
IGF-1 Genome-Edited Human MSCs Exhibit Robust Anti-Arthritogenicity in Collagen-Induced Arthritis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Targeted Knock-In of IGF-1
2.2. Characteristics of AMM/I
2.3. AMM/I Immunomodulatory Properties
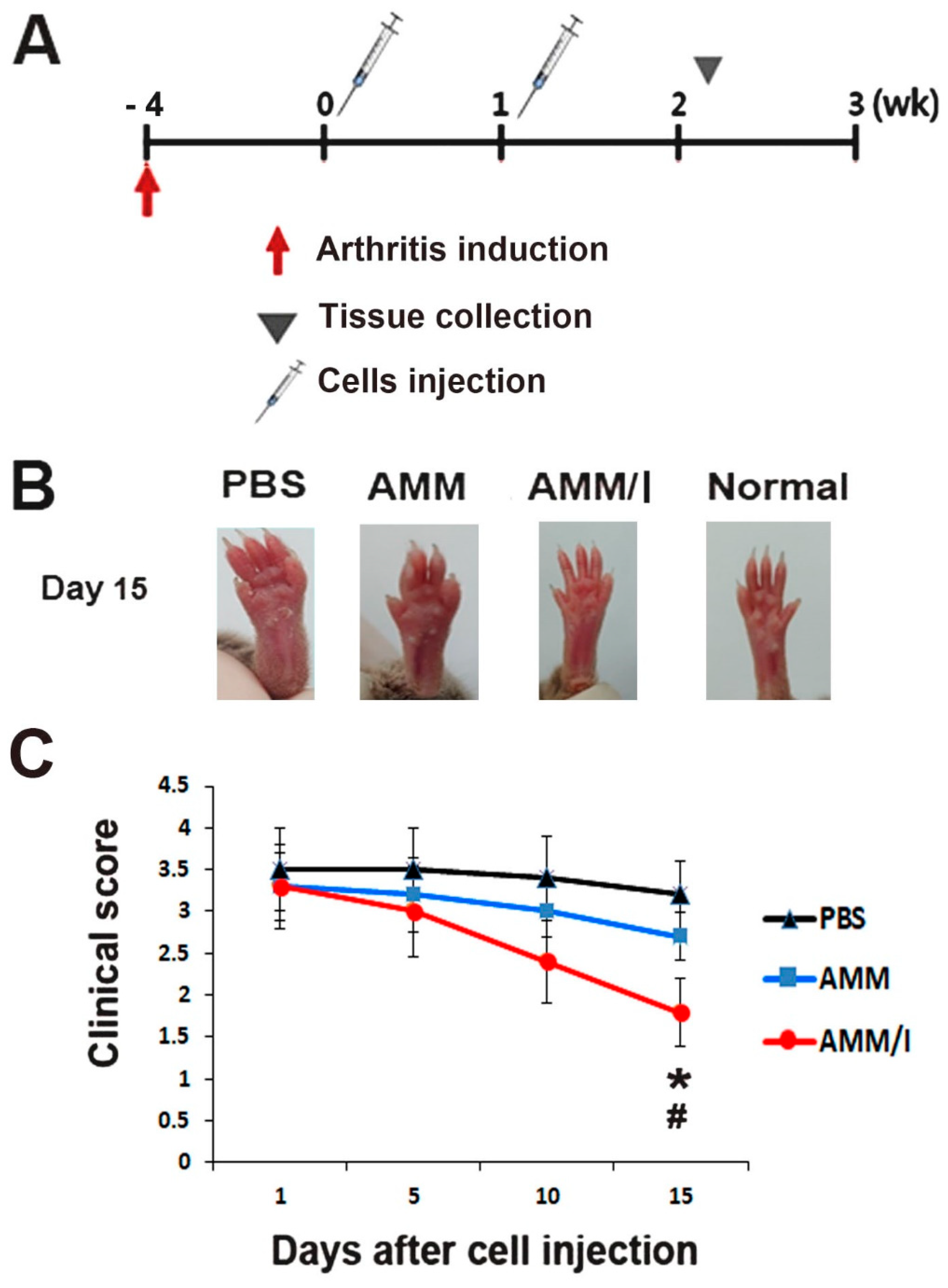
2.4. In Vivo AMM/I Anti-Arthritogenic Properties
2.5. Histological Analysis of Joints
2.6. Anti-Arthritogenic Therapeutic Mechanism
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Construction of Donor Vector, Transfection, and Selection
4.3. Junction PCR
4.4. Quantitative Reverse Transcription PCR (RT-qPCR)
4.5. Isolation of Synovial Fibroblasts
4.6. Co-Culture and Enzyme-Linked Immunosorbent Assay (ELISA)
4.7. Induction of Collagen-Induced Arthritis Model and Treatment
4.8. Flow Cytometry Analysis
4.9. Histological Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goldring, S.R. Pathogenesis of bone and cartilage destruction in rheumatoid arthritis. Rheumatology 2003, 42 (Suppl. S2), ii11–ii16. [Google Scholar] [CrossRef]
- Joshi, P.; Dhaneshwar, S.S. An update on disease modifying antirheumatic drugs. Inflamm. Allergy Drug Targets 2014, 13, 249–261. [Google Scholar] [CrossRef]
- Alghasham, A.; Rasheed, Z. Therapeutic targets for rheumatoid arthritis: Progress and promises. Autoimmunity 2014, 47, 77–94. [Google Scholar] [CrossRef]
- van den Berg, W.B.; Miossec, P. IL-17 as a future therapeutic target for rheumatoid arthritis. Nat. Rev. Rheumatol. 2009, 5, 549–553. [Google Scholar] [CrossRef]
- Melief, S.M.; Schrama, E.; Brugman, M.H.; Tiemessen, M.M.; Hoogduijn, M.J.; Fibbe, W.E.; Roelofs, H. Multipotent stromal cells induce human regulatory T cells through a novel pathway involving skewing of monocytes toward anti-inflammatory macrophages. Stem Cells 2013, 31, 1980–1991. [Google Scholar] [CrossRef]
- Debreova, M.; Culenova, M.; Smolinska, V.; Nicodemou, A.; Csobonyeiova, M.; Danisovic, L. Rheumatoid arthritis: From synovium biology to cell-based therapy. Cytotherapy 2022, 24, 365–375. [Google Scholar] [CrossRef]
- Chahal, J.; Gomez-Aristizabal, A.; Shestopaloff, K.; Bhatt, S.; Chaboureau, A.; Fazio, A.; Chisholm, J.; Weston, A.; Chiovitti, J.; Keating, A.; et al. Bone Marrow Mesenchymal Stromal Cell Treatment in Patients with Osteoarthritis Results in Overall Improvement in Pain and Symptoms and Reduces Synovial Inflammation. Stem Cells Transl. Med. 2019, 8, 746–757. [Google Scholar] [CrossRef]
- Wen, C.; Xu, L.; Xu, X.; Wang, D.; Liang, Y.; Duan, L. Insulin-like growth factor-1 in articular cartilage repair for osteoarthritis treatment. Arthritis Res. Ther. 2021, 23, 277. [Google Scholar] [CrossRef]
- Prieto-Alhambra, D.; Javaid, M.K.; Judge, A.; Maskell, J.; Cooper, C.; Arden, N.K.; Group, C.O.S. Hormone replacement therapy and mid-term implant survival following knee or hip arthroplasty for osteoarthritis: A population-based cohort study. Ann. Rheum. Dis. 2015, 74, 557–563. [Google Scholar] [CrossRef]
- Yu, X.; Xing, C.; Pan, Y.; Ma, H.; Zhang, J.; Li, W. IGF-1 alleviates ox-LDL-induced inflammation via reducing HMGB1 release in HAECs. Acta Biochim. Biophys. Sin. 2012, 44, 746–751. [Google Scholar] [CrossRef]
- Choi, J.S.; Jeong, I.S.; Han, J.H.; Cheon, S.H.; Kim, S.W. IL-10-secreting human MSCs generated by TALEN gene editing ameliorate liver fibrosis through enhanced anti-fibrotic activity. Biomater. Sci. 2019, 7, 1078–1087. [Google Scholar] [CrossRef]
- Jeong, I.S.; Park, Y.; Ryu, H.A.; An, H.S.; Han, J.H.; Kim, S.W. Dual chemotactic factors-secreting human amniotic mesenchymal stem cells via TALEN-mediated gene editing enhanced angiogenesis. Int. J. Cardiol. 2018, 260, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Cipriani, P.; Ruscitti, P.; Di Benedetto, P.; Carubbi, F.; Liakouli, V.; Berardicurti, O.; Ciccia, F.; Triolo, G.; Giacomelli, R. Mesenchymal stromal cells and rheumatic diseases: New tools from pathogenesis to regenerative therapies. Cytotherapy 2015, 17, 832–849. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Mu, R.; Wang, S.; Long, L.; Liu, X.; Li, R.; Sun, J.; Guo, J.; Zhang, X.; Yu, P.; et al. Therapeutic potential of human umbilical cord mesenchymal stem cells in the treatment of rheumatoid arthritis. Arthritis Res. Ther. 2010, 12, R210. [Google Scholar] [CrossRef]
- Choi, J.J.; Yoo, S.A.; Park, S.J.; Kang, Y.J.; Kim, W.U.; Oh, I.H.; Cho, C.S. Mesenchymal stem cells overexpressing interleukin-10 attenuate collagen-induced arthritis in mice. Clin. Exp. Immunol. 2008, 153, 269–276. [Google Scholar] [CrossRef]
- Djouad, F.; Fritz, V.; Apparailly, F.; Louis-Plence, P.; Bony, C.; Sany, J.; Jorgensen, C.; Noel, D. Reversal of the immunosuppressive properties of mesenchymal stem cells by tumor necrosis factor alpha in collagen-induced arthritis. Arthritis Rheum. 2005, 52, 1595–1603. [Google Scholar] [CrossRef]
- Chen, B.; Hu, J.; Liao, L.; Sun, Z.; Han, Q.; Song, Z.; Zhao, R.C. Flk-1+ mesenchymal stem cells aggravate collagen-induced arthritis by up-regulating interleukin-6. Clin. Exp. Immunol. 2010, 159, 292–302. [Google Scholar] [CrossRef]
- Magatti, M.; De Munari, S.; Vertua, E.; Gibelli, L.; Wengler, G.S.; Parolini, O. Human amnion mesenchyme harbors cells with allogeneic T-cell suppression and stimulation capabilities. Stem Cells 2008, 26, 182–192. [Google Scholar] [CrossRef]
- Gelse, K.; von der Mark, K.; Aigner, T.; Park, J.; Schneider, H. Articular cartilage repair by gene therapy using growth factor-producing mesenchymal cells. Arthritis Rheum. 2003, 48, 430–441. [Google Scholar] [CrossRef]
- Fosang, A.J.; Tyler, J.A.; Hardingham, T.E. Effect of interleukin-1 and insulin like growth factor-1 on the release of proteoglycan components and hyaluronan from pig articular cartilage in explant culture. Matrix 1991, 11, 17–24. [Google Scholar] [CrossRef]
- Roman-Blas, J.A.; Jimenez, S.A. NF-kappaB as a potential therapeutic target in osteoarthritis and rheumatoid arthritis. Osteoarthr. Cartil. 2006, 14, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.E.; Flierman, R.; van Duivenvoorde, L.M.; Witteveen, H.J.; van Ewijk, W.; van Laar, J.M.; de Vries, R.R.; Toes, R.E. Effective treatment of collagen-induced arthritis by adoptive transfer of CD25+ regulatory T cells. Arthritis Rheum. 2005, 52, 2212–2221. [Google Scholar] [CrossRef] [PubMed]
- Firestein, G.S. Evolving concepts of rheumatoid arthritis. Nature 2003, 423, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Suh, Y.S.; Lee, S.I.; Cheon, Y.H.; Kim, M.; Noh, H.S.; Kim, H.O. Serum IGF-1 in patients with rheumatoid arthritis: Correlation with disease activity. BMC Res. Notes 2022, 15, 128. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Adithan, A.; Alam, M.J.; Kopalli, S.R.; Kim, B.; Kang, C.W.; Hwang, K.C.; Kim, J.H. IGF-1 Facilitates Cartilage Reconstruction by Regulating PI3K/AKT, MAPK, and NF-kB Signaling in Rabbit Osteoarthritis. J. Inflamm. Res. 2021, 14, 3555–3568. [Google Scholar] [CrossRef] [PubMed]
- DiToro, D.; Harbour, S.N.; Bando, J.K.; Benavides, G.; Witte, S.; Laufer, V.A.; Moseley, C.; Singer, J.R.; Frey, B.; Turner, H.; et al. Insulin-Like Growth Factors Are Key Regulators of T Helper 17 Regulatory T Cell Balance in Autoimmunity. Immunity 2020, 52, 650–667.e10. [Google Scholar] [CrossRef]
- Erlandsson, M.C.; Toyra Silfversward, S.; Nadali, M.; Turkkila, M.; Svensson, M.N.D.; Jonsson, I.M.; Andersson, K.M.E.; Bokarewa, M.I. IGF-1R signalling contributes to IL-6 production and T cell dependent inflammation in rheumatoid arthritis. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2158–2170. [Google Scholar] [CrossRef] [PubMed]
- Miyagawa, I.; Nakayamada, S.; Kondo, M.; Tanaka, Y. Regulatory Mechanism of The Induction of Regulatory T Cells through Growth Factors Released by Human Mesenchymal Stem Cells. Crit. Rev. Immunol. 2018, 38, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Rinderknecht, E.; Humbel, R.E. The amino acid sequence of human insulin-like growth factor I and its structural homology with proinsulin. J. Biol. Chem. 1978, 253, 2769–2776. [Google Scholar] [CrossRef]
- McQuillan, D.J.; Handley, C.J.; Campbell, M.A.; Bolis, S.; Milway, V.E.; Herington, A.C. Stimulation of proteoglycan biosynthesis by serum and insulin-like growth factor-I in cultured bovine articular cartilage. Biochem. J. 1986, 240, 423–430. [Google Scholar] [CrossRef]
- Schalkwijk, J.; Joosten, L.A.; van den Berg, W.B.; van Wyk, J.J.; van de Putte, L.B. Insulin-like growth factor stimulation of chondrocyte proteoglycan synthesis by human synovial fluid. Arthritis Rheum. 1989, 32, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.E.; Kunkel, S.L.; Harlow, L.A.; Johnson, B.; Evanoff, H.L.; Haines, G.K.; Burdick, M.D.; Pope, R.M.; Strieter, R.M. Enhanced production of monocyte chemoattractant protein-1 in rheumatoid arthritis. J. Clin. Investig. 1992, 90, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Shin, T.H.; Kim, H.S.; Kang, T.W.; Lee, B.C.; Lee, H.Y.; Kim, Y.J.; Shin, J.H.; Seo, Y.; Won Choi, S.; Lee, S.; et al. Human umbilical cord blood-stem cells direct macrophage polarization and block inflammasome activation to alleviate rheumatoid arthritis. Cell Death Dis. 2016, 7, e2524. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yuan, Y.; Hu, X.; Han, R.; Chen, M.; Wang, M.; Zhang, X.; Ma, Y.; Wu, M.; Zou, Y.; et al. Low serum levels of insulin-like growth factor-1 are associated with an increased risk of rheumatoid arthritis: A systematic review and meta-analysis. Nutr. Res. 2019, 69, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.J.; Ahmed, S.; Bobrowski, P.; Haqqi, T.M. The chrondoprotective actions of a natural product are associated with the activation of IGF-1 production by human chondrocytes despite the presence of IL-1beta. BMC Complement. Altern. Med. 2006, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Fortier, L.A.; Mohammed, H.O.; Lust, G.; Nixon, A.J. Insulin-like growth factor-I enhances cell-based repair of articular cartilage. J. Bone Jt. Surg. Br. 2002, 84, 276–288. [Google Scholar] [CrossRef]
- Guo, J.; Zheng, D.; Li, W.F.; Li, H.R.; Zhang, A.D.; Li, Z.C. Insulin-like growth factor 1 treatment of MSCs attenuates inflammation and cardiac dysfunction following MI. Inflammation 2014, 37, 2156–2163. [Google Scholar] [CrossRef] [PubMed]
- Chae, D.S.; Park, Y.J.; Kim, S.W. Anti-Arthritogenic Property of Interleukin 10-Expressing Human Amniotic MSCs Generated by Gene Editing in Collagen-Induced Arthritis. Int. J. Mol. Sci. 2022, 23, 7913. [Google Scholar] [CrossRef]
- Langdon, C.; Kerr, C.; Hassen, M.; Hara, T.; Arsenault, A.L.; Richards, C.D. Murine oncostatin M stimulates mouse synovial fibroblasts in vitro and induces inflammation and destruction in mouse joints in vivo. Am. J. Pathol. 2000, 157, 1187–1196. [Google Scholar]
- Wu, C.C.; Liu, F.L.; Sytwu, H.K.; Tsai, C.Y.; Chang, D.M. CD146+ mesenchymal stem cells display greater therapeutic potential than CD146- cells for treating collagen-induced arthritis in mice. Stem Cell Res. Ther. 2016, 7, 23. [Google Scholar]
- Reisch, N.; Engler, A.; Aeschlimann, A.; Simmen, B.R.; Michel, B.A.; Gay, R.E.; Gay, S.; Sprott, H. DREAM is reduced in synovial fibroblasts of patients with chronic arthritic pain: Is it a suitable target for peripheral pain management? Arthritis Res. Ther. 2008, 10, R60. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chae, D.-S.; Han, S.; Kim, S.-W. IGF-1 Genome-Edited Human MSCs Exhibit Robust Anti-Arthritogenicity in Collagen-Induced Arthritis. Int. J. Mol. Sci. 2024, 25, 4442. https://doi.org/10.3390/ijms25084442
Chae D-S, Han S, Kim S-W. IGF-1 Genome-Edited Human MSCs Exhibit Robust Anti-Arthritogenicity in Collagen-Induced Arthritis. International Journal of Molecular Sciences. 2024; 25(8):4442. https://doi.org/10.3390/ijms25084442
Chicago/Turabian StyleChae, Dong-Sik, Seongho Han, and Sung-Whan Kim. 2024. "IGF-1 Genome-Edited Human MSCs Exhibit Robust Anti-Arthritogenicity in Collagen-Induced Arthritis" International Journal of Molecular Sciences 25, no. 8: 4442. https://doi.org/10.3390/ijms25084442
APA StyleChae, D. -S., Han, S., & Kim, S. -W. (2024). IGF-1 Genome-Edited Human MSCs Exhibit Robust Anti-Arthritogenicity in Collagen-Induced Arthritis. International Journal of Molecular Sciences, 25(8), 4442. https://doi.org/10.3390/ijms25084442