

Ferroptosis, Inflammation, and Microbiome Alterations in the Intestine in the Göttingen Minipig Model of Hematopoietic-Acute Radiation Syndrome


 and
and 
Abstract
:1. Introduction
2. Results
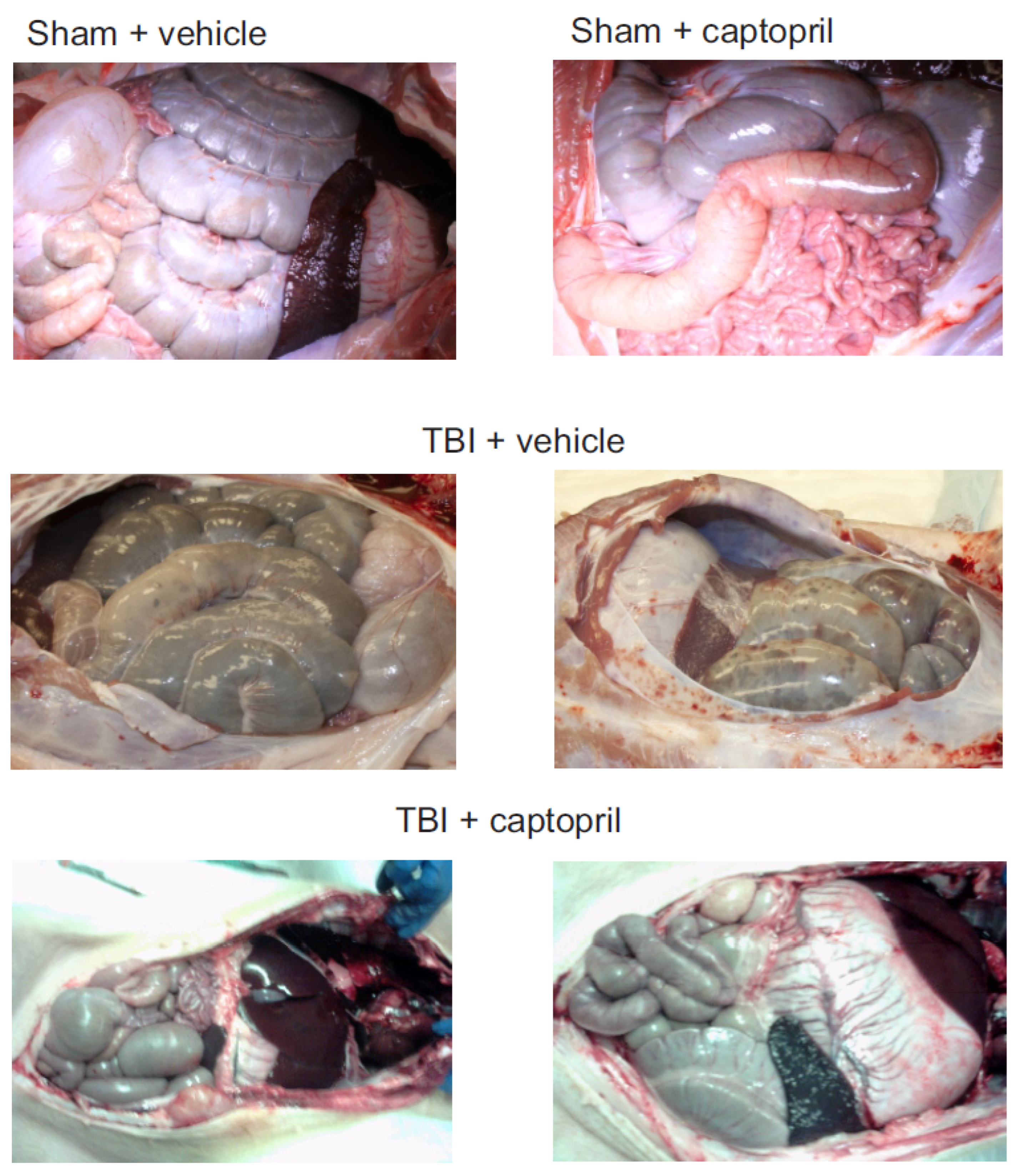
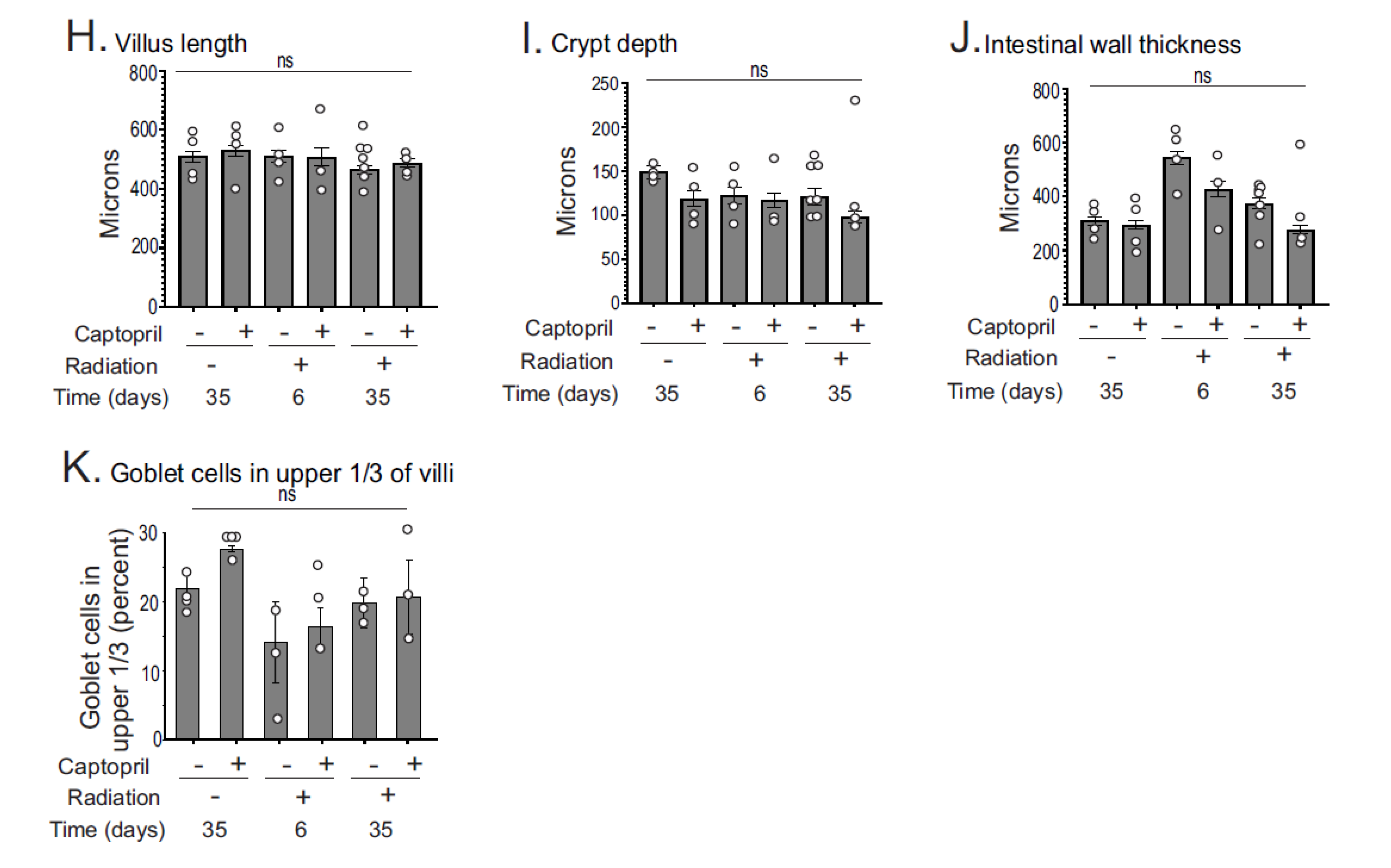
2.1. Histological Analysis of Captopril Effects on Radiation Sequelae in the Intestine
2.2. Iron-Binding Proteins and Ferroptosis-Related Enzymes Are Altered after Radiation in the GI but Are Not Significantly Affected by Captopril
2.3. Captopril Mitigates Radiation-Induced Upregulation of p21/waf1 and Some Pro-Inflammatory Cytokine Expression in the GI
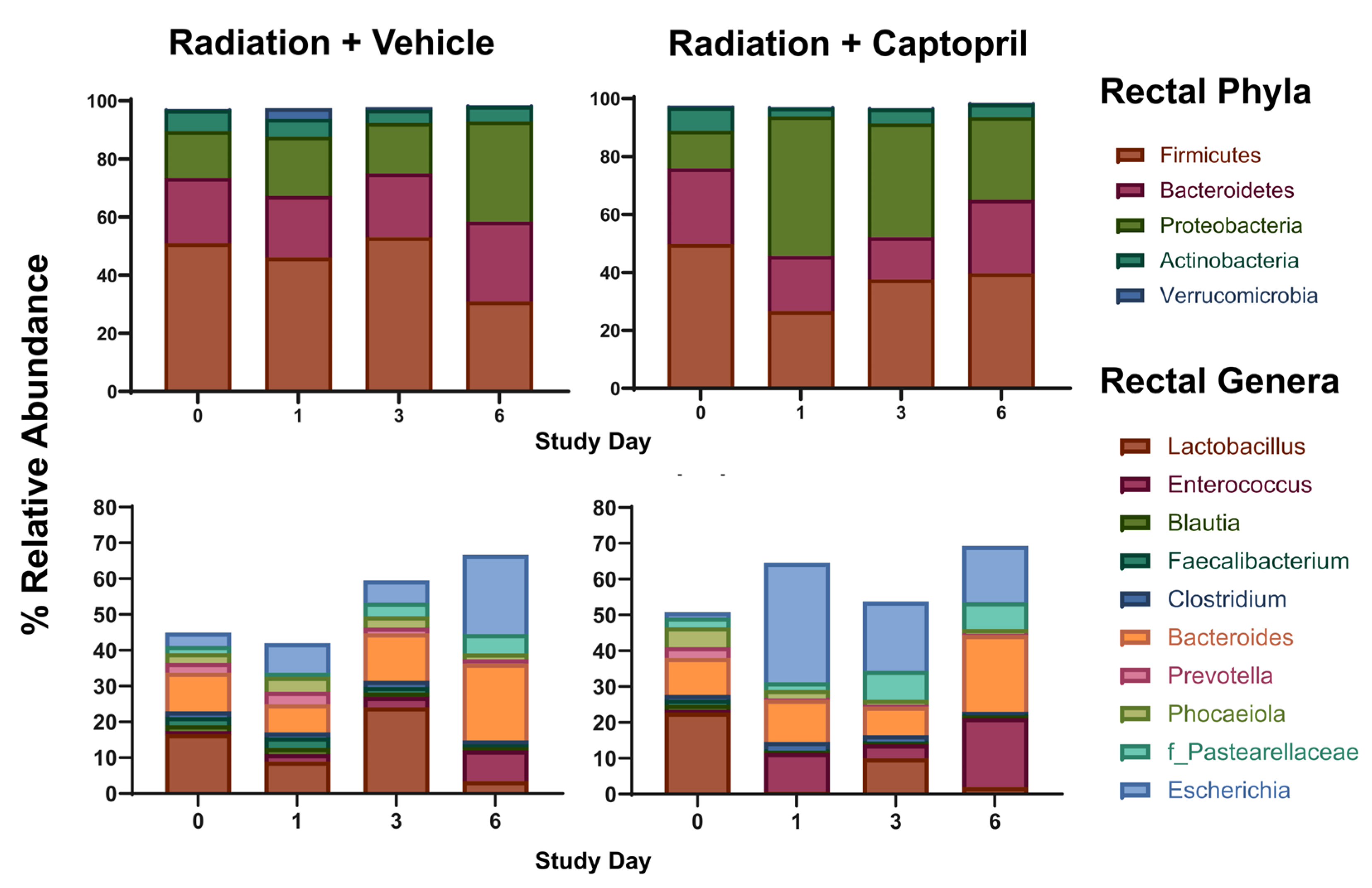
2.4. Captopril Effects on Radiation-Induced Alterations of the Gut Microbiome
3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. Animals, Irradiation, and Drug Administration
4.3. Tissue Collection and Histology
4.4. Microbiome
4.5. Western Blotting
4.6. RNA Isolation and Quantitative PCR Analysis
4.7. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garau, M.M.; Calduch, A.L.; Lopez, E.C. Radiobiology of the acute radiation syndrome. Rep. Pract. Oncol. Radiother. 2011, 16, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Acute Radiation Syndrome: A Fact Sheet for Clinicians. Available online: https://www.cdc.gov/nceh/radiation/emergencies/arsphysicianfactsheet.htm (accessed on 8 November 2022).
- Stenke, L.; Hedman, C.; Lagergren Lindberg, M.; Lindberg, K.; Valentin, J. The acute radiation syndrome-need for updated medical guidelines. J. Radiol. Prot. 2022, 42, 014004. [Google Scholar]
- Shao, L.; Luo, Y.; Zhou, D. Hematopoietic stem cell injury induced by ionizing radiation. Antioxid. Redox Signal. 2014, 20, 1447–1462. [Google Scholar] [CrossRef] [PubMed]
- Panganiban, R.A.; Snow, A.L.; Day, R.M. Mechanisms of radiation toxicity in transformed and non-transformed cells. Int. J. Mol. Sci. 2013, 14, 15931–15958. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.R.; Maity, A.; Sanzari, J.K. A review of radiation-induced coagulopathy and new findings to support potential prevention strategies and treatments. Radiat. Res. 2016, 186, 121–140. [Google Scholar] [CrossRef] [PubMed]
- Wagemaker, G. Heterogeneity of radiation sensitivity of hemopoietic stem cell subsets. Stem Cells 1995, 13 (Suppl. S1), 257–260. [Google Scholar] [CrossRef] [PubMed]
- Peslak, S.A.; Wenger, J.; Bemis, J.C.; Kingsley, P.D.; Frame, J.M.; Koniski, A.D.; Chen, Y.; Williams, J.P.; McGrath, K.E.; Dertinger, S.D.; et al. Sublethal radiation injury uncovers a functional transition during erythroid maturation. Exp. Hematol. 2011, 39, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Dorr, H.; Meineke, V. Acute radiation syndrome caused by accidental radiation exposure-therapeutic principles. BMC Med. 2011, 9, 126. [Google Scholar] [CrossRef]
- Anno, G.H.; Young, R.W.; Bloom, R.M.; Mercier, J.R. Dose response relationships for acute ionizing-radiation lethality. Health Phys. 2003, 84, 565–575. [Google Scholar] [CrossRef] [PubMed]
- McCann, D.G.C. Radiation poisoning: Current concepts in the acute radiation syndrome. Am. J. Clin. Med. 2006, 3, 13–21. [Google Scholar]
- Armed Forces Radiobiology Research Institute. Medical Management of Radiological Casualties, 4th ed.; Armed Forces Radiobiology Research Institute: Bethesda, MD, USA, 2013. [Google Scholar]
- Bensemmane, L.; Squiban, C.; Demarquay, C.; Mathieu, N.; Benderitter, M.; Le Guen, B.; Milliat, F.; Linard, C. The stromal vascular fraction mitigates radiation-induced gastrointestinal syndrome in mice. Stem Cell Res. Ther. 2021, 12, 309. [Google Scholar] [CrossRef]
- Booth, C.; Tudor, G.; Tudor, J.; Katz, B.P.; MacVittie, T.J. Acute gastrointestinal syndrome in high-dose irradiated mice. Health Phys. 2012, 103, 383–399. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Chen, Y.Z.; Zhang, Z.X.; Yang, Z.X.; Duan, G.X.; Qin, L.Q.; Zhao, L.; Xu, J.Y. At what dose can total body and whole abdominal irradiation cause lethal intestinal injury among C57BL/6J mice? Dose Response 2020, 18, 1559325820956783. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Chou, W.C.; Lai, Y.; Liang, K.; Tam, J.W.; Brickey, W.J.; Chen, L.; Montgomery, N.D.; Li, X.; Bohannon, L.M.; et al. Multi-omics analyses of radiation survivors identify radioprotective microbes and metabolites. Science 2020, 370, eaay9097. [Google Scholar] [CrossRef] [PubMed]
- Moroni, M.; Coolbaugh, T.V.; Lombardini, E.; Mitchell, J.M.; Moccia, K.D.; Shelton, L.J.; Nagy, V.; Whitnall, M.H. Hematopoietic radiation syndrome in the Gottingen minipig. Radiat. Res. 2011, 176, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Moroni, M.; Elliott, T.B.; Deutz, N.E.; Olsen, C.H.; Owens, R.; Christensen, C.; Lombardini, E.D.; Whitnall, M.H. Accelerated hematopoietic syndrome after radiation doses bridging hematopoietic (H-ARS) and gastrointestinal (GI-ARS) acute radiation syndrome: Early hematological changes and systemic inflammatory response syndrome in minipig. Int. J. Radiat. Biol. 2014, 90, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Daniel, A.R.; Luo, L.; Lee, C.L.; Kirsch, D.G. Investigating the role of inflammasome caspases 1 and 11 in the acute radiation syndrome. Radiat. Res. 2021, 196, 686–689. [Google Scholar] [CrossRef]
- Huang, S.; Huang, Y.; Lin, W.; Wang, L.; Yang, Y.; Li, P.; Xiao, L.; Chen, Y.; Chu, Q.; Yuan, X. Sitagliptin alleviates radiation-induced intestinal injury by activating NRF2-antioxidant axis, mitigating NLRP3 inflammasome activation, and reversing gut microbiota disorder. Oxid. Med. Cell Longev. 2022, 2022, 2586305. [Google Scholar] [PubMed]
- Wang, L.; Wang, A.; Fu, Q.; Shi, Z.; Chen, X.; Wang, Y.; Xu, W.; Wang, T.; Zhang, S.; Hu, S. Ferroptosis plays an important role in promoting ionizing radiation-induced intestinal injuries. Biochem. Biophys. Res. Commun. 2022, 595, 7–13. [Google Scholar] [CrossRef]
- Li, J.; Cao, F.; Yin, H.L.; Huang, Z.J.; Lin, Z.T.; Mao, N.; Sun, B.; Wang, G. Ferroptosis: Past, present and future. Cell Death Dis. 2020, 11, 88. [Google Scholar] [CrossRef]
- Elliott, T.B.; Bolduc, D.L.; Ledney, G.D.; Kiang, J.G.; Fatanmi, O.O.; Wise, S.Y.; Romaine, P.L.; Newman, V.L.; Singh, V.K. Combined immunomodulator and antimicrobial therapy eliminates polymicrobial sepsis and modulates cytokine production in combined injured mice. Int. J. Radiat. Biol. 2015, 91, 690–702. [Google Scholar] [CrossRef]
- Ciorba, M.A.; Riehl, T.E.; Rao, M.S.; Moon, C.; Ee, X.; Nava, G.M.; Walker, M.R.; Marinshaw, J.M.; Stappenbeck, T.S.; Stenson, W.F. Lactobacillus probiotic protects intestinal epithelium from radiation injury in a TLR-2/cyclo-oxygenase-2-dependent manner. Gut 2012, 61, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Riehl, T.E.; Alvarado, D.; Ee, X.; Zuckerman, A.; Foster, L.; Kapoor, V.; Thotala, D.; Ciorba, M.A.; Stenson, W.F. Lactobacillus rhamnosus GG protects the intestinal epithelium from radiation injury through release of lipoteichoic acid, macrophage activation and the migration of mesenchymal stem cells. Gut 2019, 68, 1003–1013. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Dong, J.; Xiao, H.; Zhang, S.; Wang, B.; Cui, M.; Fan, S. Gut commensal derived-valeric acid protects against radiation injuries. Gut Microbes 2020, 11, 789–806. [Google Scholar] [CrossRef] [PubMed]
- Ericsson, A.C. The use of non-rodent model species in microbiota studies. Lab. Anim. 2019, 53, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Ericsson, A.C.; Busi, S.B.; Amos-Landgraf, J.M. Characterization of the rat gut microbiota via 16S rRNA amplicon library sequencing. Methods Mol. Biol. 2019, 2018, 195–212. [Google Scholar] [PubMed]
- Hansen, A.K.; Hansen, C.H.F. The microbiome and rodent models of immune mediated diseases. Mamm. Genome 2021, 32, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.; Zhang, X. Alien species invasion of deep-sea bacteria into mouse gut microbiota. J. Adv. Res. 2023, 45, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Charrier, S.; Michaud, A.; Badaoui, S.; Giroux, S.; Ezan, E.; Sainteny, F.; Corvol, P.; Vainchenker, W. Inhibition of angiotensin I-converting enzyme induces radioprotection by preserving murine hematopoietic short-term reconstituting cells. Blood 2004, 104, 978–985. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.A.; Landauer, M.R.; Mog, S.R.; Barshishat-Kupper, M.; Zins, S.R.; Amare, M.F.; Day, R.M. Timing of captopril administration determines radiation protection or radiation sensitization in a murine model of total body irradiation. Exp. Hematol. 2010, 38, 270–281. [Google Scholar] [CrossRef]
- Barshishat-Kupper, M.; Mungunsukh, O.; Tipton, A.J.; McCart, E.A.; Panganiban, R.A.; Davis, T.A.; Landauer, M.R.; Day, R.M.l. Captopril modulates hypoxia-inducible factors and erythropoietin responses in a murine model of total body irradiation. Exp. Hematol. 2011, 39, 293–304. [Google Scholar] [CrossRef]
- McCart, E.A.; Lee, Y.H.; Jha, J.; Mungunsukh, O.; Rittase, W.B.; Summers, T.A., Jr.; Muir, J.; Day, R.M. Delayed captopril administration mitigates hematopoietic injury in a murine model of total body irradiation. Sci. Rep. 2019, 9, 2198. [Google Scholar] [CrossRef] [PubMed]
- Rittase, W.B.; McCart, E.A.; Muir, J.M.; Bouten, R.M.; Slaven, J.E.; Mungunsukh, O.; Bylicky, M.A.; Wilkins, W.L.; Lee, S.H.; Gudmundsson, K.O.; et al. Effects of captopril against radiation injuries in the Göttingen minipig model of hematopoietic-acute radiation syndrome. PLoS ONE 2021, 16, e0256208. [Google Scholar] [CrossRef]
- Rodgers, K.E.; Espinoza, T.; Roda, N.; Meeks, C.J.; Hill, C.; Louie, S.G.; Dizerega, G.S. Accelerated hematopoietic recovery with angiotensin-(1-7) after total body radiation. Int. J. Radiat. Biol. 2012, 88, 466–476. [Google Scholar] [CrossRef]
- Kim, Y.C.; Mungunsukh, O.; Day, R.M. Erythropoietin regulation by angiotensin II. Vitam. Horm. 2017, 105, 57–77. [Google Scholar] [PubMed]
- Hubert, C.; Savary, K.; Gasc, J.M.; Corvol, P. The hematopoietic system: A new niche for the renin-angiotensin system. Nat. Clin. Pract. Cardiovasc. Med. 2006, 3, 80–85. [Google Scholar] [CrossRef]
- Julien, E.; Biasch, K.; El Omar, R.; Freund, J.N.; Gachet, C.; Lanza, F.; Tavian, M. Renin-angiotensin system is involved in embryonic emergence of hematopoietic stem/progenitor cells. Stem Cells 2021, 39, 636–649. [Google Scholar] [CrossRef] [PubMed]
- Day, R.M.; Davis, T.A.; Barshishat-Kupper, M.; McCart, E.A.; Tipton, A.J.; Landauer, M.R. Enhanced hematopoietic protection from radiation by the combination of genistein and captopril. Int. Immunopharmacol. 2013, 15, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Schorpp, K.; Jin, J.; Yozwiak, C.E.; Hoffstrom, B.G.; Decker, A.M.; Rajbhandari, P.; Stokes, M.E.; Bender, H.G.; Csuka, J.M.; et al. Transferrin receptor is a specific ferroptosis marker. Cell Rep. 2020, 30, 3411–3423.e7. [Google Scholar] [CrossRef]
- Lei, P.; Bai, T.; Sun, Y. Mechanisms of ferroptosis and relations with regulated cell death: A Review. Front. Physiol. 2019, 10, 139. [Google Scholar] [CrossRef]
- Rittase, W.B.; Slaven, J.E.; Suzuki, Y.J.; Muir, J.M.; Lee, S.H.; Rusnak, M.; Brehm, G.V.; Bradfield, D.T.; Symes, A.J.; Day, R.M. Iron deposition and ferroptosis in the spleen in a murine model of acute radiation syndrome. Int. J. Mol. Sci. 2022, 23, 11029. [Google Scholar] [CrossRef]
- Adedoyin, O.; Boddu, R.; Traylor, A.; Lever, J.M.; Bolisetty, S.; George, J.F.; Agarwal, A. Heme oxygenase-1 mitigates ferroptosis in renal proximal tubule cells. Am. J. Physiol. Renal Physiol. 2018, 314, F702–F714. [Google Scholar] [CrossRef] [PubMed]
- Lei, G.; Zhang, Y.; Koppula, P.; Liu, X.; Zhang, J.; Lin, S.H.; Ajani, J.A.; Xiao, Q.; Liao, Z.; Wang, H.; et al. The role of ferroptosis in ionizing radiation-induced cell death and tumor suppression. Cell Res. 2020, 30, 146–162. [Google Scholar] [CrossRef] [PubMed]
- Imai, H.; Matsuoka, M.; Kumagai, T.; Sakamoto, T.; Koumura, T. Lipid peroxidation-dependent cell death regulated by GPX4 and ferroptosis. Curr. Top. Microbiol. Immunol. 2017, 403, 143–170. [Google Scholar] [PubMed]
- Hong, S.H.; Lee, D.H.; Lee, Y.S.; Jo, M.J.; Jeong, Y.A.; Kwon, W.T.; Choudry, H.A.; Bartlett, D.L.; Lee, Y.J. Molecular crosstalk between ferroptosis and apoptosis: Emerging role of ER stress-induced p53-independent PUMA expression. Oncotarget 2017, 8, 115164–115178, Correction in Oncotarget 2018, 9, 24869. [Google Scholar] [CrossRef]
- Hyoh, Y.; Ishizaka, S.; Horii, T.; Fujiwara, A.; Tegoshi, T.; Yamada, M.; Arizono, N. Activation of caspases in intestinal villus epithelial cells of normal and nematode infected rats. Gut 2002, 50, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Bernardini, N.; Colucci, R.; Mattii, L.; Segnani, C.; Fornai, M.; de Giorgio, R.; Barbara, G.; Castagna, M.; Nardini, V.; Dolfi, A.; et al. Constitutive expression of cyclooxygenase-2 in the neuromuscular compartment of normal human colon. Neurogastroenterol. Motil. 2006, 18, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Garg, S.; Boerma, M.; Wang, J.; Fu, Q.; Loose, D.S.; Kumar, K.S.; Hauer-Jensen, M. Influence of sublethal total-body irradiation on immune cell populations in the intestinal mucosa. Radiat. Res. 2010, 173, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Hull, L.; Zizzo, A.; Wang, L.; Lin, B.; Zhai, M.; Xiao, M. Pharmacokinetic study of rhIL-18BP and its effect on radiation-induced cytokine changes in mouse serum and intestine. Toxics 2022, 11, 35. [Google Scholar] [CrossRef]
- Wang, A.; Ling, Z.; Yang, Z.; Kiela, P.R.; Wang, T.; Wang, C.; Cao, L.; Geng, F.; Shen, M.; Ran, X.; et al. Gut microbial dysbiosis may predict diarrhea and fatigue in patients undergoing pelvic cancer radiotherapy: A pilot study. PLoS ONE 2015, 10, e0126312. [Google Scholar]
- Pabst, R. The pig as a model for immunology research. Cell Tissue Res. 2020, 380, 287–304. [Google Scholar] [CrossRef]
- Zhang, J.; Zheng, L.; Wang, Z.; Hailong, P.; Hu, W.; Nie, J.; Shang, P.; Li, B.; Hei, T.K.; Zhou, G. Lowering iron levels protects against bone loss in focally irradiated and contralateral femurs through distinct mechanisms. Bone 2019, 120, 50–60. [Google Scholar] [CrossRef]
- Zhang, X.; Xing, X.; Liu, H.; Feng, J.; Tian, M.; Chang, S.; Liu, P.; Zhang, H. Ionizing radiation induces ferroptosis in granulocyte-macrophage hematopoietic progenitor cells of murine bone marrow. Int. J. Radiat. Biol. 2020, 96, 584–595. [Google Scholar] [CrossRef] [PubMed]
- Rittase, W.B.; Muir, J.M.; Slaven, J.E.; Bouten, R.M.; Bylicky, M.A.; Wilkins, W.L.; Day, R.M. Deposition of iron in the bone marrow of a murine model of hematopoietic acute radiation syndrome. Exp. Hematol. 2020, 84, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.H.; Liu, J.H.Z.; Liu, C.Y.; Sun, W.Y.; Duan, W.J.; Wang, G.; Kurihara, H.; He, R.R.; Li, F.; Chen, Y.; et al. ALOX15-launched PUFA-phospholipids peroxidation increases the susceptibility of ferroptosis in ischemia-induced myocardial damage. Signal Transduct. Target. Ther. 2022, 7, 288. [Google Scholar] [CrossRef] [PubMed]
- Mungunsukh, O.; George, J.; McCart, E.A.; Snow, A.L.; Mattapallil, J.J.; Mog, S.R.; Panganiban, R.A.M.; Bolduc, D.L.; Rittase, W.B.; Bouten, R.M.; et al. Captopril reduces lung inflammation and accelerated senescence in response to thoracic radiation in mice. J. Radiat. Res. 2021, 62, 236–248. [Google Scholar] [CrossRef] [PubMed]
- Tsujiguchi, T.; Hirouchi, T.; Monzen, S.; Tabuchi, Y.; Takasaki, I.; Kondo, T.; Kashiwakura, I. Expression analysis of radiation-responsive genes in human hematopoietic stem/progenitor cells. J. Radiat. Res. 2016, 57, 35–43. [Google Scholar] [CrossRef]
- Sato, Y.; Yoshino, H.; Ishikawa, J.; Monzen, S.; Yamaguchi, M.; Kashiwakura, I. Prediction of hub genes and key pathways associated with the radiation response of human hematopoietic stem/progenitor cells using integrated bioinformatics methods. Sci. Rep. 2023, 13, 10762. [Google Scholar] [CrossRef] [PubMed]
- Shankar, G.; Cohen, D.A. Idiopathic pneumonia syndrome after bone marrow transplantation: The role of pre-transplant radiation conditioning and local cytokine dysregulation in promoting lung inflammation and fibrosis. Int. J. Exp. Pathol. 2001, 82, 101–113. [Google Scholar]
- Bouten, R.M.; Dalgard, C.L.; Soltis, A.R.; Slaven, J.E.; Day, R.M. Transcriptomic profiling and pathway analysis of cultured human lung microvascular endothelial cells following ionizing radiation exposure. Sci. Rep. 2021, 11, 24214. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Jian, Y.; Wang, F.; Yu, S.; Guo, J.; Kan, J.; Guo, W. NLRP3 and gut microbiota homeostasis: Progress in research. Cells 2022, 11, 3758. [Google Scholar] [CrossRef]
- Hollingsworth, B.A.; Cassatt, D.R.; DiCarlo, A.L.; Rios, C.I.; Satyamitra, M.M.; Winters, T.A.; Taliaferro, L.P. Acute radiation syndrome and the microbiome: Impact and review. Front. Pharmacol. 2021, 12, 643283. [Google Scholar] [CrossRef]
- Cheema, A.K.; Li, Y.; Singh, J.; Johnson, R.; Girgis, M.; Wise, S.Y.; Fatanmi, O.O.; Kaytor, M.D.; Singh, V.K. Microbiome study in irradiated mice treated with BIO 300, a promising radiation countermeasure. Anim. Microbiome 2021, 3, 71. [Google Scholar] [CrossRef] [PubMed]
- Carbonero, F.; Mayta, A.; Bolea, M.; Yu, J.Z.; Lindeblad, M.; Lyubimov, A.; Neri, F.; Szilagyi, E.; Smith, B.; Halliday, L.; et al. Specific members of the gut microbiota are reliable biomarkers of irradiation intensity and lethality in large animal models of human health. Radiat. Res. 2019, 191, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Carbonero, F.; Mayta-Apaza, A.C.; Yu, J.Z.; Lindeblad, M.; Lyubimov, A.; Neri, F.; Szilagyi, E.; Bartholomew, A. A comparative analysis of gut microbiota disturbances in the Gottingen minipig and rhesus macaque models of acute radiation syndrome following bioequivalent radiation exposures. Radiat. Environ. Biophys. 2018, 57, 419–426. [Google Scholar] [CrossRef]
- Kalkeri, R.; Walters, K.; Van Der Pol, W.; McFarland, B.C.; Fisher, N.; Koide, F.; Morrow, C.D.; Singh, V.K. Changes in the gut microbiome community of nonhuman primates following radiation injury. BMC Microbiol. 2021, 21, 93. [Google Scholar] [CrossRef] [PubMed]
- Yao, T.; Li, L. The influence of microbiota on ferroptosis in intestinal diseases. Gut Microbes 2023, 15, 2263210. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Ullah, N.; Zha, L.; Bai, Y.; Khan, A.; Zhao, T.; Che, T.; Zhang, C. Alteration of gut microbiota in inflammatory bowel disease (IBD): Cause or consequence? IBD treatment targeting the gut microbiome. Pathogens 2019, 8, 126. [Google Scholar] [CrossRef] [PubMed]
- Touchefeu, Y.; Montassier, E.; Nieman, K.; Gastinne, T.; Potel, G.; Bruley des Varannes, S.; Le Vacon, F.; de La Cochetiere, M.F. Systematic review: The role of the gut microbiota in chemotherapy- or radiation-induced gastrointestinal mucositis—Current evidence and potential clinical applications. Aliment Pharmacol Ther. 2014, 40, 409–421. [Google Scholar] [CrossRef]
- Hu, C.; Liu, M.; Tang, L.; Liu, H.; Sun, B.; Chen, L. Probiotic intervention mitigates the metabolic disturbances of perfluorobutanesulfonate along the gut-liver axis of zebrafish. Chemosphere 2021, 284, 131374. [Google Scholar] [CrossRef]
- Deng, F.; Zhao, B.C.; Yang, X.; Lin, Z.B.; Sun, Q.S.; Wang, Y.F.; Yan, Z.Z.; Liu, W.F.; Li, C.; Hu, J.J.; et al. The gut microbiota metabolite capsiate promotes GPX4 expression by activating TRPV1 to inhibit intestinal ischemia reperfusion-induced ferroptosis. Gut Microbes 2021, 13, 1902719. [Google Scholar] [CrossRef]
- Gu, K.; Wu, A.; Yu, B.; Zhang, T.; Lai, X.; Chen, J.; Yan, H.; Zheng, P.; Luo, Y.; Luo, J. Iron overload induces colitis by modulating ferroptosis and interfering gut microbiota in mice. Sci. Total Environ. 2023, 905, 167043. [Google Scholar]
- Yang, T.; Aquino, V.; Lobaton, G.O.; Li, H.; Colon-Perez, L.; Goel, R.; Qi, Y.; Zubcevic, J.; Febo, M.; Richards, E.M.; et al. Sustained captopril-induced reduction in blood pressure is associated with alterations in gut-brain axis in the spontaneously hypertensive rat. J. Am. Heart Assoc. 2019, 8, e010721. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Kang, H.; Zhang, W.; Wang, J.; Liu, Z.; Jing, J.; Han, L.; Gao, A. Probiotics ameliorate benzene-induced systemic inflammation and hematopoietic toxicity by inhibiting Bacteroidaceae-mediated ferroptosis. Sci. Total Environ. 2023, 899, 165678. [Google Scholar] [CrossRef] [PubMed]
- Center for Drug Evaluation and Research. Product Development Under the Animal Rule. Guidance for Industry; U.S. Department of Health and Human Services, Food and Drug Administration: Silver Spring, MD, USA, 2015. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/product-development-under-animal-rule (accessed on 21 October 2023).
- Kenchegowda, D.; Seed, T.M.; Singh, V.K. What are the practical, ethical, and pathobiological considerations in the use of minipigs as an animal model in drug discovery for acute radiation syndrome and delayed effects of acute radiation exposure? Expert. Opin. Drug Discov. 2021, 16, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Moroni, M.; Coolbaugh, T.V.; Mitchell, J.M.; Lombardini, E.; Moccia, K.D.; Shelton, L.J.; Nagy, V.; Whitnall, M.H. Vascular access port implantation and serial blood sampling in a Gottingen minipig (Sus scrofa domestica) model of acute radiation injury. J. Am. Assoc. Lab. Anim. Sci. 2011, 50, 65–72. [Google Scholar] [PubMed]
- Moroni, M.; Ngudiankama, B.F.; Christensen, C.; Olsen, C.H.; Owens, R.; Lombardini, E.D.; Holt, R.K.; Whitnall, M.H. The Göttingen minipig is a model of the hematopoietic acute radiation syndrome: G-colony stimulating factor stimulates hematopoiesis and enhances survival from lethal total-body gamma-irradiation. Int. J. Radiat. Oncol. Biol. Phys. 2013, 86, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Krakau, S.; Straub, D.; Gourle, H.; Gabernet, G.; Nahnsen, S. nf-core/mag: A best-practice pipeline for metagenome hybrid assembly and binning. NAR Genom. Bioinform. 2022, 4, lqac007. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef]
- Youngblut, N.D.; Ley, R.E. Struo2: Efficient metagenome profiling database construction for ever-expanding microbial genome datasets. PeerJ 2021, 9, e12198. [Google Scholar] [CrossRef]
- Lu, J.; Breitwieser, F.P.; Thielen, P.; Salzberg, S.L. Bracken: Estimating species abundance in metagenomics data. PeerJ Comp. Sci. 2017, 3, e104. [Google Scholar] [CrossRef]
- Dabdoub, S.M. Kraken-biom: Enabling Interoperative Format Conversion for Kraken Results (Version 1.2). 2016. Available online: https://github.com/smdabdoub/kraken-biom (accessed on 21 October 2023).
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Bisanz, J.E. qiime2R: Importing QIIME2 Artifacts and Associated Data into R Sessions. Version 0.99. 2018. Available online: https://github.com/jbisanz/qiime2R (accessed on 21 October 2023).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vehicle Group (% of Phyla) | Captopril Group (% of Phyla) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Day a | 0 | 1 | 3 | 6 | 0 | 1 | 3 | 6 | |
| Phylum | |||||||||
| Firmicutes | 50.97 (9.49) | 46.11 (2.06) (p = 0.99) c | 53.10 (11.82) (p = 0.99) c | 30.96 (10.72) (p = 0.99) c | 49.79 (3.34) (p = 0.99) b | 26.66 (4.14) (p = 0.22) b (p = 0.17) c | 37.60 (10.89) (p = 0.82) b (p = 0.24) c | 39.61 (5.46) (p = 1) b (p = 1) c | |
| Bacteriodetes | 2.46 (5.57) | 21.15 (4.39) (p = 0.99) c | 21.87 (9.49) (p = 0.99) c | 27.41 (9.26) (p = 0.99) c | 26.13 (2.33) (p = 0.99) b | 19.07 (1.02) (p = 0.09) b (p = 0.30) c | 14.58 (2.39) (p = 0.71) b (p = 0.10) c | 25.52 (6.02) (p = 1) b (p = 1) c | |
| Proteobacteria | 16.14 (3.53) | 20.40 (1.88) (p = 0.99) c | 17.46 (4.31) (p = 0.99) c | 34.52 (17.03) (p = 0.99) c | 12.95 (1.78) (p = 0.99) b | 48.08 (6.72) (p = 0.23) b (p = 0.27) c | 39.22 (12.12) (p = 1) b (p = 0.94) c | 28.51 (4.56) (p = 1) b (p = 0.27) c | |
| Actinobacteria | 7.29 (0.83) | 6.16 (0.29) (p = 0.99) c | 4.52 (2.22) (p = 0.99) c | 5.31 (2.38) (p = 0.99) c | 8.15 (0.65) (p = 0.99) b | 3.05 (1.64) (p = 0.44) b (p = 0.06) c | 5.14 (2.83) (p = 1) b (p = 0.14) c | 4.54 (2.37) (p = 1) b (p = 0.23) c | |
| Verrucomicrobia | 0.371 (0.12) | 3.69 (1.96) (p = 0.99) c | 0.90 (0.18) (p = 0.99) c | 0.29 (0.23) (p = 0.99) c | 0.54 (0.43) (p = 0.99) b | 0.23 (0.14) (p = 0.003) b (p = 0.19) c | 0.15 (0.11) (p = 0.49) b (p = 0.19) c | 0.35 (0.21) (p = 1) b (p = 1) c | |
| Vehicle (% of Total Genera) | Captopril (% of Total Genera) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Day a | 0 | 1 | 3 | 6 | 0 | 1 | 3 | 6 | |
| Genus | |||||||||
| Lactobacillus | 16.61 (5.51) | 8.95 (2.39) | 24.09 (12.50) | 3.49 (2.86) | 22.79 (6.39) (p = 0.99) b | 0.64 (0.34) (p = 0.016) b | 10.00 (7.20) (p = 0.99) b | 1.95 (1.42) (p = 0.99) b | |
| Streptococcus | 1.77 (0.40) | 1.64 (1.13) | 0.84 (0.21) | 0.92 (0.52) | 1.79 (0.08) (p = 0.99) b | 0.81 (0.60) (p = 0.88) b | 0.81 (0.25) (p = 0.99) b | 1.10 (0.44) (p = 0.99) b | |
| Enterococcus | 0.80 (0.33) | 2.07 (1.10) | 2.94 (2.31) | 8.56 (3.87) | 0.76 (0.07) (p = 0.87) b | 10.86 (3.35) (p = 0.59) b | 3.91 (2.19) (p = 0.99) b | 19.28 (3.73) (p = 0.99) b | |
| Paenibacillus | 1.30 (0.29) | 1.26 (0.28) | 0.89 (0.39) | 0.66 (0.57) | 1.13 (0.25) (p = 0.99) b | 0.47 (0.24) (p = 0.083) b | 1.0 (0.52) (p = 0.99) b | 0.20 (0.10) (p = 0.99) b | |
| Blautia | 1.71 (0.37) | 1.77 (0.46) | 1.21 (0.47) | 0.88 (0.68) | 1.43 (0.09) (p = 0.99) b | 0.46 (0.17) (p = 0.088) b | 0.38 (0.24) (p = 0.99) b | 0.65 (0.21) (p = 0.99) b | |
| Roseburia | 1.73 (0.49) | 1.36 (0.34) | 1.10 (0.63) | 0.73 (0.66) | 1.17 (0.44) (p = 0.99) b | 0.47 (0.26) (p = 0.15) b | 0.28 (0.21) (p = 0.99) b | 0.11 (0.08) (p = 0.99) b | |
| f Lachnospiraceae | 1.34 (0.35) | 1.27 (0.33) | 0.94 (0.45) | 0.68 (0.60) | 0.95 (0.22) (p = 0.99) b | 0.29 (0.11) (p = 0.049) b | 0.25 (0.18) (p = 0.99) b | 0.12 (0.08) (p = 0.99) b | |
| Faecalibacterium | 2.27 (0.54) | 2.93 (0.61) | 1.55 (0.84) | 0.84 (0.76) | 1.41 (0.29) (p = 0.99) b | 0.20 (0.07) (p = 0.049) b | 0.40 (0.33) (p = 0.99) b | 0.16 (0.12) (p = 0.99) b | |
| Clostridium | 1.61 (0.26) | 1.36 (0.23) | 1.68 (0.17) | 1.06 (0.40) | 1.33 (0.27) (p = 0.99) b | 2.44 (0.94) (p = 0.18) b | 1.77 (0.35) (p = 0.99) b | 0.94 (0.20) (p = 0.99) b | |
| Corynebacterium | 1.548 (0.71) | 0.32 (0.03) | 0.18 (0.05) | 2.04 (1.53) | 2.16 (0.13) (p = 0.99) b | 1.08 (0.77) (p = 0.16) b | 0.60 (0.26) (p = 0.99) b | 3.20 (1.87) (p = 0.99) b | |
| Bacteroides | 10.74 (6.26) | 7.91 (2.45) | 13.29 (10.27) | 21.49 (10.81) | 10.35(4.74) (p = 0.99) b | 11.81 (2.23) (p = 0.58) b | 7.96 (2.66) (p = 0.99) b | 21.47 (5.55) (p = 0.99) b | |
| Prevotella | 2.79 (0.77) | 3.42 (1.33) | 1.59 (0.77) | 1.20 (1.11) | 2.98 (0.24) (p = 0.99) b | 0.32 (0.11) (p = 0.045) b | 0.53 (0.44) (p = 0.99) b | 0.36 (0.21) (p = 0.99) b | |
| Phocaeiola | 2.71 (0.40) | 4.24 (0.88) | 3.18 (0.61) | 1.67 (1.14) | 5.50 (0.71) (p = 0.99) b | 2.36 (1.12) (p = 0.62) b | 1.45 (1.24) (p = 0.99) b | 1.32 (0.51) (p = 0.99) b | |
| f Pastearellaceae | 1.99 (0.87) | 1.09 (0.74) | 3.77 (2.53) | 5.34 (3.41) | 2.75 (0.80) (p = 0.99) b | 2.13 (1.24) (p = 0.31) b | 8.04 (4.51) (p = 0.99) b | 7.42 (2.00) (p = 0.99) b | |
| Escherichia | 3.76 (3.31) | 8.32 (2.87) | 6.27 (3.40) | 22.11 (19.60) | 1.47 (0.94) (p = 0.99) b | 33.42 10.31) (p = 0.30 )b | 19.34 10.79) (p = 0.99) b | 15.75 (5.01) (p = 0.99) b | |
| Ruminococcus | 1.22 (0.42) | 1.08 (0.20) | 0.72 (0.36) | 0.48 (0.44) | 0.77 (0.06) (p = 0.99) b | 0.12 (0.04) (p = 0.053) b | 0.26 (0.22) (p = 0.99) b | 0.12 (0.09) (p = 0.99) b | |
| Forward Primer | Reverse Primer | ||
|---|---|---|---|
| Gene (Amplicon Size) | |||
| CCL2 (121 bp) | 5′-AGAAGATCTCGATGCAGCGG-3′ | 5′-TTCTGCTTGGGTTCTGCACA-3′ | |
| CXCL8 (166 bp) | 5′-TGCAGAACTTCGATGCCAGT-3′ | 5′-CCACTTTTCCTTGGGGTCCA-3′ | |
| GAPDH (196 bp) | 5′-GTCGGAGTGAACGGATTTG-3′ | 5′-CATTGATGACAAGCTTCCCG-3′ | |
| GPX4 (252 bp) | 5′-GAATTCGGCACGAGAGGAGC-3′ | 5′-TTGGTGACGATGCACACGTA-3′ | |
| HMOX1 (277 bp) | 5′-CGCCTTCCTGCTCAACATTC-3′ | 5′-ACGGTTGCATTCACAGGGTT-3′ | |
| IL1B (162 bp) | 5′-TGTCTGTGATGCCAACGTG-3′ | 5′-TCATCTCCTTGCACAAAGCTC-3′ | |
| IL6 (101 bp) | 5′-GTCGAGGCCGTGCAGATTAG-3′ | 5′-GCATTTGTGGTGGGGTTAGG-3′ | |
| IL10 (not disclosed) | Prime PCR assay, Bio-Rad primer | ||
| IL18 (147 bp) | 5′-GGCAGTAACCATCTCTGTGCA-3′ | 5′-TGTCCAGGAACACTTCTCTGA-3′ | |
| NLRP3 (70 bp) | 5′-TTCTTCCATGGCTCAGGACAC-3′ | 5′-AGGGCATAGGTCCACACAAAA-3′ | |
| NRF2 (268 bp) | 5′-CTAAGGGTGCTCCTTTGCGA-3′ | 5′-CATGCTCCTTCCGTCGTTGA-3′ | |
| PTGS2 (281 bp) | 5′-AGGACCAGCTTTCACCAAAGG-3 | 5′-TATGTTCCCGCAGCCAGATTG-3 | |
| SLC7A11 (287 bp) | 5′-CCTGGGCAGGAGAAAGTTGT-3′ | 5′-CAGACTCGCACAAAAGCTGG-3′ | |
| SLC40A1 (268 bp) | 5′-TAAAGTGGCCCAGACGTCAC-3′ | 5′-TCGTATTGTAGCATTCATATCTGC-3′ | |
| TNFA (129 bp) | 5′-GATTCAGGGATGTGTGGCCT-3′ | 5′-GCCACATTCCAGATGTCCCA-3′ | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horseman, T.; Rittase, W.B.; Slaven, J.E.; Bradfield, D.T.; Frank, A.M.; Anderson, J.A.; Hays, E.C.; Ott, A.C.; Thomas, A.E.; Huppmann, A.R.; et al. Ferroptosis, Inflammation, and Microbiome Alterations in the Intestine in the Göttingen Minipig Model of Hematopoietic-Acute Radiation Syndrome. Int. J. Mol. Sci. 2024, 25, 4535. https://doi.org/10.3390/ijms25084535
Horseman T, Rittase WB, Slaven JE, Bradfield DT, Frank AM, Anderson JA, Hays EC, Ott AC, Thomas AE, Huppmann AR, et al. Ferroptosis, Inflammation, and Microbiome Alterations in the Intestine in the Göttingen Minipig Model of Hematopoietic-Acute Radiation Syndrome. International Journal of Molecular Sciences. 2024; 25(8):4535. https://doi.org/10.3390/ijms25084535
Chicago/Turabian StyleHorseman, Timothy, W. Bradley Rittase, John E. Slaven, Dmitry T. Bradfield, Andrew M. Frank, Joseph A. Anderson, Evelyn C. Hays, Andrew C. Ott, Anjali E. Thomas, Alison R. Huppmann, and et al. 2024. "Ferroptosis, Inflammation, and Microbiome Alterations in the Intestine in the Göttingen Minipig Model of Hematopoietic-Acute Radiation Syndrome" International Journal of Molecular Sciences 25, no. 8: 4535. https://doi.org/10.3390/ijms25084535
APA StyleHorseman, T., Rittase, W. B., Slaven, J. E., Bradfield, D. T., Frank, A. M., Anderson, J. A., Hays, E. C., Ott, A. C., Thomas, A. E., Huppmann, A. R., Lee, S. -H., Burmeister, D. M., & Day, R. M. (2024). Ferroptosis, Inflammation, and Microbiome Alterations in the Intestine in the Göttingen Minipig Model of Hematopoietic-Acute Radiation Syndrome. International Journal of Molecular Sciences, 25(8), 4535. https://doi.org/10.3390/ijms25084535