
Upregulation of EMR1 (ADGRE1) by Tumor-Associated Macrophages Promotes Colon Cancer Progression by Activating the JAK2/STAT1,3 Signaling Pathway in Tumor Cells

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
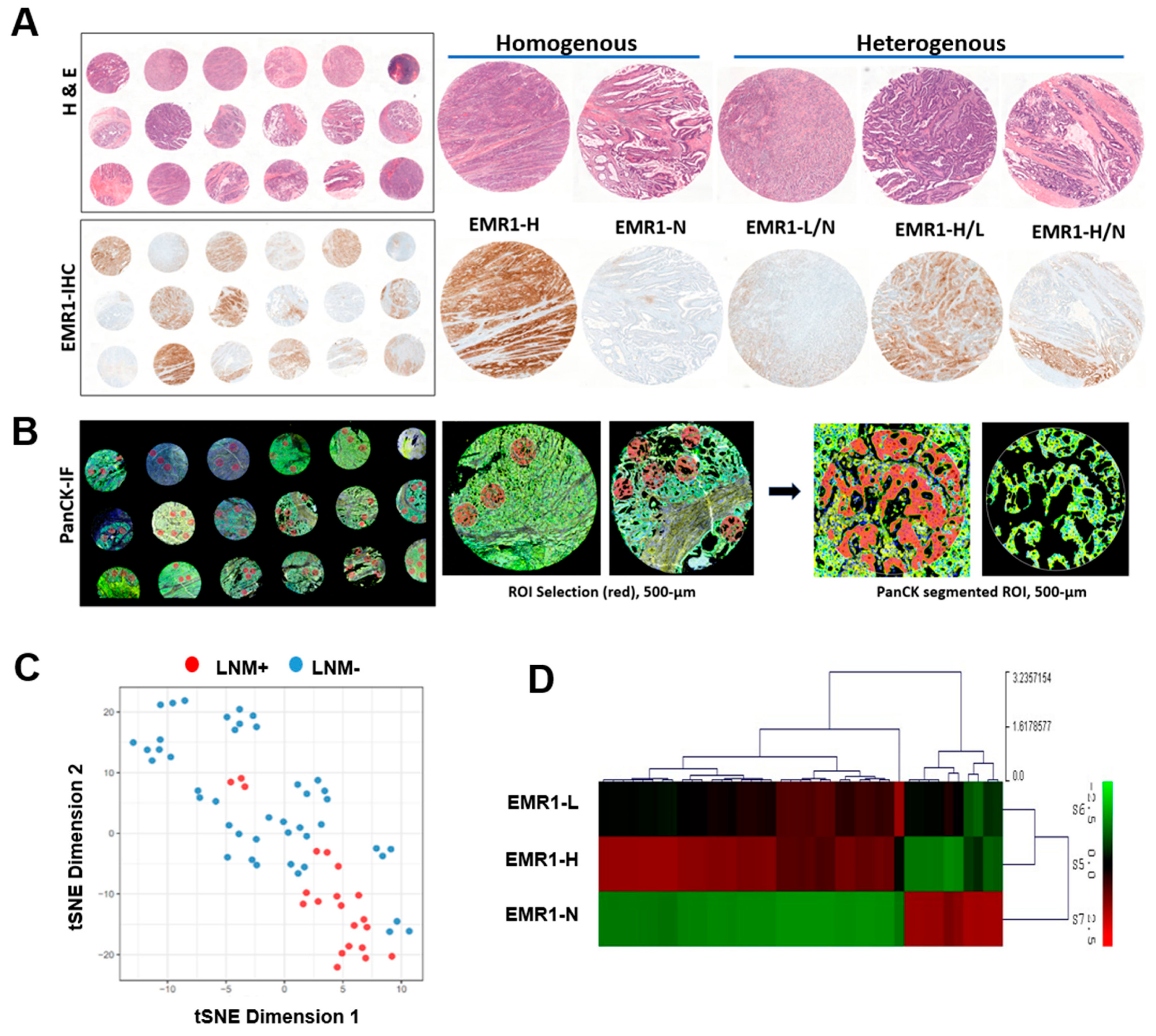
2.1. Whole Transcriptomic Profiling by GeoMx
2.1.1. Data Reduction for Comparative Spatial Transcriptomic Profiling of CC
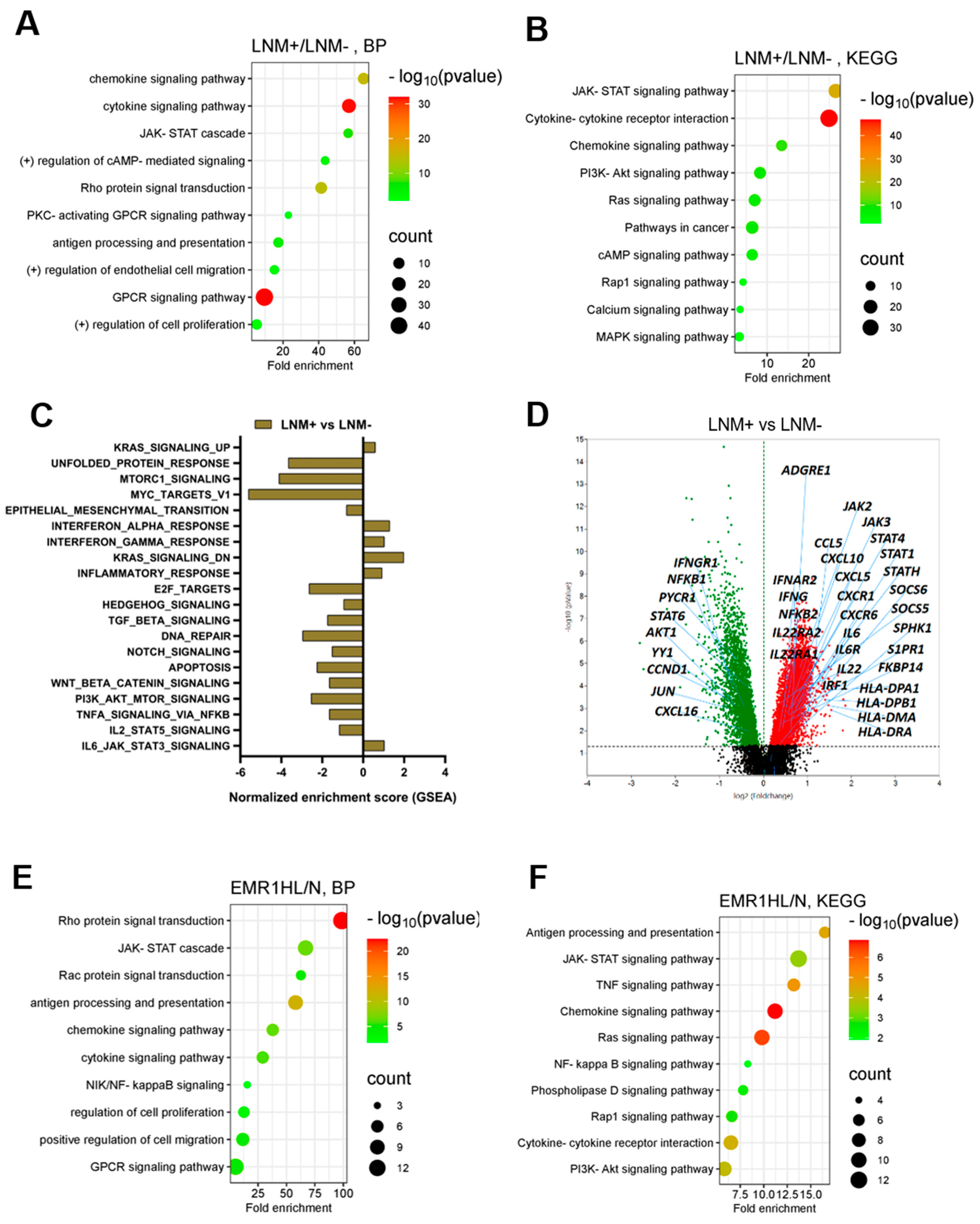
2.1.2. Comparison of Differential Gene Expression in CC According to LNM Using Spatial Transcriptomic Profiling
2.1.3. Comparison of DEGs in CC According to EMR1 Expression Using Spatial Transcriptomic Profiling
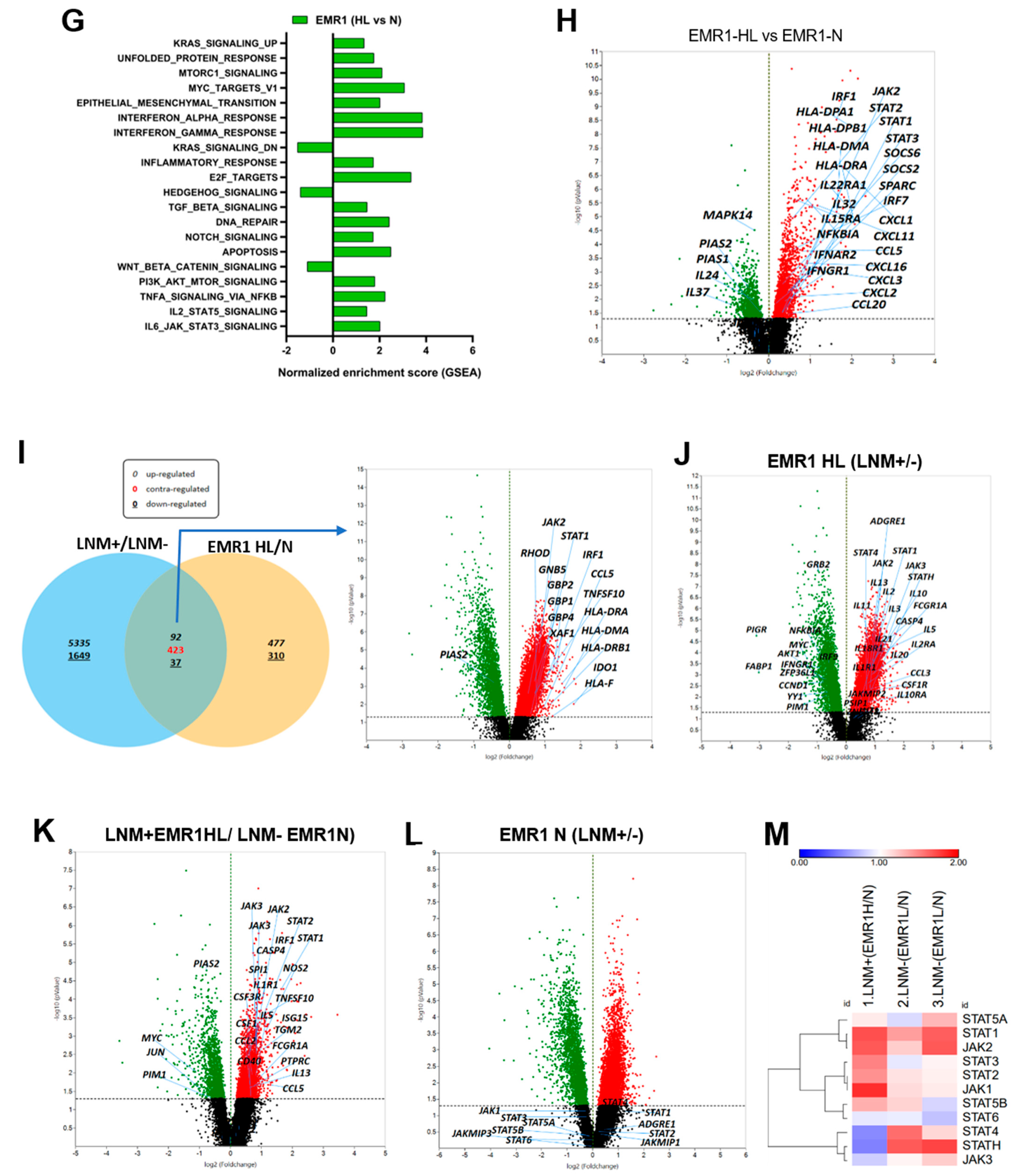
2.1.4. DEGs Involved in LNM and EMR1 Expression in CC
2.2. In Vitro Analysis of CC Cells Based on EMR1 Expression Using a Coculture System with Macrophages
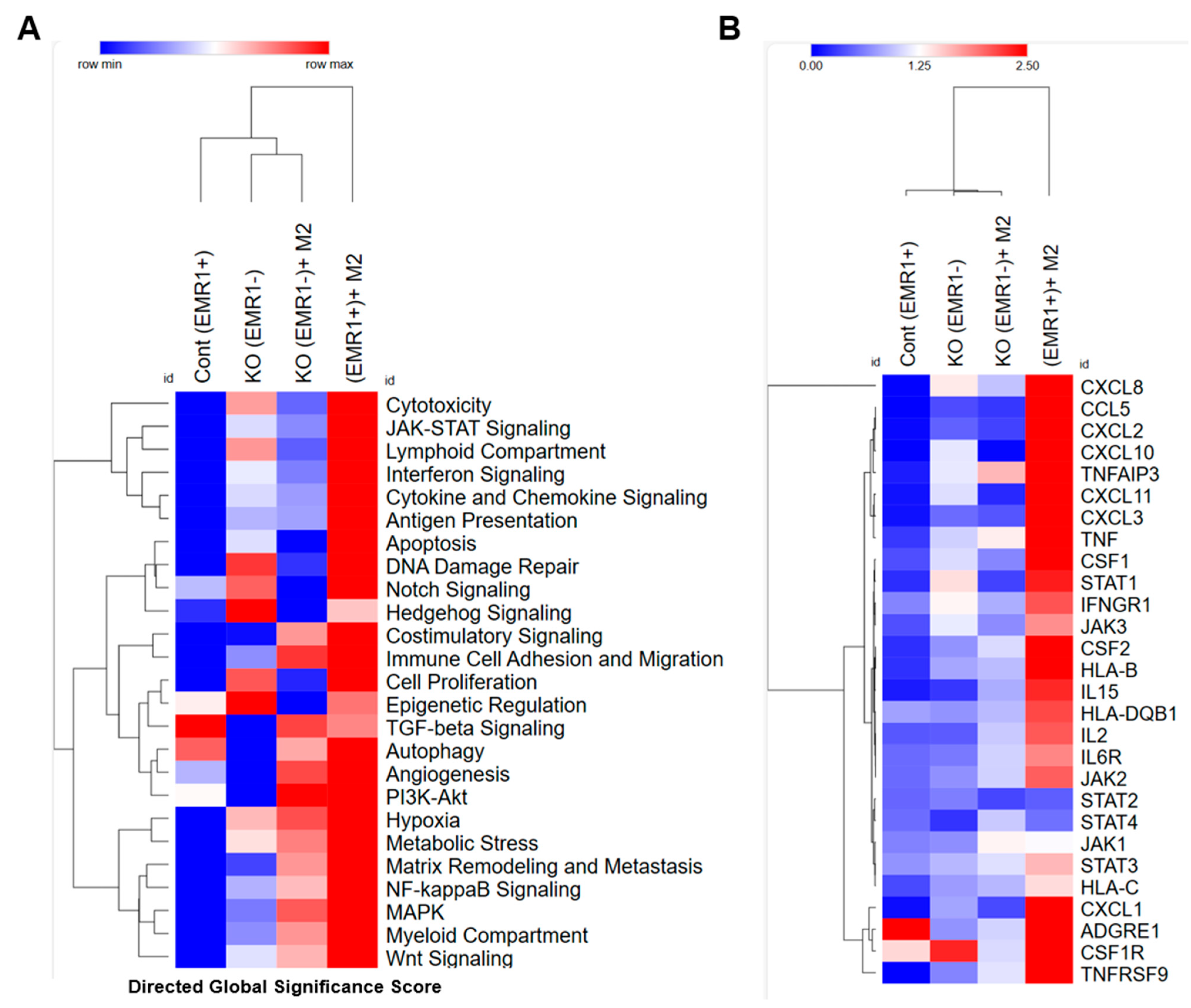
2.2.1. Molecular Changes between EMR1+ or EMR1KO CC Cells Cocultured with Macrophages In Vitro Based on the nCounter System
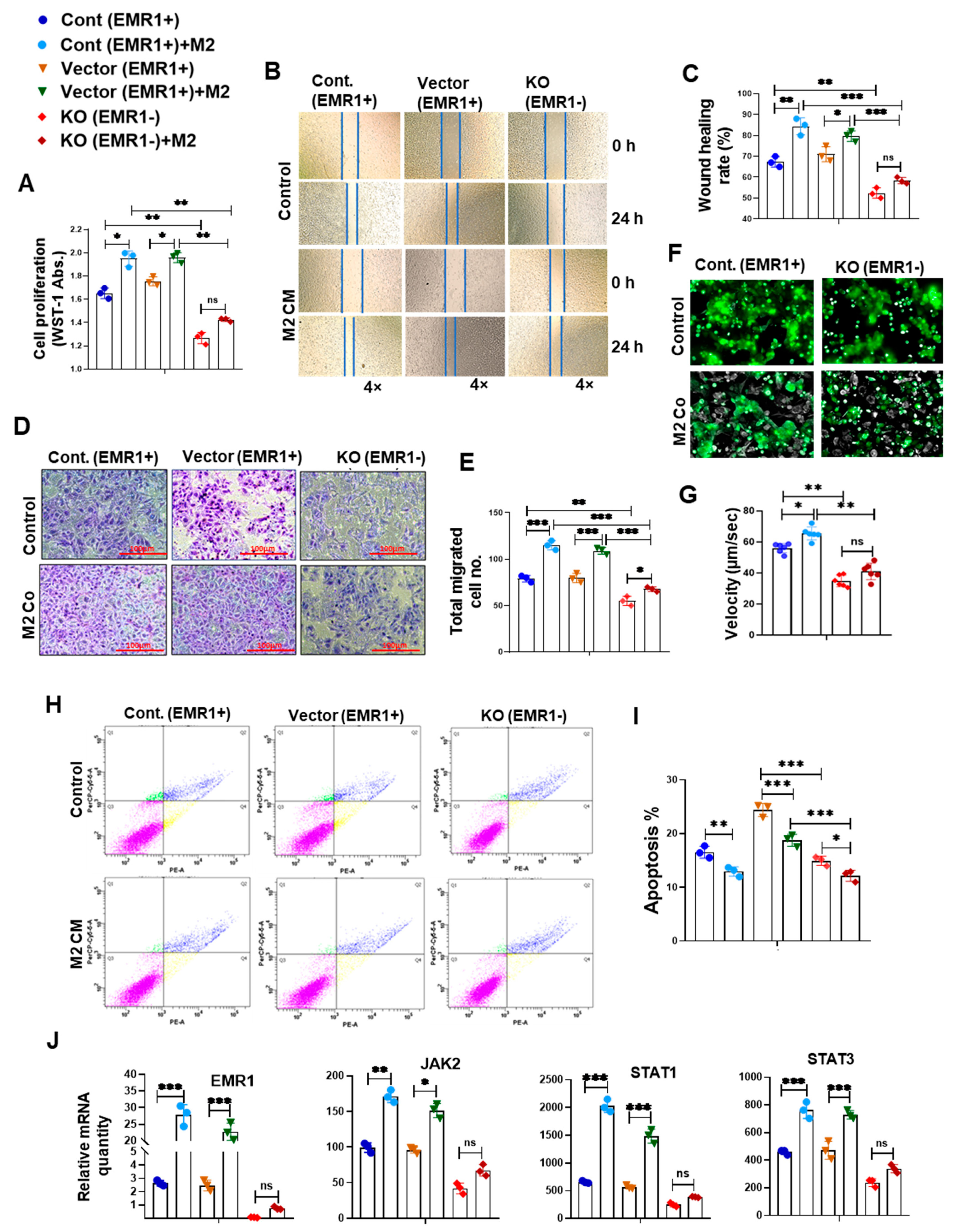
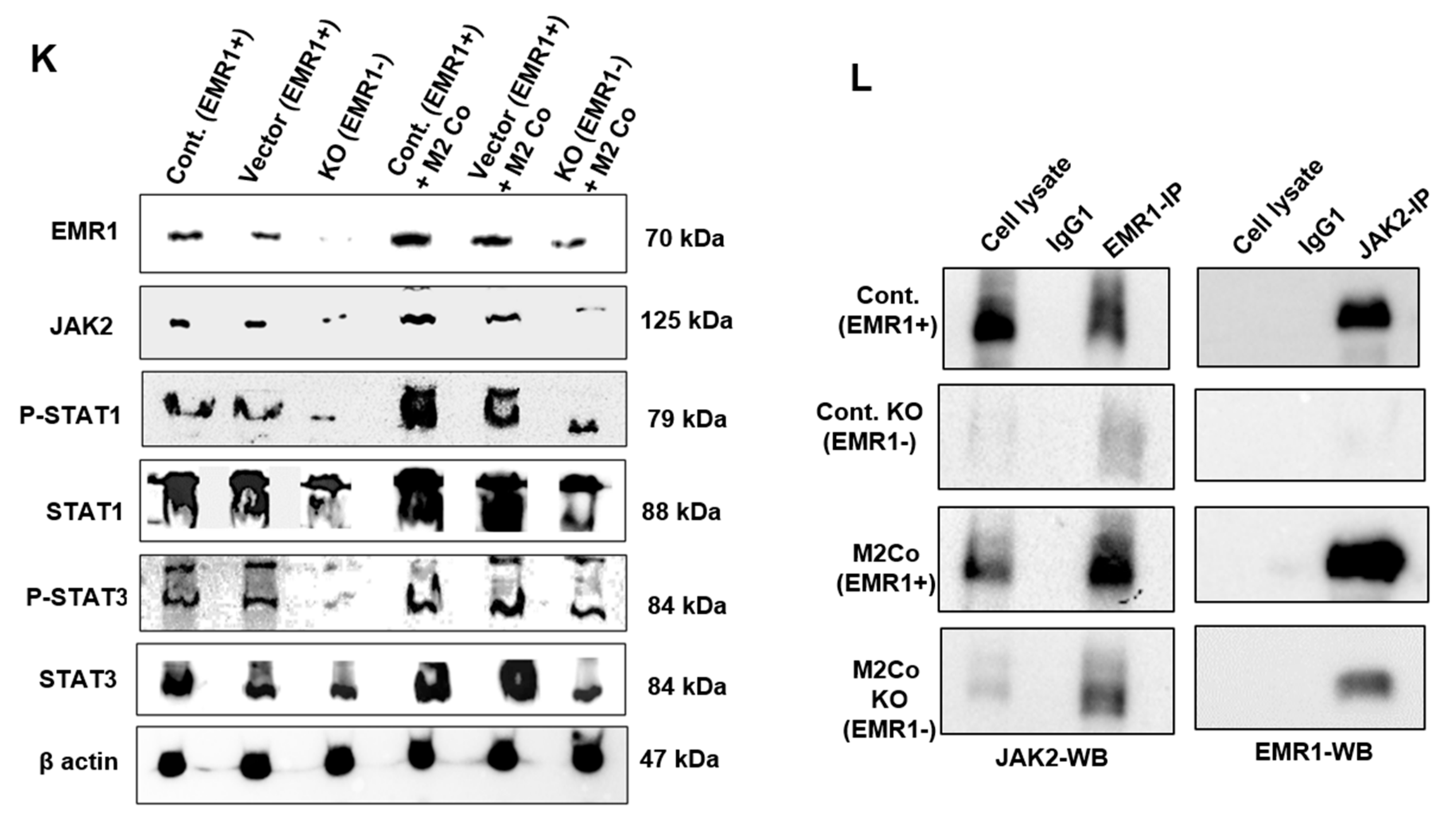
2.2.2. Biological Behavior and Molecular Changes of CC Cells Due to EMR1 Expression in a Coculture System with Macrophages
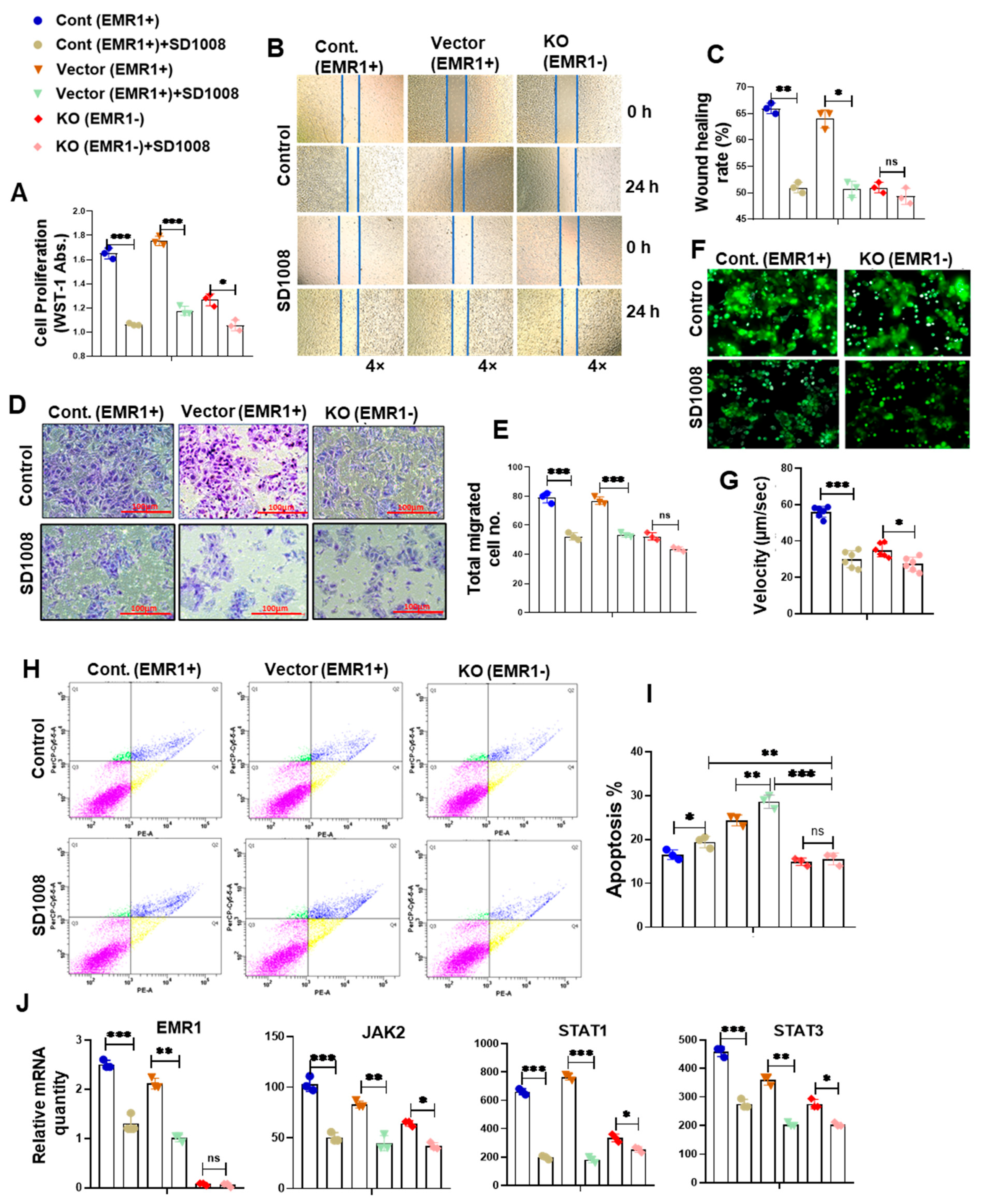
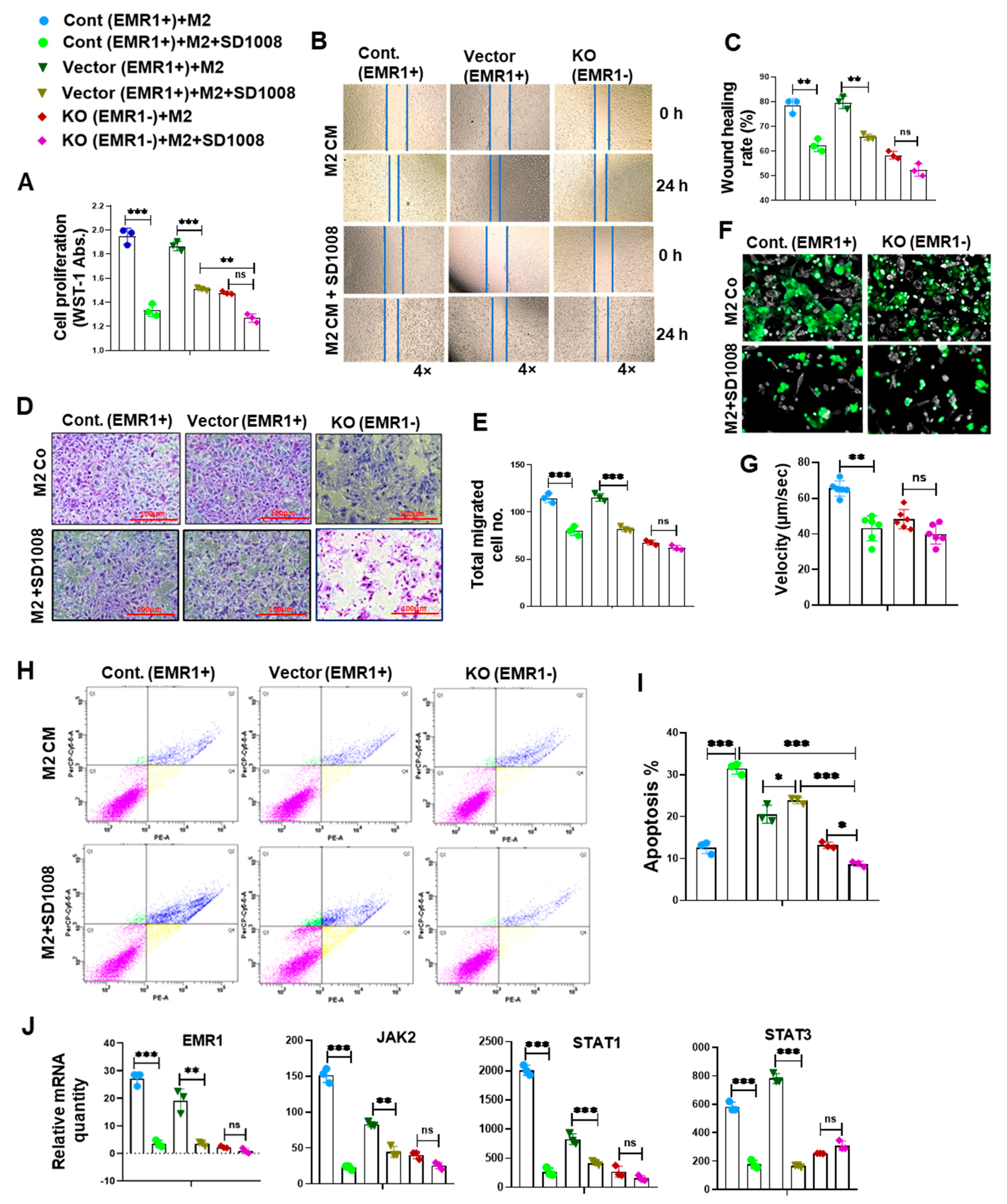
2.2.3. Effect of JAK2/STAT3 Pathway Inhibitor (SD1008) on the Molecular and Biologic Characteristics of CC Cells According to EMR1 Expression
3. Discussion
4. Materials and Methods
4.1. Ethics Approval
4.2. Whole Transcriptome Analysis Using GeoMx Digital Spatial Profiling (DSP)
4.2.1. Tissue Sample Preparation and Procedures for GeoMx DSP
4.2.2. Dimension-Reduction Using t-Distributed Stochastic Neighbor Embedding (t-SNE)
4.2.3. Functional Enrichment Analysis
4.3. In Vitro Validation of EMR1-Related Differential Gene Expression in CC Cells Using a Coculture System with Macrophages
4.3.1. Cell Culture and Reagents
4.3.2. Generation of EMR1KO HCT15 CC Cells with the CRISPR-Cas9 Gene Editing System, and Validation via Western Blotting and Flow Cytometry
4.3.3. Polarization of Macrophages
4.3.4. Coculture Procedures
4.3.5. Exploring EMR1-Related RNA Expression in CC Cells after Coculture with Macrophages Using an nCounter System
4.3.6. Exploring EMR1-Related Protein Expression after Coculture with Macrophages Using Western Blotting
4.3.7. Exploring EMR1-Related mRNA Expression after Coculture with Macrophages Using Real-Time Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
4.3.8. Exploring EMR1-Related Protein-Protein Interaction after Coculture with Macrophages Using Co-Immunoprecipitation (Co-IP)
4.4. Evaluation of the Biological Changes of Cancer Cells Due to EMR1 Expression after Coculture with Macrophages
4.4.1. Cell Proliferation Assay
4.4.2. Transwell Migration Assay
4.4.3. Wound Healing Assay
4.4.4. Cell Motility Assay
4.4.5. Apoptosis Assay
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ranasinghe, R.; Mathai, M.; Zulli, A. A synopsis of modern-day colorectal cancer: Where we stand. Biochim. Biophys. Acta Rev. Cancer 2022, 1877, 188699. [Google Scholar] [CrossRef] [PubMed]
- Currais, P.; Rosa, I.; Claro, I. Colorectal cancer carcinogenesis: From bench to bedside. World J. Gastrointest. Oncol. 2022, 14, 654–663. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Yang, M. Molecular network of colorectal cancer and current therapeutic options. Front. Oncol. 2022, 12, 852927. [Google Scholar] [CrossRef] [PubMed]
- Zafari, N.; Khosravi, F.; Rezaee, Z.; Esfandyari, S.; Bahiraei, M.; Bahramy, A.; Ferns, G.A.; Avan, A. The role of the tumor microenvironment in colorectal cancer and the potential therapeutic approaches. J. Clin. Lab. Anal. 2022, 36, e24585. [Google Scholar] [CrossRef] [PubMed]
- Baghban, R.; Roshangar, L.; Jahanban-Esfahlan, R.; Seidi, K.; Ebrahimi-Kalan, A.; Jaymand, M.; Kolahian, S.; Javaheri, T.; Zare, P. Tumor microenvironment complexity and therapeutic implications at a glance. Cell Commun. Signal. 2020, 18, 59. [Google Scholar] [CrossRef] [PubMed]
- Akter, R.; Kim, K.; Kwon, H.Y.; Kim, Y.; Eom, Y.W.; Cho, H.-M.; Cho, M.-Y. EMR1/ADGRE1 Expression in Cancer Cells Upregulated by Tumor-Associated Macrophages Is Related to Poor Prognosis in Colorectal Cancer. Biomedicines 2022, 10, 3121. [Google Scholar] [CrossRef] [PubMed]
- Gad, A.A.; Balenga, N. The emerging role of adhesion GPCRs in cancer. ACS Pharmacol. Transl. Sci. 2020, 3, 29–42. [Google Scholar] [CrossRef]
- Hamann, J.; Koning, N.; Pouwels, W.; Ulfman, L.H.; van Eijk, M.; Stacey, M.; Lin, H.H.; Gordon, S.; Kwakkenbos, M.J. EMR1, the human homolog of F4/80, is an eosinophil-specific receptor. Eur. J. Immunol. 2007, 37, 2797–2802. [Google Scholar] [CrossRef]
- Hall, R.A.; Lala, T. Adhesion G Protein-Coupled Receptors: Structure, Signaling, Physiology and Pathophysiology. Physiol. Rev. 2022, 102, 1587–1624. [Google Scholar]
- Lin, H.-H.; Hsiao, C.-C.; Pabst, C.; Hébert, J.; Schöneberg, T.; Hamann, J. Adhesion GPCRs in regulating immune responses and inflammation. Adv. Immunol. 2017, 136, 163–201. [Google Scholar] [PubMed]
- Aust, G.; Zhu, D.; Van Meir, E.G.; Xu, L. Adhesion GPCRs in tumorigenesis. In Adhesion G Protein-Coupled Receptors: Molecular, Physiological and Pharmacological Principles in Health and Disease; Springer: Berlin/Heidelberg, Germany, 2016; pp. 369–396. [Google Scholar]
- Steinert, M.; Wobus, M.; Boltze, C.; Schütz, A.; Wahlbuhl, M.; Hamann, J.; Aust, G. Expression and regulation of CD97 in colorectal carcinoma cell lines and tumor tissues. Am. J. Pathol. 2002, 161, 1657–1667. [Google Scholar] [CrossRef] [PubMed]
- Wobus, M.; Huber, O.; Hamann, J.; Aust, G. CD97 overexpression in tumor cells at the invasion front in colorectal cancer (CC) is independently regulated of the canonical Wnt pathway. Mol. Carcinog. 2006, 45, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Hilbig, D.; Dietrich, N.; Wandel, E.; Gonsior, S.; Sittig, D.; Hamann, J.; Aust, G. The interaction of CD97/ADGRE5 With β-Catenin in adherens junctions Is lost during colorectal carcinogenesis. Front. Oncol. 2018, 8, 182. [Google Scholar] [CrossRef]
- Ji, B.; Feng, Y.; Sun, Y.; Ji, D.; Qian, W.; Zhang, Z.; Wang, Q.; Zhang, Y.; Zhang, C.; Sun, Y. GPR56 promotes proliferation of colorectal cancer cells and enhances metastasis via epithelial-mesenchymal transition through PI3K/AKT signaling activation. Oncol. Rep. 2018, 40, 1885–1896. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, Z.; Chen, J.; Xue, M.; Pan, X. ELTD1 promotes invasion and metastasis by activating MMP2 in colorectal cancer. Int. J. Biol. Sci. 2021, 17, 3048. [Google Scholar] [CrossRef]
- Waddell, L.A.; Lefevre, L.; Bush, S.J.; Raper, A.; Young, R.; Lisowski, Z.M.; McCulloch, M.E.B.; Muriuki, C.; Sauter, K.A.; Clark, E.L.; et al. ADGRE1 (EMR1, F4/80) Is a Rapidly-Evolving Gene Expressed in Mammalian Monocyte-Macrophages. Front. Immunol. 2018, 9, 2246. [Google Scholar] [CrossRef] [PubMed]
- Legrand, F.; Tomasevic, N.; Simakova, O.; Lee, C.-C.R.; Wang, Z.; Raffeld, M.; Makiya, M.A.; Palath, V.; Leung, J.; Baer, M. The eosinophil surface receptor epidermal growth factor–like module containing mucin-like hormone receptor 1 (EMR1): A novel therapeutic target for eosinophilic disorders. J. Allergy Clin. Immunol. 2014, 133, 1439–1447.e1438. [Google Scholar] [CrossRef] [PubMed]
- Marx, V. Method of the Year: Spatially resolved transcriptomics. Nat. Methods 2021, 18, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Merritt, C.R.; Ong, G.T.; Church, S.E.; Barker, K.; Danaher, P.; Geiss, G.; Hoang, M.; Jung, J.; Liang, Y.; McKay-Fleisch, J. Multiplex digital spatial profiling of proteins and RNA in fixed tissue. Nat. Biotechnol. 2020, 38, 586–599. [Google Scholar] [CrossRef] [PubMed]
- Zollinger, D.R.; Lingle, S.E.; Sorg, K.; Beechem, J.M.; Merritt, C.R. GeoMx™ RNA assay: High multiplex, digital, spatial analysis of RNA in FFPE tissue. In Situ Hybrid. Protoc. 2020, 2148, 331–345. [Google Scholar]
- Brouwer, N.P.; Webbink, L.; Haddad, T.S.; Rutgers, N.; van Vliet, S.; Wood, C.S.; Jansen, P.W.; Lafarge, M.W.; de Wilt, J.H.; imCMS Consortium. Transcriptomics and proteomics reveal distinct biology for lymph node metastases and tumour deposits in colorectal cancer. J. Pathol. 2023, 261, 410–412. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.J.; Zavras, J.P.; Veziroglu, E.M.; Nasir-Moin, M.; Kolling, F.W.; Christensen, B.C.; Salas, L.A.; Barney, R.E.; Palisoul, S.M.; Ren, B. Identification of Spatial Proteomic Signatures of Colon Tumor Metastasis: A Digital Spatial Profiling Approach. Am. Pathol. 2023, 193, 778–795. [Google Scholar] [CrossRef] [PubMed]
- Rah, B.; Rather, R.A.; Bhat, G.R.; Baba, A.B.; Mushtaq, I.; Farooq, M.; Yousuf, T.; Dar, S.B.; Parveen, S.; Hassan, R. JAK/STAT signaling: Molecular targets, therapeutic opportunities, and limitations of targeted inhibitions in solid malignancies. Front. Pharmacol. 2022, 13, 821344. [Google Scholar] [CrossRef] [PubMed]
- Erdogan, F.; Radu, T.B.; Orlova, A.; Qadree, A.K.; de Araujo, E.D.; Israelian, J.; Valent, P.; Mustjoki, S.M.; Herling, M.; Moriggl, R. JAK-STAT core cancer pathway: An integrative cancer interactome analysis. J. Cell. Mol. Med. 2022, 26, 2049–2062. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Yuan, X.; Song, J.; Chen, Y.; Tan, X.; Li, Q. Association analyses of the JAK/STAT signaling pathway with the progression and prognosis of colon cancer. Oncol. Lett. 2019, 17, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Low, J.T.; Silke, J.; O’Reilly, L.A. Digesting the role of JAK-STAT and cytokine signaling in oral and gastric cancers. Front. Immunol. 2022, 13, 835997. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.A.; Nisar, S.; Singh, M.; Ashraf, B.; Masoodi, T.; Prasad, C.P.; Sharma, A.; Maacha, S.; Karedath, T.; Hashem, S. Cytokine-and chemokine-induced inflammatory colorectal tumor microenvironment: Emerging avenue for targeted therapy. Cancer Commun. 2022, 42, 689–715. [Google Scholar] [CrossRef] [PubMed]
- Erkasap, N.; Özyurt, R.; Özkurt, M.; Yaşar, F.; Erkasap, S.; Ihtiyar, E. The role of JAK/STAT signaling pathway and TNF-α crosstalk in human colorectal cancer. Gene Rep. 2016, 3, 1–4. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, F.; Li, G.; Li, G.; Yang, X.; Liu, L.; Zhang, R.; Zhang, B.; Feng, Y. Human colorectal cancer-derived mesenchymal stem cells promote colorectal cancer progression through IL-6/JAK2/STAT3 signaling. Cell Death Dis. 2018, 9, 25. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.-H.; Chen, K.-H.; Tsai, J.-H.; Cheng, Y.-M.; Lee, C.-C.; Kao, C.-H.; Chan, K.-Y.; Chen, Y.-T.; Hsu, W.-L.; Yeh, K.-H. Proteasome inhibitors restore the STAT1 pathway and enhance the expression of MHC class I on human colon cancer cells. J. Biomed. Sci. 2021, 28, 75. [Google Scholar] [CrossRef] [PubMed]
- Tarafdar, A.; Hopcroft, L.E.; Gallipoli, P.; Pellicano, F.; Cassels, J.; Hair, A.; Korfi, K.; Jørgensen, H.G.; Vetrie, D.; Holyoake, T.L. CML cells actively evade host immune surveillance through cytokine-mediated downregulation of MHC-II expression. Blood J. Am. Soc. Hematol. 2017, 129, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Stickel, N.; Hanke, K.; Marschner, D.; Prinz, G.; Köhler, M.; Melchinger, W.; Pfeifer, D.; Schmitt-Graeff, A.; Brummer, T.; Heine, A. MicroRNA-146a reduces MHC-II expression via targeting JAK/STAT signaling in dendritic cells after stem cell transplantation. Leukemia 2017, 31, 2732–2741. [Google Scholar] [CrossRef]
- Lei, P.-J.; Pereira, E.R.; Andersson, P.; Amoozgar, Z.; Van Wijnbergen, J.W.; O’Melia, M.J.; Zhou, H.; Chatterjee, S.; Ho, W.W.; Posada, J.M. Cancer cell plasticity and MHC-II-mediated immune tolerance promote breast cancer metastasis to lymph nodes. J. Exp. Med. 2023, 220, e20221847. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Song, H.; Yin, H.; Jiang, T.; Xu, Y.; Liu, L.; Wang, H.; Gao, H.; Wang, R. The circular RNA circSPARC enhances the migration and proliferation of colorectal cancer by regulating the JAK/STAT pathway. Mol. Cancer 2021, 20, 81. [Google Scholar] [CrossRef]
- Long, J.; Yao, Z.; Sui, Y.; Fang, S. SphK1 promotes cancer progression through activating JAK/STAT pathway and up-regulating S1PR1 expression in colon cancer cells. Anti-Cancer Agents Med. Chem. 2022, 22, 254–260. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, R.; Yang, J.; Bi, T.; Zhou, S. FKBP14 promotes the proliferation and migration of colon carcinoma cells through targeting IL-6/STAT3 signaling pathway. OncoTargets Ther. 2019, 12, 9069. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Zhang, Z.-G.; Tian, X.-Q.; Sun, D.-F.; Liang, Q.-C.; Zhang, Y.-J.; Lu, R.; Chen, Y.-X.; Fang, J.-Y. Inhibition of JAK1, 2/STAT3 signaling induces apoptosis, cell cycle arrest, and reduces tumor cell invasion in colorectal cancer cells. Neoplasia 2008, 10, 287–297. [Google Scholar] [CrossRef]
- Du, W.; Hong, J.; Wang, Y.C.; Zhang, Y.J.; Wang, P.; Su, W.Y.; Lin, Y.W.; Lu, R.; Zou, W.P.; Xiong, H. Inhibition of JAK2/STAT3 signalling induces colorectal cancer cell apoptosis via mitochondrial pathway. J. Cell. Mol. Med. 2012, 16, 1878–1888. [Google Scholar] [CrossRef]
- Li, W.; Lee, M.-R.; Choi, E.; Cho, M.-Y. Clinicopathologic significance of survivin expression in relation to CD133 expression in surgically resected stage II or III colorectal cancer. J. Pathol. Transl. Med. 2017, 51, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Eom, Y.W.; Akter, R.; Li, W.; Lee, S.; Hwang, S.; Kim, J.; Cho, M.-Y. M1 macrophages promote trail expression in adipose tissue-derived stem cells, which suppresses colitis-associated colon cancer by increasing apoptosis of cd133+ cancer stem cells and decreasing m2 macrophage population. Int. J. Mol. Sci. 2020, 21, 3887. [Google Scholar] [CrossRef]
- Li, W.L.; Lee, M.-R.; Cho, M.-Y. The small molecule survivin inhibitor YM155 may be an effective treatment modality for colon cancer through increasing apoptosis. Biochem. Biophys. Res. Commun. 2016, 471, 309–314. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | LNM+ | LNM− | ||
| Age | ≤50:>50 | 0:4 | 1:9 | |
| Sex | Male:Female | 1:3 | 5:5 | |
| Location of CC | Right:Left | 4:0 | 9:1 | |
| EMR1 | High:Negative | 1:0 | 2:3 | |
| Heterogeneous | L/N | 0 | 2 | |
| H/L | 2 | 3 | ||
| H/N | 1 | 0 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akter, R.; Park, R.; Lee, S.K.; Han, E.j.; Park, K.-S.; Park, J.; Cho, M.-Y. Upregulation of EMR1 (ADGRE1) by Tumor-Associated Macrophages Promotes Colon Cancer Progression by Activating the JAK2/STAT1,3 Signaling Pathway in Tumor Cells. Int. J. Mol. Sci. 2024, 25, 4388. https://doi.org/10.3390/ijms25084388
Akter R, Park R, Lee SK, Han Ej, Park K-S, Park J, Cho M-Y. Upregulation of EMR1 (ADGRE1) by Tumor-Associated Macrophages Promotes Colon Cancer Progression by Activating the JAK2/STAT1,3 Signaling Pathway in Tumor Cells. International Journal of Molecular Sciences. 2024; 25(8):4388. https://doi.org/10.3390/ijms25084388
Chicago/Turabian StyleAkter, Rokeya, Rackhyun Park, Soo Kyung Lee, Eun ju Han, Kyu-Sang Park, Junsoo Park, and Mee-Yon Cho. 2024. "Upregulation of EMR1 (ADGRE1) by Tumor-Associated Macrophages Promotes Colon Cancer Progression by Activating the JAK2/STAT1,3 Signaling Pathway in Tumor Cells" International Journal of Molecular Sciences 25, no. 8: 4388. https://doi.org/10.3390/ijms25084388
APA StyleAkter, R., Park, R., Lee, S. K., Han, E. j., Park, K. -S., Park, J., & Cho, M. -Y. (2024). Upregulation of EMR1 (ADGRE1) by Tumor-Associated Macrophages Promotes Colon Cancer Progression by Activating the JAK2/STAT1,3 Signaling Pathway in Tumor Cells. International Journal of Molecular Sciences, 25(8), 4388. https://doi.org/10.3390/ijms25084388