Figure 1.
Schematics and table showing cellular energy production systems under various conditions. (A), A schematic showing the different energy systems in the striatum with inhibitors to block the different energy production pathways. (B), A table showing how the different conditions affect the various energy production systems associated with evoked DA release. 2DG—2 deoxy-D-glucose, GLC—glucose, P—phosphate, ROT—rotenone, Lac—lactate, IA—iodoacetic Acid, Pyu—pyruvate, 4CIN—α-Cyano-4-hydroxycinnamic acid, Gly—glycolysis, MCT—monocarboxylate transport, OxPhos—oxidative phosphorylation.
Figure 1.
Schematics and table showing cellular energy production systems under various conditions. (A), A schematic showing the different energy systems in the striatum with inhibitors to block the different energy production pathways. (B), A table showing how the different conditions affect the various energy production systems associated with evoked DA release. 2DG—2 deoxy-D-glucose, GLC—glucose, P—phosphate, ROT—rotenone, Lac—lactate, IA—iodoacetic Acid, Pyu—pyruvate, 4CIN—α-Cyano-4-hydroxycinnamic acid, Gly—glycolysis, MCT—monocarboxylate transport, OxPhos—oxidative phosphorylation.
Figure 2.
Evoked DA release is higher in the dSTR than in the NAcc shell. Evoked DA release in the dSTR is significantly higher than that in the NAcc shell. (A), Representative FSCV traces of 1p-stimulation-evoked DA release recorded in the dSTR and the NAcc shell. (B), Plot showing average amplitudes of 1p-evoked DA release (dSTR, n = 8; NAcc shell, n = 9 NAcc shell, **** p < 0.0001, Student t-test). (C), Representative traces of 1p-evoked DA release over 40 min taken every 10 min under control conditions. (D), Normalized evoked DA release over 40 min measured every 2 min in the dSTR and the NAcc shell (dSTR, n = 6; NAcc shell, n = 6, n.s., two-way ANOVA with Bonferroni post hoc test).
Figure 2.
Evoked DA release is higher in the dSTR than in the NAcc shell. Evoked DA release in the dSTR is significantly higher than that in the NAcc shell. (A), Representative FSCV traces of 1p-stimulation-evoked DA release recorded in the dSTR and the NAcc shell. (B), Plot showing average amplitudes of 1p-evoked DA release (dSTR, n = 8; NAcc shell, n = 9 NAcc shell, **** p < 0.0001, Student t-test). (C), Representative traces of 1p-evoked DA release over 40 min taken every 10 min under control conditions. (D), Normalized evoked DA release over 40 min measured every 2 min in the dSTR and the NAcc shell (dSTR, n = 6; NAcc shell, n = 6, n.s., two-way ANOVA with Bonferroni post hoc test).
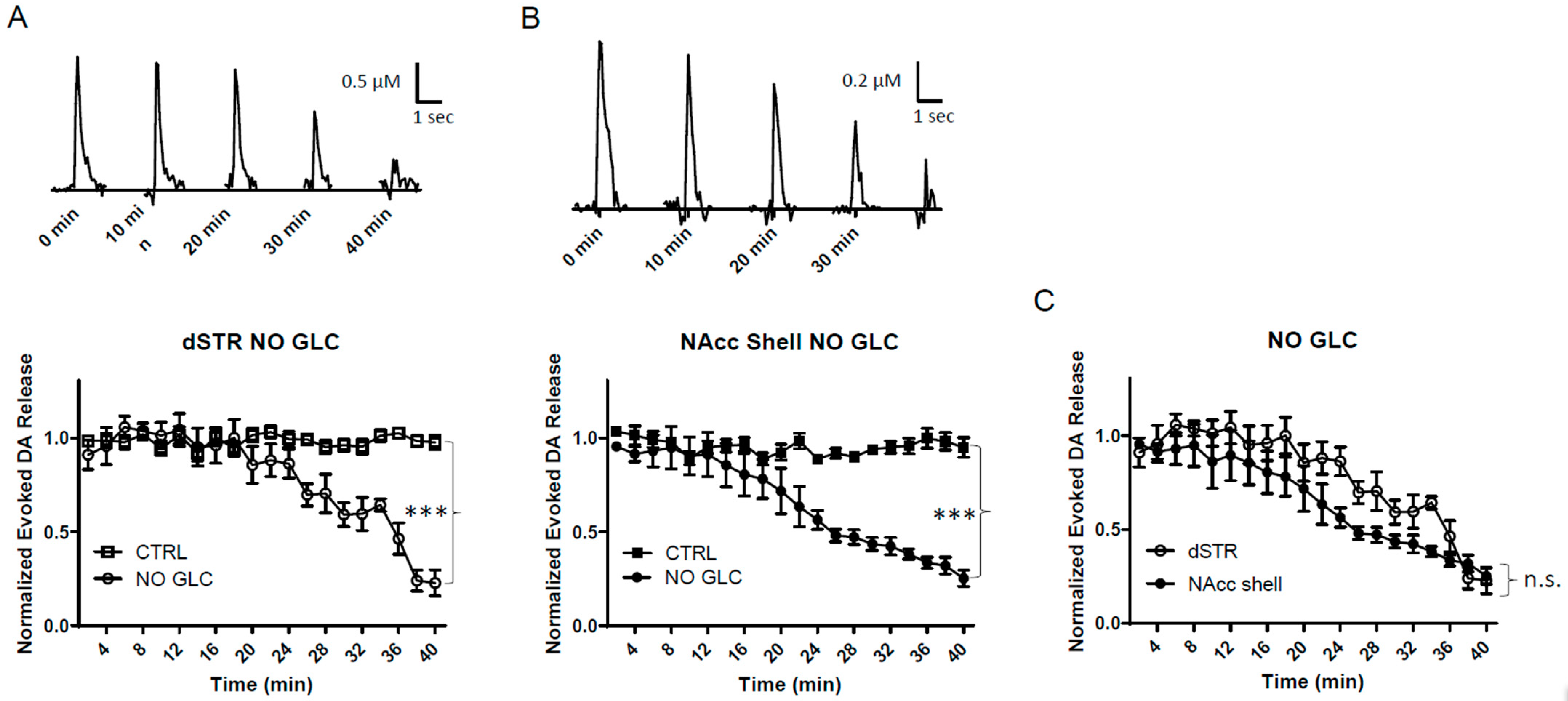
Figure 3.
Similar diminishment of evoked DA release in the dSTR and the NAcc shell under glucose-deprived conditions. (A), Representative traces of 1p-evoked DA release in the dSTR over 40 min taken every 10 min under glucose-deprived conditions (NO GLC, top) and plot of the normalized evoked DA release over 40 min measured every 2 min under glucose-deprived conditions (bottom, NO GLC, n = 6; control (CTRL), n = 6, NO GLC vs. CTRL, *** p < 0.001, two-way ANOVA with Bonferroni post hoc test). (B), Representative traces of 1p-evoked DA release in the NAcc shell over 40 min taken every 10 min under glucose-deprived conditions (top) and the plot of the normalized evoked DA release over 40 min measured every 2 min under glucose-deprived conditions (bottom, NO GLC, n = 6; CTRL, n = 6, NO GLC vs. CTRL, *** p < 0.001, two-way ANOVA with Bonferroni post hoc test). (C), Comparison of normalized evoked DA release in the dSTR and NAcc shell over 40 min under glucose-deprived conditions showing no statistical significance, dSTR vs. NAcc shell, n.s., two-way ANOVA with Bonferroni post hoc test.
Figure 3.
Similar diminishment of evoked DA release in the dSTR and the NAcc shell under glucose-deprived conditions. (A), Representative traces of 1p-evoked DA release in the dSTR over 40 min taken every 10 min under glucose-deprived conditions (NO GLC, top) and plot of the normalized evoked DA release over 40 min measured every 2 min under glucose-deprived conditions (bottom, NO GLC, n = 6; control (CTRL), n = 6, NO GLC vs. CTRL, *** p < 0.001, two-way ANOVA with Bonferroni post hoc test). (B), Representative traces of 1p-evoked DA release in the NAcc shell over 40 min taken every 10 min under glucose-deprived conditions (top) and the plot of the normalized evoked DA release over 40 min measured every 2 min under glucose-deprived conditions (bottom, NO GLC, n = 6; CTRL, n = 6, NO GLC vs. CTRL, *** p < 0.001, two-way ANOVA with Bonferroni post hoc test). (C), Comparison of normalized evoked DA release in the dSTR and NAcc shell over 40 min under glucose-deprived conditions showing no statistical significance, dSTR vs. NAcc shell, n.s., two-way ANOVA with Bonferroni post hoc test.
![Ijms 25 04580 g003]()
Figure 4.
Glycolysis inhibition diminishes evoked DA release in the NAcc shell more rapidly than in the dSTR. (A), Representative traces of 1p-evoked DA release in the dSTR over 40 min taken every 10 min when glycolysis was inhibited with 1 mM IA (top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, IA, n = 6; CTRL, n = 6, IA vs. CTRL, **** p < 0.0001, two-way ANOVA with Bonferroni post hoc test). (B), Representative traces of 1p-evoked DA release in the NAcc over 40 min taken every 10 min when glycolysis was inhibited with 1 mM IA (top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, IA, n = 6; CTRL, n = 6, IA vs. CTRL, **** p < 0.0001, two-way ANOVA with Bonferroni post hoc test). (C), Comparison of normalized evoked DA release in the dSTR and NAcc shell over 40 min under glycolysis inhibition conditions, dSTR vs. NAcc shell, ** p < 0.01, two-way ANOVA with Bonferroni post hoc test. (D), Comparison of normalized evoked DA release in the dSTR over 40 min under glycolysis inhibition and glucose-deprived conditions, showing no significant difference, IA, n = 6; NO GLC, n = 6, IA vs. NO GLC, ** p < 0.01, n.s., two-way ANOVA with Bonferroni post hoc test. (E), Comparison of normalized evoked DA release in the NAcc over 40 min under glycolysis inhibition and glucose-deprived conditions, showing glycolysis inhibition diminishes evoked DA release more rapidly than glucose-deprived conditions, IA, n = 6; NO GLC, n = 6, IA vs. NO GLC, ** p < 0.01, two-way ANOVA with Bonferroni post hoc test.
Figure 4.
Glycolysis inhibition diminishes evoked DA release in the NAcc shell more rapidly than in the dSTR. (A), Representative traces of 1p-evoked DA release in the dSTR over 40 min taken every 10 min when glycolysis was inhibited with 1 mM IA (top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, IA, n = 6; CTRL, n = 6, IA vs. CTRL, **** p < 0.0001, two-way ANOVA with Bonferroni post hoc test). (B), Representative traces of 1p-evoked DA release in the NAcc over 40 min taken every 10 min when glycolysis was inhibited with 1 mM IA (top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, IA, n = 6; CTRL, n = 6, IA vs. CTRL, **** p < 0.0001, two-way ANOVA with Bonferroni post hoc test). (C), Comparison of normalized evoked DA release in the dSTR and NAcc shell over 40 min under glycolysis inhibition conditions, dSTR vs. NAcc shell, ** p < 0.01, two-way ANOVA with Bonferroni post hoc test. (D), Comparison of normalized evoked DA release in the dSTR over 40 min under glycolysis inhibition and glucose-deprived conditions, showing no significant difference, IA, n = 6; NO GLC, n = 6, IA vs. NO GLC, ** p < 0.01, n.s., two-way ANOVA with Bonferroni post hoc test. (E), Comparison of normalized evoked DA release in the NAcc over 40 min under glycolysis inhibition and glucose-deprived conditions, showing glycolysis inhibition diminishes evoked DA release more rapidly than glucose-deprived conditions, IA, n = 6; NO GLC, n = 6, IA vs. NO GLC, ** p < 0.01, two-way ANOVA with Bonferroni post hoc test.
![Ijms 25 04580 g004]()
Figure 5.
Bypassing glycolysis diminishes evoked release of DA in the NAcc shell more than in the dSTR. (A), Representative traces of 1p-evoked DA release in the dSTR over 40 min taken every 10 min under when 10 mM pyruvate was administered under glucose-deprived conditions (PYU + NO GLC, top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, PYU + NO GLC, n = 5; CTRL; n = 6, PYU + NO GLC vs. CTRL, n.s., two-way ANOVA with Bonferroni post hoc test). (B), Representative traces of 1p-evoked DA release in the NAcc shell over 40 min taken every 10 min under glucose-deprived conditions (top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, PYU + NO GLC, n = 5; CTRL, n = 6, PYU + NO GLC vs. CTRL, **** p < 0.0001, two-way ANOVA with Bonferroni post hoc test). (C), Comparison of normalized evoked DA release in the dSTR and NAcc shell over 40 min under condition bypassing glycolysis, dSTR vs. NAcc shell, **** p < 0.0001, two-way ANOVA with Bonferroni post hoc test.
Figure 5.
Bypassing glycolysis diminishes evoked release of DA in the NAcc shell more than in the dSTR. (A), Representative traces of 1p-evoked DA release in the dSTR over 40 min taken every 10 min under when 10 mM pyruvate was administered under glucose-deprived conditions (PYU + NO GLC, top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, PYU + NO GLC, n = 5; CTRL; n = 6, PYU + NO GLC vs. CTRL, n.s., two-way ANOVA with Bonferroni post hoc test). (B), Representative traces of 1p-evoked DA release in the NAcc shell over 40 min taken every 10 min under glucose-deprived conditions (top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, PYU + NO GLC, n = 5; CTRL, n = 6, PYU + NO GLC vs. CTRL, **** p < 0.0001, two-way ANOVA with Bonferroni post hoc test). (C), Comparison of normalized evoked DA release in the dSTR and NAcc shell over 40 min under condition bypassing glycolysis, dSTR vs. NAcc shell, **** p < 0.0001, two-way ANOVA with Bonferroni post hoc test.
![Ijms 25 04580 g005]()
Figure 6.
Specific inhibition of OxPhos diminishes evoked release of DA in the dSTR more than the NAcc shell. (A), Representative traces of 1p-evoked DA release in the dSTR over 40 min taken every 10 min when complex I inhibited with 10 µM rotenone (ROT, top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, ROT, n = 5; CTRL, n = 6, ROT vs. CTRL, **** p < 0.0001, two-way ANOVA with Bonferroni post hoc test). (B), Representative traces of 1p-evoked DA release in the NAcc shell over 40 min taken every 10 min when complex I inhibited with 10 µM ROT (top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, ROT, n = 5; CTRL, n = 6, ROT vs. CTRL, * p < 0.05, two-way ANOVA with Bonferroni post hoc test). (C), Comparison of normalized evoked DA release in the dSTR and NAcc shell over 40 min under specific inhibition of OxPhos, dSTR vs. NAcc shell, * p < 0.05, two-way ANOVA with Bonferroni post hoc test.
Figure 6.
Specific inhibition of OxPhos diminishes evoked release of DA in the dSTR more than the NAcc shell. (A), Representative traces of 1p-evoked DA release in the dSTR over 40 min taken every 10 min when complex I inhibited with 10 µM rotenone (ROT, top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, ROT, n = 5; CTRL, n = 6, ROT vs. CTRL, **** p < 0.0001, two-way ANOVA with Bonferroni post hoc test). (B), Representative traces of 1p-evoked DA release in the NAcc shell over 40 min taken every 10 min when complex I inhibited with 10 µM ROT (top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, ROT, n = 5; CTRL, n = 6, ROT vs. CTRL, * p < 0.05, two-way ANOVA with Bonferroni post hoc test). (C), Comparison of normalized evoked DA release in the dSTR and NAcc shell over 40 min under specific inhibition of OxPhos, dSTR vs. NAcc shell, * p < 0.05, two-way ANOVA with Bonferroni post hoc test.
Figure 7.
MCT inhibition reduces evoked DA release in the dSTR but not in the NAcc shell. (A), Representative traces of 1p-evoked DA release in the dSTR over 40 min taken every 10 min when MCT inhibited with 100 µM 4CIN (top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, 4CIN, n = 6; CTRL, n = 6, 4CIN vs. CTRL, ** p < 0.01, two-way ANOVA with Bonferroni post hoc test). (B), Representative traces of 1p-evoked DA release in the NAcc shell over 40 min taken every 10 min MCT inhibited with 100 µM 4CIN (top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, 4CIN, n = 6; CTRL, n = 6, 4CIN vs. CTRL, n.s., two-way ANOVA with Bonferroni post hoc test). (C), Comparison of normalized evoked DA release in the dSTR and NAcc shell over 40 min under inhibition of MCT, dSTR vs. NAcc shell, * p < 0.05, two-way ANOVA with Bonferroni post hoc test.
Figure 7.
MCT inhibition reduces evoked DA release in the dSTR but not in the NAcc shell. (A), Representative traces of 1p-evoked DA release in the dSTR over 40 min taken every 10 min when MCT inhibited with 100 µM 4CIN (top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, 4CIN, n = 6; CTRL, n = 6, 4CIN vs. CTRL, ** p < 0.01, two-way ANOVA with Bonferroni post hoc test). (B), Representative traces of 1p-evoked DA release in the NAcc shell over 40 min taken every 10 min MCT inhibited with 100 µM 4CIN (top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, 4CIN, n = 6; CTRL, n = 6, 4CIN vs. CTRL, n.s., two-way ANOVA with Bonferroni post hoc test). (C), Comparison of normalized evoked DA release in the dSTR and NAcc shell over 40 min under inhibition of MCT, dSTR vs. NAcc shell, * p < 0.05, two-way ANOVA with Bonferroni post hoc test.
Figure 8.
MCT inhibition under glucose-deprived conditions reduces evoked DA release in the striatum more rapidly compared to glucose-deprived conditions. (A), Representative traces of 1p-evoked DA release in the dSTR over 40 min taken every 10 min when MCT inhibited with 100 µM 4CIN under glucose-deprived conditions (top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, 4CIN + NO GLC, n = 6; NO GLC, n = 6, 4CIN + NO GLC vs. NO GLC, ** p < 0.01, two-way ANOVA with Bonferroni post hoc test). (B), Representative traces of 1p-evoked DA release in the dSTR over 40 min taken every 10 min when MCT inhibited with 100 µM 4CIN under glucose-deprived conditions (top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, 4CIN + NO GLC, n = 6; NO GLC, n = 6, 4CIN + NO GLC vs. NO GLC, * p < 0.05, two-way ANOVA with Bonferroni post hoc test). (C), Comparison of normalized evoked DA release in the dSTR and NAcc shell over 40 min under MCT inhibition under glucose-deprived conditions, dSTR vs. NAcc shell, n.s., two-way ANOVA with Bonferroni post hoc test.
Figure 8.
MCT inhibition under glucose-deprived conditions reduces evoked DA release in the striatum more rapidly compared to glucose-deprived conditions. (A), Representative traces of 1p-evoked DA release in the dSTR over 40 min taken every 10 min when MCT inhibited with 100 µM 4CIN under glucose-deprived conditions (top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, 4CIN + NO GLC, n = 6; NO GLC, n = 6, 4CIN + NO GLC vs. NO GLC, ** p < 0.01, two-way ANOVA with Bonferroni post hoc test). (B), Representative traces of 1p-evoked DA release in the dSTR over 40 min taken every 10 min when MCT inhibited with 100 µM 4CIN under glucose-deprived conditions (top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, 4CIN + NO GLC, n = 6; NO GLC, n = 6, 4CIN + NO GLC vs. NO GLC, * p < 0.05, two-way ANOVA with Bonferroni post hoc test). (C), Comparison of normalized evoked DA release in the dSTR and NAcc shell over 40 min under MCT inhibition under glucose-deprived conditions, dSTR vs. NAcc shell, n.s., two-way ANOVA with Bonferroni post hoc test.
![Ijms 25 04580 g008]()
Figure 9.
Energy production via the MCT system can maintain evoked DA release in the dSTR but not in the NAcc shell. (A), Representative traces of 1p-evoked DA release in the dSTR over 40 min taken every 10 min when 10 mM lactate was administered under glucose-deprived condition (Lac + NO GLC, top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, Lac + NO GLC, n = 6; CTRL, n = 6, Lac + NO GLC vs. CTRL, n.s., two-way ANOVA with Bonferroni post hoc test). (B), Representative traces of 1p-evoked DA release in the NAcc shell over 40 min taken every 10 min when 10 mM lactate was administered under glucose-deprived condition (Lac + NO GLC, top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, Lac + NO GLC, n = 6; CTRL, n = 6, Lac + NO GLC vs. CTRL, **** p < 0.0001, two-way ANOVA with Bonferroni post hoc test). (C), Comparison of normalized evoked DA release in the dSTR and NAcc shell over 40 min when 10 mM lactate was administered under glucose-deprived condition, dSTR vs. NAcc shell, *** p < 0.001, two-way ANOVA with Bonferroni post hoc test.
Figure 9.
Energy production via the MCT system can maintain evoked DA release in the dSTR but not in the NAcc shell. (A), Representative traces of 1p-evoked DA release in the dSTR over 40 min taken every 10 min when 10 mM lactate was administered under glucose-deprived condition (Lac + NO GLC, top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, Lac + NO GLC, n = 6; CTRL, n = 6, Lac + NO GLC vs. CTRL, n.s., two-way ANOVA with Bonferroni post hoc test). (B), Representative traces of 1p-evoked DA release in the NAcc shell over 40 min taken every 10 min when 10 mM lactate was administered under glucose-deprived condition (Lac + NO GLC, top) and a plot of the normalized evoked DA release over 40 min measured every 2 min (bottom, Lac + NO GLC, n = 6; CTRL, n = 6, Lac + NO GLC vs. CTRL, **** p < 0.0001, two-way ANOVA with Bonferroni post hoc test). (C), Comparison of normalized evoked DA release in the dSTR and NAcc shell over 40 min when 10 mM lactate was administered under glucose-deprived condition, dSTR vs. NAcc shell, *** p < 0.001, two-way ANOVA with Bonferroni post hoc test.
![Ijms 25 04580 g009]()
Figure 10.
Oxidation level is higher in the dSTR than in the NAcc shell. (A), Representative images of roEGFP signals in the dSTR and NAcc excited at 900 nm and 800 nm. (B), Plot of average oxidation level (800/900) in the dSTR (11 slices) and the NAcc (11 slices) from four mice aged 4–7 months old, dSTR vs. NAcc, **** p < 0.0001, Student t-test.
Figure 10.
Oxidation level is higher in the dSTR than in the NAcc shell. (A), Representative images of roEGFP signals in the dSTR and NAcc excited at 900 nm and 800 nm. (B), Plot of average oxidation level (800/900) in the dSTR (11 slices) and the NAcc (11 slices) from four mice aged 4–7 months old, dSTR vs. NAcc, **** p < 0.0001, Student t-test.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









