
Exogenous Melatonin Enhances Dihydrochalcone Accumulation in Lithocarpus litseifolius Leaves via Regulating Hormonal Crosstalk and Transcriptional Profiling
 ,
, 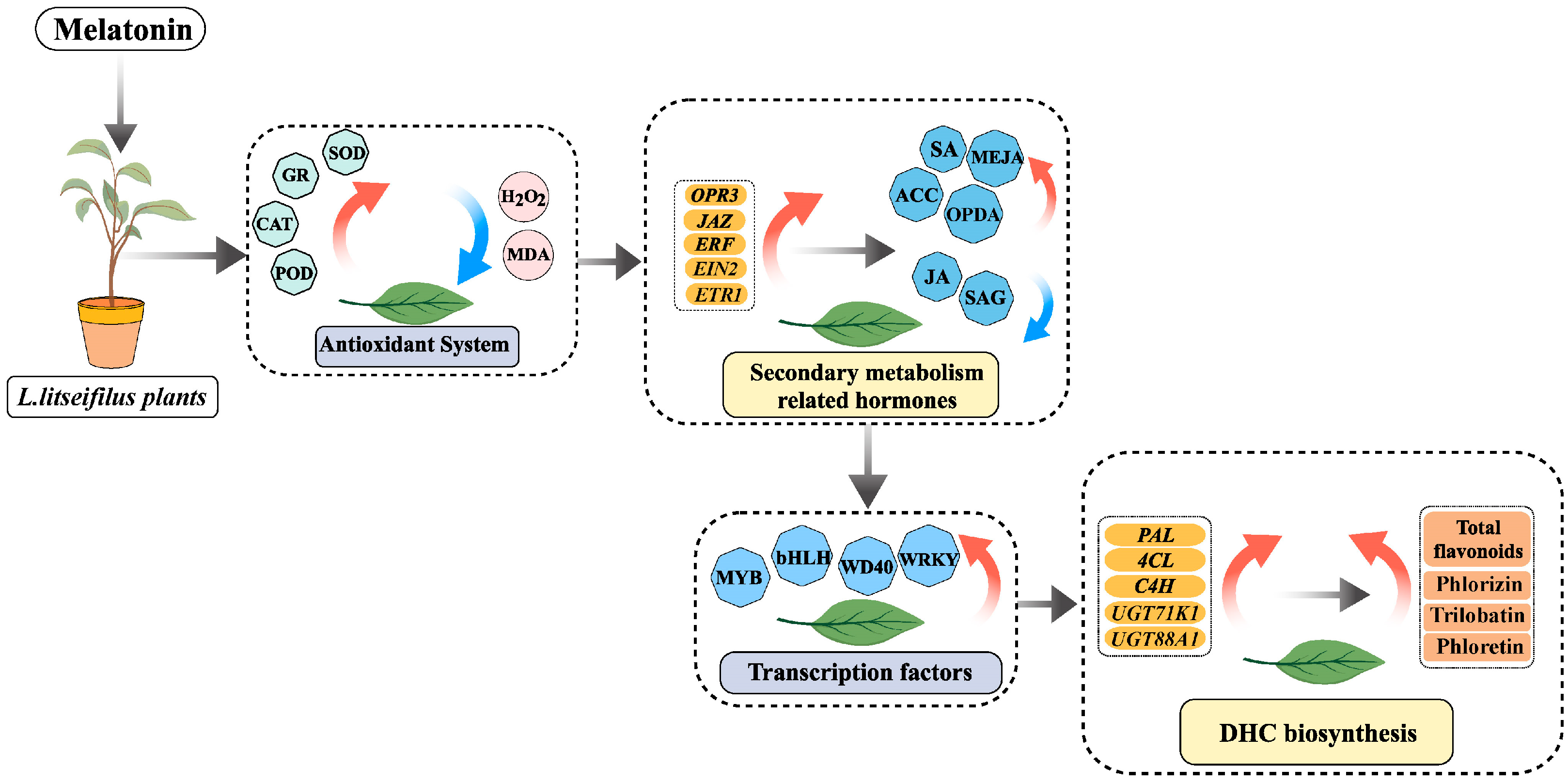{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
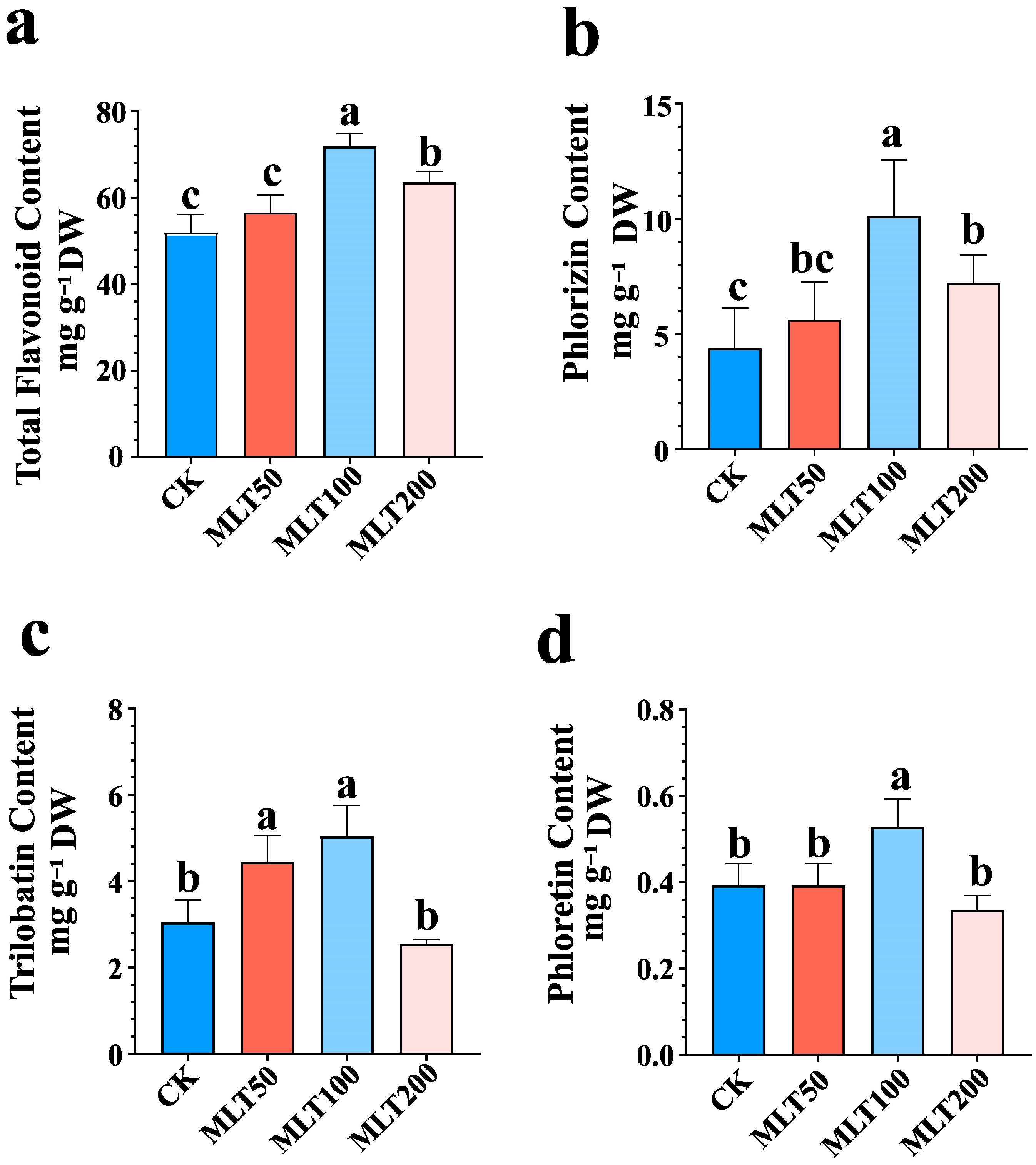
2.1. Effect of MLT Treatment on DHC Biosynthesis
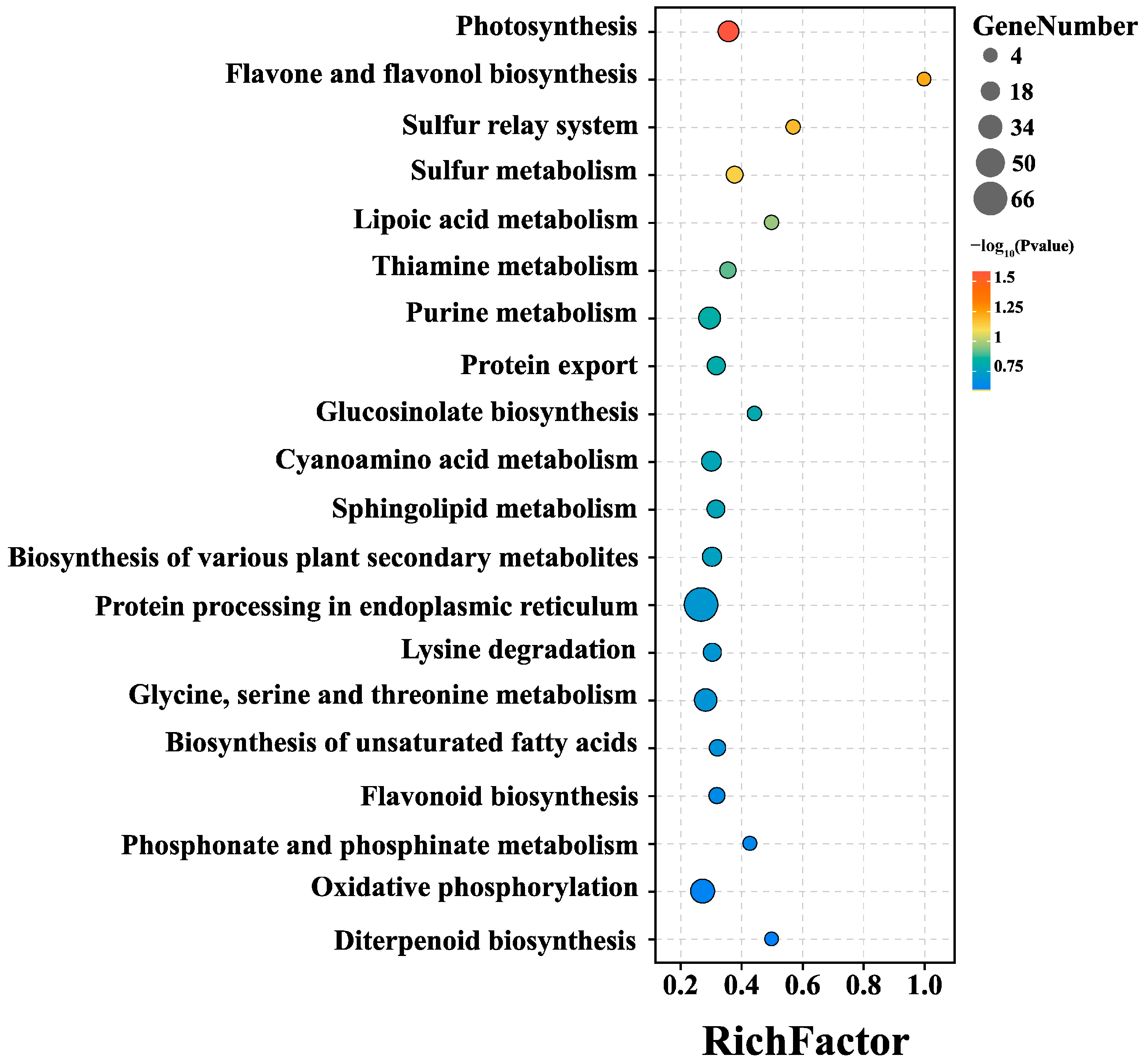
2.2. Transcriptome Sequencing and DEG Pathway Analysis
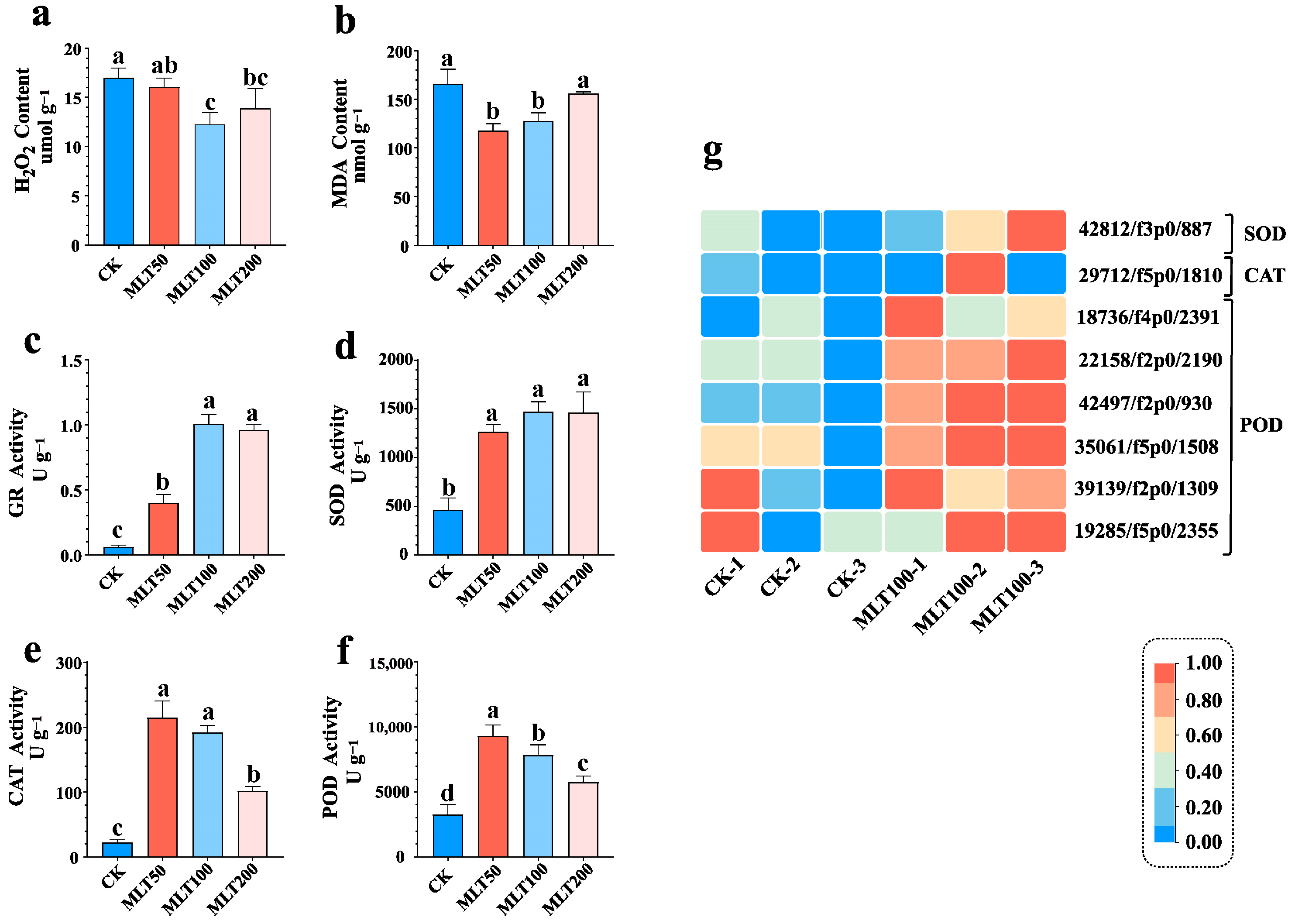
2.3. Effects of MLT on the Antioxidant System and Related Gene Expression
2.4. Effects of MLT on Phytohormone Crosstalks and Related Gene Expression
2.5. Effects of MLT on TF Families
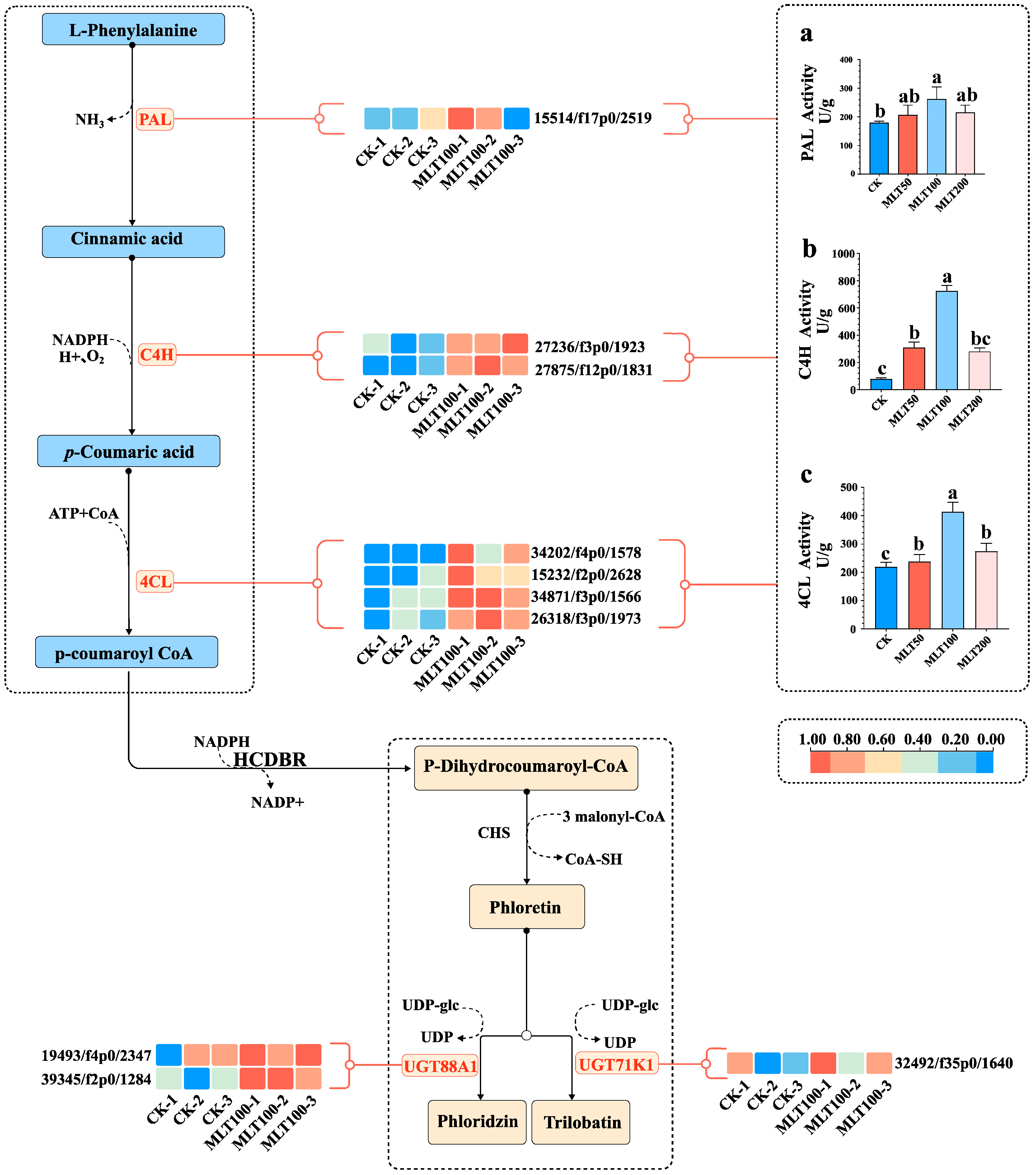
2.6. Effects of MLT on DHC Biosynthesis and Related Gene Expression
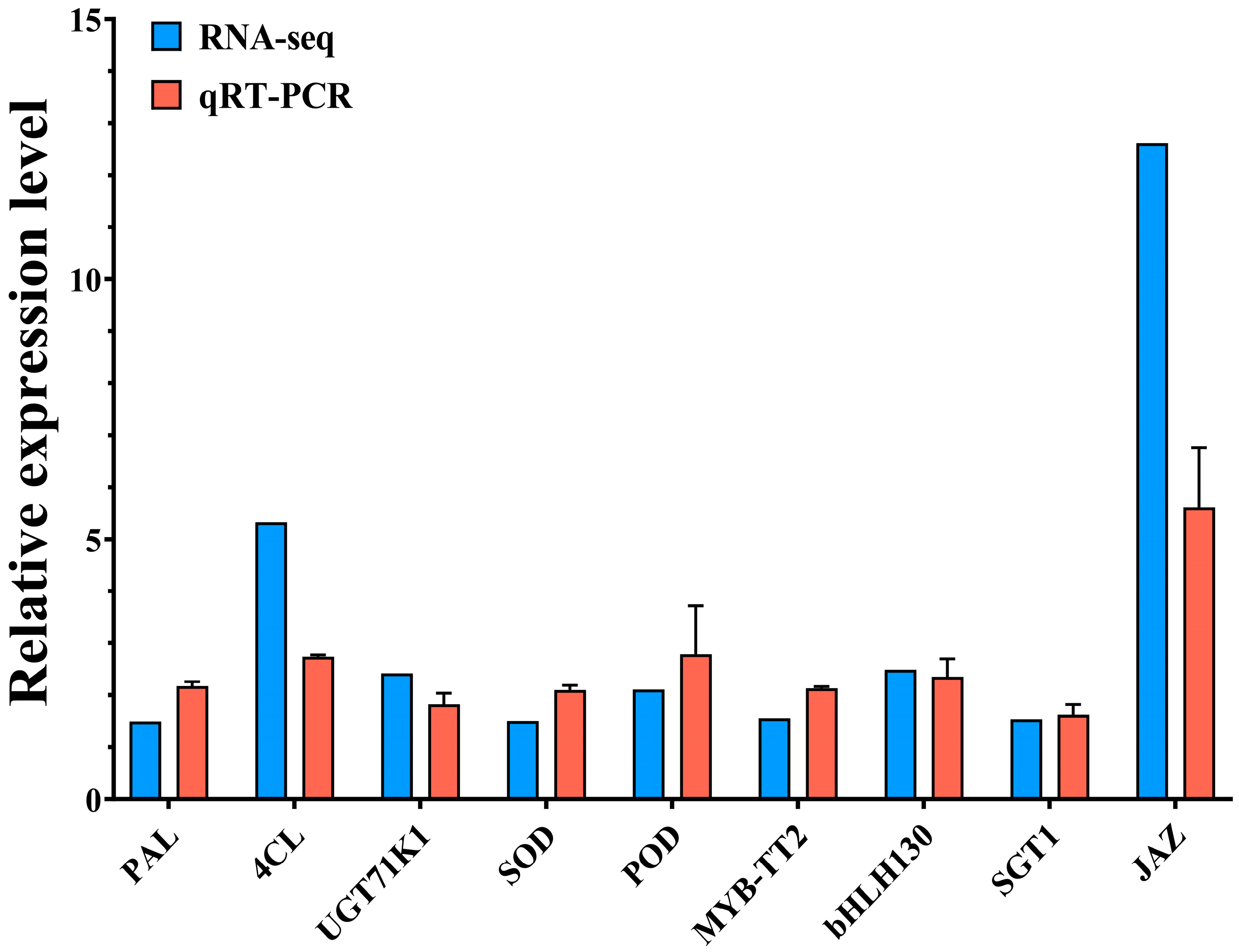
2.7. Quantitative Real-Time PCR (qRT–PCR) Validation of DEGs
2.8. Correlation Analysis
3. Discussion
3.1. MLT Elicitation of DHC Levels
3.2. MLT Triggers the Antioxidant Capacity in DHC Accumulation
3.3. MLT Modulates Phytohormone Crosstalks in DHC Accumulation
3.4. MLT Regulates Transcriptional Profiles in DHC Biosynthesis
4. Materials and Methods
4.1. Plant Material
4.2. MLT Elicitation Treatment
4.3. DHC Extraction and Quantification
4.4. Determination of H2O2 and MDA Levels and Antioxidant Enzyme Activities
4.5. Determination of Phytohormone Levels
4.6. Determination of Flavonoid Biosynthesis-Related Enzyme Activities
4.7. Transcriptome Sequencing Analysis
4.7.1. RNA Extraction, Illumina Sequencing, and Functional Annotation
4.7.2. DEGs and Pathway Enrichment Analysis
4.7.3. qRT–PCR Validation of DEGs
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Shang, A.; Liu, H.-Y.; Luo, M.; Xia, Y.; Yang, X.; Li, H.-Y.; Wu, D.-T.; Sun, Q.; Geng, F.; Gan, R.-Y. Sweet Tea (Lithocarpus polystachyus Rehd.) as a New Natural Source of Bioactive Dihydrochalcones with Multiple Health Benefits. Crit. Rev. Food Sci. Nutr. 2022, 62, 917–934. [Google Scholar] [CrossRef] [PubMed]
- Ibdah, M.; Martens, S.; Gang, D.R. Biosynthetic Pathway and Metabolic Engineering of Plant Dihydrochalcones. J. Agric. Food Chem. 2018, 66, 2273–2280. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.M. SGLT2 Inhibitors: Physiology and Pharmacology. Kidney360 2021, 2, 2027–2037. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yauk, Y.-K.; Zhao, Q.; Hamiaux, C.; Xiao, Z.; Gunaseelan, K.; Zhang, L.; Tomes, S.; López-Girona, E.; Cooney, J.; et al. Biosynthesis of the Dihydrochalcone Sweetener Trilobatin Requires Phloretin Glycosyltransferase2. Plant Physiol. 2020, 184, 738–752. [Google Scholar] [CrossRef]
- Yang, J.; Huang, Y.; Yang, Z.; Zhou, C.; Hu, X. Identification and Quantitative Evaluation of Major Sweet Ingredients in Sweet Tea (Lithocarpus Polystachyus Rehd.) Based upon Location, Harvesting Time, Leaf Age. J. Chem. Soc. Pak. 2018, 40, 158–164. [Google Scholar]
- Ramirez-Estrada, K.; vidal-Limon, H.; Hidalgo, D.; Moyano, E.; Golenioswki, M.; Cusidó, R.; Palazon, J. Elicitation, an Effective Strategy for the Biotechnological Production of Bioactive High-Added Value Compounds in Plant Cell Factories. Molecules 2016, 21, 182. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: A New Plant Hormone and/or a Plant Master Regulator? Trends Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Tian, X.; He, X.; Yang, J.; Yang, Z.; Fang, W. Exogenous Melatonin Stimulated Isoflavone Biosynthesis in NaCl-Stressed Germinating Soybean (Glycine max L.). Plant Physiol. Biochem. 2022, 185, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tian, J.; Wang, S.; Song, T.; Zhang, J.; Yao, Y. Application of Melatonin Promotes Anthocyanin Accumulation in Crabapple Leaves. Plant Physiol. Biochem. 2019, 142, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Xie, Y.; Zhang, Z.; Chen, L. Melatonin: A Multifunctional Factor in Plants. Int. J. Mol. Sci. 2018, 19, 1528. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Xiao, Q.; Chen, Z.; Han, Y. Crosstalk between Melatonin and Reactive Oxygen Species in Plant Abiotic Stress Responses: An Update. Int. J. Mol. Sci. 2022, 23, 5666. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Sun, Q.; Li, H.; Li, X.; Cao, Y.; Zhang, H.; Li, S.; Zhang, L.; Qi, Y.; Ren, S.; et al. Melatonin Improved Anthocyanin Accumulation by Regulating Gene Expressions and Resulted in High Reactive Oxygen Species Scavenging Capacity in Cabbage. Front. Plant Sci. 2016, 7, 197. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin and Its Relationship to Plant Hormones. Ann. Bot-Lond. 2018, 121, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Xu, X.; Li, L.; Sun, Q.; Wang, Q.; Huang, H.; Tong, Z.; Zhang, J. Melatonin-Mediated Development and Abiotic Stress Tolerance in Plants. Front. Plant Sci. 2023, 14, 1100827. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Yang, M.; Wu, X.; Wang, Y.; Zhang, R. Physiological and Transcriptomic Analyses of the Effects of Exogenous Melatonin on Drought Tolerance in Maize (Zea mays L.). Plant Physiol. Biochem. 2021, 168, 128–142. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, A.; Brockman, A.; Aguirre, L.; Campbell, A.; Bean, A.; Cantero, A.; Gonzalez, A. Advances in the MYB–bHLH–WD Repeat (MBW) Pigment Regulatory Model: Addition of a WRKY Factor and Co-Option of an Anthocyanin MYB for Betalain Regulation. Plant Cell Physiol. 2017, 58, 1431–1441. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Fang, H.; Chen, Z.; Liu, Z.; Yu, X.; Liang, C. Ectopic Expression of a R2R3-MYB Transcription Factor Gene LjaMYB12 from Lonicera japonica Increases Flavonoid Accumulation in Arabidopsis Thaliana. Int. J. Mol. Sci. 2019, 20, 4494. [Google Scholar] [CrossRef] [PubMed]
- Kajla, M.; Roy, A.; Singh, I.K.; Singh, A. Regulation of the Regulators: Transcription Factors Controlling Biosynthesis of Plant Secondary Metabolites during Biotic Stresses and Their Regulation by miRNAs. Front. Plant Sci. 2023, 14, 1126567. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The Flavonoid Biosynthesis Network in Plants. Int. J. Mol. Sci. 2021, 22, 12824. [Google Scholar] [CrossRef] [PubMed]
- Gosch, C.; Halbwirth, H.; Schneider, B.; Hölscher, D.; Stich, K. Cloning and Heterologous Expression of Glycosyltransferases from Malus x domestica and Pyrus communis, Which Convert Phloretin to Phloretin 2′-O-Glucoside (Phloridzin). Plant Sci. 2010, 178, 299–306. [Google Scholar] [CrossRef]
- Han, Z.; Zhang, J.; Cai, S.; Chen, X.; Quan, X.; Zhang, G. Association Mapping for Total Polyphenol Content, Total Flavonoid Content and Antioxidant Activity in Barley. BMC Genom. 2018, 19, 81. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lin, L.; Long, Y.; Guo, H.; Wang, Z.; Cui, M.; Huang, J.; Xing, Z. Comprehensive Transcriptome Analysis Revealed the Effects of the Light Quality, Light Intensity, and Photoperiod on Phlorizin Accumulation in Lithocarpus polystachyus Rehd. Forests 2019, 10, 995. [Google Scholar] [CrossRef]
- Baek, G.; Lee, H.; Ko, J.; Choi, H.-K. Exogenous Melatonin Enhances the Growth and Production of Bioactive Metabolites in Lemna aequinoctialis Culture by Modulating Metabolic and Lipidomic Profiles. BMC Plant Biol. 2022, 22, 545. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Wang, L.; Belwal, T.; Zhang, X.; Lu, H.; Chen, C.; Li, L. Exogenous Melatonin and Abscisic Acid Expedite the Flavonoids Biosynthesis in Grape Berry of Vitis vinifera Cv. Kyoho. Molecules 2019, 25, 12. [Google Scholar] [CrossRef] [PubMed]
- Chang, Q.; Zhang, L.; Chen, S.; Gong, M.; Liu, L.; Hou, X.; Mi, Y.; Wang, X.; Wang, J.; Zhang, Y.; et al. Exogenous Melatonin Enhances the Yield and Secondary Metabolite Contents of Prunella vulgaris by Modulating Antioxidant System, Root Architecture and Photosynthetic Capacity. Plants 2023, 12, 1129. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gao, X.; Wang, X.; Song, W.; Wang, Q.; Wang, X.; Li, S.; Fu, B. Exogenous Melatonin Ameliorates Drought Stress in Agropyron Mongolicum by Regulating Flavonoid Biosynthesis and Carbohydrate Metabolism. Front. Plant Sci. 2022, 13, 1051165. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Wang, J.; Xu, D.; Tao, S.; Chong, S.; Yan, D.; Li, Z.; Yuan, H.; Zheng, B. Melatonin Regulates the Functional Components of Photosynthesis, Antioxidant System, Gene Expression, and Metabolic Pathways to Induce Drought Resistance in Grafted Carya cathayensis Plants. Sci. Total Environ. 2020, 713, 136675. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin as a Regulatory Hub of Plant Hormone Levels and Action in Stress Situations. Plant Biol. 2021, 23, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ruiz, J.; Arnao, M. Relationship of Melatonin and Salicylic Acid in Biotic/Abiotic Plant Stress Responses. Agronomy 2018, 8, 33. [Google Scholar] [CrossRef]
- Sarrou, E.; Chatzopoulou, P.; Dimassi-Theriou, K.; Therios, I.; Koularmani, A. Effect of Melatonin, Salicylic Acid and Gibberellic Acid on Leaf Essential Oil and Other Secondary Metabolites of Bitter Orange Young Seedlings. J. Essent. Oil Res. 2015, 27, 487–496. [Google Scholar] [CrossRef]
- Arabia, A.; Munné-Bosch, S.; Muñoz, P. Melatonin Triggers Tissue-Specific Changes in Anthocyanin and Hormonal Contents during Postharvest Decay of Angeleno Plums. Plant Sci. 2022, 320, 111287. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, L.; Morreel, K.; De Witte, E.; Lammertyn, F.; Van Montagu, M.; Boerjan, W.; Inzé, D.; Goossens, A. Mapping Methyl Jasmonate-Mediated Transcriptional Reprogramming of Metabolism and Cell Cycle Progression in Cultured Arabidopsis Cells. Proc. Natl. Acad. Sci. USA 2008, 105, 1380–1385. [Google Scholar] [CrossRef] [PubMed]
- Buer, C.S.; Sukumar, P.; Muday, G.K. Ethylene Modulates Flavonoid Accumulation and Gravitropic Responses in Roots of Arabidopsis. Plant Physiol. 2006, 140, 1384–1396. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yue, Q.; Bian, F.; Sun, H.; Zhai, H.; Yao, Y. Melatonin Enhances Phenolics Accumulation Partially via Ethylene Signaling and Resulted in High Antioxidant Capacity in Grape Berries. Front. Plant Sci. 2017, 8, 1426. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Chen, L.; Xie, Z.M.; Peng, X. Combined Transcriptome and Metabolome Analysis Revealed Pathways Involved in Improved Salt Tolerance of Gossypium hirsutum L. Seedlings in Response to Exogenous Melatonin Application. BMC Plant Biol. 2022, 22, 552. [Google Scholar] [CrossRef] [PubMed]
- Kajal; Ojha, R.; Lohani, P.; Deshmukh, R.; Salvi, P. Engineering the Transcriptional Regulatory Network to Improve Abiotic Stress Tolerance in Crop Plants: Taming the Tough Time. J. Plant Growth Regul. 2023, 43, 25–37. [Google Scholar]
- Wu, M.; Liu, J.; Song, L.; Li, X.; Cong, L.; Yue, R.; Yang, C.; Liu, Z.; Xu, L.; Wang, Z. Differences among the Anthocyanin Accumulation Patterns and Related Gene Expression Levels in Red Pears. Plants 2019, 8, 100. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Peng, J.; Wu, J.; Zhou, P.; He, R.; Allan, A.C.; Zeng, L. NtbHLH1, a JAF13-like bHLH, Interacts with NtMYB6 to Enhance Proanthocyanidin Accumulation in Chinese Narcissus. BMC Plant Biol. 2021, 21, 275. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hou, H.; Jiang, X.; Wang, P.; Dai, X.; Chen, W.; Gao, L.; Xia, T. A WD40 Repeat Protein from Camellia sinensis Regulates Anthocyanin and Proanthocyanidin Accumulation through the Formation of MYB–bHLH–WD40 Ternary Complexes. Int. J. Mol. Sci. 2018, 19, 1686. [Google Scholar] [CrossRef]
- Li, L.; Liu, Q.; Liu, T.; Cui, X.; Ning, W. Expression of Putative Luteolin Biosynthesis Genes and WRKY Transcription Factors in Taraxacum antungense Kitag. Plant. Cell Tiss. Organ. Cult. 2021, 145, 649–665. [Google Scholar] [CrossRef]
- Wang, J.; Cao, K.; Wang, L.; Dong, W.; Zhang, X.; Liu, W. Two MYB and Three bHLH Family Genes Participate in Anthocyanin Accumulation in the Flesh of Peach Fruit Treated with Glucose, Sucrose, Sorbitol, and Fructose In Vitro. Plants 2022, 11, 507. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Wang, C.; Han, X.; Tang, S.; Liu, S.; Xia, X.; Yin, W. A Novel bHLH Transcription Factor PebHLH35 from Populus Euphratica Confers Drought Tolerance through Regulating Stomatal Development, Photosynthesis and Growth in Arabidopsis. Biochem. Biophys. Res. Commun. 2014, 450, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Wan, D.; Chen, Y.; Wang, J. Determination of Total Flavonoids in Three Sedum Crude Drugs by UV-Vis Spectrophotometry. Pharmacogn. Mag. 2010, 6, 259. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Tuo, Y.; Zhang, Y.; Deng, Q.; Shi, C.; Chen, X.; Zhang, X. Evaluation of Two Parts of Lithocarpus polystachyus Rehd. from Different Chinese Areas by Multicomponent Content Determination and Pattern Recognition. J. Anal. Methods Chem. 2020, 2020, 8837526. [Google Scholar] [CrossRef] [PubMed]
- Niu, Q.; Zong, Y.; Qian, M.; Yang, F.; Teng, Y. Simultaneous Quantitative Determination of Major Plant Hormones in Pear Flowers and Fruit by UPLC/ESI-MS/MS. Anal. Methods 2014, 6, 1766–1773. [Google Scholar] [CrossRef]
- Cui, K.; Lin, Y.; Zhou, X.; Li, S.; Liu, H.; Zeng, F.; Zhu, F.; Ouyang, G.; Zeng, Z. Comparison of Sample Pretreatment Methods for the Determination of Multiple Phytohormones in Plant Samples by Liquid Chromatography–Electrospray Ionization-Tandem Mass Spectrometry. Microchem. J. 2015, 121, 25–31. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Yi, Z.; Chen, J.; Sun, H.; Rosli, H.G.; Pombo, M.A.; Zhang, P.; Banf, M.; Dai, X.; Martin, G.B.; Giovannoni, J.J.; et al. iTAK: A Program for Genome-Wide Prediction and Classification of Plant Transcription Factors, Transcriptional Regulators, and Protein Kinases. Mol. Plant 2016, 9, 1667–1670. [Google Scholar]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinf. 2011, 12, 323. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Chen, M.; Huang, X.; Zhang, G.; Zeng, L.; Zhang, G.; Wu, S.; Wang, Y. SRplot: A Free Online Platform for Data Visualization and Graphing. PLoS ONE 2023, 18, e0294236. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Wang, Y.; Yan, Y.; Wang, H. Transcriptome Analysis of the Development of Pedicel Abscission Zone in Tomato. Horticulturae 2022, 8, 865. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Sun, Y.; Wang, H.; Xu, M.; He, C.; Wang, C.; Yu, Y.; Zhang, Z.; Su, L. Exogenous Melatonin Enhances Dihydrochalcone Accumulation in Lithocarpus litseifolius Leaves via Regulating Hormonal Crosstalk and Transcriptional Profiling. Int. J. Mol. Sci. 2024, 25, 4592. https://doi.org/10.3390/ijms25094592
Zhang W, Sun Y, Wang H, Xu M, He C, Wang C, Yu Y, Zhang Z, Su L. Exogenous Melatonin Enhances Dihydrochalcone Accumulation in Lithocarpus litseifolius Leaves via Regulating Hormonal Crosstalk and Transcriptional Profiling. International Journal of Molecular Sciences. 2024; 25(9):4592. https://doi.org/10.3390/ijms25094592
Chicago/Turabian StyleZhang, Wenlong, Yuqi Sun, Hongfeng Wang, Mingfeng Xu, Chunmei He, Congcong Wang, Yongli Yu, Zongshen Zhang, and Lingye Su. 2024. "Exogenous Melatonin Enhances Dihydrochalcone Accumulation in Lithocarpus litseifolius Leaves via Regulating Hormonal Crosstalk and Transcriptional Profiling" International Journal of Molecular Sciences 25, no. 9: 4592. https://doi.org/10.3390/ijms25094592
APA StyleZhang, W., Sun, Y., Wang, H., Xu, M., He, C., Wang, C., Yu, Y., Zhang, Z., & Su, L. (2024). Exogenous Melatonin Enhances Dihydrochalcone Accumulation in Lithocarpus litseifolius Leaves via Regulating Hormonal Crosstalk and Transcriptional Profiling. International Journal of Molecular Sciences, 25(9), 4592. https://doi.org/10.3390/ijms25094592