
The Inhibition Activity of Natural Methoxyflavonoid from Inula britannica on Soluble Epoxide Hydrolase and NO Production in RAW264.7 Cells

 and
and
Abstract
:1. Introduction
2. Results
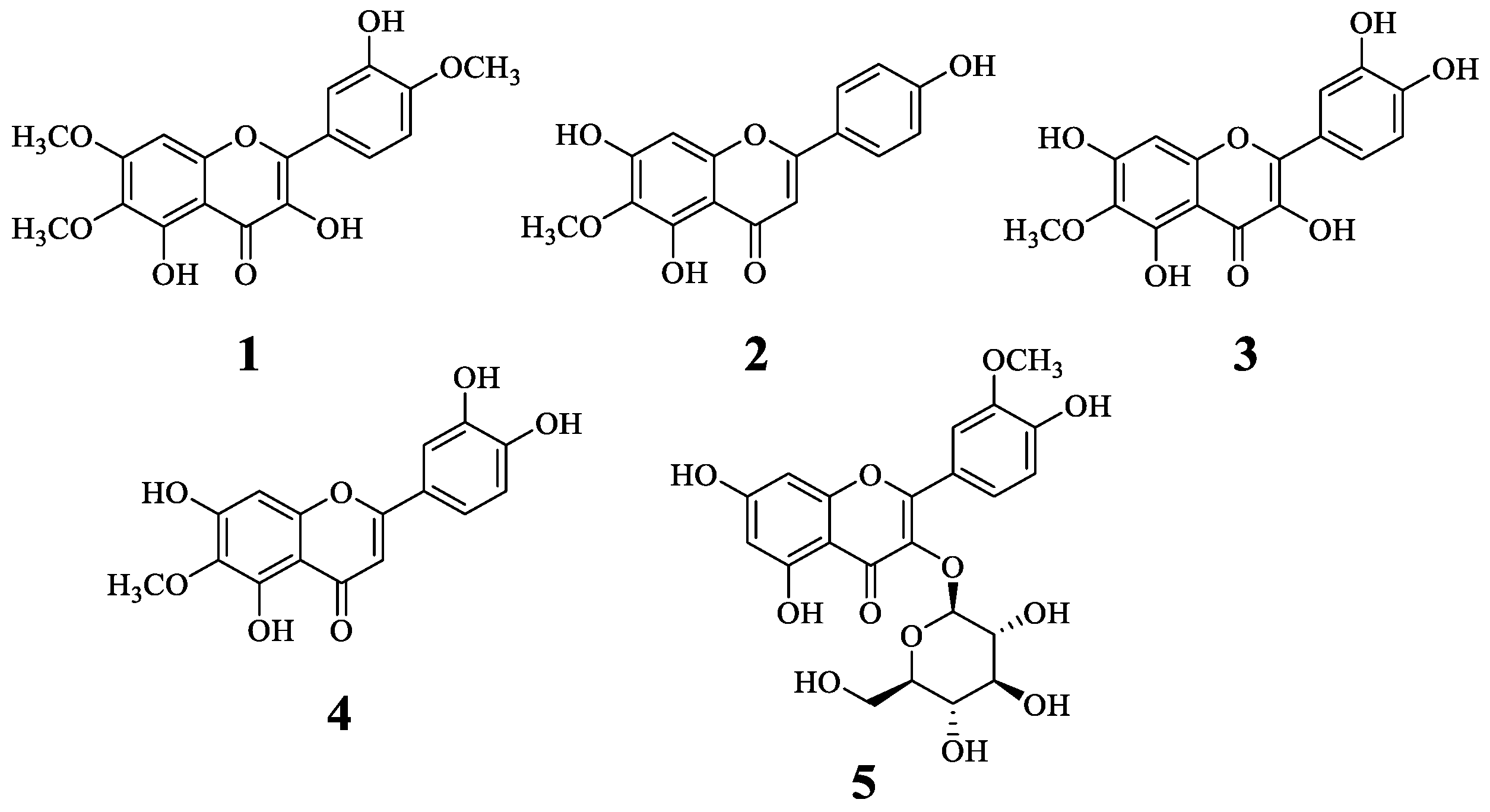
2.1. Isolation and Identification of Compounds 1–5
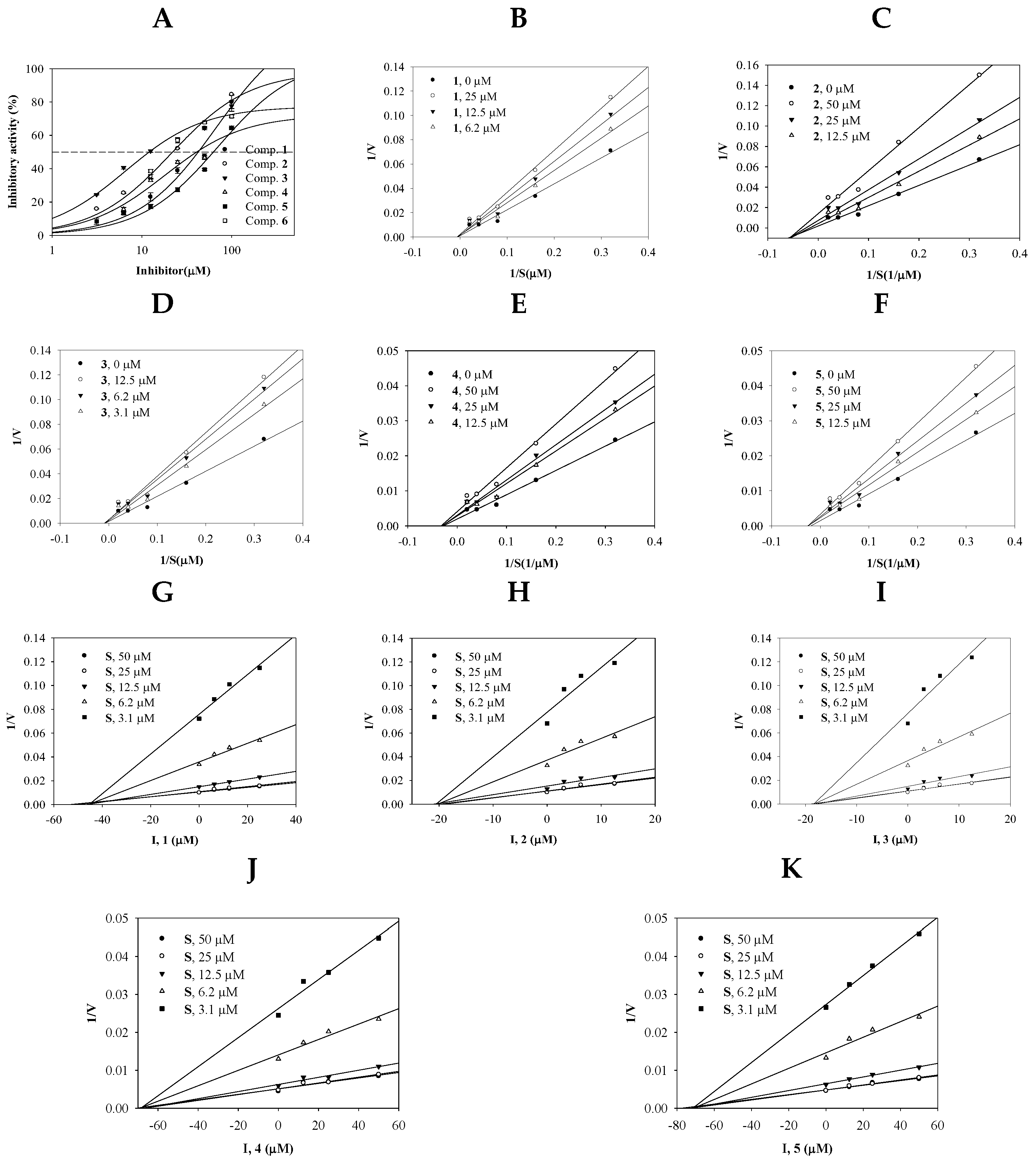
2.2. Inhibitory Activity of Compounds 1–5 on sEH
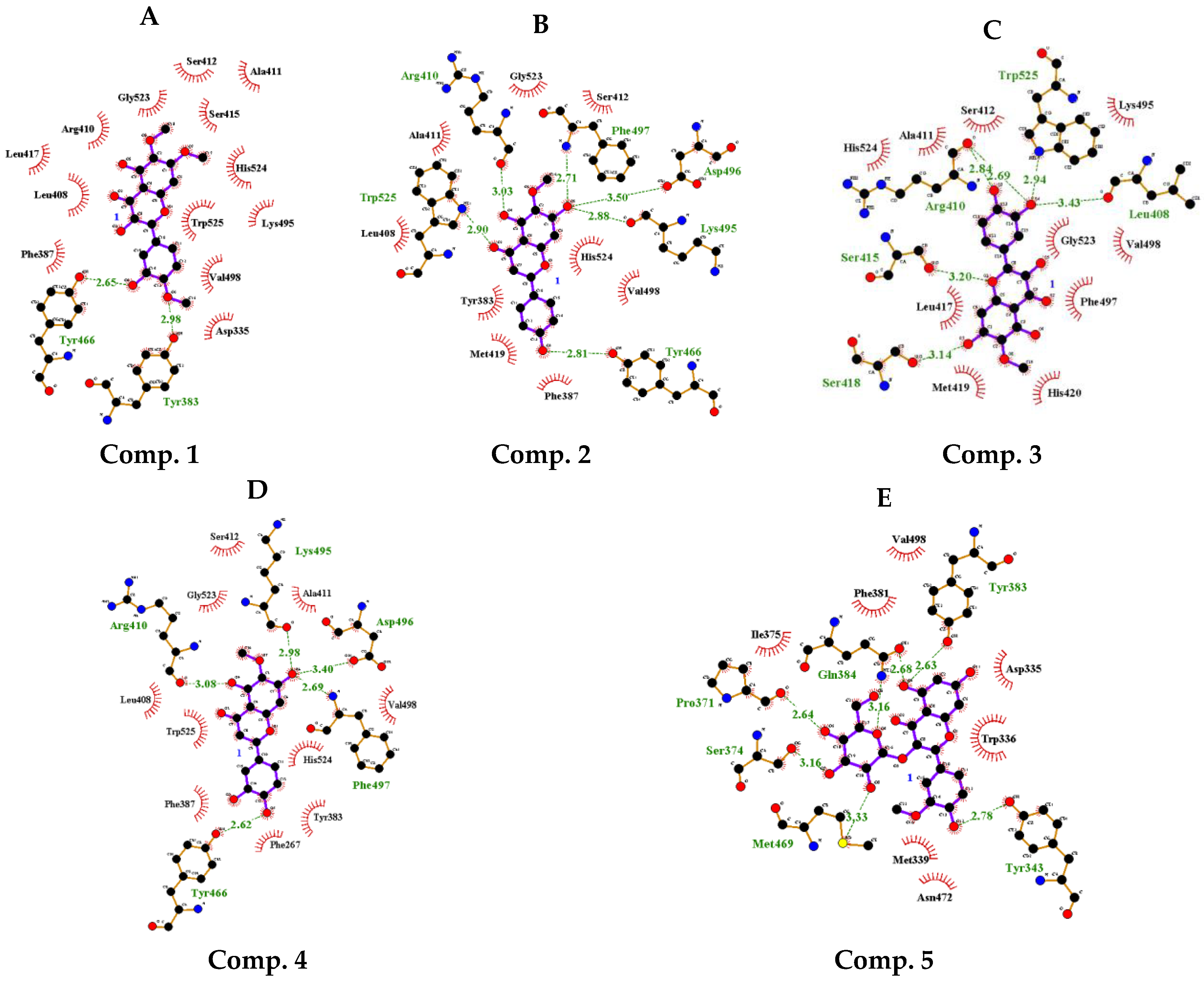
2.3. Static Molecular Modeling between Compounds 1–5 and sEH
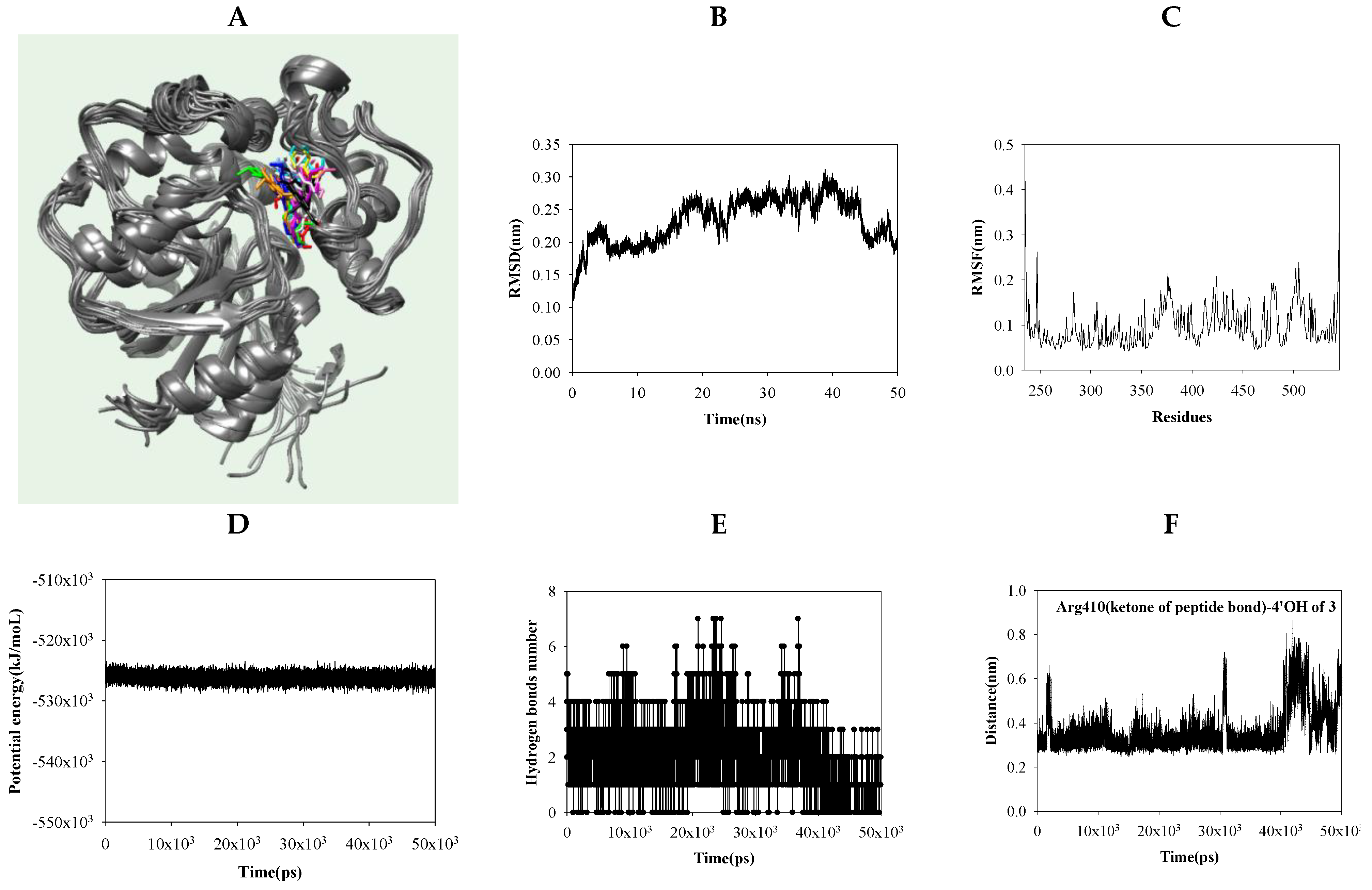
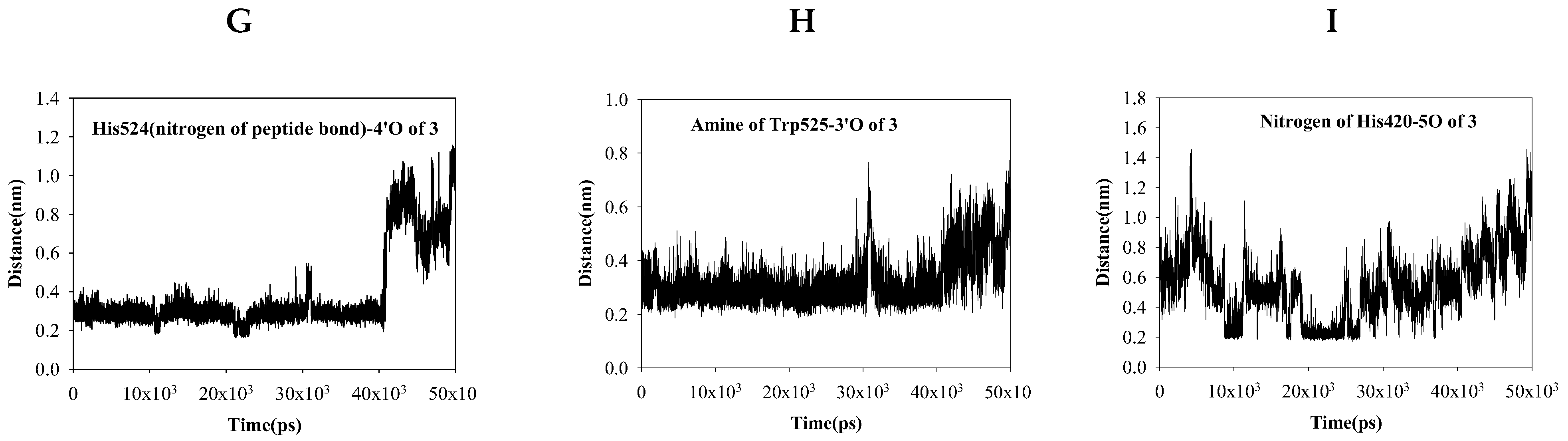
2.4. Dynamic Molecular Modeling between Compound 3 and sEH
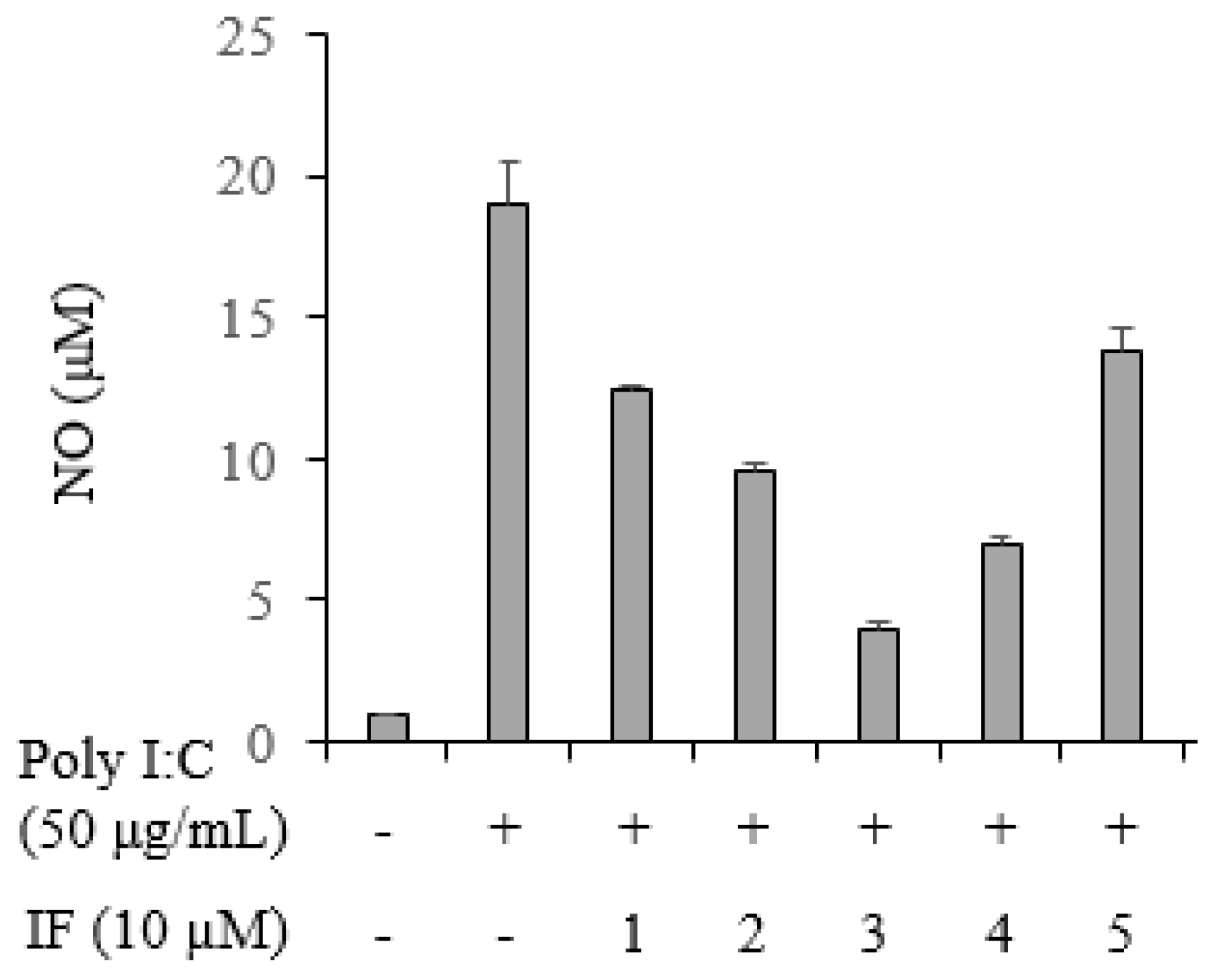
2.5. Inhibition of NO Production by Compounds 1–5 in Poly(I:C)-Stimulated RAW264.7 Cells
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Material
4.3. Extraction and Isolation
4.4. sEH Activity Assay
4.5. Molecular Docking
4.6. Molecular Dynamics
4.7. Cell Culture and Viability
4.8. Nitric Oxide Assay
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khan, A.L.; Hussain, J.; Hamayun, M.; Gilani, S.A.; Ahmad, S.; Rehman, G.; Kim, Y.H.; Kang, S.M.; Lee, I.J. Secondary metabolites from Inlula britannica L. and their biological activities. Molecules 2010, 15, 1562–1577. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, X.; Hou, A.; Zhang, J.; Wang, S.; Man, W.; Yu, H.; Zheng, S.; Wang, Q.; Jiang, H.; et al. A review of the botany, traditional uses, phytochemistry, and pharmacology of the Flos Inulae. J. Ethnopharmacol. 2021, 276, 114125. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.M.; Zhang, M.L.; Shi, Q.W.; Kiyota, H. Chemical constituents of plants from the genus Inula. Chem. Biodivers. 2006, 3, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Seca, A.M.; Grigore, A.; Pinto, D.C.; Silva, A.M. The genus Inula and their metabolites: From ethnopharmacological to medicinal uses. J. Ethnopharmacol. 2014, 154, 286–310. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.I.; Kim, J.H.; Lee, S.; Lee, I.S.; Park, J. Eupatin, a flavonoid, inhibits coronavirus 3CL protease and replication. Int. J. Mol. Sci. 2023, 24, 9211. [Google Scholar] [CrossRef]
- Sun, C.-P.; Zhang, X.-Y.; Morisseau, C.; Hwang, S.H.; Zhang, Z.-J.; Hammock, B.D.; Ma, X.-C. Discovery of soluble epoxide hydrolase inhibitors from chemical synthesis and natural products. J. Med. Chem. 2021, 64, 184–215. [Google Scholar] [CrossRef]
- Morisseau, C.; Hammock, B.D. Epoxide hydrolases: Mechanisms, inhibitor designs, and biological roles. Annu. Rev. Pharmacol. 2005, 45, 311–333. [Google Scholar] [CrossRef] [PubMed]
- Imig, J.D.; Hammock, B.D. Soluble epoxide hydrolase as a therapeutic target for cardiovascular diseases. Nat. Rev. Drug Discov. 2009, 8, 794–805. [Google Scholar] [CrossRef] [PubMed]
- Spector, A.A.; Fang, X.; Snyder, G.D.; Weintraub, N.L. Epoxyeicosatrienoic acids (EETs): Metabolism and biochemical function. Prog. Lipid Res. 2004, 43, 55–90. [Google Scholar] [CrossRef]
- Jung, O.; Brandes, R.P.; Kim, I.-H.; Schweda, F.; Schmidt, R.; Hammock, B.D.; Busse, R.; Fleming, I. Soluble epoxide hydrolase is a main effector of angiotensin Ⅱ–Induced hypertension. Hypertension 2005, 45, 759–765. [Google Scholar] [CrossRef]
- Morisseau, C.; Hammock, B.D. Impact of soluble epoxide hydrolase and epoxyeicosaoids on human health. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.-Y.; Zhangm, X.-Y.; Zhou, M.-R.; Tian, X.-G.; Lv, X.; Zhang, H.-L.; Deng, S.; Zhang, B.-J.; Sun, C.-P.; Ma, X.-C. Natural soluble epoxide hydrolase inhibitors from alisma orientale and their potential mechanism with soluble epoxide hydrolase. Inter. J. Biol. Macromol. 2021, 183, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Huh, Y.-C.; Hur, M.; Park, W.T.; Moon, Y.-H.; Kim, T.I.; Kim, S.M.; Koo, S.C. Inhibitory activity of lignanamides isolated from hemp seed hulls (Cannabis sativa L.) against soluble epoxide hydrolase. Heliyon 2023, 9, e19772. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Zhao, W.-Y.; Shao, B.; Zhang, B.-J.; Liu, T.-T.; Sun, C.-P.; Huang, H.-L.; Wu, J.-R.; Liang, J.-H.; Ma, X.-C. Natural soluble epoxide hydrolase inhibitors from Inula helenium and their interactions with soluble epoxide hydrolase. Int. J. Biol. Macromol. 2020, 158, 1362–1368. [Google Scholar] [CrossRef] [PubMed]
- Iwata, M.; Inoue, T.; Asai, Y.; Hori, K.; Fujiwara, M.; Matsuo, S.; Tsuchida, W.; Suzukim, S. The protective role of localized nitric oxide production during inflammation may be mediated by the heme oxygenase-1/carbon monoxide pathway. Biochem. Biophys. Rep. 2020, 23, 100790. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.L.; Megson, I.L.; Haslett, C.; Rossi, A.G. Nitric oxide: A key regulator of yeloid inflammatory cell apoptosis. Cell Death Differ. 2003, 10, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Maurice, D.A.; Pierre, T.; Hiroyuki, M. Isolation and structure characterization of flavonoids. In Flavonoids; Goncalo, C.J., Ed.; IntechOpen: Rijeka, Croatia, 2017; p. 3. [Google Scholar]
- Zeng, W.; Quesheng; Zhang, Q.; Liang, H. Flavonoids from Artemisia gmelinii Web. ex Stechm. J. Chin. Pharm. Sci. 2014, 23, 496–499. [Google Scholar] [CrossRef]
- Bai, N.S.; Zhou, Z.; Zhu, N.Q.; Zhang, L.; Quan, Z.; He, K.; Zheng, Q.Y.; Ho, C.T. Antioxidative flavonoids from the flower of Inula britannica. J. Food Lipids. 2005, 12, 141–149. [Google Scholar] [CrossRef]
- El-Hawiet, A.M.; Toaima, S.M.; Asaad, A.M.; Radwan, M.M.; El-Sebakhy, N.A. Chemical constituents from Astragalus annularis Forssk. and A. trimestris L., Fabaceae. Rev. Bras. Farmacogn. 2010, 20, 860–865. [Google Scholar] [CrossRef]
- Robinson, P.K. Enzyme: Principles and biotechnology applications. Essays Biochem. 2015, 59, 1–41. [Google Scholar] [CrossRef]
- Kim, J.H.; Cho, C.W.; Hur, M.; Park, W.T.; Moon, Y.H.; Koo, S.C.; Hur, Y.C.; Kang, J.S.; Lee, I.S. Inhibitory activity of quaternary isoquinoline alkaloids on soluble epoxide hydrolase. Curr. Issues Mol. Biol. 2022, 44, 4282–4289. [Google Scholar] [CrossRef]
- Kim, J.H.; Jin, C.H. Inhibitory activity of flavonoids, chrysoeriol and luteolin-7-O-glucopyranoside, on soluble epoxide hydrolase from Capsicum chinense. Biomolecules 2020, 10, 180. [Google Scholar] [CrossRef]
- Kim, J.H.; Thao, N.P.; Han, Y.K.; Lee, Y.S.; Luyen, B.T.T.; Oanh, H.V.; Kim, Y.H.; Yang, S.Y. The insight of in vitro and in silico studies on cholinesterase inhibitors from the roots of Cimicifuga dahurica (Turcz.) Maxim. J. Enzyme Inhib. Med. Chem. 2018, 33, 1174–1180. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.S.; Kim, J.H.; Lin, Y.; Su, X.D.; Kang, J.S.; Yang, S.Y.; Kim, Y.H. Inhibitory activity of quercetin 3-O-arabinofuranoside and 2-oxopomolic acid derived from Malus domestica on Soluble Epoxide Hydrolase. Molecules 2020, 25, 4352. [Google Scholar] [CrossRef]
- Bai, M.M.; Shi, W.; Tian, J.M.; Lei, M.; Kim, J.H.; Sun, Y.N.; Kim, Y.H.; Gao, J.M. Soluble epoxide hydrolase inhibitory and anti-inflammatory components from the leaves of Eucommia ulmoides Oliver (duzhong). J. Agric. Food Chem. 2015, 63, 2198–2205. [Google Scholar] [CrossRef]
- Dabeek, W.M.; Marra, M.V. Dietary qquercetin and kaempferol: Bioavailability and potential cardiovascular-related bioactivity in humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef]
- Sun, C.-P.; Zhang, J.; Zhao, W.-Y.; Yi, J.; Yan, J.-K.; Wang, Y.-L.; Morisseau, C.; Liu, Z.-B.; Hammock, B.D.; Ma, X.-C. Protostane-type triterpenoids as natural soluble epoxide hydrolase inhibitors: Inhibition potentials and molecular dynamics. Bioorg. Chem. 2020, 96, 103637. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, M.; Zhang, W.-H.; Zhum, Q.-M.; Hou, X.-K.; Sun, C.-P.; Ma, X.-C.; Xiao, H.-T. Total flavonoids of Inula japonica alleviated the inflammatory response and oxidative stress in LPS-induced acute lung injury via inhibiting the sEH activity: Insights from lipid metabolomics. Phytomedicine 2022, 107, 154380. [Google Scholar] [CrossRef] [PubMed]
- Leem, H.H.; Lee, G.Y.; Lee, J.S.; Lee, H.; Kim, J.H.; Kim, Y.H. Soluble epoxide hydrolase inhibitory activity of components from Leonurus japonicas. Int. J. Biol. Macromol. 2017, 103, 451–457. [Google Scholar] [CrossRef]
- Azuma, S.; Murakami, Y.; Azuma, E.; Baba, K.; Taniguchi, M. Anti-flammatory effects of flavonoids in Citrus jabara fruit peels. Trends Immunother. 2020, 4, 5–14. [Google Scholar] [CrossRef]
- Zhao, W.-Y.; Yan, J.-J.; Zhang, M.; Wang, C.; Feng, L.; Lv, X.; Hou, X.-K.; Sum, C.-P.; Chen, L.-X.; Ma, X.-C. Natural soluble epoxide hydrolase inhibitors from Inula britanica and their potential interactions with soluble epoxide hydrolase: Insight from inhibition kinetics and molecular dynamics. Chem. Biol. Interact. 2021, 345, 109571. [Google Scholar] [CrossRef] [PubMed]
- Rosselli Marinella RJ Keller Raghvendra, K. Role of nitric oxide in the biology, physiology and pathophysiology of reproduction. Hum. Reprod. Update 1998, 4, 3–24. [Google Scholar]
- Inkanuwat, A.; Sukaboon, R.; Reamtong, O.; Asawanonda, P.; Pattaratanakun, A.; Saisavoey, T.; Sangtanoo, P.; Karnchanatat, A. Nitric oxide synthesis inhibition and anti-inflammatory effect of polypeptide isolated from chicken feather meal in lipopolysaccharide-stimulated RAW264.7 macrophages. Food Technol. Biotechnol. 2019, 57, 200–212. [Google Scholar] [CrossRef] [PubMed]
- Jabeen, A.; Mesaik, M.A.; Simjee, S.U.; Lubna; Bano, S.; Faizi, S. Anti-TNF-α and anti-arthritic effect of patuletin: A rare flavonoid from tagetes patula. Inter. Immunopharm. 2016, 36, 232–240. [Google Scholar] [CrossRef]
- Arand, M.; Wager, H.; Oesch, F. Asp333, Asp495, and His523 form the catalytic triad of rat soluble epoxide hydrolase. J. Biol. Chem. 1996, 271, 4223–4229. [Google Scholar] [CrossRef]
- RCSB Protein Data Bank. Available online: https://www.rcsb.org/3d-view/3ANS (accessed on 11 April 2024).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 (µM) a | Binding Mode (µM) | |
|---|---|---|
| 1 | 42.6 ± 0.8 | non-competitive (43.9) |
| 2 | 22.2 ± 0.3 | non-competitive (19.8) |
| 3 | 12.1 ± 0.1 | non-competitive (17.5) |
| 4 | 40.9 ± 0.6 | non-competitive (63.5) |
| 5 | 62.8 ± 1.8 | non-competitive (68.7) |
| AUDA b | 12.3 ± 0.3 nM |
| Inhibitor | Hydrogen Bonds (Å) | Autodock Score (kcal/mol) |
|---|---|---|
| 1 | Tyr383(2.98), Tyr466(2.65) | −8.16 |
| 2 | Arg410(3.03), Tyr466(2.81), Lys495(2.88), Asp496(3.50), Phe497(2.71), Trp525(2.90) | −8.34 |
| 3 | Leu408(3.43), Arg410(2.84, 2.69), Ser415(3.20), Ser418(3.14), Trp525(2.94) | −8.59 |
| 4 | Arg410(3.08), Tyr466(2.62), Lys495(2.98), Asp496(3.40), Phe497(2.69), | −8.18 |
| 5 | Tyr343(2.78), Pro371(2.64), Ser374(3.16), Tyr383(2.63), Gln384(3.16), Met469(3.33) | −7.53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.H.; Han, K.-S.; Lee, E.-S.; Kim, Y.-G.; Kim, Y.-I.; Cho, B.O.; Lee, I.S. The Inhibition Activity of Natural Methoxyflavonoid from Inula britannica on Soluble Epoxide Hydrolase and NO Production in RAW264.7 Cells. Int. J. Mol. Sci. 2024, 25, 4357. https://doi.org/10.3390/ijms25084357
Kim JH, Han K-S, Lee E-S, Kim Y-G, Kim Y-I, Cho BO, Lee IS. The Inhibition Activity of Natural Methoxyflavonoid from Inula britannica on Soluble Epoxide Hydrolase and NO Production in RAW264.7 Cells. International Journal of Molecular Sciences. 2024; 25(8):4357. https://doi.org/10.3390/ijms25084357
Chicago/Turabian StyleKim, Jang Hoon, Kyung-Sook Han, Eun-Song Lee, Yong-Goo Kim, Yong-Il Kim, Byoung Ok Cho, and Ik Soo Lee. 2024. "The Inhibition Activity of Natural Methoxyflavonoid from Inula britannica on Soluble Epoxide Hydrolase and NO Production in RAW264.7 Cells" International Journal of Molecular Sciences 25, no. 8: 4357. https://doi.org/10.3390/ijms25084357
APA StyleKim, J. H., Han, K. -S., Lee, E. -S., Kim, Y. -G., Kim, Y. -I., Cho, B. O., & Lee, I. S. (2024). The Inhibition Activity of Natural Methoxyflavonoid from Inula britannica on Soluble Epoxide Hydrolase and NO Production in RAW264.7 Cells. International Journal of Molecular Sciences, 25(8), 4357. https://doi.org/10.3390/ijms25084357