
In Vitro and In Silico Studies on Cytotoxic Properties of Oxythiamine and 2′-Methylthiamine

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
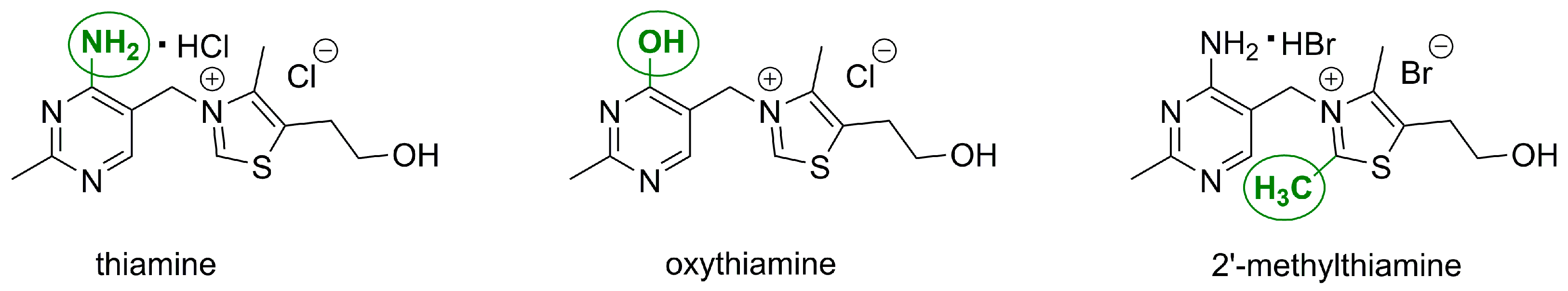
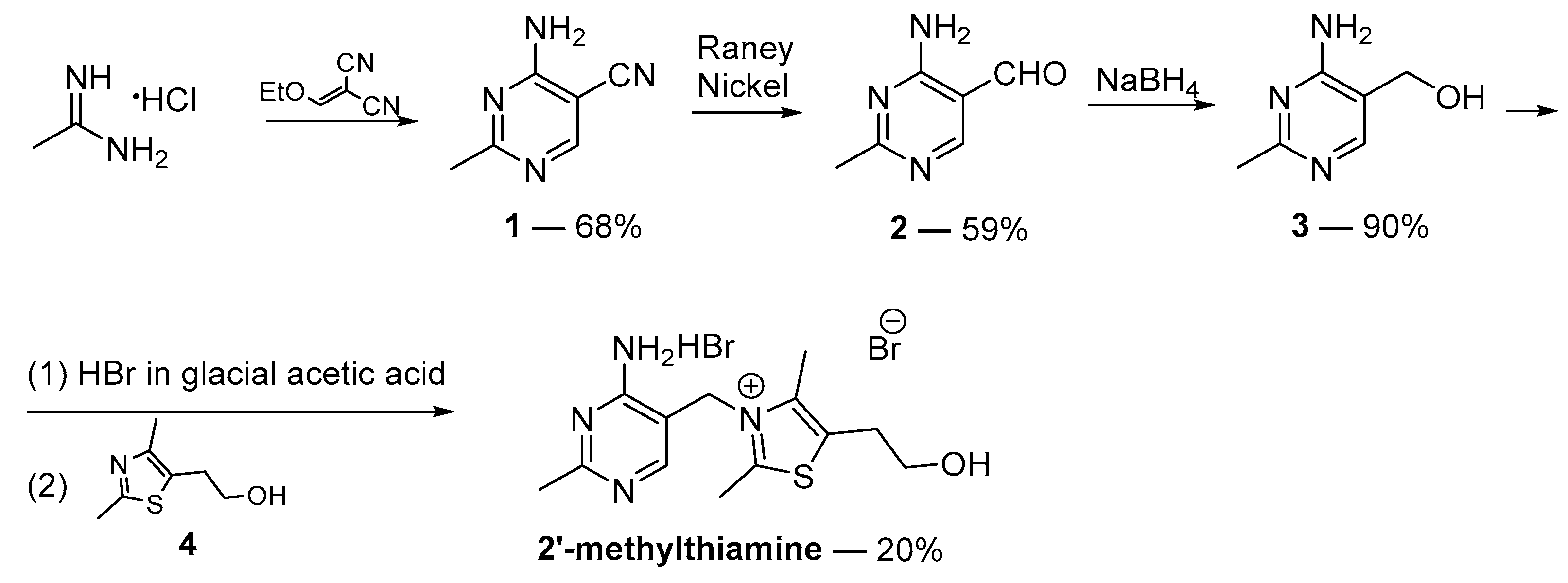
2.1. Synthesis of 2′-Methylthiamine
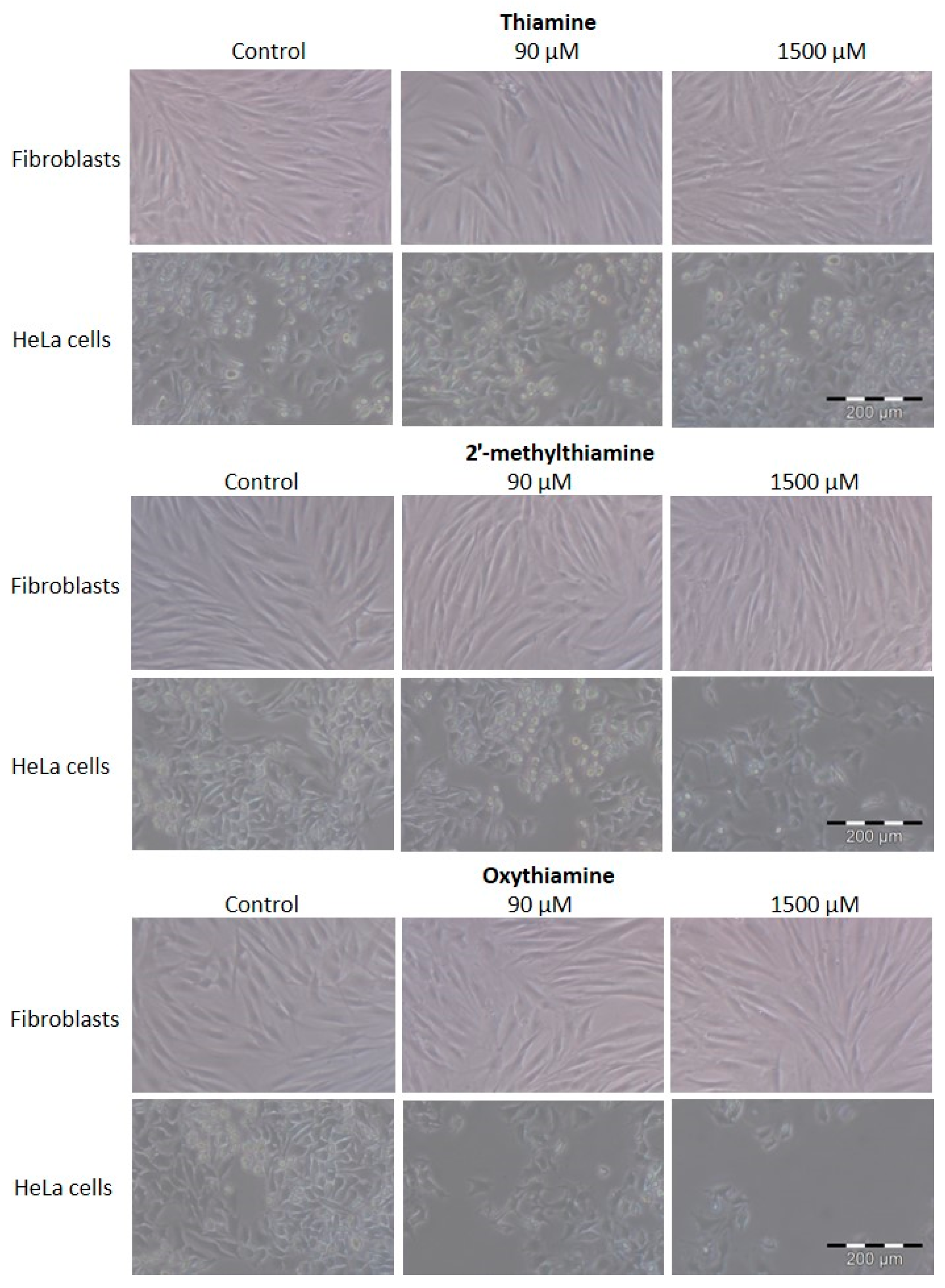
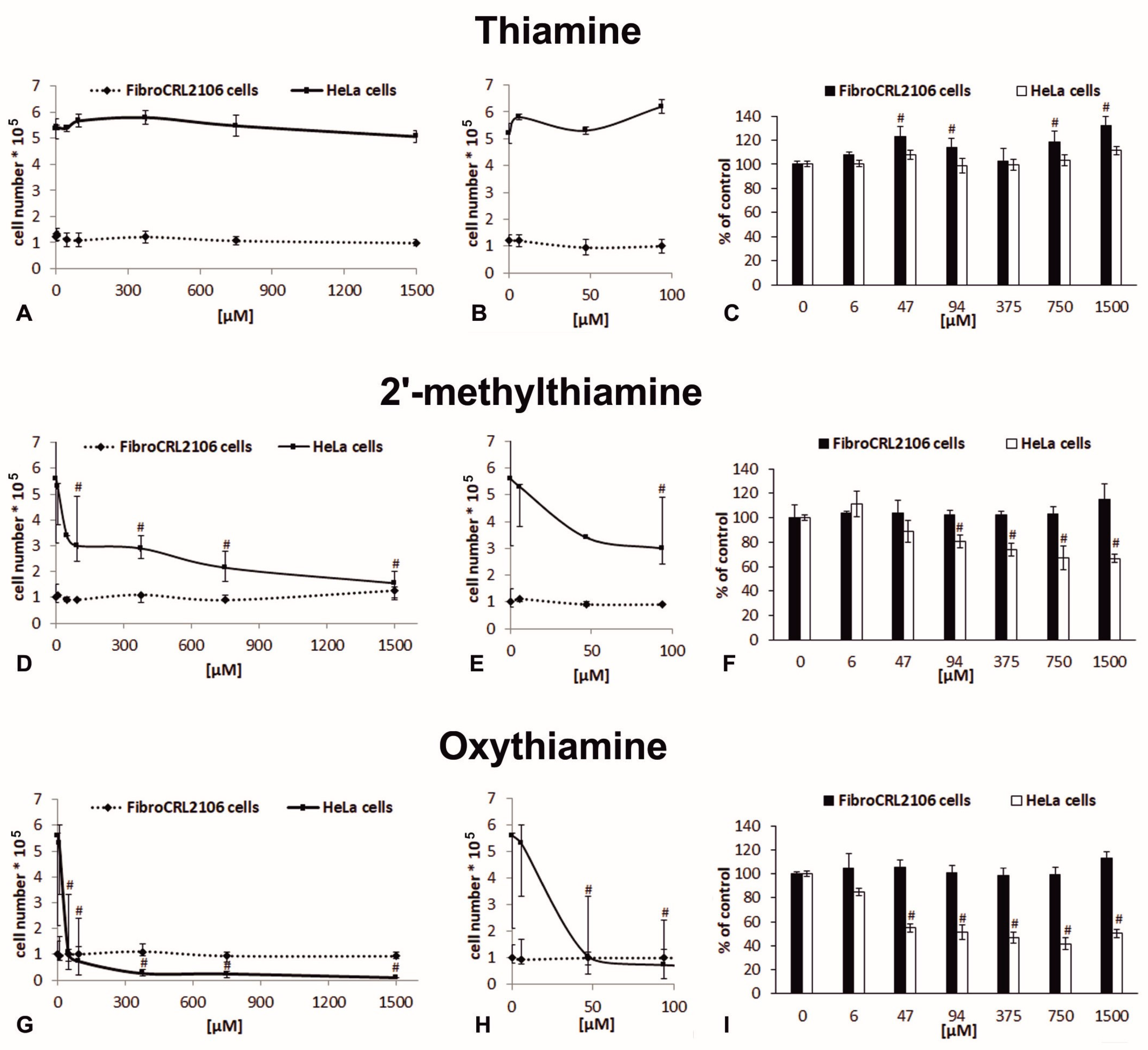
2.2. The Effects of Oxythiamine and 2′-Methylthiamine on Cells In Vitro
2.3. Molecular Docking
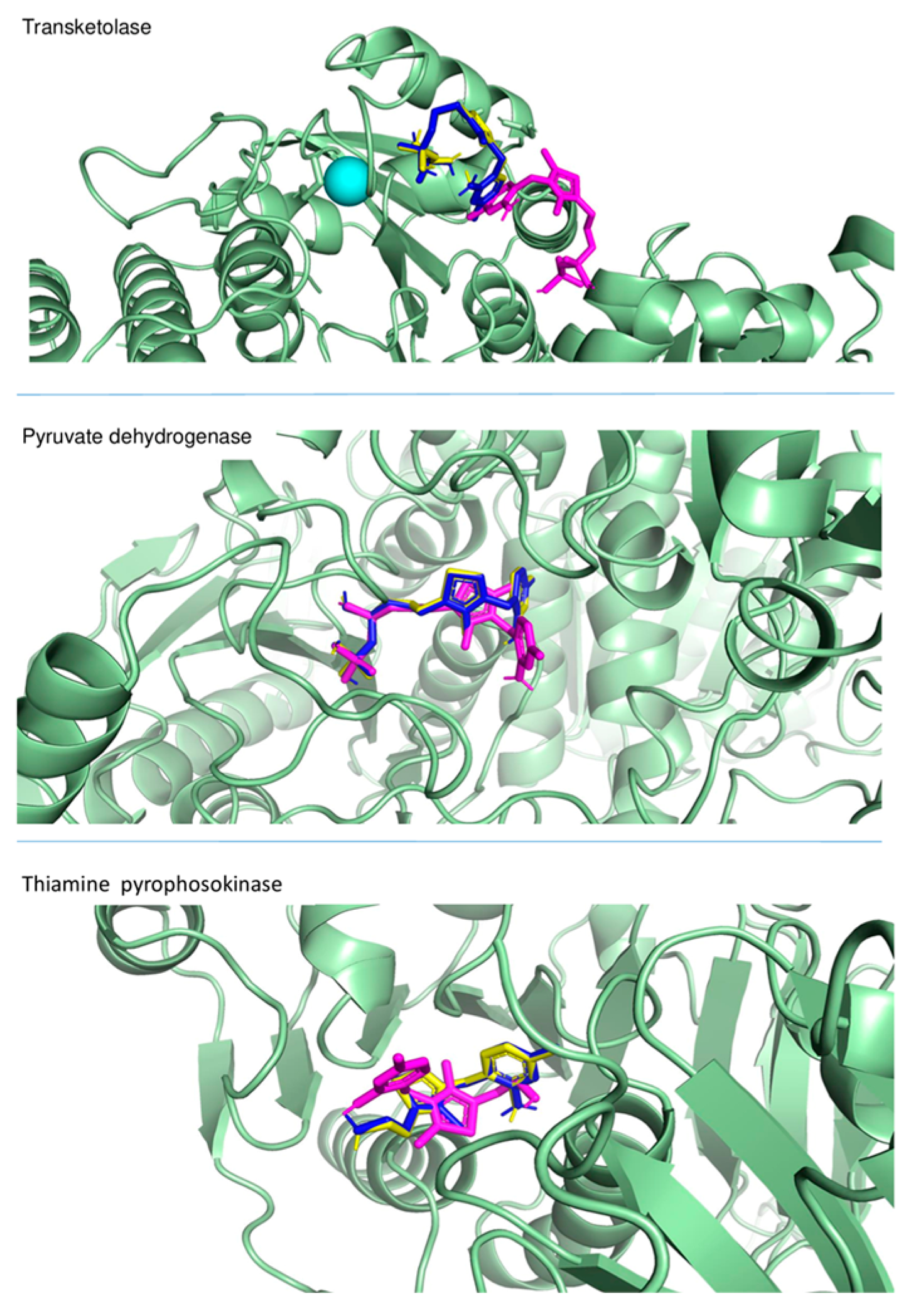
2.3.1. Docking to Enzymes—Affinities
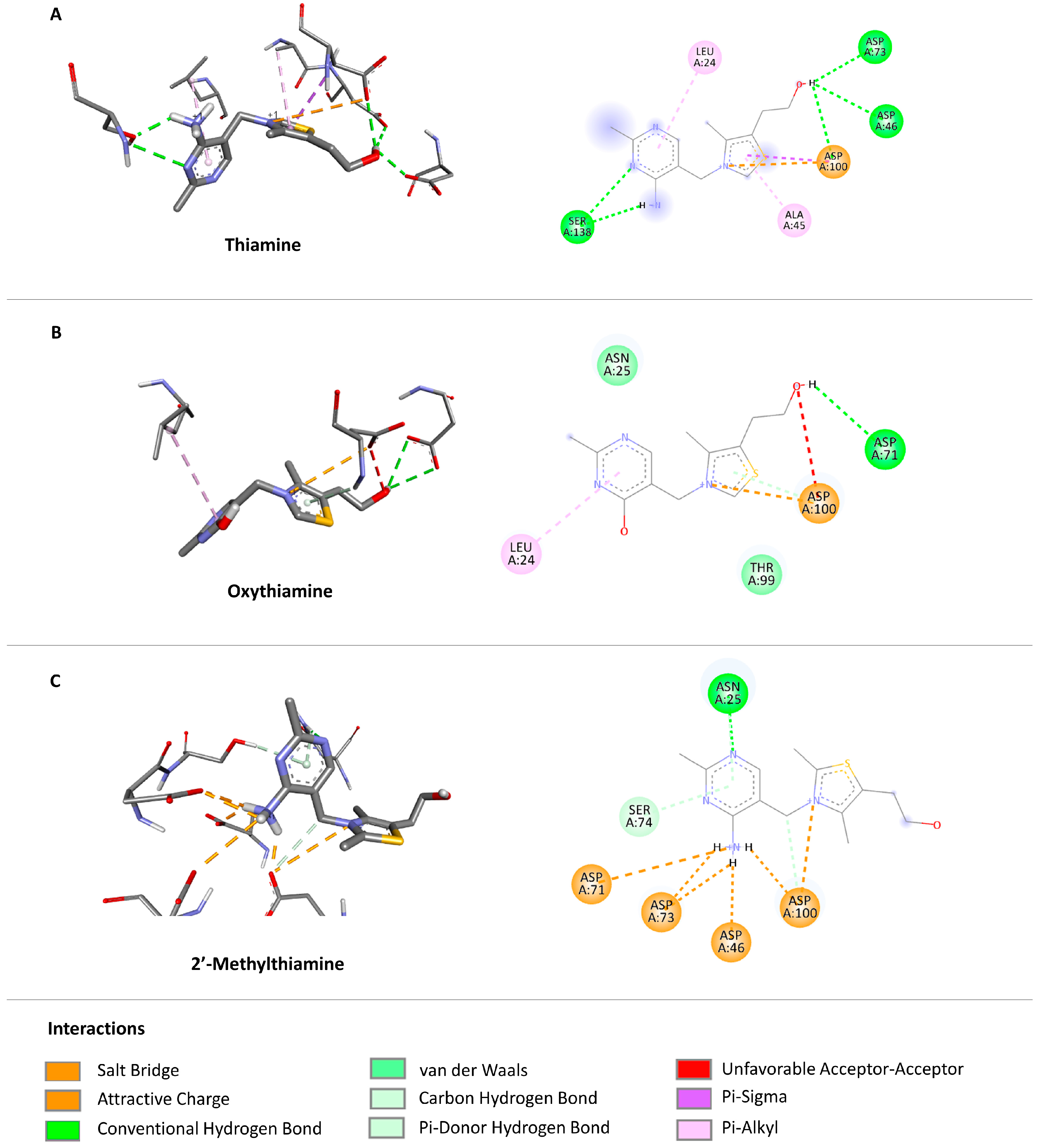
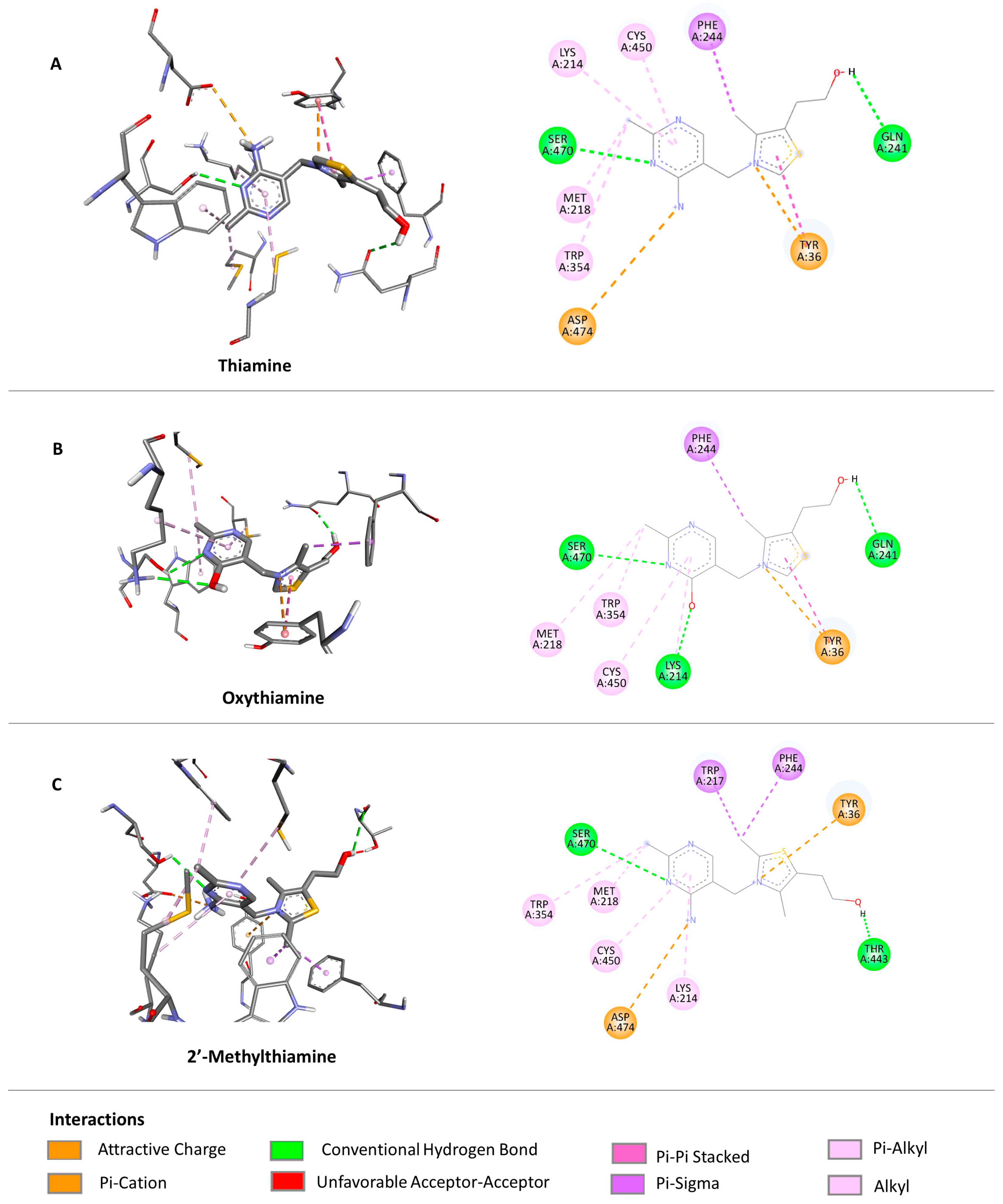
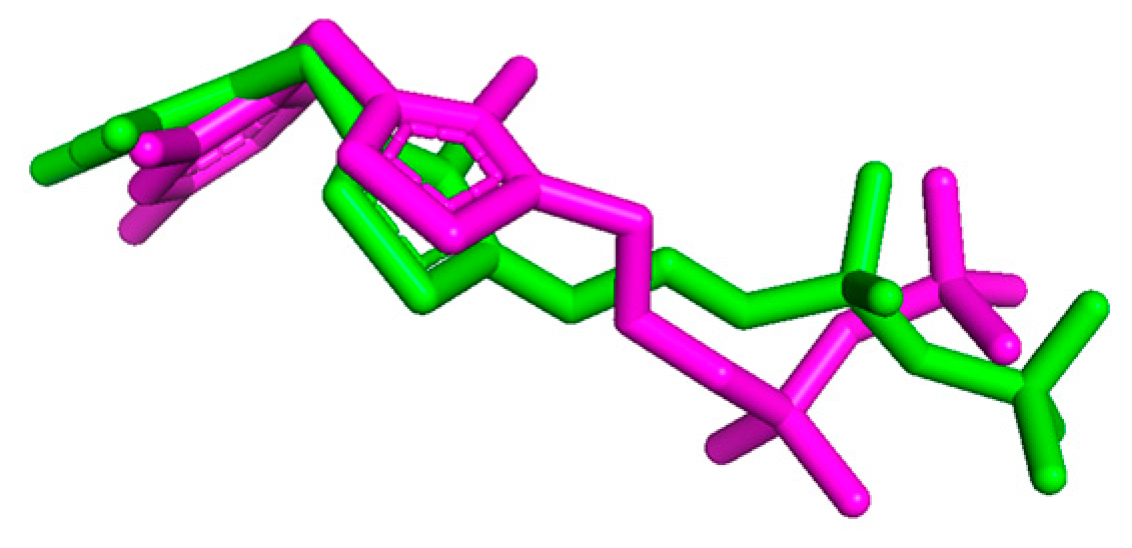
2.3.2. Docking to Enzymes—Ligand Arrangements
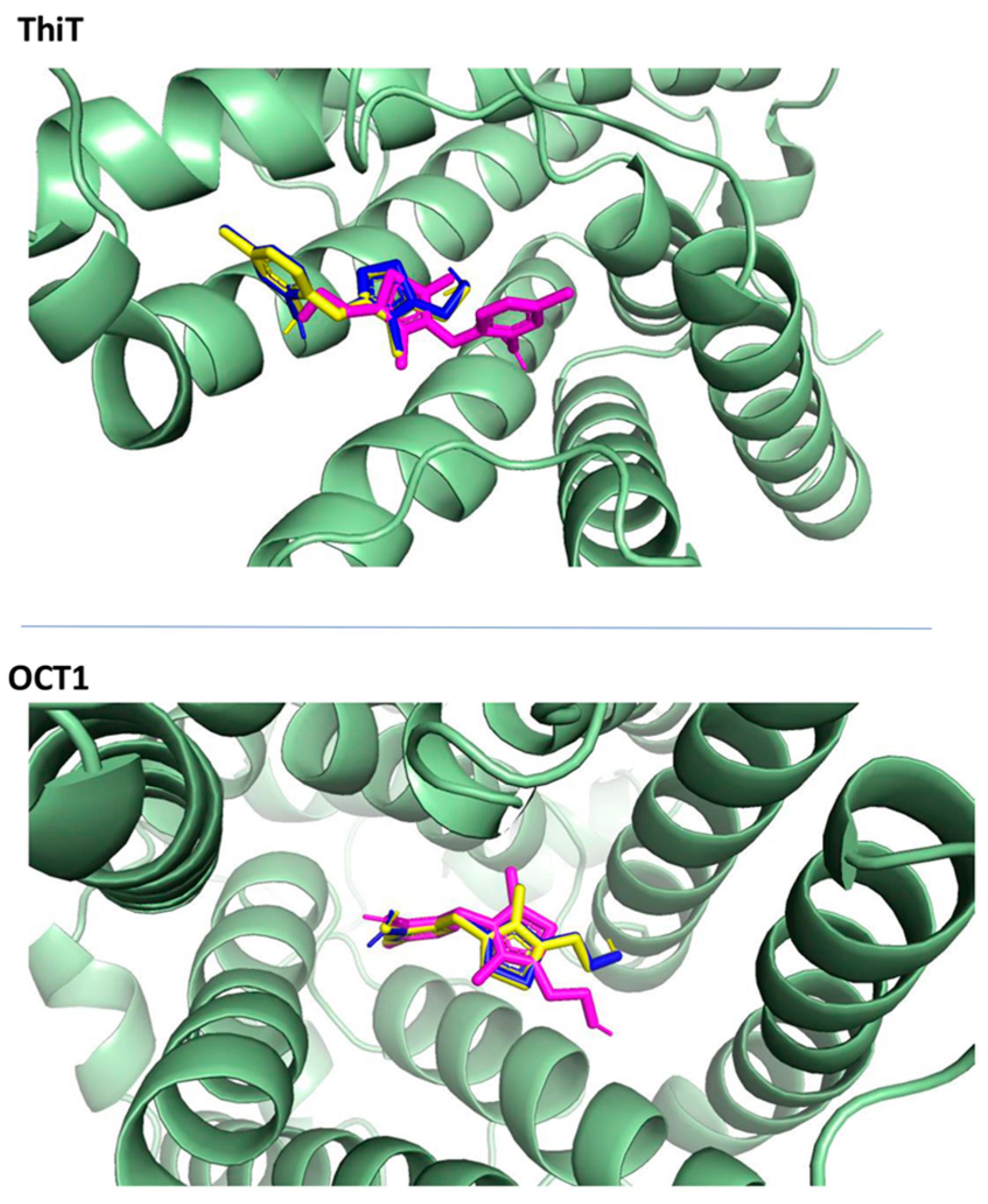
2.3.3. Docking to Transporters
2.3.4. Docking Statistics
2.3.5. Docking—Discussion
2.3.6. Docking—Future Directions
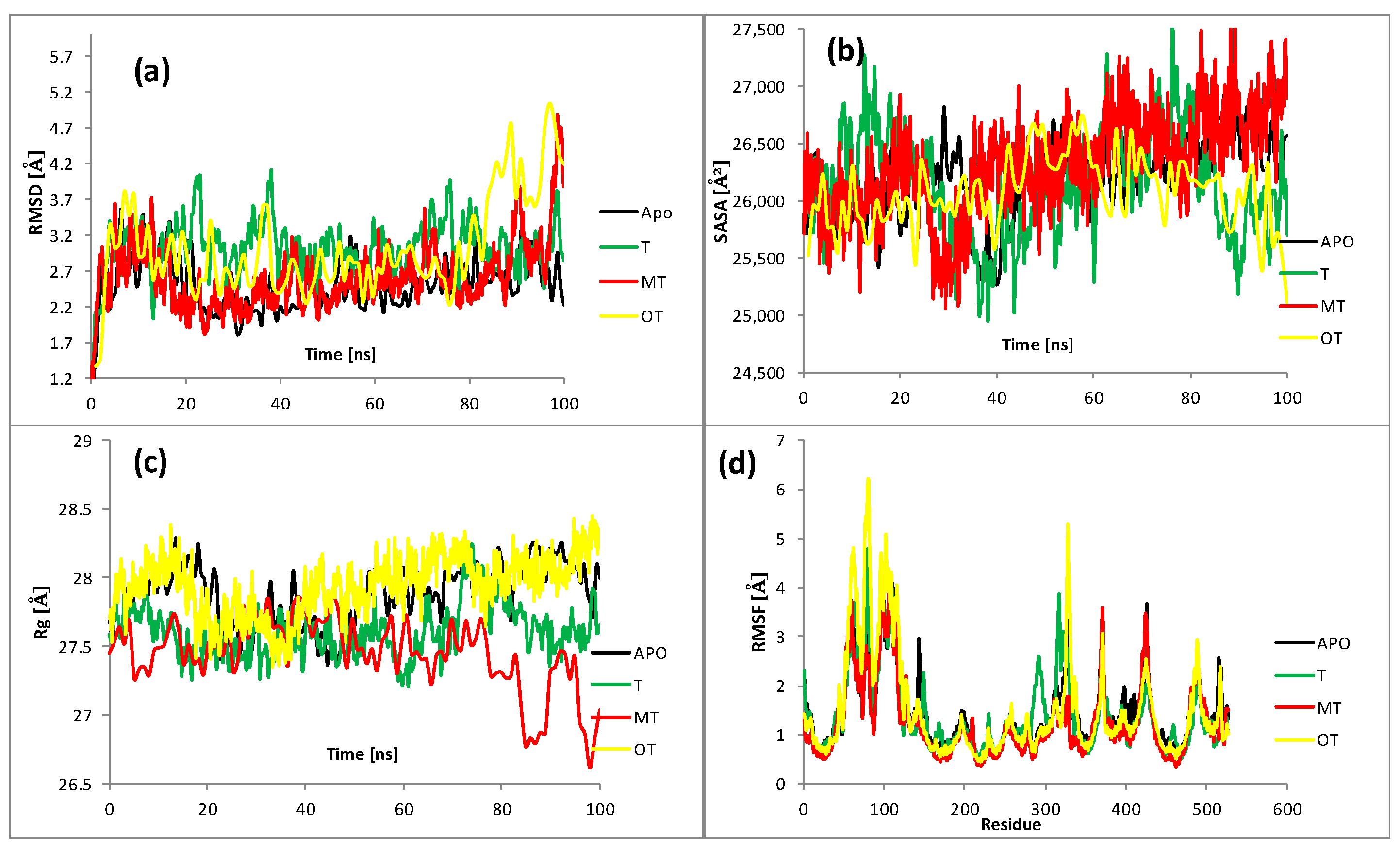
2.4. Molecular Dynamics
3. Materials and Methods
3.1. General Information on the Synthesis of 2′-Methylthiamine
3.1.1. 4-Amino-2-Methylpyrimidine-5-Carbonitrile (1) [56]
3.1.2. 4-Amino-2-Methylpyrimidine-5-Carbaldehyde (2) [57]
3.1.3. (4-Amino-2-Methylpyrimidin-5-yl)Methanol (3) [58]
3.1.4. 2-[3-[(4-Amino-2-Methylpyrimidin-5-yl)methyl]-2,4-dimethyl-1,3-thiazol-3-ium-5-yl]ethanol (2′-Methylthiamine)
3.2. In Vitro Cell Culture and Cytotoxicity Test
3.3. Molecular Docking
3.4. Molecular Dynamics
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mrowicka, M.; Mrowicki, J.; Dragan, G.; Majsterek, I. The Importance of Thiamine (Vitamin B1) in Humans. Biosci. Rep. 2023, 43, BSR20230374. [Google Scholar] [CrossRef]
- Nauton, L.; Hélaine, V.; Théry, V.; Hecquet, L. Insights into the Thiamine Diphosphate Enzyme Activation Mechanism: Computational Model for Transketolase Using a Quantum Mechanical/Molecular Mechanical Method. Biochemistry 2016, 55, 2144–2152. [Google Scholar] [CrossRef]
- Solovjeva, O.N. New Role of Water in Transketolase Catalysis. Int. J. Mol. Sci. 2023, 24, 2068. [Google Scholar] [CrossRef]
- Park, S.; Jeon, J.H.; Min, B.K.; Ha, C.M.; Thoudam, T.; Park, B.Y.; Lee, I.K. Role of the Pyruvate Dehydrogenase Complex in Metabolic Remodeling: Differential Pyruvate Dehydrogenase Complex Functions in Metabolism. Diabetes Metab. J. 2018, 42, 270. [Google Scholar] [CrossRef]
- Nasiri, A.; Sadeghi, M.; Vaisi-Raygani, A.; Kiani, S.; Aghelan, Z.; Khodarahmi, R. Emerging Regulatory Roles of Mitochondrial Sirtuins on Pyruvate Dehydrogenase Complex and the Related Metabolic Diseases: Review. Biomed. Res. Ther. 2020, 7, 3645–3658. [Google Scholar] [CrossRef]
- Hansen, G.E.; Gibson, G.E. The α-Ketoglutarate Dehydrogenase Complex as a Hub of Plasticity in Neurodegeneration and Regeneration. Int. J. Mol. Sci. 2022, 23, 12403. [Google Scholar] [CrossRef]
- Kochetov, G.A.; Solovjeva, O.N. Structure and Functioning Mechanism of Transketolase. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2014, 1844, 1608–1618. [Google Scholar] [CrossRef]
- Hao, S.; Meng, Q.; Sun, H.; Li, Y.; Li, Y.; Gu, L.; Liu, B.; Zhang, Y.; Zhou, H.; Xu, Z.; et al. The Role of Transketolase in Human Cancer Progression and Therapy. Biomed. Pharmacother. 2022, 154, 113607. [Google Scholar] [CrossRef]
- Budnik, V.I.; Tylicki, A.; Lukashev, N.V. Thiamin Diphosphate-dependent Enzymes: From Enzymology to Metabolic Regulation, Drug Design and Disease Models. FEBS J. 2013, 280, 6412–6442. [Google Scholar] [CrossRef]
- Tylicki, A.; Łotowski, Z.; Siemieniuk, M.; Ratkiewicz, A. Thiamine and Selected Thiamine Antivitamins—Biological Activity and Methods of Synthesis. Biosci. Rep. 2018, 38, BSR20171148. [Google Scholar] [CrossRef]
- Bettendorff, L. Synthetic Thioesters of Thiamine: Promising Tools for Slowing Progression of Neurodegenerative Diseases. Int. J. Mol. Sci. 2023, 24, 11296. [Google Scholar] [CrossRef]
- Szoka, L.; Karna, E.; Palka, J. The Mechanism of Oxythiamine-Induced Collagen Biosynthesis in Cultured Fibroblasts. Mol. Cell Biochem. 2015, 403, 51–60. [Google Scholar] [CrossRef]
- Yang, C.-M.; Liu, Y.-Z.; Liao, J.-W.; Hu, M.-L. The in Vitro and in Vivo Anti-Metastatic Efficacy of Oxythiamine and the Possible Mechanisms of Action. Clin. Exp. Metastasis 2010, 27, 341–349. [Google Scholar] [CrossRef]
- Bai, L.; Zhu, H. A Dose- and Time-Dependent Effect of Oxythiamine on Cell Growth Inhibition in Non-Small Cell Lung Cancer. Cogn. Neurodyn 2022, 16, 633–641. [Google Scholar] [CrossRef]
- Boros, L.G.; Puigjaner, J.; Cascante, M.; Lee, W.-N.P.; Brandes, J.L.; Bassilian, S.; Yusuf, F.I.; Williams, R.D.; Muscarella, P.; Melvin, W.S.; et al. Oxythiamine and Dehydroepiandrosterone Inhibit the Nonoxidative Synthesis of Ribose and Tumor Cell Proliferation1. Cancer Res. 1997, 57, 4242–4248. [Google Scholar]
- Wang, J.; Zhang, X.; Ma, D.; Lee, W.-N.P.; Xiao, J.; Zhao, Y.; Go, V.L.; Wang, Q.; Yen, Y.; Recker, R.; et al. Inhibition of Transketolase by Oxythiamine Altered Dynamics of Protein Signals in Pancreatic Cancer Cells. Exp. Hematol. Oncol. 2013, 2, 18. [Google Scholar] [CrossRef]
- Raïs, B.; Comin, B.; Puigjaner, J.; Brandes, J.L.; Creppy, E.; Saboureau, D.; Ennamany, R.; Paul Lee, W.-N.; Boros, L.G.; Cascante, M. Oxythiamine and Dehydroepiandrosterone Induce a G1 Phase Cycle Arrest in Ehrlich’s Tumor Cells through Inhibition of the Pentose Cycle. FEBS Lett. 1999, 456, 113–118. [Google Scholar] [CrossRef]
- Siemieniuk, M.; Sosnowska, K.; Czerniecki, J.; Czyzewska, U.; Winnicka, K.; Tylicki, A. Oxythiamine Improves Antifungal Activity of Ketoconazole Evaluated in Canine Malassezia Pachydermatis Strains. Vet. Dermatol. 2018, 29, 476-e160. [Google Scholar] [CrossRef]
- Yan, M.; Smeets, R.; Gosau, M.; Vollkommer, T.; Fuest, S.; Stetzer, E.; Kluwe, L.; Coy, J.F.; Burg, S. Tolerance of Human Fibroblasts to Benfo-Oxythiamine In Vitro. Int. J. Environ. Res. Public Health 2022, 19, 4112. [Google Scholar] [CrossRef]
- Chan, A.H.Y.; Ho, T.C.S.; Fathoni, I.; Pope, R.; Saliba, K.J.; Leeper, F.J. Inhibition of Thiamine Diphosphate-Dependent Enzymes by Triazole-Based Thiamine Analogues. ACS Med. Chem. Lett. 2023, 14, 621–628. [Google Scholar] [CrossRef]
- Schellenberger, A.; Winter, K.; Hübner, G.; Schwaiberger, R.; Helbig, D.; Schumacher, S.; Thieme, R.; Bouillon, G.; Rädler, K.-P. Zur Theorie Der Thiaminpyrophosphat-Wirkung, III. Rekombinations-Und Funktionskinetik Der Hefe-Pyruvat-Decarboxylase. Biol. Chem. 1966, 346, 123–147. [Google Scholar] [CrossRef]
- Bag, S.; Tawari, N.R.; Degani, M.S.; Queener, S.F. Design, Synthesis, Biological Evaluation and Computational Investigation of Novel Inhibitors of Dihydrofolate Reductase of Opportunistic Pathogens. Bioorganic Med. Chem. 2010, 18, 3187–3197. [Google Scholar] [CrossRef] [PubMed]
- Baxter, R.L.; Hartley, A.B.; Chan, H.W.S. Thiamine Biosynthesis in Yeast—Evaluation of 4-Hydroxy-5-Hydroxymethyl-2-Methylpyrimidine as a Precursor. J. Chem. Soc. Perkin Trans. 1990, 11, 2963–2966. [Google Scholar] [CrossRef]
- Piper, J.R.; Johnson, C.A.; Maddry, J.A.; Malik, N.D.; McGuire, J.J.; Otter, G.M.; Sirotnak, F.M. Studies on Analogs of Classical Antifolates Bearing the Naphthoyl Group in Place of Benzoyl in the Side Chain. J. Med. Chem. 1993, 36, 4161–4171. [Google Scholar] [CrossRef] [PubMed]
- Archer, S.; Perianayagam, C. An Attempt to Apply Lethal Synthesis to the Design of Chemotherapeutic Agents. Fluorinated 5.Beta.-(Hydroxyethyl)-4-Methylthiazoles. J. Med. Chem. 1979, 22, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Boyd, S.A.; Condroski, K.R.; Thomas, A.; Gonzales, S.S.; Gunawardana, I.W.; Le Huerou, Y.; Romoff, T.T.; Sullivan, F.X.; Thioalkeneamides as Transketolase Inhibitors. WO/2005/095344, Publication Date 13 October 2005. Available online: https://patentscope.wipo.int/search/en/detail.jsf?docId=WO2005095344 (accessed on 25 February 2024).
- Thomas, A.A.; Le Huerou, Y.; De Meese, J.; Gunawardana, I.; Kaplan, T.; Romoff, T.T.; Gonzales, S.S.; Condroski, K.; Boyd, S.A.; Ballard, J.; et al. Synthesis, in Vitro and in Vivo Activity of Thiamine Antagonist Transketolase Inhibitors. Bioorganic Med. Chem. Lett. 2008, 18, 2206–2210. [Google Scholar] [CrossRef] [PubMed]
- Hanberry, B.S.; Berger, R.; Zastre, J.A. High Dose Vitamin B1 Reduces Proliferation in Cancer Cell Lines Analogous to Dichloroacetate. Cancer Chemother. Pharmacol. 2014, 73, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-Y.; Timm, D.E.; Hurley, T.D. Pyrithiamine as a Substrate for Thiamine Pyrophosphokinase. J. Biol. Chem. 2006, 281, 6601–6607. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Masania, J.; Anwar, A.; Xue, M.; Zehnder, D.; Kanji, H.; Rabbani, N.; Thornalley, P.J. The Uremic Toxin Oxythiamine Causes Functional Thiamine Deficiency in End-Stage Renal Disease by Inhibiting Transketolase Activity. Kidney Int. 2016, 90, 396–403. [Google Scholar] [CrossRef]
- Ricciardelli, C.; Lokman, N.A.; Cheruvu, S.; Tan, I.A.; Ween, M.P.; Pyragius, C.E.; Ruszkiewicz, A.; Hoffmann, P.; Oehler, M.K. Transketolase Is Upregulated in Metastatic Peritoneal Implants and Promotes Ovarian Cancer Cell Proliferation. Clin. Exp. Metastasis 2015, 32, 441–455. [Google Scholar] [CrossRef]
- Xu, I.M.-J.; Lai, R.K.-H.; Lin, S.-H.; Tse, A.P.-W.; Chiu, D.K.-C.; Koh, H.-Y.; Law, C.-T.; Wong, C.-M.; Cai, Z.; Wong, C.C.-L.; et al. Transketolase Counteracts Oxidative Stress to Drive Cancer Development. Proc. Natl. Acad. Sci. USA 2016, 113, E725–E734. [Google Scholar] [CrossRef]
- Tseng, C.-W.; Kuo, W.-H.; Chan, S.-H.; Chan, H.-L.; Chang, K.-J.; Wang, L.-H. Transketolase Regulates the Metabolic Switch to Control Breast Cancer Cell Metastasis via the α-Ketoglutarate Signaling Pathway. Cancer Res. 2018, 78, 2799–2812. [Google Scholar] [CrossRef]
- Gorbach, Z.V.; Kubyshin, V.L.; Maglysh, S.S.; Zabrodskaia, S.V. Metabolism of transketolase coenzyme in the rat liver. Biokhimiia 1986, 51, 1093–1099. [Google Scholar]
- Datta, A.G.; Racker, E. Mechanism of Action of Transketolase. I. Properties of the Crystalline Yeast Enzyme. J. Biol. Chem. 1961, 236, 617–623. [Google Scholar] [CrossRef]
- Peterson, J.W.; Gubler, C.J.; Kuby, S.A. Partial Purification and Properties of Thiamine Pyrophosphokinase from Pig Brain. Biochim. Biophys. Acta (BBA)-Enzymol. 1975, 397, 377–394. [Google Scholar] [CrossRef]
- Bettendorff, L.; Wins, P. Mechanism of Thiamine Transport in Neuroblastoma Cells. Inhibition of a High Affinity Carrier by Sodium Channel Activators and Dependence of Thiamine Uptake on Membrane Potential and Intracellular ATP. J. Biol. Chem. 1994, 269, 14379–14385. [Google Scholar] [CrossRef]
- Zhao, R.; Gao, F.; Goldman, I.D. Molecular Cloning of Human Thiamin pyrophosphokinase1The Sequence Data in This Paper Have Been Submitted to GenBank under the Accession Number AF297710.1. Biochim. Biophys. Acta (BBA)-Gene Struct. Expr. 2001, 1517, 320–322. [Google Scholar] [CrossRef]
- Zastre, J.A.; Sweet, R.L.; Hanberry, B.S.; Ye, S. Linking Vitamin B1 with Cancer Cell Metabolism. Cancer Metab. 2013, 1, 16. [Google Scholar] [CrossRef]
- Zhao, R.; Goldman, I.D. Folate and Thiamine Transporters Mediated by Facilitative Carriers (SLC19A1-3 and SLC46A1) and Folate Receptors. Mol. Asp. Med. 2013, 34, 373–385. [Google Scholar] [CrossRef]
- Palmieri, F.; Monné, M.; Fiermonte, G.; Palmieri, L. Mitochondrial Transport and Metabolism of the Vitamin B-Derived Cofactors Thiamine Pyrophosphate, Coenzyme A, FAD and NAD+, and Related Diseases: A Review. IUBMB Life 2022, 74, 592–617. [Google Scholar] [CrossRef]
- Lozano, E.; Herraez, E.; Briz, O.; Robledo, V.S.; Hernandez-Iglesias, J.; Gonzalez-Hernandez, A.; Marin, J.J.G. Role of the Plasma Membrane Transporter of Organic Cations OCT1 and Its Genetic Variants in Modern Liver Pharmacology. BioMed Res. Int. 2013, 2013, e692071. [Google Scholar] [CrossRef]
- Chen, L.; Shu, Y.; Liang, X.; Chen, E.C.; Yee, S.W.; Zur, A.A.; Li, S.; Xu, L.; Keshari, K.R.; Lin, M.J.; et al. OCT1 Is a High-Capacity Thiamine Transporter That Regulates Hepatic Steatosis and Is a Target of Metformin. Proc. Natl. Acad. Sci. USA 2014, 111, 9983–9988. [Google Scholar] [CrossRef]
- Liang, X.; Yee, S.W.; Chien, H.-C.; Chen, E.C.; Luo, Q.; Zou, L.; Piao, M.; Mifune, A.; Chen, L.; Calvert, M.E.; et al. Organic Cation Transporter 1 (OCT1) Modulates Multiple Cardiometabolic Traits through Effects on Hepatic Thiamine Content. PLoS Biol. 2018, 16, e2002907. [Google Scholar] [CrossRef]
- Jensen, O.; Matthaei, J.; Blome, F.; Schwab, M.; Tzvetkov, M.V.; Brockmöller, J. Variability and Heritability of Thiamine Pharmacokinetics With Focus on OCT1 Effects on Membrane Transport and Pharmacokinetics in Humans. Clin. Pharmacol. Ther. 2020, 107, 628–638. [Google Scholar] [CrossRef]
- Otter, M.; Csader, S.; Keiser, M.; Oswald, S. Expression and Functional Contribution of Different Organic Cation Transporters to the Cellular Uptake of Doxorubicin into Human Breast Cancer and Cardiac Tissue. Int. J. Mol. Sci. 2022, 23, 255. [Google Scholar] [CrossRef]
- Heise, M.; Lautem, A.; Knapstein, J.; Schattenberg, J.M.; Hoppe-Lotichius, M.; Foltys, D.; Weiler, N.; Zimmermann, A.; Schad, A.; Gründemann, D.; et al. Downregulation of Organic Cation Transporters OCT1 (SLC22A1) and OCT3 (SLC22A3) in Human Hepatocellular Carcinoma and Their Prognostic Significance. BMC Cancer 2012, 12, 109. [Google Scholar] [CrossRef]
- Lautem, A.; Heise, M.; Gräsel, A.; Hoppe-Lotichius, M.; Weiler, N.; Foltys, D.; Knapstein, J.; Schattenberg, J.M.; Schad, A.; Zimmermann, A.; et al. Downregulation of Organic Cation Transporter 1 (SLC22A1) Is Associated with Tumor Progression and Reduced Patient Survival in Human Cholangiocellular Carcinoma. Int. J. Oncol. 2013, 42, 1297–1304. [Google Scholar] [CrossRef]
- Edemir, B. Identification of Prognostic Organic Cation and Anion Transporters in Different Cancer Entities by In Silico Analysis. Int. J. Mol. Sci. 2020, 21, 4491. [Google Scholar] [CrossRef]
- Monjas, L.; Swier, L.J.Y.M.; Setyawati, I.; Slotboom, D.J.; Hirsch, A.K.H. Dynamic Combinatorial Chemistry to Identify Binders of ThiT, an S-Component of the Energy-Coupling Factor Transporter for Thiamine. ChemMedChem 2017, 12, 1693–1696. [Google Scholar] [CrossRef]
- Swier, L.J.Y.M.; Monjas, L.; Reeßing, F.; Oudshoorn, R.C.; Primke, T.; Bakker, M.M.; van Olst, E.; Ritschel, T.; Faustino, I.; Marrink, S.J.; et al. Insight into the Complete Substrate-Binding Pocket of ThiT by Chemical and Genetic Mutations. Med. Chem. Commun. 2017, 8, 1121–1130. [Google Scholar] [CrossRef]
- Swier, L.J.Y.M.; Monjas, L.; Guskov, A.; de Voogd, A.R.; Erkens, G.B.; Slotboom, D.J.; Hirsch, A.K.H. Structure-Based Design of Potent Small-Molecule Binders to the S-Component of the ECF Transporter for Thiamine. ChemBioChem 2015, 16, 819–826. [Google Scholar] [CrossRef]
- Kim, H.J.; Li, Y.; Zimmermann, M.; Lee, Y.; Lim, H.W.; Leong Tan, A.S.; Choi, I.; Ko, Y.; Lee, S.; Seo, J.J.; et al. Pharmacological Perturbation of Thiamine Metabolism Sensitizes Pseudomonas Aeruginosa to Multiple Antibacterial Agents. Cell Chem. Biol. 2022, 29, 1317–1324.e5. [Google Scholar] [CrossRef]
- Rindi, G.; de Giuseppe, L.; Ventura, U. Distribution and Phosphorylation of Oxythiamine in Rat Tissues. J. Nutr. 1963, 81, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Tylicki, A.; Czerniecki, J.; Dobrzyn, P.; Matanowska, A.; Olechno, A.; Strumilo, S. Modification of Thiamine Pyrophosphate Dependent Enzyme Activity by Oxythiamine in Saccharomyces Cerevisiae Cells. Can. J. Microbiol. 2005, 51, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Ma, X.-D.; Chen, F.-E. Development of Two Scalable Syntheses of 4-Amino-5-Aminomethyl-2-Methylpyrimidine: Key Intermediate for Vitamin B1. Org. Process Res. Dev. 2012, 16, 57–60. [Google Scholar] [CrossRef]
- Yoshinaga, H.; Uruno, Y.; Nagata, H.; Hashimoto, M.; Kato, T. Cyclic Aminomethyl Pyrimidine Derivative 2016.
- Price, D.; May, E.L.; Pickel, F.D. Studies on Pyrimidines Related to Vitamin B1. I. A New Synthesis of 2-Methyl-6-Aminopyrimidine-5-Aldehyde. J. Am. Chem. Soc. 1940, 62, 2818–2820. [Google Scholar] [CrossRef]
- Plumb, J.A.; Milroy, R.; Kaye, S.B. Effects of the pH Dependence of 3-(4,5-Dimethylthiazol-2-Yl)-2,5-Diphenyltetrazolium Bromide-Formazan Absorption on Chemosensitivity Determined by a Novel Tetrazolium-Based Assay1. Cancer Res. 1989, 49, 4435–4440. [Google Scholar] [PubMed]
- Cui, J.; Wei, M.; Pang, L.; Gan, C.; Xiao, J.; Shi, H.; Zhan, J.; Liu, Z.; Huang, Y. Synthesis and Antiproliferative Evaluation of Novel Steroid-Benzisoselenazolone Hybrids. Steroids 2019, 152, 108502. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- The PyMOL Molecular Graphics System, Version 2.2.3; Copyright Schrodinger LLC: New York, NY, USA, 2021.
- Biovia, Discovery Studio. BIOVIA Discovery Studio Visualizer, Version 21.1.0; Dassault Systèmes: San Diego, CA, USA, 2022.
- Kato, M.; Max Wynn, R.; Chuang, J.L.; Tso, S.-C.; Machius, M.; Li, J.; Chuang, D.T. Structural Basis for Inactivation of the Human Pyruvate Dehydrogenase Complex by Phosphorylation: Role of Disordered Phosphorylation Loops. Structure 2008, 16, 1849–1859. [Google Scholar] [CrossRef]
- Mitschke, L.; Parthier, C.; Schröder-Tittmann, K.; Coy, J.; Lüdtke, S.; Tittmann, K. The Crystal Structure of Human Transketolase and New Insights into Its Mode of Action. J. Biol. Chem. 2010, 285, 31559–31570. [Google Scholar] [CrossRef] [PubMed]
- Erkens, G.B.; Berntsson, R.P.-A.; Fulyani, F.; Majsnerowska, M.; Vujičić-Žagar, A.; ter Beek, J.; Poolman, B.; Slotboom, D.J. The Structural Basis of Modularity in ECF-Type ABC Transporters. Nat. Struct. Mol. Biol. 2011, 18, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Suo, Y.; Wright, N.J.; Guterres, H.; Fedor, J.G.; Butay, K.J.; Borgnia, M.J.; Im, W.; Lee, S.-Y. Molecular Basis of Polyspecific Drug and Xenobiotic Recognition by OCT1 and OCT2. Nat. Struct. Mol. Biol. 2023, 30, 1001–1011. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Kissling, R.M.; Letellier, M.-A.; Nolan, S.P. Transesterification/Acylation of Secondary Alcohols Mediated by N-Heterocyclic Carbene Catalysts. J. Org. Chem. 2004, 69, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.C.; Hardy, D.J.; Maia, J.D.C.; Stone, J.E.; Ribeiro, J.V.; Bernardi, R.C.; Buch, R.; Fiorin, G.; Hénin, J.; Jiang, W.; et al. Scalable Molecular Dynamics on CPU and GPU Architectures with NAMD. J. Chem. Phys. 2020, 153, 044130. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual Molecular Dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A Web-Based Graphical User Interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef] [PubMed]
- Buck, M.; Bouguet-Bonnet, S.; Pastor, R.W.; MacKerell, A.D. Importance of the CMAP Correction to the CHARMM22 Protein Force Field: Dynamics of Hen Lysozyme. Biophys. J. 2006, 90, L36–L38. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, J.; Jo, S.; Brooks III, C.L.; Lee, H.S.; Im, W. CHARMM-GUI Ligand Reader and Modeler for CHARMM Force Field Generation of Small Molecules. J. Comput. Chem. 2017, 38, 1879–1886. [Google Scholar] [CrossRef]
- Maia, J.A.; Amaral, L.A.N.; Scheurer, R.M. Improving the Efficiency and Equilibration of Molecular Dynamics Simulations: Coupling Gaussian Accelerated Molecular Dynamics with a High-Energy Collective Variable Replica Exchange. J. Chem. Theory Comput. 2020, 16, 1264. [Google Scholar]
- Milewska, S.; Siemiaszko, G.; Wilczewska, A.Z.; Misztalewska-Turkowicz, I.; Markiewicz, K.H.; Szymczuk, D.; Sawicka, D.; Car, H.; Lazny, R.; Niemirowicz-Laskowska, K. Folic-Acid-Conjugated Thermoresponsive Polymeric Particles for Targeted Delivery of 5-Fluorouracil to CRC Cells. Int. J. Mol. Sci. 2023, 24, 1364. [Google Scholar] [CrossRef]
- Fan, Y.; Marioli, M.; Zhang, K. Analytical Characterization of Liposomes and Other Lipid Nanoparticles for Drug Delivery. J. Pharm. Biomed. Anal. 2021, 192, 113642. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line/Compound | GI50 (µM) | IC50 (µM) | SI |
|---|---|---|---|
| fibroblasts/OT | >1500 | >1500 | OT 150 ± 20.83 |
| HeLa cells/OT | 39 * ± 6.50 | 51 * ± 2.89 | |
| fibroblasts/MT | >1500 | >1500 | MT 186 ± 35.48 |
| HeLa cells/MT | 83 * ± 14.50 | 112 * ± 20.36 | |
| fibroblasts/T | >1500 | >1500 | (-) |
| HeLa cells/T | >1500 | >1500 |
| Enzyme | Number and Percent of Matches Per Docking Position for a Particular Ligand | |||||||
|---|---|---|---|---|---|---|---|---|
| T-PP or T | OT-PP or OT | MT-PP or MT | ||||||
| Position 1 | Position 2 | Position 3 | Position 1 | Position 2 | Position 1 | Position 2 | Position 3 | |
| Transketolase | 120 | 47 | (-) | 119 | 62 | 129 | 51 | (-) |
| 71.9% | 28.1% | 65.7% | 34.3% | 71.7% | 28.3% | |||
| Pyruvate dehydrogenase | 122 | 1 | (-) | 113 | 1 | 116 | 1 | (-) |
| 99.2% | 0.8% | 99.1% | 0.9% | 99.1% | 0.9% | |||
| Thiamine pyrophosphokinase | 117 | 29 | 2 | 123 | 10 | 123 | 24 | 1 |
| 79.1% | 19.6% | 1.3% | 92.5% | 7.5% | 83.1% | 16.2% | 0.7% | |
| Enzyme 1 | Average ΔG for Particular Ligand in Addition to Docking Position (kcal/mol) | Minimum ΔG Including Position (kcal/mol) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| T-PP or T | OT-PP or OT | MT-PP or MT | |||||||||
| Position 1 | Position 2 | Position 3 | Position 1 | Position 2 | Position 1 | Position 2 | Position 3 | T-PP or T | OT-PP or OT | MT-PP or MT | |
| TK 3 | −5.5 (−6.2/25.0) | −5.5 (−5.9/32.4) | (-) | −5.6 (−6.2/24.3) | −5.6 (−6.1/41.7) | −5.3 6 (−6.0/29.2) | −5.5 (−5.8/44.8) | (-) | −6.2 Pos. 1 | −6.2 Pos. 1 | −6.1 Pos. 1 |
| PDH 4 | −9.1 (±0.25) | −7.5 2 | (-) | −9.1 (±0.23) | −7.7 2 | −8.3 7 (±0.19) | −7.8 2 | (-) | −9.8 Pos. 1 | −9.5 Pos. 1 | −9.2 Pos. 1 |
| TPP 5 | −7.0 (−7.5/−6.7) | −7.0 (−7.4/−6.8) | −7.0 (−7.2/−6.8) | −7.1 (−7.4/−6.7) | −7.0 (−7.2/−6.8) | −7.1 8 (−8.2/−6.9) | −7.0 (−8.1/−6.9) | −6.8 2 | −7.5 Pos. 1 | −7.0 Pos. 1 | −8.2 Pos. 1 |
| Thiamine Transporter | Number and Percentage of Matches per Docking Position for Particular Thiamine Transporter | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| T | OT | MT | |||||||||
| Position 1 | Position 2 | Position 3 | Position 4 | Position 1 | Position 2 | Position 3 | Position 4 | Position 1 | Position 2 | Position 3 | |
| ThiT S-component | 10 (4.1%) | 189 (77.8%) | 44 (18.1%) | (-) | 113 (46.7%) | 129 (53.3%) | (-) | (-) | 129 (100%) | (-) | (-) |
| OTC1 | 47 (13.9%) | 245 (72.3%) | 37 (10.9%) | 10 (2.9%) | 212 (61.8%) | 100 (29.2%) | 24 (7.0%) | 7 (2.0%) | 2 (0.6%) | 99 (29.3%) | 237 (70.1%) |
| Transporter | Average ΔG for Particular Ligand in Addition to Docking Position (kcal/mol) 1 | Minimum ΔG Including Position (kcal/mol) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T | OT | MT | ||||||||||||
| Position 1 | Position 2 | Position 3 | Position 4 | Position 1 | Position 2 | Position 3 | Position 4 | Position 1 | Position 2 | Position 3 | T | OT | MT | |
| ThiT 2 | −6.9 (−7.4/−6.9) | −7.8 (−8.1/−7.4) | −7.3 (−7.4/−7.1) | (-) | −7.6 (−8.1/−6.9) | −7.9 (−8.2/−7.3) | (-) | (-) | −7.0 4 (−7.6/−6.6) | (-) | (-) | −8.1 Pos.2 | −8.2 Pos. 2 | −7.6 Pos. 1 |
| OCT1 3 | −6.3 (−6.3/−6.2) | −6.5 (−6.6/−6.3) | −6.5 (−6.5/−6.4) | −6.3 (−6.4/−6.3) | −6.5 (−6.6/−6.4) | −6.5 (−6.5/−6.4) | −6.2 (−6.3/−6.2) | −6.3 (−6.3/−6.3) | −6.8 (−6.8/−6.8) | −6.8 (−6.9/−6.7) | −7.1 5 (−7.1/−7.0) | −6.6 Pos. 2 | −6.6 Pos. 2 | −7.1 Pos. 3 |
| Residue, Type (D/A) | Occupancy % | ||
|---|---|---|---|
| Thiamine | Oxythiamine | 2′-Methylthiamine | |
| SER 470 (D) | 39.2 | 2.3 | 62 |
| SER 470 (A) | 0.2 | ||
| TYR 36 (A) | 11 | 2.4 | |
| GLN 241 (A) | 2.3 | 0.3 | 0.1 |
| GLN 447 (D) | 4.9 | 0.2 | 0.4 |
| THR 443 (D) | 1.5 | ||
| THR 443 (D) | 0.8 | ||
| GLU 386(A) | 6.1 | 0.4 | |
| THR 245 (A) | 10.3 | ||
| LYS 124 (D) | 5.8 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malinowska, M.; Czerniecka, M.; Jastrzebska, I.; Ratkiewicz, A.; Tylicki, A.; Wawrusiewicz-Kurylonek, N. In Vitro and In Silico Studies on Cytotoxic Properties of Oxythiamine and 2′-Methylthiamine. Int. J. Mol. Sci. 2024, 25, 4359. https://doi.org/10.3390/ijms25084359
Malinowska M, Czerniecka M, Jastrzebska I, Ratkiewicz A, Tylicki A, Wawrusiewicz-Kurylonek N. In Vitro and In Silico Studies on Cytotoxic Properties of Oxythiamine and 2′-Methylthiamine. International Journal of Molecular Sciences. 2024; 25(8):4359. https://doi.org/10.3390/ijms25084359
Chicago/Turabian StyleMalinowska, Marta, Magdalena Czerniecka, Izabella Jastrzebska, Artur Ratkiewicz, Adam Tylicki, and Natalia Wawrusiewicz-Kurylonek. 2024. "In Vitro and In Silico Studies on Cytotoxic Properties of Oxythiamine and 2′-Methylthiamine" International Journal of Molecular Sciences 25, no. 8: 4359. https://doi.org/10.3390/ijms25084359
APA StyleMalinowska, M., Czerniecka, M., Jastrzebska, I., Ratkiewicz, A., Tylicki, A., & Wawrusiewicz-Kurylonek, N. (2024). In Vitro and In Silico Studies on Cytotoxic Properties of Oxythiamine and 2′-Methylthiamine. International Journal of Molecular Sciences, 25(8), 4359. https://doi.org/10.3390/ijms25084359