as studied by diethyldithiocarbamate spin trapping. Curr. Top. Biophys. 2005, 29, 73–82. [Google Scholar]
- Szczygiel, D.; Pawlus, J.; Plonka, P.M.; Elas, M.; Szczygiel, M.; Plonka, B.K.; Lukiewicz, S.J. Nitric oxide complexes in the interaction between primary and secondary tumor of L5178Y lymphoma. Nitric Oxide 2004, 11, 279–289. [Google Scholar] [CrossRef]
- Plonka, P.M.; Plonka, B.K.; Pajak, S.; Lukiewicz, S.J. ESR studies on the generation of nitric oxide in rejecting rat heart allografts and in situ growing tumors. Curr. Top. Biophys. 1996, 20, 46–52. [Google Scholar]
- Lancaster Jr, J.R.; Xie, K. Tumors Face NO Problems? Cancer Res. 2006, 66, 6459–6462. [Google Scholar] [CrossRef]
- Morgan, R.A.; Dudley, M.E.; Wunderlich, J.R.; Hughes, M.S.; Yang, J.C.; Sherry, R.M.; Royal, R.E.; Topalían, S.L.; Kammula, U.S.; Restifo, N.P.; et al. Cancer regression in patients after transfer of genetically engineered lymphocytes. Science 2006, 314, 126–129. [Google Scholar] [CrossRef]
- Veszelovszky, E.; Thomsen, L.L.; Zhuang, L.; Baguley, B.C. Flavone acetic acid and 5,6-dimethylxanthenone-4-acetic acid: Relationship between plasma nitrate elevation and the induction of tumour necrosis. Eur. J. Cancer 1993, 29A, 404–408. [Google Scholar] [CrossRef] [PubMed]
- Winsborrow, B.A.G.; Grondey, H.; Savoie, H.; Fyfe, C.A.; Dolphin, D. Magnetic resonance imaging evaluation of photodynamic therapy-induced hemorrhagic necrosis in the murine M1 tumor model. Photochem. Photobiol. 1997, 66, 847–852. [Google Scholar] [CrossRef] [PubMed]
- Plonka, P.M. Electron paramagnetic resonance as a unique tool for skin and hair research. Exp. Dermatol. 2009, 18, 472–482. [Google Scholar] [CrossRef]
- Pajak, S.; Cieszka, K.; Plonka, P.M.; Lukiewicz, S.J.; Mihm, M.; Slominski, A. Transplantable melanomas in gerbils (Meriones unguiculatus). I. Origin, morphology and growth rate. Anticancer. Res. 1996, 16, 1203–1208. [Google Scholar]
- Plonka, P.M.; Plonka, B.K.; Lukiewicz, S.J. Evaluation of implant—Recipient interactions by ESR spectroscopy. Curr. Top. Biophys. 1994, 18, 46–53. [Google Scholar]
- Kepp, O.; Galluzzi, L.; Zitvogel, L.; Kroemer, G. Pyroptosis—A cell death modality of its kind? Eur. J. Immunol. 2010, 40, 627–630. [Google Scholar] [CrossRef] [PubMed]
- Bergsbaken, T.; Fink, S.L.; Cookson, B.T. Pyroptosis: Host cell death and inflammation. Nat. Rev. Microbiol. 2009, 7, 99–109. [Google Scholar] [CrossRef]
- Fink, S.L.; Cookson, B.T. Caspase-1-dependent pore formation during pyroptosis leads to osmotic lysis of infected host macrophages. Cell. Microbiol. 2006, 8, 1812–1825. [Google Scholar] [CrossRef] [PubMed]
- Szumiel, I.; Kapiszewska, M.; Kruszewski, M.; Iwanenko, T.; Lange, C.S. Content of iron and copper in the nuclei and induction of pH 9-labile lesions in L5178Y sublines inversely cross-sensitive to H2O2 and X-rays. Radiat. Environ. Biophys. 1995, 34, 113–119. [Google Scholar] [CrossRef]
- Kubrina, L.N.; Caldwell, W.S.; Mordvintcev, P.I.; Malenkova, I.V.; Vanin, A.F. EPR evidence for nitric oxide production from guanidino nitrogens of L-arginine in animal tissues in vivo. Biochim. Biophys. Acta (BBA) Bioenerg. 1992, 1099, 233–237. [Google Scholar] [CrossRef]
- Andriambeloson, E.; Kleschyov, A.L.; Muller, B.; Beretz, A.; Stoclet, J.C.; Andriantsitohaina, R. Nitric oxide production and endothelium-dependent vasorelaxation induced by wine polyphenols in rat aorta. Br. J. Pharmacol. 1997, 120, 1053–1058. [Google Scholar] [CrossRef]
- Alimoradi, H.; Greish, K.; Gamble, A.B.; Giles, G.I. Controlled delivery of nitric oxide for cancer therapy. Pharm. Nanotechnol. 2019, 7, 279–303. [Google Scholar] [CrossRef]
- Matuszak, Z.; Chignell, C.F.; Reszka, K.J. The acid-catalyzed interaction of melanin with nitrite ions. An EPR investigation. Nukleonika 2015, 60, 475–481. [Google Scholar] [CrossRef]
- Godechal, Q.; Gallez, B. The contribution of electron paramagnetic resonance to melanoma research. J. Ski. Skin. Cancer 2011, 2011, 273280. [Google Scholar] [CrossRef]
- Yarlagadda, K.; Hassani, J.; Foote, I.P.; Markowitz, J. The role of nitric oxide in melanoma. Biochim. Biophys. Acta (BBA) Rev. Cancer 2017, 1868, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Challis, G.B.; Stam, H.J. The spontaneous regression of cancer: A review of cases from 1900 to 1987. Acta Oncol. 1990, 29, 545–550. [Google Scholar] [CrossRef]
- Yang, Z.; Misner, B.; Ji, H.; Poulos, T.L.; Silverman, R.B.; Meyskens, F.L.; Yang, S. Targeting nitric oxide signaling with nNOS inhibitors as a novel strategy for the therapy and prevention of human melanoma. Antioxid. Redox Signal. 2013, 19, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Gupta, M.; Rundle, P.A.; Rennie, I.G. Spontaneous regression of choroidal melanoma. Eye 2006, 20, 959–961. [Google Scholar] [CrossRef]
- Kalialis, L.V.; Drzewiecki, K.T.; Mohammadi, M.; Mehlsen, A.B. Spontaneous regression of metastases from malignant melanoma: A case report. Melanoma Res. 2008, 18, 279–283. [Google Scholar] [CrossRef]
- Muniesa, C.; Ferreres, J.R.; Moreno, A.; Marcoval, J. Completely regressed primary cutaneous malignant melanoma with metastases. J. Eur. Acad. Dermatol. Venereol. 2009, 23, 327–328. [Google Scholar] [CrossRef]
- Pierret, L.; Van Baren, N.; Bonehill, A.; Corthals, J.; Van Nuffel, A.M.T.; Heirman, C.; Roelandt, T.; De Coninck, A.; Van Riet, I.; Degreef, E.; et al. Hemorrhagic regression of melanoma metastases during therapeutic vaccination: A report of three cases. Melanoma Res. 2009, 19, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Kalialis, L.V.; Drzewiecki, K.T.; Klyver, H. Spontaneous regression of metastases from melanoma: Review of the literature. Melanoma Res. 2009, 19, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Dillon, P.; Thomas, N.; Sharpless, N.; Collichio, F. Regression of advanced melanoma upon withdrawal of immunosuppression: Case series and literature review. Med. Oncol. 2010, 27, 1127–1132. [Google Scholar] [CrossRef] [PubMed]
- Loustalot, P.; Algire, G.H.; Legallais, F.Y.; Anderson, B.F. Growth and histopathology of melanotic and amelanotic derivatives of the Cloudman melanoma S91. J. Natl. Cancer Inst. 1952, 12, 1079–1117. [Google Scholar] [PubMed]
- Cloudman, A.M. The effect of an extra-chromosomal influence upon transplanted spontaneous tumors in mice. Science 1941, 93, 380–381. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, E. B16 murine melanoma: Historical perspective on the development of a solid tumor model. In Tumor Models in Cancer Research; Teicher, B.A., Ed.; Humana Press: Totowa, NJ, USA, 2002; pp. 73–89. [Google Scholar]
- Green, E.L. Handbook of Genetically Standardized JAX Mice, 1st ed.; Bar Harbor Times Publishing Company: Bar Harbor, ME, USA, 1962. [Google Scholar]
- Stetson, C.A.; Good, R.A. Studies on the mechanism of the Shwartzman-phenomenon; evidence for the participation of polymorphonuclear leucocytes in the phenomenon. J. Exp. Med. 1951, 93, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Shwartzman, G. Studies on Bacillus typhosus toxic substances: I. Phenomenon of local skin reactivity to B. typhosus culture filtrate. J. Exp. Med. 1928, 48, 247–268. [Google Scholar] [CrossRef] [PubMed]
- Shwartzman, G. Studies on streptococcus bacteriophage: II. The influence of lytic principles upon the agglutination of hemolytic streptococci. J. Exp. Med. 1928, 47, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Shwartzman, G. Hemorrhagic necrosis and regression of Sarcoma 180. Science 1935, 82, 201. [Google Scholar] [CrossRef]
- MacPherson, G.G.; North, R.J. Endotoxin-mediated necrosis and regression of established tumours in the mouse. A correlative study of quantitative changes in blood flow and ultrastructural morphology. Cancer Immunol. Immunother. 1986, 21, 209–216. [Google Scholar] [CrossRef]
- Freudenberg, N.; Joh, K.; Westphal, O.; Mittermayer, C.; Freudenberg, M.A.; Galanos, C. Haemorrhagic tumour necrosis following endotoxin administration. I. Communication: Morphological investigation on endotoxin-induced necrosis of the methylcholanthrene (Meth A) tumour in the mouse. Virchows Arch. A Pathol. Anat. Histopathol. 1984, 403, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Carswell, E.A.; Old, L.J.; Kassel, R.L.; Green, S.; Fiore, N.; Williamson, B. An endotoxin-induced serum factor that causes tumor necrosis. Proc. Natl. Acad. Sci. USA 1975, 72, 3666–3670. [Google Scholar] [CrossRef]
- Buehler, P.W.; D’Agnillo, F. Toxicological consequences of extracellular hemoglobin: Biochemical and physiological perspectives. Antioxid. Redox Signal. 2010, 12, 275–291. [Google Scholar] [CrossRef]
- Beer, J.Z.; Budzicka, E.; Niepokojczycka, E.; Rosiek, O.; Szumiel, I.; Walicka, M. Loss of tumorigenicity with simultaneous changes in radiosensitivity and photosensitivity during in vitro growth of L5178Y murine lymphoma cells. Cancer Res. 1983, 43, 4736–4742. [Google Scholar]
- Alexander, P. Mouse lymphoma cells with different radiosensitivities. Nature 1961, 192, 572–573. [Google Scholar] [CrossRef]
- Fischer, G.A.; Welch, A.D. Effect of citrovorum factor and peptones on mouse leukemia cells L-5178 in tissue culture. Science 1957, 126, 1018–1019. [Google Scholar] [CrossRef]
- Ehrlich, P.; Apolant, H. Beobachtungen über maligne Mäusetumoren. Berl. Klin. Wochenschr. 1905, 42, 871–874. [Google Scholar]
- Schrek, R. Further quantitative methods for the study of transplantable tumors. the growth of R39 sarcoma and Brown-Pearce carcinoma. Am. J. Cancer 1936, 28, 345–363. [Google Scholar]
- Vanin, A.F.; Kubrina, L.N.; Kurbanov, I.S.; Mordvintsev, P.I.; Khrapova, N.V.; Galagan, M.E.; Matkhanov, E.I. Iron as an inducer of nitric oxide formation in animal organisms. Biokhimiia 1989, 54, 1974–1979. [Google Scholar]
- Baguley, B.C.; Calveley, S.B.; Crowe, K.K.; Fray, L.M.; O’Rourke, S.A.; Smith, G.P. Comparison of the effects of flavone acetic acid, fostriecin, homoharringtonine and tumour necrosis factor α on Colon 38 tumours in mice. Eur. J. Cancer Clin. Oncol. 1989, 25, 263–269. [Google Scholar] [CrossRef]
- Romeis, B. Mikroskopische Technik; Urban & Schwarzenberg: Munich, Germany, 1991. [Google Scholar]
- Wertz, J.E.; Bolton, J.R. Electron Spin Resonance: Elementary Theory and Practical Applications, 1st ed.; Chapman and Hall: New York, NY, USA, 1986. [Google Scholar]
- Weidner, N.; Carroll, P.R.; Flax, J.; Blumenfeld, W.; Folkman, J. Tumor angiogenesis correlates with metastasis in invasive prostate carcinoma. Am. J. Pathol. 1993, 143, 401–409. [Google Scholar] [PubMed]
- Weidner, N.; Semple, J.P.; Welch, W.R.; Folkman, J. Tumor angiogenesis and metastasis—Correlation in invasive breast carcinoma. N. Engl. J. Med. 1991, 324, 1–8. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor Growth Parameters | Tumor Vascularization Parameters | ||||
|---|---|---|---|---|---|
| Solid Tumors | Host | Time [Days] | TVA | MVD | MAM |
| Ehrlich carcinoma | Swiss | 18 | 0.6 ± 0.7 | 23 ± 29 | 230 ± 121 |
| L5178Y lymphoma | DBA/2 | 5 | 3.3 ± 0.6 | 185 ± 25 | 176 ± 71 |
| Swiss | 5 | 4.9 ± 0.6 | 265 ± 11 | 190 ± 112 | |
| DBA/2 | 8 | 5.4 ± 0.6 | 354 ± 26 | 150 ± 97 | |
| S91 melanoma | DBA/2 | 27 | 8.0 ± 2.6 | 199 ± 73 | 490 ± 143 |
| Spleen (control) | DBA/2 | - | 2.8 ± 0.1 | 841 ± 120 | 40 ± 18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szczygieł, D.; Szczygieł, M.; Łaś, A.; Elas, M.; Zuziak, R.; Płonka, B.K.; Płonka, P.M. Spin Trapping of Nitric Oxide by Hemoglobin and Ferrous Diethyldithiocarbamate in Model Tumors Differing in Vascularization. Int. J. Mol. Sci. 2024, 25, 4172. https://doi.org/10.3390/ijms25084172
Szczygieł D, Szczygieł M, Łaś A, Elas M, Zuziak R, Płonka BK, Płonka PM. Spin Trapping of Nitric Oxide by Hemoglobin and Ferrous Diethyldithiocarbamate in Model Tumors Differing in Vascularization. International Journal of Molecular Sciences. 2024; 25(8):4172. https://doi.org/10.3390/ijms25084172
Chicago/Turabian StyleSzczygieł, Dariusz, Małgorzata Szczygieł, Anna Łaś, Martyna Elas, Roxana Zuziak, Beata K. Płonka, and Przemysław M. Płonka. 2024. "Spin Trapping of Nitric Oxide by Hemoglobin and Ferrous Diethyldithiocarbamate in Model Tumors Differing in Vascularization" International Journal of Molecular Sciences 25, no. 8: 4172. https://doi.org/10.3390/ijms25084172
APA StyleSzczygieł, D., Szczygieł, M., Łaś, A., Elas, M., Zuziak, R., Płonka, B. K., & Płonka, P. M. (2024). Spin Trapping of Nitric Oxide by Hemoglobin and Ferrous Diethyldithiocarbamate in Model Tumors Differing in Vascularization. International Journal of Molecular Sciences, 25(8), 4172. https://doi.org/10.3390/ijms25084172