
The Importance and Essentiality of Natural and Synthetic Chelators in Medicine: Increased Prospects for the Effective Treatment of Iron Overload and Iron Deficiency


Abstract
:1. Introduction
2. Iron Metabolism and Iron Imbalance
2.1. Iron Metabolism and Regulation of Iron in Humans
2.2. Iron Distribution and Iron Balance
2.3. Genetic and Other Diseases of Iron Imbalance and Distribution
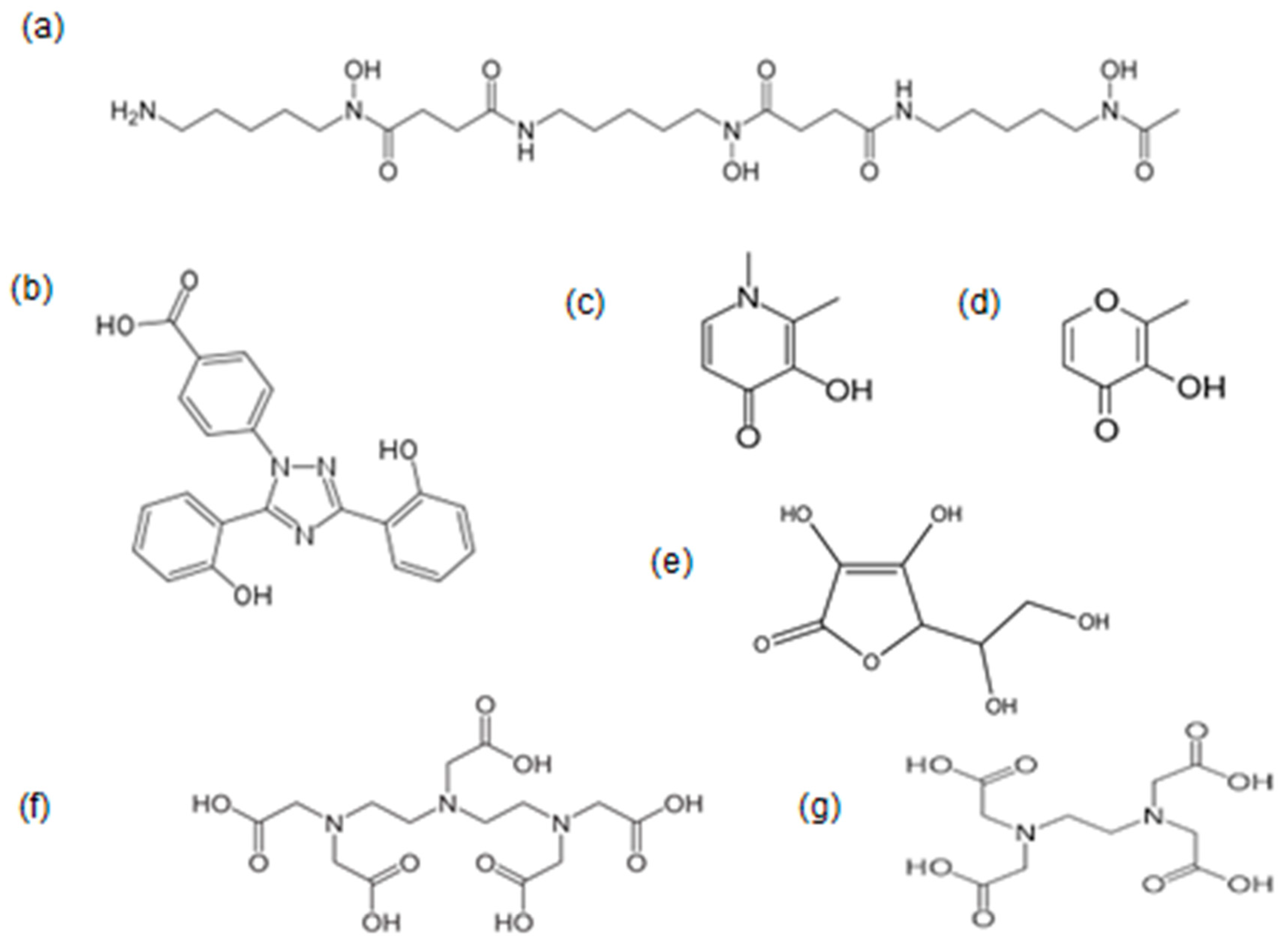
3. Iron Chelation, Natural Chelators and Chelating Drugs
3.1. Natural Biomolecules with Iron-Chelating Potential
3.2. Properties and Effects of Natural Chelator–Iron Complexes
3.3. The Properties and Effects of Iron-Chelating Drugs
4. Recent Advances in Chelator–Iron Complexes for the Treatment of Iron Deficiency Anaemia
5. Recent Advances in Iron-Chelation Protocols for the Complete Treatment of Iron Overload
6. Future Strategies in the Treatment of Iron Deficiency Anaemia and Iron Overload Diseases
7. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADMET | absorption, distribution, metabolism, excretion and toxicity |
| DF | deferoxamine |
| DFRA | deferasirox |
| DMT1 | divalent metal transported protein |
| DTPA | diethylenetriaminepentaacetic acid |
| EDTA | ethylenediaminetetraacetic acid |
| FR | free radical |
| ICOC | international committee on chelation |
| IDA | iron deficiency anaemia |
| IP6 | phytic acid |
| L1 | deferiprone |
| LMWt | low molecular weight |
| MRI | magnetic resonance imaging |
| RBC | red blood cell |
| ROS | reactive oxygen species |
| sc | subcutaneous |
| T2* | (magnetic resonance imaging) relaxation time |
| TI | thalassemia intermedia |
| TM | beta thalassemia major |
References
- Cairo, G.; Bernuzzi, F.; Recalcati, S.A. A precious metal: Iron, an essential nutrient for all cells. Genes Nutr. 2006, 1, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Katsarou, A.; Pantopoulos, K. Basics and principles of cellular and systemic iron homeostasis. Mol. Asp. Med. 2020, 75, 100866. [Google Scholar] [CrossRef] [PubMed]
- Gozzelino, R.; Arosio, P. Iron Homeostasis in Health and Disease. Int. J. Mol. Sci. 2016, 17, 130. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S. Zinc: An overview. Nutrition 1995, 11 (Suppl. 1), 93–99. [Google Scholar] [PubMed]
- Coleman, J.E. Zinc proteins: Enzymes, storage proteins, transcription factors, and replication proteins. Annu. Rev. Biochem. 1992, 61, 897–946. [Google Scholar] [CrossRef]
- Daniel, K.G.; Harbach, R.H.; Guida, W.C.; Dou, Q.P. Copper storage diseases: Menkes, Wilsons, and cancer. Front. Biosci. 2004, 9, 2652–2662. [Google Scholar] [CrossRef]
- Prasad, A.S. Zinc deficiency. BMJ 2003, 326, 409–410. [Google Scholar] [CrossRef]
- Baldari, S.; Di Rocco, G.; Toietta, G. Current Biomedical Use of Copper Chelation Therapy. Int. J. Mol. Sci. 2020, 21, 1069. [Google Scholar] [CrossRef] [PubMed]
- Balsano, C.; Porcu, C.; Sideri, S. Is copper a new target to counteract the progression of chronic diseases? Metallomics 2018, 10, 1712–1722. [Google Scholar] [CrossRef] [PubMed]
- Andrews, N.C. Disorders of iron metabolism. N. Engl. J. Med. 1999, 341, 1986–1995, Erratum in: N. Engl. J. Med. 2000, 342, 364. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Kontoghiorghe, C.N. Iron and Chelation in Biochemistry and Medicine: New Approaches to Controlling Iron Metabolism and Treating Related Diseases. Cells 2020, 9, 1456. [Google Scholar] [CrossRef] [PubMed]
- McLean, E.; Cogswell, M.; Egli, I.; Wojdyla, D.; De Benoist, B. Worldwide Prevalence of Anaemia, WHO Vitamin and Mineral Nutrition Information System, 1993–2005. Public Health Nutr. 2009, 12, 444–454. [Google Scholar] [CrossRef]
- Pasricha, S.R.; Tye-Din, J.; Muckenthaler, M.U.; Swinkels, D.W. Iron deficiency. Lancet 2021, 397, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Tardy, A.L.; Pouteau, E.; Marquez, D.; Yilmaz, C.; Scholey, A. Vitamins and Minerals for Energy, Fatigue and Cognition: A Narrative Review of the Biochemical and Clinical Evidence. Nutrients 2020, 12, 228. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Anemia of Inflammation. N. Engl. J. Med. 2019, 381, 1148–1157. [Google Scholar] [CrossRef] [PubMed]
- Pham, J.; Laven-Law, G.; Winter, J.M.; Wassie, M.M.; Cock, C.; Symonds, E.L. The Diagnostic Accuracy of a Fecal Immunochemical Test in Detecting Colorectal Cancer and Advanced Precancerous Colorectal Neoplasia in Patients with Iron Deficiency: A Protocol for Systematic Review and Meta-Analysis. Gastroenterol. Res. Pract. 2023, 2023, 5982580. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.; Wang, Y.; Xiong, H.; Xia, Y.; Cui, Z.; Liu, L. Mechanism of Iron Ion Homeostasis in Intestinal Immunity and Gut Microbiota Remodeling. Int. J. Mol. Sci. 2024, 25, 727. [Google Scholar] [CrossRef] [PubMed]
- Rohr, M.; Brandenburg, V.; Brunner-La Rocca, H.P. How to diagnose iron deficiency in chronic disease: A review of current methods and potential marker for the outcome. Eur. J. Med. Res. 2023, 28, 15. [Google Scholar] [CrossRef] [PubMed]
- Macdougall, I.C. Anaemia in CKD—Treatment standard. Nephrol. Dial. Transplant. 2023, gfad250. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Kolnagou, A. Molecular factors and mechanisms affecting iron and other metal excretion or absorption in health and disease. The role of natural and synthetic chelators. Curr. Med. Chem. 2005, 12, 2695–2709. [Google Scholar] [CrossRef]
- Teixeira Farinha, H.; Bouriez, D.; Grimaud, T.; Rotariu, A.M.; Collet, D.; Mantziari, S.; Gronnier, C. Gastro-Intestinal Disorders and Micronutrient Deficiencies following Oncologic Esophagectomy and Gastrectomy. Cancers 2023, 15, 3554. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghe, C.N.; Kolnagou, A.; Kontoghiorghes, G.J. Dietary and Pharmacological Factors Affecting Iron Absorption in Mice and Man (Comment for a Letter to the Editor). Haematologica 2016, 101, 120–121. [Google Scholar] [CrossRef] [PubMed]
- Barton, J.C.; Edwards, C.Q. (Eds.) Hemochromatosis: Genetics, Pathophysiology, Diagnosis and Treatment; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Adams, P.C. Epidemiology and diagnostic testing for hemochromatosis and iron overload. Int. J. Lab. Hematol. 2015, 37, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Feder, J.N.; Gnirke, A.; Thomas, W.; Tsuchihashi, Z.; Ruddy, D.A.; Basava, A.; Dormishian, F.; Domingo, R., Jr.; Ellis, M.C.; Fullan, A.; et al. A novel MHC class I-like gene is mutated in patients with hereditary haemochromatosis. Nat. Genet. 1996, 13, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Pietrangelo, A. Hereditary hemochromatosis—A new look at an old disease. N. Engl. J. Med. 2004, 350, 2383–2397. [Google Scholar] [CrossRef] [PubMed]
- Dubois, S.; Kowdley, K.V. Targeted screening for hereditary haemochromatosis in high-risk groups. Aliment. Pharmacol. Ther. 2004, 20, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Community control of hereditary anaemias: Memorandum from a WHO meeting. Bull. World Health Org. 1983, 61, 63–80. [Google Scholar]
- Modell, B.; Berdoukas, V. The Clinical Approach to Thalassaemia; Grune and Stratton: New York, NY, USA, 1984; pp. 165–169. [Google Scholar]
- Weatherall, D.; Clegg, J.B. Inherited haemoglobin disorders: An increasing global health problem. Bull. World Health Organ. 2001, 79, 704–712. [Google Scholar] [PubMed]
- Hokland, P.; Daar, S.; Khair, W.; Sheth, S.; Taher, A.T.; Torti, L.; Hantaweepant, C.; Rund, D. Thalassaemia—A global view. Br. J. Haematol. 2023, 201, 199–214. [Google Scholar] [CrossRef]
- Cattoni, A.; Capitoli, G.; Casagranda, S.; Corti, P.; Adavastro, M.; Molinaro, A.; Di Gennaro, F.; Bonanomi, S.; Biondi, A.; Galimberti, S.; et al. Iron Overload Following Hematopoietic Stem Cell Transplantation: Prevalence, Severity, and Management in Children and Adolescents with Malignant and Nonmalignant Diseases. Transplant. Cell. Ther. 2023, 29, 271.e1–271.e12. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. How to manage iron toxicity in post-allogeneic hematopoietic stem cell transplantation? Expert Rev. Hematol. 2020, 13, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, S.; Angelucci, E.; Arnold, S.D.; Baker, K.S.; Bhatia, M.; Bresters, D.; Dietz, A.C.; De La Fuente, J.; Duncan, C.; Gaziev, J.; et al. Current Results and Future Research Priorities in Late Effects after Hematopoietic Stem Cell Transplantation for Children with Sickle Cell Disease and Thalassemia: A Consensus Statement from the Second Pediatric Blood and Marrow Transplant Consortium International Conference on Late Effects after Pediatric Hematopoietic Stem Cell Transplantation. Biol. Blood Marrow Transplant. 2017, 23, 552–561. [Google Scholar] [PubMed]
- Gu, Y.; Estcourt, L.J.; Doree, C.; Hopewell, S.; Vyas, P. Comparison of a restrictive versus liberal red cell transfusion policy for patients with myelodysplasia, aplastic anaemia, and other congenital bone marrow failure disorders. Cochrane Database Syst. Rev. 2015, 2015, CD011577. [Google Scholar]
- Tang, A.Y.; Zhou, M.; Maillis, A.N.; Lai, K.W.; Lane, P.A.; Snyder, A.B. Trends in blood transfusion, hydroxyurea use, and iron overload among children with sickle cell disease enrolled in Medicaid, 2004–2019. Pediatr. Blood Cancer 2023, 70, e30152. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C.; Cross, C.E. Free radicals, antioxidants and human disease: Where are we now? J. Lab. Clin. Med. 1992, 119, 598–620. [Google Scholar] [PubMed]
- Galaris, D.; Barbouti, A.; Pantopoulos, K. Iron homeostasis and oxidative stress: An intimate relationship. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 118535. [Google Scholar] [CrossRef]
- Ďuračková, Z. Some current insights into oxidative stress. Physiol. Res. 2010, 59, 459–469. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. Prospects for introducing deferiprone as potent pharmaceutical antioxidant. Front. Biosci. 2009, 1, 161–178. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Naguro, I.; Ichijo, H. Iron homeostasis and iron-regulated ROS in cell death, senescence and human diseases. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 1398–1409. [Google Scholar] [CrossRef] [PubMed]
- Fibach, E. The Redox Balance and Membrane Shedding in RBC Production, Maturation, and Senescence. Front. Physiol. 2021, 12, 604738. [Google Scholar] [CrossRef]
- Denisov, E.T.; Afanas’ev, I.B. Oxidation and Antioxidants in Organic Chemistry and Biology; CRC Press: Boca Raton, FL, USA; Taylor and Francis Group: Abingdon, UK, 2005. [Google Scholar]
- Shah, S.V.; Rajapurkar, M.M. The role of labile iron in kidney disease and treatment with chelation. Hemoglobin 2009, 33, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Reeder, B.; Wilson, M.T. Hemoglobin and myoglobin associated oxidative stress: From molecular mechanisms to disease States. Curr. Med. Chem. 2005, 12, 2741–2751. [Google Scholar] [CrossRef] [PubMed]
- Reeder, B.J. Redox and Peroxidase Activities of the Hemoglobin Superfamily: Relevance to Health and Disease. Antioxid. Redox Signal. 2017, 26, 763–776. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.Y.; Dixon, S.J. Mechanisms of ferroptosis. Cell Mol. Life Sci. 2016, 73, 2195–2209. [Google Scholar] [CrossRef]
- Li, G.; Liang, Y.; Yang, H.; Zhang, W.; Xie, T. The Research Landscape of Ferroptosis in Cancer: A Bibliometric Analysis. Front. Cell Dev. Biol. 2022, 10, 841724. [Google Scholar] [CrossRef]
- Park, E.; Chung, S.W. ROS-mediated autophagy increases intracellular iron levels and ferroptosis by ferritin and transferrin receptor regulation. Cell Death Dis. 2019, 10, 822. [Google Scholar] [CrossRef]
- Gao, W.; Wang, X.; Zhou, Y.; Wang, X.; Yu, Y. Autophagy, ferroptosis, pyroptosis, and necroptosis in tumor immunotherapy. Signal Transduct. Target. Ther. 2022, 7, 196. [Google Scholar] [CrossRef]
- Xie, Y.; Hou, W.; Song, X.; Yu, Y.; Huang, J.; Sun, X.; Kang, R.; Tang, D. Ferroptosis: Process and function. Cell Death Differ. 2016, 23, 369–379. [Google Scholar] [CrossRef]
- Wang, P.; Lu, Y.-Q. Ferroptosis: A Critical Moderator in the Life Cycle of Immune Cells. Front. Immunol. 2022, 13, 877634. [Google Scholar] [CrossRef]
- Gao, M.; Monian, P.; Pan, Q.; Zhang, W.; Xiang, J.; Jiang, X. Ferroptosis is an autophagic cell death process. Cell Res. 2016, 26, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, Q.; Song, H.; Zhang, Y.; Chen, H.; Liu, P.; Sun, T.; Jiang, C. A Triple Therapeutic Strategy with Antiexosomal Iron Efflux for Enhanced Ferroptosis Therapy and Immunotherapy. Small 2022, 18, e2201704. [Google Scholar] [CrossRef]
- Hou, W.; Xie, Y.; Song, X.; Sun, X.; Zeh, H.J., 3rd; Kang, R.; Tang, D. Autophagy promotes ferroptosis by degradation of ferritin. Autophagy 2016, 12, 1425–1428. [Google Scholar] [CrossRef]
- Liu, L.; Li, L.; Li, M.; Luo, Z. Autophagy-Dependent Ferroptosis as a Therapeutic Target in Cancer. ChemMedChem 2021, 16, 2942–2950. [Google Scholar] [CrossRef] [PubMed]
- de Souza, I.; Ramalho, M.C.C.; Guedes, C.B.; Osawa, I.Y.A.; Monteiro, L.K.S.; Gomes, L.R.; Rocha, C.R.R. Ferroptosis Modulation: Potential Therapeutic Target for Glioblastoma Treatment. Int. J. Mol. Sci. 2022, 23, 6879. [Google Scholar] [CrossRef] [PubMed]
- González-Montero, J.; Chichiarelli, S.; Eufemi, M.; Altieri, F.; Saso, L.; Rodrigo, R. Ascorbate as a Bioactive Compound in Cancer Therapy: The Old Classic Strikes Back. Molecules 2022, 27, 3818. [Google Scholar] [CrossRef]
- Yin, L.; Tang, Y. Predicting prognosis and clinical features of the tumor microenvironment based on ferroptosis score in patients with breast cancer. Sci. Rep. 2022, 12, 10611. [Google Scholar] [CrossRef]
- Wang, S.; Wei, W.; Ma, N.; Qu, Y.; Liu, Q. Molecular mechanisms of ferroptosis and its role in prostate cancer therapy. Crit. Rev. Oncol. 2022, 176, 103732. [Google Scholar] [CrossRef]
- Balachander, K.; Paramasivam, A. Ferroptosis: An emerging therapeutic target for oral cancer. Oral Oncol. 2022, 131, 105970. [Google Scholar] [CrossRef]
- Si, C.; Zhou, X.; Deng, J.; Ye, S.; Kong, L.; Zhang, B.; Wang, W. Role of ferroptosis in gastrointestinal tumors: From mechanisms to therapies. Cell Biol. Int. 2022, 46, 997–1008. [Google Scholar] [CrossRef]
- Chen, J.; Wei, Z.; Fu, K.; Duan, Y.; Zhang, M.; Li, K.; Guo, T.; Yin, R. Non-apoptotic cell death in ovarian cancer: Treatment, resistance and prognosis. Biomed. Pharmacother. 2022, 150, 112929. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.K.; Chu, Y.H.; Syue, W.J.; Lin, H.Y.; Chang, W.T.; Chen, J.Y.; Wu, C.Y.; Yen, C.H.; Cheng, K.C.; Chiu, C.C. The Role of Nonapoptotic Programmed Cell Death—Ferroptosis, Necroptosis, and Pyroptosis—In Pancreatic Ductal Adenocarcinoma Treatment. Front. Oncol. 2022, 12, 872883. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Adzavon, Y.M.; Wen, X.; Zhao, P.; Xie, F.; Liu, M.; Ma, X. Novel Insights in the Regulatory Mechanisms of Ferroptosis in Hepatocellular Carcinoma. Front. Cell Dev. Biol. 2022, 10, 873029. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Shi, Y.; Gao, Z.; Sun, Y.; Yao, F.; Ma, L. Ferroptosis at the crossroads of tumor-host interactions, metastasis, and therapy response. Am. J. Physiol. Physiol. 2022, 323, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Nie, Z.; Chen, M.; Gao, Y.; Huang, D.; Cao, H.; Peng, Y.; Guo, N.; Wang, F.; Zhang, S. Ferroptosis and Tumor Drug Resistance: Current Status and Major Challenges. Front. Pharmacol. 2022, 13, 879317. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Meng, X.; Peng, L.; Xie, W.; Liu, X.; He, W.; Li, S. Ferroptosis and cancer immunotherapy. Curr. Mol. Med. 2022, 23, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Xu, B.; Hu, N.; Guo, Z.; Bao, W.; Shao, B.; Yang, W. Targeting the Macrophage-Ferroptosis Crosstalk: A Novel Insight into Tumor Immunotherapy. Front. Biosci. 2022, 27, 203. [Google Scholar] [CrossRef]
- Guan, X.Y.; Guan, X.L.; Zhu, J.R. Mechanisms and applications of ferroptosis-associated regulators in cancer therapy and drug resistance. J. Chemother. 2023, 35, 671–688. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Miao, J. Ferroptosis: Mechanism and potential applications in cervical cancer. Front. Mol. Biosci. 2023, 10, 1164398. [Google Scholar] [CrossRef] [PubMed]
- Lei, G.; Zhuang, L.; Gan, B. Targeting ferroptosis as a vulnerability in cancer. Nat. Rev. Cancer 2022, 22, 381–396. [Google Scholar] [CrossRef]
- Wang, D.; Le Tang, L.; Zhang, Y.; Ge, G.; Jiang, X.; Mo, Y.; Wu, P.; Deng, X.; Li, L.; Zuo, S.; et al. Regulatory pathways and drugs associated with ferroptosis in tumors. Cell Death Dis. 2022, 13, 544. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Huang, H.; Fan, S.; Zheng, B.; Wu, J.; Zhang, J.; Pi, J.; Xu, J.-F. Ferroptosis: A mixed blessing for infectious diseases. Front. Pharmacol. 2022, 13, 992734. [Google Scholar] [CrossRef]
- Lin, Y.; Xu, W.; Hou, Y.; Wang, S.; Zhang, H.; Ran, M.; Huang, Y.; Wang, Y.; Yang, G. The multifaceted role of ferroptosis in kidney diseases. Chem. Interact. 2022, 365, 110107. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Chen, X.-Z.; Wang, Y.-H.; Cheng, X.-L.; Zhao, Y.; Zhou, L.-Y.; Wang, K. Emerging roles of ferroptosis in cardiovascular diseases. Cell Death Discov. 2022, 8, 394. [Google Scholar] [CrossRef] [PubMed]
- Thapa, K.; Khan, H.; Kanojia, N.; Singh, T.G.; Kaur, A.; Kaur, G. Therapeutic Insights on Ferroptosis in Parkinson’s disease. Eur. J. Pharmacol. 2022, 930, 175133. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, W.; Lammens, M.; Kerckhofs, A.; Voets, E.; Van San, E.; Van Coillie, S.; Peleman, C.; Mergeay, M.; Sirimsi, S.; Matheeussen, V.; et al. Fatal lymphocytic cardiac damage in coronavirus disease 2019 (COVID-19): Autopsy reveals a ferroptosis signature. ESC Heart Fail. 2020, 7, 3772–3781. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J. Deferiprone: A Forty-Year-Old Multi-Targeting Drug with Possible Activity against COVID-19 and Diseases of Similar Symptomatology. Int. J. Mol. Sci. 2022, 23, 6735. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Lv, Z.; Li, S.; Jiang, H.; Han, B.; Zheng, X.; Liu, Y.; Zhang, Z. Chronic arsenic exposure induces ferroptosis via enhancing ferritinophagy in chicken livers. Sci. Total Environ. 2023, 890, 164172. [Google Scholar] [CrossRef]
- Costa, I.; Barbosa, D.J.; Benfeito, S.; Silva, V.; Chavarria, D.; Borges, F.; Remião, F.; Silva, R. Molecular mechanisms of ferroptosis and their involvement in brain diseases. Pharmacol. Ther. 2023, 244, 108373. [Google Scholar] [CrossRef]
- Rayatpour, A.; Foolad, F.; Heibatollahi, M.; Khajeh, K.; Javan, M. Ferroptosis inhibition by deferiprone, attenuates myelin damage and promotes neuroprotection in demyelinated optic nerve. Sci. Rep. 2022, 12, 19630. [Google Scholar] [CrossRef]
- Wang, C.; Xie, L.; Xing, Y.; Liu, M.; Yang, J.; Gao, N.; Cai, Y. Iron-overload-induced ferroptosis in mouse cerebral toxoplasmosis promotes brain injury and could be inhibited by Deferiprone. PLoS Neglected Trop. Dis. 2023, 17, e0011607. [Google Scholar] [CrossRef]
- Asperti, M.; Cantamessa, L.; Gryzik, M.; Bugatti, M.; Codenotti, S.; Denardo, A.; Vermi, W.; Fanzani, A.; Poli, M. The modulation of iron metabolism affects the Rhabdomyosarcoma tumor growth in vitro and in vivo. Clin. Exp. Med. 2023, 23, 2487–2502. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J. New Iron Metabolic Pathways and Chelation Targeting Strategies Affecting the Treatment of All Types and Stages of Cancer. Int. J. Mol. Sci. 2022, 23, 13990. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, L.; Mo, Y.; Li, J.; Yang, J.; Wang, J.; Karrow, N.A.; Wu, H.; Sun, L. Ferroptosis is involved in deoxynivalenol-induced intestinal damage in pigs. J. Anim. Sci. Biotechnol. 2023, 14, 29. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J. Advances on Chelation and Chelator Metal Complexes in Medicine. Int. J. Mol. Sci. 2020, 21, 2499. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, L.N.; Kontoghiorghes, G.J. Competition between deferiprone, desferrioxamine and other chelators for iron and the effect of other metals. Arzneimittelforschung 1993, 43, 659–663. [Google Scholar] [PubMed]
- Kontoghiorghe, C.N.; Kolnagou, A.; Kontoghiorghes, G.J. Phytochelators Intended for Clinical Use in Iron Overload, Other Diseases of Iron Imbalance and Free Radical Pathology. Molecules 2015, 20, 20841–20872. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, H. Transferrin and transferrin receptors update. Free. Radic. Biol. Med. 2019, 133, 46–54. [Google Scholar] [CrossRef]
- Sargent, P.J.; Farnaud, S.; Evans, R.W. Structure/function overview of proteins involved in iron storage and transport. Curr. Med. Chem. 2005, 12, 2683–2693. [Google Scholar] [CrossRef] [PubMed]
- Gomme, P.T.; McCann, K.B.; Bertolini, J. Transferrin: Structure, function and potential therapeutic actions. Drug Discov. Today 2005, 10, 267–273. [Google Scholar] [CrossRef]
- Pantopoulos, K. TfR2 links iron metabolism and erythropoiesis. Blood 2015, 125, 1055–1056. [Google Scholar] [CrossRef] [PubMed]
- Makey, D.G.; Seal, U.S. The detection of four molecular forms of human transferrin during the iron binding process. Biochim. Biophys. Acta 1976, 453, 250–256. [Google Scholar] [CrossRef]
- Jacobs, A. An intracellular transit iron pool. In Ciba Foundation Symposium 51-Iron Metabolism; John Wiley & Sons, Ltd.: Chichester, UK, 1976; Volume 51, pp. 91–106. [Google Scholar]
- Arosio, P.; Elia, L.; Poli, M. Ferritin, cellular iron storage and regulation. IUBMB Life 2017, 69, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Theil, E.C. Ferritin: The protein nanocage and iron biomineral in health and in disease. Inorg. Chem. 2013, 52, 12223–12233. [Google Scholar] [CrossRef] [PubMed]
- La, A.; Nguyen, T.; Tran, K.; Sauble, E.; Tu, D.; Gonzalez, A.; Kidane, T.Z.; Soriano, C.; Morgan, J.; Doan, M.; et al. Mobilization of iron from ferritin: New steps and details. Metallomics 2018, 10, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Mehlenbacher, M.; Poli, M.; Arosio, P.; Santambrogio, P.; Levi, S.; Chasteen, N.D.; Bou-Abdallah, F. Iron Oxidation and Core Formation in Recombinant Heteropolymeric Human Ferritins. Biochemistry 2017, 56, 3900–3912. [Google Scholar] [CrossRef] [PubMed]
- Saito, H. Storage Iron Turnover from a New Perspective. Acta Haematol. 2019, 141, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Iancu, T.C. Ferritin and hemosiderin in pathological tissues. Electron Microsc. Rev. 1992, 5, 209–229. [Google Scholar] [CrossRef] [PubMed]
- Brissot, P.; Troadec, M.B.; Loréal, O.; Brissot, E. Pathophysiology and classification of iron overload diseases; update 2018. Transfus. Clin. Biol. 2019, 26, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Camaschella, C.; Nai, A. Ineffective erythropoiesis and regulation of iron status in iron loading anaemias. Br. J. Haematol. 2016, 172, 512–523. [Google Scholar] [CrossRef]
- Gozzelino, R.; Poli, M.; Arosio, P. Iron as Therapeutic Target in Human Diseases. Pharmaceuticals 2019, 12, 178. [Google Scholar] [CrossRef]
- Iancu, T.C. Ultrastructural aspects of iron storage, transport and metabolism. J. Neural Transm. 2011, 118, 329–335. [Google Scholar] [CrossRef]
- Kolnagou, A.; Natsiopoulos, K.; Kleanthous, M.; Ioannou, A.; Kontoghiorghes, G.J. Liver iron and serum ferritin levels are misleading for estimating cardiac, pancreatic, splenic and total body iron load in thalassemia patients: Factors influencing the heterogenic distribution of excess storage iron in organs as identified by MRI T2*. Toxicol. Mech. Methods 2013, 23, 48–56. [Google Scholar] [CrossRef]
- Brewer, C.J.; Coates, T.D.; Wood, J.C. Spleen R2 and R2* in iron-overloaded patients with sickle cell disease and thalassemia major. J. Magn. Reson. Imaging 2009, 29, 357–364. [Google Scholar] [CrossRef]
- Byrnes, V.; Barrett, S.; Ryan, E.; Kelleher, T.; O’Keane, C.; Coughlan, B.; Crowe, J. Increased duodenal DMT-1 expression and unchanged HFE mRNA levels in HFE-associated hereditary hemochromatosis and iron deficiency. Blood Cells Mol. Dis. 2002, 29, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Aschemeyer, S.; Qiao, B.; Stefanova, D.; Valore, E.V.; Sek, A.C.; Ruwe, T.A.; Vieth, K.R.; Jung, G.; Casu, C.; Rivella, S.; et al. Structure-function analysis of ferroportin defines the binding site and an alternative mechanism of action of hepcidin. Blood 2018, 131, 899–910. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Drakesmith, H.; Nemeth, E.; Ganz, T. Ironing out Ferroportin. Cell Metab. 2015, 22, 777–787. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Billesbølle, C.B.; Azumaya, C.M.; Kretsch, R.C.; Powers, A.S.; Gonen, S.; Schneider, S.; Arvedson, T.; Dror, R.O.; Cheng, Y.; Manglik, A. Structure of hepcidin-bound ferroportin reveals iron homeostatic mechanisms. Nature 2020, 586, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, E.; Ganz, T. Hepcidin and Iron in Health and Disease. Annu. Rev. Med. 2023, 74, 261–277. [Google Scholar] [CrossRef]
- Girelli, D.; Nemeth, E.; Swinkels, D.W. Hepcidin in the diagnosis of iron disorders. Blood 2016, 127, 2809–2813. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Alvarenga, A.M.; Brissot, P.; Santos, P.C.J.L. Haemochromatosis revisited. World J. Hepatol. 2022, 14, 1931–1939. [Google Scholar] [CrossRef]
- Pantopoulos, K. Function of the hemochromatosis protein HFE: Lessons from animal models. World J. Gastroenterol. 2008, 14, 6893–6901. [Google Scholar] [CrossRef]
- Pietrangelo, A. Ferroportin disease: Pathogenesis, diagnosis and treatment. Haematologica 2017, 102, 1972–1984. [Google Scholar] [CrossRef]
- Zhang, D.L.; Ghosh, M.C.; Ollivierre, H.; Li, Y.; Rouault, T.A. Ferroportin deficiency in erythroid cells causes serum iron deficiency and promotes hemolysis due to oxidative stress. Blood 2018, 132, 2078–2087. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Worwood, M. Inborn errors of metabolism: Iron. Br. Med. Bull. 1999, 55, 556–567. [Google Scholar] [CrossRef]
- Moustarah, F.; Daley, S.F. Dietary Iron. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar] [PubMed]
- Sim, M.; Garvican-Lewis, L.A.; Cox, G.R.; Govus, A.; McKay, A.K.A.; Stellingwerff, T.; Peeling, P. Iron considerations for the athlete: A narrative review. Eur. J. Appl. Physiol. 2019, 119, 1463–1478. [Google Scholar] [CrossRef]
- Salvin, H.E.; Pasricha, S.R.; Marks, D.C.; Speedy, J. Iron deficiency in blood donors: A national cross-sectional study. Transfusion 2014, 54, 2434–2444. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Kolnagou, A.; Demetriou, T.; Neocleous, M.; Kontoghiorghe, C.N. New Era in the Treatment of Iron Deficiency Anaemia Using Trimaltol Iron and Other Lipophilic Iron Chelator Complexes: Historical Perspectives of Discovery and Future Applications. Int. J. Mol. Sci. 2021, 22, 5546. [Google Scholar] [CrossRef]
- Kobayashi, M.; Suhara, T.; Baba, Y.; Kawasaki, N.K.; Higa, J.K.; Matsui, T. Pathological Roles of Iron in Cardiovascular Disease. Curr. Drug Targets 2018, 19, 1068–1076. [Google Scholar] [CrossRef]
- Man, C.D.; Maideen, S.F.K.; Rashid, A. Knowledge, Attitude and Practice towards Dietary Iron among Patients with Thalassemia and Their Caregivers in Peninsular Malaysia. Med. J. Malays 2019, 74, 365–371. [Google Scholar]
- Mantadakis, E.; Chatzimichael, E.; Zikidou, P. Iron Deficiency Anemia in Children Residing in High and Low-Income Countries: Risk Factors, Prevention, Diagnosis and Therapy. Mediterr. J. Hematol. Infect. Dis. 2020, 12, e2020041. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Grover, K.; Kumar, T.; Doda, A.; Bhutani, R.; Yadav, S.; Kaushal, P.; Kapoor, R.; Sharma, S. Prevalence of anaemia and its association with dietary habits among pregnant women in the urban area of Haryana. J. Fam. Med. Prim. Care 2020, 9, 783–787. [Google Scholar] [PubMed Central]
- Savva, S.C.; Kafatos, A. Is red meat required for the prevention of iron deficiency among children and adolescents? Curr. Pediatr. Rev. 2014, 10, 177–183. [Google Scholar] [CrossRef]
- Wessells, K.R.; Young, R.R.; Ferguson, E.L.; Ouédraogo, C.T.; Faye, M.T.; Hess, S.Y. Assessment of Dietary Intake and Nutrient Gaps, and Development of Food-Based Recommendations, Among Pregnant and Lactating Women in Zinder, Niger: An Optifood Linear Programming Analysis. Nutrients 2019, 11, 72. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Rawal, S. Dietary Iron Intake, Iron Status, and Gestational Diabetes. Am. J. Clin. Nutr. 2017, 106, 1672–1680. [Google Scholar] [CrossRef] [PubMed]
- Senba, M.; Nakamura, T.; Itakura, H. Relationships among iron accumulation, cirrhosis, and hepatitis B virus infection in Bantu siderosis. Ann. Soc. Belg. Med. Trop. 1989, 69, 77–78. [Google Scholar] [PubMed]
- Robertson, A.; Tenenbein, M. Hepatotoxicity in acute iron poisoning. Hum. Exp. Toxicol. 2005, 24, 559–562. [Google Scholar] [CrossRef]
- Kapoor, R.; Sabharwal, M.; Ghosh-Jerath, S. Diet Quality, Nutritional Adequacy and Anthropometric Status among Indigenous Women of Reproductive Age Group (15–49 Years) in India: A Narrative Review. Dietetics 2022, 2, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Assaf, E.A.; Al Sabbah, H.; Al-Jawadleh, A. Analysis of the nutritional status in the Palestinian territory: A review study. Front. Nutr. 2023, 10, 1206090. [Google Scholar] [CrossRef]
- Harrison, A.V.; Lorenzo, F.R.; McClain, D.A. Iron and the Pathophysiology of Diabetes. Annu. Rev. Physiol. 2023, 85, 339–362. [Google Scholar] [CrossRef]
- Baschant, U.; Altamura, S.; Steele-Perkins, P.; Muckenthaler, M.U.; Spasić, M.V.; Hofbauer, L.C.; Steinbicker, A.U.; Rauner, M. Iron effects versus metabolic alterations in hereditary hemochromatosis driven bone loss. Trends Endocrinol. Metab. 2022, 33, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Calori, S.; Comisi, C.; Mascio, A.; Fulchignoni, C.; Pataia, E.; Maccauro, G.; Greco, T.; Perisano, C. Overview of Ankle Arthropathy in Hereditary Hemochromatosis. Med. Sci. 2023, 11, 51. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Peterson, S.J.; Parikh, M.A.; Frishman, W.H. Cardiovascular Manifestations of Hemochromatosis: A Review of Pathophysiology, Mechanisms, and Treatment Options. Cardiol. Rev. 2023. [Google Scholar] [CrossRef] [PubMed]
- Haider, M.B.; Al Sbihi, A.; Chaudhary, A.J.; Haider, S.M.; Edhi, A.I. Hereditary hemochromatosis: Temporal trends, sociodemographic characteristics, and independent risk factor of hepatocellular cancer—Nationwide population-based study. World J. Hepatol. 2022, 14, 1804–1816. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Spyrou, A.; Kolnagou, A. Iron chelation therapy in hereditary hemochromatosis and thalassemia intermedia: Regulatory and non regulatory mechanisms of increased iron absorption. Hemoglobin 2010, 34, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Pippard, M.J.; Callender, S.T.; Warner, G.T.; Weatherall, D.J. Iron absorption and loading in beta-thalassaemia intermedia. Lancet 1979, 2, 819–821. [Google Scholar] [CrossRef] [PubMed]
- Pootrakul, P.; Kitcharoen, K.; Yansukon, P.; Wasi, P.; Fucharoen, S.; Charoenlarp, P.; Brittenham, G.; Pippard, M.J.; Finch, C.A. The effect of erythroid hyperplasia on iron balance. Blood 1988, 71, 1124–1129. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghe, C.N.; Kontoghiorghes, G.J. Efficacy and safety of iron-chelation therapy with deferoxamine, deferiprone, and deferasirox for the treatment of iron-loaded patients with non-transfusion-dependent thalassemia syndromes. Drug Des. Devel Ther. 2016, 10, 465–481. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J. Iron Load Toxicity in Medicine: From Molecular and Cellular Aspects to Clinical Implications. Int. J. Mol. Sci. 2023, 24, 12928. [Google Scholar] [CrossRef]
- Kolnagou, A.; Kleanthous, M.; Kontoghiorghes, G.J. Benefits and Risks in Polypathology and Polypharmacotherapy Challenges in the Era of the Transition of Thalassaemia from a Fatal to a Chronic or Curable Disease. Front. Biosci. 2022, 14, 18. [Google Scholar] [CrossRef]
- Guarente, J.; Tormey, C. Transfusion Support of Patients with Myelodysplastic Syndromes. Clin. Lab. Med. 2023, 43, 669–683. [Google Scholar] [CrossRef] [PubMed]
- Lok, C.T.H.; Ser, A.J.T.; Oligbu, P.; Pervaiz, M.; Oligbu, G. Risk of Streptococcus pneumoniae-associated haemolytic uraemic syndrome in industrialised nations: A systematic review of the literature. Epidemiol. Mikrobiol. Imunol. 2023, 72, 213–220. [Google Scholar]
- Friberg, N.; Arvidsson, I.; Tontanahal, A.; Kristoffersson, A.C.; Gram, M.; Kaplan, B.S.; Karpman, D. Red blood cell-derived arginase release in hemolytic uremic syndrome. J. Transl. Med. 2024, 22, 17. [Google Scholar] [CrossRef] [PubMed]
- Anderson, F.W.; Hiller, M.C. (Eds.) Development of Iron Chelators for Clinical Use; DHEW Publication No. (NIH) 77-994; DHEW: Bethesda, MA, USA, 1975; pp. 1–275. [Google Scholar]
- Kontoghiorghes, G.J. The Design of Orally Active Iron Chelators for the Treatment of Thalassaemia. Ph.D. Thesis, University of Essex, Colchester, UK, 1982; pp. 1–243, British Library Microfilm No D66194/86. Available online: https://www.pri.ac.cy/files/KGJ_thesis_1982.pdf (accessed on 1 March 2023).
- Kontoghiorghes, G.J. Design, properties, and effective use of the oral chelator L1 and other alpha-ketohydroxypyridines in the treatment of transfusional iron overload in thalassemia. Ann. N. Y. Acad. Sci. 1990, 612, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Pattichis, K.; Neocleous, K.; Kolnagou, A. The design and development of deferiprone (L1) and other iron chelators for clinical use: Targeting methods and application prospects. Curr. Med. Chem. 2004, 11, 2161–2183. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Eracleous, E.; Economides, C.; Kolnagou, A. Advances in iron overload therapies. Prospects for effective use of deferiprone (L1), deferoxamine, the new experimental chelators ICL670, GT56-252, L1NAll and their combination. Curr. Med. Chem. 2005, 12, 2663–2681. [Google Scholar] [CrossRef] [PubMed]
- Levina, A.; Lay, P.A. Transferrin Cycle and Clinical Roles of Citrate and Ascorbate in Improved Iron Metabolism. ACS Chem. Biol. 2019, 14, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Pratt, R.; Handelman, G.J.; Edwards, T.E.; Gupta, A. Ferric pyrophosphate citrate: Interactions with transferrin. Biometals 2018, 31, 1081–1089. [Google Scholar] [CrossRef]
- Luna, B.M.; Ershova, K.; Yan, J.; Ulhaq, A.; Nielsen, T.B.; Hsieh, S.; Pantapalangkoor, P.; Vanscoy, B.; Ambrose, P.; Rudin, S.; et al. Adjunctive transferrin to reduce the emergence of antibiotic resistance in Gram-negative bacteria. J. Antimicrob. Chemother. 2019, 74, 2631–2639. [Google Scholar] [CrossRef]
- Kontoghiorghe, C.N.; Kolnagou, A.; Kontoghiorghes, G.J. Potential clinical applications of chelating drugs in diseases targeting transferrin-bound iron and other metals. Expert Opin. Investig. Drugs 2013, 22, 591–618. [Google Scholar] [CrossRef]
- Ott, D.B.; Hartwig, A.; Stillman, M.J. Competition between Al3+ and Fe3+ binding to human transferrin and toxicological implications: Structural investigations using ultra-high resolution ESI MS and CD spectroscopy. Metallomics 2019, 11, 968–981. [Google Scholar] [CrossRef]
- González-Chávez, S.A.; Arévalo-Gallegos, S.; Rascón-Cruz, Q. Lactoferrin: Structure, function and applications. Int. J. Antimicrob. Agents 2009, 33, 301.e1–301.e8. [Google Scholar] [CrossRef]
- Wang, B.; Timilsena, Y.P.; Blanch, E.; Adhikari, B. Lactoferrin: Structure, function, denaturation and digestion. Crit. Rev. Food Sci. Nutr. 2019, 59, 580–596. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, K.E.; Weeks, K.; Carter, D.A. Lactoferrin Is Broadly Active against Yeasts and Highly Synergistic with Amphotericin B. Antimicrob. Agents Chemother. 2020, 64, e02284-19. [Google Scholar] [CrossRef] [PubMed]
- Rosa, L.; Cutone, A.; Lepanto, M.S.; Paesano, R.; Valenti, P. Lactoferrin: A Natural Glycoprotein Involved in Iron and Inflammatory Homeostasis. Int. J. Mol. Sci. 2017, 18, 1985. [Google Scholar] [CrossRef]
- Van Gorkom, G.N.Y.; Lookermans, E.L.; Van Elssen, C.H.M.J.; Bos, G.M.J. The effect of vitamin C (Ascorbic acid) in the treatment of patients with cancer: A systematic review. Nutrients 2019, 11, 977. [Google Scholar] [CrossRef] [PubMed]
- Hager, D.N.; Hinson, J.S.; Rothman, R.E. Vitamin C for Sepsis and Acute Respiratory Failure. JAMA-J. Am. Med. Assoc. 2020, 323, 791–792. [Google Scholar] [CrossRef]
- Nielsen, T.K.; Højgaard, M.; Andersen, J.T.; Poulsen, H.E.; Lykkesfeldt, J.; Mikines, K.J. Elimination of ascorbic acid after high-dose infusion in prostate cancer patients: A pharmacokinetic evaluation. Basic Clin. Pharmacol. Toxicol. 2015, 116, 343–348. [Google Scholar] [CrossRef]
- Hemilä, H.; Chalker, E. Vitamin C reduces the severity of common colds: A meta-analysis. BMC Public Health 2023, 23, 2468. [Google Scholar] [CrossRef]
- Bhoot, H.R.; Zamwar, U.M.; Chakole, S.; Anjankar, A. Dietary Sources, Bioavailability, and Functions of Ascorbic Acid (Vitamin C) and Its Role in the Common Cold, Tissue Healing, and Iron Metabolism. Cureus 2023, 15, e49308. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Kolnagou, A.; Kontoghiorghe, C.N.; Mourouzidis, L.; Timoshnikov, V.A.; Polyakov, N.E. Trying to Solve the Puzzle of the Interaction of Ascorbic Acid and Iron: Redox, Chelation and Therapeutic Implications. Medicines 2020, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Manolis, A.A.; Manolis, T.A.; Melita, H.; Manolis, A.S. Role of Vitamins in Cardiovascular Health: Know Your Facts-Part 2. Curr. Vasc. Pharmacol. 2023, 21, 399–423. [Google Scholar] [CrossRef]
- Timoshnikov, V.A.; Selyutina, O.Y.; Polyakov, N.E.; Didichenko, V.; Kontoghiorghes, G.J. Mechanistic Insights of Chelator Complexes with Essential Transition Metals: Antioxidant/Pro-Oxidant Activity and Applications in Medicine. Int. J. Mol. Sci. 2022, 23, 1247. [Google Scholar] [CrossRef] [PubMed]
- Sourabh, S.; Bhatia, P.; Jain, R. Favourable Improvement in Haematological Parameters in Response to Oral Iron and Vitamin C Combination in Children with Iron Refractory Iron Deficiency Anemia (IRIDA) Phenotype. Blood Cells Mol. Dis. 2019, 75, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, C.; Olivares, M.; Brito, A.; Hamilton-West, C.; Pizarro, F. Is a 40% Absorption of Iron from a Ferrous ascorbate Reference Dose Appropriate to Assess Iron Absorption Independent of Iron Status? Biol. Trace Elem. Res. 2013, 155, 322–326. [Google Scholar] [CrossRef] [PubMed]
- Patil, P.; Geevarghese, P.; Khaire, P.; Joshi, T.; Suryawanshi, A.; Mundada, S.; Pawar, S.; Farookh, A. Comparison of Therapeutic Efficacy of Ferrous Ascorbate and Iron Polymaltose Complex in Iron Deficiency Anemia in Children: A Randomized Controlled Trial. Indian J. Pediatr. 2019, 86, 1112–1117. [Google Scholar] [CrossRef] [PubMed]
- Elalfy, M.S.; Saber, M.M.; Adly, A.A.; Ismail, E.A.; Tarif, M.; Ibrahim, F.; Elalfy, O.M. Role of vitamin C as an adjuvant therapy to different iron chelators in young β-thalassemia major patients: Efficacy and safety in relation to tissue iron overload. Eur. J. Haematol. 2016, 96, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Conte, D.; Brunelli, L.; Ferrario, L.; Mandelli, C.; Quatrini, M.; Velio, P.; Bianchi, P.A. Effect of ascorbic acid on desferrioxamine-induced urinary iron excretion in idiopathic hemochromatosis. Acta Haematol. 1984, 72, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Perron, N.R.; Brumaghim, J.L. A review of the antioxidant mechanisms of polyphenol compounds related to iron binding. Cell Biochem. Biophys. 2009, 53, 75–100. [Google Scholar] [CrossRef]
- Leopoldini, M.; Russo, N.; Toscano, M. The molecular basis of working mechanism of natural polyphenolic antioxidants. Food Chem. 2011, 125, 288–306. [Google Scholar] [CrossRef]
- Sajadi Hezaveh, Z.; Azarkeivan, A.; Janani, L.; Hosseini, S.; Shidfar, F. The effect of quercetin on iron overload and inflammation in β-thalassemia major patients: A double-blind randomized clinical trial. Complement. Ther. Med. 2019, 46, 24–28. [Google Scholar] [CrossRef]
- Salehi, B.; Stojanović-Radić, Z.; Matejić, J.; Sharifi-Rad, M.; Anil Kumar, N.V.; Martins, N.; Sharifi-Rad, J. The therapeutic potential of curcumin: A review of clinical trials. Eur. J. Med. Chem. 2019, 163, 527–545. [Google Scholar] [CrossRef]
- Gillessen, A.; Schmidt, H.H. Silymarin as Supportive Treatment in Liver Diseases: A Narrative Review. Adv. Ther. 2020, 37, 1279–1301. [Google Scholar] [CrossRef] [PubMed]
- Neilands, J.B. Siderophores: Structure and function of microbial iron transport compounds. J. Biol. Chem. 1995, 270, 26723–26726. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.R.; Bogdan, A.R.; Miyazawa, M.; Hashimoto, K.; Tsuji, Y. Siderophores in Iron Metabolism: From Mechanism to Therapy Potential. Trends Mol. Med. 2016, 22, 1077–1090. [Google Scholar] [CrossRef]
- Page, M.G.P. The Role of Iron and Siderophores in Infection, and the Development of Siderophore Antibiotics. Clin. Infect. Dis. 2019, 69 (Suppl. 7), S529–S537. [Google Scholar] [CrossRef]
- Sousa, L.; Oliveira, M.M.; Pessôa, M.T.C.; Barbosa, L.A. Iron overload: Effects on cellular biochemistry. Clin. Chim. Acta 2020, 504, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Consoli, V.; Sorrenti, V.; Grosso, S.; Vanella, L. Heme Oxygenase-1 Signaling and Redox Homeostasis in Physiopathological Conditions. Biomolecules 2021, 11, 589. [Google Scholar] [CrossRef] [PubMed]
- Ryter, S.W.; Alam, J.; Choi, A.M. Heme oxygenase-1/carbon monoxide: From basic science to therapeutic applications. Physiol. Rev. 2006, 86, 583–650. [Google Scholar] [CrossRef]
- Grace, R.F.; Glader, B. Red Blood Cell Enzyme Disorders. Pediatr. Clin. N. Am. 2018, 65, 579–595. [Google Scholar] [CrossRef]
- Leung, A.K.C.; Lam, J.; Wong, A.H.C.; Hon, K.L.; Li, X. Iron deficiency anemia: An updated review. Curr. Pediatr. Rev. 2023, 20, 339–356. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Gao, S.; Zhang, X.; Zang, J. Dietary Heme-Containing Proteins: Structures, Applications, and Challenges. Foods 2022, 11, 3594. [Google Scholar] [CrossRef] [PubMed]
- Piskin, E.; Cianciosi, D.; Gulec, S.; Tomas, M.; Capanoglu, E. Iron Absorption: Factors, Limitations, and Improvement Methods. ACS Omega 2022, 7, 20441–20456. [Google Scholar] [CrossRef] [PubMed]
- Bouvard, V.; Loomis, D.; Guyton, K.Z.; Grosse, Y.; El Ghissassi, F.; Benbrahim-Tallaa, L.; Guha, N.; Mattock, H.; Straif, K.; Stewart, B.W.; et al. Carcinogenicity of Consumption of Red and Processed Meat. Lancet Oncol. 2015, 16, 1599–1600. [Google Scholar] [CrossRef] [PubMed]
- Tolosano, E.; Altruda, F. Hemopexin: Structure, Function, and Regulation. DNA Cell Boil. 2002, 21, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Shih, A.W.; McFarlane, A.; Verhovsek, M. Haptoglobin testing in hemolysis: Measurement and interpretation. Am. J. Hematol. 2014, 89, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J. Chelators affecting iron absorption in mice. Arzneimittelforschung 1990, 40, 1332–1335. [Google Scholar] [PubMed]
- Yamamoto, R.S.; Williams, G.M.; Frangel, H.H.; Weisburger, J.H. 8-hydroxyquinoline: Chronic toxicity and inhibitory effect on the carcinogenicity of N-2- fluorenylacetamide. Toxicol. Appl. Pharmacol. 1971, 19, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Fernández, S.; Castro, P.; Nogué, S.; Nicolás, J.M. Acute iron intoxication: Change in urine color during chelation therapy with deferoxamine. Intensive Care Med. 2014, 40, 104. [Google Scholar] [CrossRef] [PubMed]
- Gumber, M.R.; Kute, V.B.; Shah, P.R.; Vanikar, A.V.; Patel, H.V.; Balwani, M.R.; Ghuge, P.P.; Trivedi, H.L. Successful treatment of severe iron intoxication with gastrointestinal decontamination, deferoxamine, and hemodialysis. Ren. Fail. 2013, 35, 729–731. [Google Scholar] [CrossRef]
- White, G.P.; Jacobs, A.; Grady, R.W.; Cerami, A. The effect of chelating agents on iron mobilization in Chang cell cultures. Blood 1976, 48, 923–929. [Google Scholar] [CrossRef]
- Forsbeck, K.; Nilsson, K.; Kontoghiorghes, G.J. Variation in iron accumulation, transferrin membrane binding and DNA synthesis in the K-562 and U-937 cell lines induced by chelators and their iron complexes. Eur. J. Haematol. 1987, 39, 318–325. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. Structure/red blood cell permeability activity of iron (III) chelator complexes. Inorganica Chim. Acta 1988, 151, 101–106. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. The aim of iron chelation therapy in thalassaemia. Eur. J. Haematol. 2017, 99, 465–466. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J. Advances in oral iron chelation in man. Int. J. Hematol. 1992, 55, 27–38. [Google Scholar] [PubMed]
- Aydinok, Y.; Kattamis, A.; Cappellini, M.D.; El-Beshlawy, A.; Origa, R.; Elalfy, M.; Kilinç, Y.; Perrotta, S.; Karakas, Z.; Viprakasit, V.; et al. Effects of deferasirox-deferoxamine on myocardial and liver iron in patients with severe transfusional iron overload. Blood 2015, 125, 3868–3877. [Google Scholar] [CrossRef] [PubMed]
- Elalfy, M.S.; Adly, A.M.; Wali, Y.; Tony, S.; Samir, A.; Elhenawy, Y.I. Efficacy and safety of a novel combination of two oral chelators deferasirox/deferiprone over deferoxamine/deferiprone in severely iron overloaded young beta thalassemia major patients. Eur. J. Haematol. 2015, 95, 411–420. [Google Scholar] [CrossRef]
- Campbell, N.R.; Hasinoff, B.B. Iron supplements: A common cause of drug interactions. Br. J. Clin. Pharmacol. 1991, 31, 251–255. [Google Scholar] [CrossRef]
- Djaldetti, M.; Fishman, P.; Notti, I.; Bessler, H. The effect of tetracycline administration on iron absorption in mice. Biomedicine 1981, 35, 150–152. [Google Scholar] [PubMed]
- Leyden, J.J. Absorption of minocycline hydrochloride and tetracycline hydrochloride: Effect of food, milk, and iron. J. Am. Acad. Dermatol. 1985, 12 Pt 1, 308–312. [Google Scholar] [CrossRef]
- Korać Jačić, J.; Dimitrijević, M.; Bajuk-Bogdanović, D.; Stanković, D.; Savić, S.; Spasojević, I.; Milenković, M.R. The formation of Fe3+-doxycycline complex is pH dependent: Implications to doxycycline bioavailability. J. Biol. Inorg. Chem. 2023, 28, 679–687. [Google Scholar] [CrossRef] [PubMed]
- McQuilten, Z.K.; Thao, L.T.P.; Pasricha, S.R.; Artz, A.S.; Bailey, M.; Chan, A.T.; Cohen, H.J.; Lockery, J.E.; Murray, A.M.; Nelson, M.R.; et al. Effect of Low-Dose Aspirin Versus Placebo on Incidence of Anemia in the Elderly: A Secondary Analysis of the Aspirin in Reducing Events in the Elderly Trial. Ann. Intern. Med. 2023, 176, 913–921, Erratum in: Ann. Intern. Med. 2023, 176, 1288. [Google Scholar] [CrossRef] [PubMed]
- Stěrba, M.; Popelová, O.; Vávrová, A.; Jirkovský, E.; Kovaříková, P.; Geršl, V.; Simůnek, T. Oxidative stress, redox signaling, and metal chelation in anthracycline cardiotoxicity and pharmacological cardioprotection. Antioxid. Redox Signal. 2013, 18, 899–929. [Google Scholar] [CrossRef] [PubMed]
- Perera, D.N.; Palliyaguruge, C.L.; Eapasinghe, D.D.; Liyanage, D.M.; Seneviratne, R.A.C.H.; Demini, S.M.D.; Jayasinghe, J.A.S.M.; Faizan, M.; Rajagopalan, U.; Galhena, B.P.; et al. Factors affecting iron absorption and the role of fortification in enhancing iron levels. Nutr. Bull. 2023, 48, 442–457. [Google Scholar] [CrossRef] [PubMed]
- Hansen, R.; Sejer, E.P.F.; Holm, C.; Schroll, J.B. Iron supplements in pregnant women with normal iron status: A systematic review and meta-analysis. Acta Obstet. Gynecol. Scand. 2023, 102, 1147–1158. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yan, F.; Yan, H.; Wang, Z. Impact of iron supplementation on gestational diabetes mellitus: A literature review. Diabetes Obes. Metab. 2023, 25, 342–353. [Google Scholar] [CrossRef]
- Alaunyte, I.; Stojceska, V.; Plunkett, A. Iron and the female athlete: A review of dietary treatment methods for improving iron status and exercise performance. J. Int. Soc. Sports Nutr. 2015, 12, 38. [Google Scholar] [CrossRef]
- Saldanha, L.G.; Dwyer, J.T.; Andrews, K.W.; Brown, L.L. The Chemical Forms of Iron in Commercial Prenatal Supplements Are Not Always the Same as Those Tested in Clinical Trials. J. Nutr. 2019, 149, 890–893. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. Deferiprone and Iron-Maltol: Forty Years since Their Discovery and Insights into Their Drug Design, Development, Clinical Use and Future Prospects. Int. J. Mol. Sci. 2023, 24, 4970. [Google Scholar] [CrossRef]
- Harvey, R.S.; Reffitt, D.M.; Doig, L.A.; Meenan, J.; Ellis, R.D.; Thompson, R.P.; Powell, J.J. Ferric trimaltol corrects iron deficiency anaemia in patients intolerant of iron. Aliment. Pharmacol. Ther. 1998, 12, 845–848. [Google Scholar] [CrossRef]
- Reffitt, D.M.; Burden, T.J.; Seed, P.T.; Wood, J.; Thompson, R.P.; Powell, J.J. Assessment of iron absorption from ferric trimaltol. Ann. Clin. Biochem. 2000, 37, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Stallmach, A.; Büning, C. Ferric maltol (ST10): A novel oral iron supplement for the treatment of iron deficiency anemia in inflammatory bowel disease. Expert Opin. Pharmacother. 2015, 16, 2859–2867. [Google Scholar] [CrossRef] [PubMed]
- Khoury, A.; Pagan, K.A.; Farland, M.Z. Ferric Maltol: A New Oral Iron Formulation for the Treatment of Iron Deficiency in Adults. Ann. Pharmacother. 2021, 55, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Bokemeyer, B.; Krummenerl, A.; Maaser, C.; Howaldt, S.; Mroß, M.; Mallard, N. Randomized Open-Label Phase 1 Study of the Pharmacokinetics of Ferric Maltol in Inflammatory Bowel Disease Patients with Iron Deficiency. Eur. J. Drug Metab. Pharmacokinet. 2017, 42, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Gasche, C.; Ahmad, T.; Tulassay, Z.; Baumgart, D.C.; Bokemeyer, B.; Büning, C.; Howaldt, S.; Stallmach, A. AEGIS Study Group. Ferric maltol is effective in correcting iron deficiency anemia in patients with inflammatory bowel disease: Results from a phase-3 clinical trial program. Inflamm. Bowel Dis. 2015, 21, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.; Ahmad, T.; Tulassay, Z.; Baumgart, D.C.; Bokemeyer, B.; Howaldt, S.; Stallmach, A.; Büning, C.; the AEGIS Study Group. Ferric maltol therapy for iron deficiency anaemia in patients with inflammatory bowel disease: Long-term extension data from a Phase 3 study. Aliment. Pharmacol. Ther. 2016, 44, 259–270. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kopyt, N.P. AEGIS-CKD Study Group. Efficacy and safety of oral ferric maltol (FM) in treating iron-deficiency anemia (IDA) in patients with CKD: Randomized controlled trial [FR-OR120]. J. Am. Soc. Nephrol. 2018, 29, 70–71. [Google Scholar]
- Olsson, K.M.; Fuge, J.; Brod, T.; Kamp, J.C.; Schmitto, J.; Kempf, T.; Bauersachs, J.; Hoeper, M.M. Oral iron supplementation with ferric maltol in patients with pulmonary hypertension. Eur. Respir. J. 2020, 56, 2000616. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; May, A. Uptake and intracellular distribution of iron from transferrin and chelators in erythroid cells. BioMetals 1990, 3, 183–187. [Google Scholar] [CrossRef]
- Nataatmadja, M.S.; Francis, R. Recurrent severe hypophosphatemia following intravenous iron administration. Clin. Case Rep. 2020, 8, 243–246. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kshirsagar, A.V.; Li, X. Long-Term Risks of Intravenous Iron in End-Stage Renal Disease Patients. Adv. Chronic Kidney Dis. 2019, 26, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Punj, S.; Ghafourian, K.; Ardehali, H. Iron deficiency and supplementation in heart failure and chronic kidney disease. Mol. Asp. Med. 2020, 75, 100873. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Liabeuf, S.; Gras, V.; Moragny, J.; Laroche, M.L.; Andrejak, M.; French National Network of Pharmacovigilance Centers. Ulceration of the oral mucosa following direct contact with ferrous sulfate in elderly patients: A case report and a review of the French National Pharmacovigilance Database. Clin. Interv. Aging 2014, 9, 737–740. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ferrari, P.; Nicolini, A.; Manca, M.L.; Rossi, G.; Anselmi, L.; Conte, M.; Carpi, A.; Bonino, F. Treatment of mild non-chemotherapy-induced iron deficiency anemia in cancer patients: Comparison between oral ferrous bisglycinate chelate and ferrous sulfate. Biomed. Pharmacother. 2012, 66, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J. Drug Selection and Posology, Optimal Therapies and Risk/Benefit Assessment in Medicine: The Paradigm of Iron-Chelating Drugs. Int. J. Mol. Sci. 2023, 24, 16749. [Google Scholar] [CrossRef] [PubMed]
- Aoyagi, N.; Kimura, R.; Murata, T. Studies on passiflora incarnata dry extract. I. Isolation of maltol and pharmacological action of maltol and ethyl maltol. Chem. Pharm. Bull. 1974, 22, 1008–1013. [Google Scholar] [CrossRef] [PubMed]
- Rohleder, A.R.; Scherf, K.A.; Schieberle, P.; Koehler, P. Quantitative Analyses of Key Odorants and Their Precursors Reveal Differences in the Aroma of Gluten-Free Rice Bread and Wheat Bread. J. Agric. Food Chem. 2019, 67, 11179–11186. [Google Scholar] [CrossRef] [PubMed]
- Rennhard, H.H. The metabolism of ethyl maltol and maltol in the dog. J. Agric. Food Chem. 1971, 19, 152–154. [Google Scholar] [CrossRef] [PubMed]
- Chitambar, C.R. Gallium Complexes as Anticancer Drugs. Met. Ions Life Sci. 2018, 18, 281–301. [Google Scholar] [CrossRef]
- Arnold, C.; Chaffin, M.K.; Cohen, N.; Fajt, V.R.; Taylor, R.J.; Bernstein, L.R. Pharmacokinetics of gallium maltolate after intragastric administration in adult horses. Am. J. Vet. Res. 2010, 71, 1371–1376. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Kleanthous, M.; Kontoghiorghe, C.N. The History of Deferiprone (L1) and the Paradigm of the Complete Treatment of Iron Overload in Thalassaemia. Mediterr. J. Hematol. Infect. Dis. 2020, 12, e2020011. [Google Scholar] [CrossRef] [PubMed]
- Kolnagou, A.; Michaelides, Y.; Kontos, C.; Kyriacou, K.; Kontoghiorghes, G.J. Myocyte damage and loss of myofibers is the potential mechanism of iron overload toxicity in congestive cardiac failure in thalassemia. Complete reversal of the cardiomyopathy and normalization of iron load by deferiprone. Hemoglobin 2008, 32, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Zurlo, M.G.; De Stefano, P.; Borgna-Pignatti, C.; Di Palma, A.; Piga, A.; Melevendi, C.; Di Gregorio, F.; Burattini, M.G.; Terzoli, S. Survival and causes of death in thalassaemia major. Lancet 1989, 334, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Modell, B.; Khan, M.; Darlison, M.; Westwood, M.A.; Ingram, D.; Pennell, D.J.; Modell, B.; Khan, M.; Darlison, M.; Westwood, M.A.; et al. Improved survival of thalassaemia major in the UK and relation to T2* cardiovascular magnetic resonance. J. Cardiovasc. Magn. Reson. 2008, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Telfer, P.; Coen, P.G.; Christou, S.; Hadjigavriel, M.; Kolnakou, A.; Pangalou, E.; Pavlides, N.; Psiloines, M.; Simamonian, K.; Skordos, G.; et al. Survival of medically treated thalassemia patients in Cyprus. Trends and risk factors over the period 1980–2004. Haematologica 2006, 91, 1187–1192. [Google Scholar] [PubMed]
- Au, W.Y.; Lee, V.; Lau, C.W.; Yau, J.; Chan, D.; Chan, E.Y.; Cheung, W.W.; Ha, S.Y.; Kho, B.; Lee, C.Y.; et al. A synopsis of current care of thalassaemia major patients in Hong Kong. Hong Kong Med. J. 2011, 17, 261–266. [Google Scholar]
- Maggio, A.; Filosa, A.; Vitrano, A.; Aloj, G.; Kattamis, A.; Ceci, A.; Fucharoen, S.; Cianciulli, P.; Grady, R.W.; Prossomariti, L.; et al. Iron chelation therapy in thalassemia major: A systematic review with meta-analyses of 1520 patients included on randomized clinical trials. Blood Cells Mol. Dis. 2011, 47, 166–175. [Google Scholar] [CrossRef]
- Verma, I.C. Burden of genetic disorders in India. Indian J. Pediatr. 2000, 67, 893–898. [Google Scholar] [CrossRef]
- Teawtrakul, N.; Chansung, K.; Sirijerachai, C.; Wanitpongpun, C.; Thepsuthammarat, K. The impact and disease burden of thalassemia in Thailand: A population-based study in 2010. J. Med. Assoc. Thail. 2012, 95, 95. [Google Scholar]
- Kolnagou, A.; Kontoghiorghe, C.N.; Kontoghiorghes, G.J. Transition of Thalassaemia and Friedreich ataxia from fatal to chronic diseases. World J. Methodol. 2014, 4, 197–218. [Google Scholar] [CrossRef]
- Fragodimitri, C.; Schiza, V.; Giakoumis, A.; Drakaki, K.; Salichou, A.; Karampatsos, F.; Yousef, J.; Karageorga, M.; Berdoukas, V.; Aessopos, A. Successful chelation in beta-thalassemia major in the 21st century. Medicine 2023, 102, e35455. [Google Scholar] [CrossRef] [PubMed]
- Aessopos, A.; Kati, M.; Farmakis, D.; Polonifi, E.; Deftereos, S.; Tsironi, M. Intensive chelation therapy in beta-thalassemia and possible adverse cardiac effects of desferrioxamine. Int. J. Hematol. 2007, 86, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Kolnagou, A.; Peng, C.T.; Shah, S.V.; Aessopos, A. Safety issues of iron chelation therapy in patients with normal range iron stores including thalassaemia, neurodegenerative, renal and infectious diseases. Expert Opin. Drug Saf. 2010, 9, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Mavrogeni, S.I.; Gotsis, E.D.; Markussis, V.; Tsekos, N.; Politis, C.; Vretou, E.; Kermastinos, D. T2 relaxation time study of iron overload in b-thalassemia. Magn. Reson. Mater. Phys. Biol. Med. 1998, 6, 7–12. [Google Scholar] [CrossRef]
- Kolnagou, A.; Kleanthous, M.; Kontoghiorghes, G.J. Reduction of body iron stores to normal range levels in thalassaemia by using a deferiprone/deferoxamine combination and their maintenance thereafter by deferiprone monotherapy. Eur. J. Haematol. 2010, 85, 430–438. [Google Scholar] [CrossRef]
- Farmaki, K.; Tzoumari, I.; Pappa, C.; Chouliaras, G.; Berdoukas, V. Normalisation of total body iron load with very intensive combined chelation reverses cardiac and endocrine complications of thalassaemia major. Br. J. Haematol. 2010, 148, 466–475. [Google Scholar] [CrossRef]
- Kolnagou, A.; Kontoghiorghe, C.N.; Kontoghiorghes, G.J. Prevention of Iron Overload and Long Term Maintenance of Normal Iron Stores in Thalassaemia Major Patients using Deferiprone or Deferiprone Deferoxamine Combination. Drug Res. 2017, 67, 404–411. [Google Scholar] [CrossRef]
- Kolnagou, A.; Kontoghiorghes, G.J. Chelation protocols for the elimination and prevention of iron overload in thalassaemia. Front. Biosci. 2018, 23, 1082–1098. [Google Scholar]
- Karimi, M.; Haghpanah, S.; Bahoush, G.; Ansari, S.; Azarkeivan, A.; Shahsavani, A.; Bazrafshan, A.; Jangjou, A. Evaluation of Efficacy, Safety, and Satisfaction Taking Deferasirox Twice Daily Versus Once Daily in Patients With Transfusion-Dependent Thalassemia. J. Pediatr. Hematol. Oncol. 2020, 42, 23–26. [Google Scholar] [CrossRef]
- Pepe, A.; Meloni, A.; Pistoia, L.; Cuccia, L.; Gamberini, M.R.; Lisi, R.; D’Ascola, D.G.; Rosso, R.; Allò, M.; Spasiano, A.; et al. MRI multicentre prospective survey in thalassaemia major patients treated with deferasirox versus deferiprone and desferrioxamine. Br. J. Haematol. 2018, 183, 783–795. [Google Scholar] [CrossRef]
- Lin, C.H.; Chen, X.; Wu, C.C.; Wu, K.H.; Song, T.S.; Weng, T.F.; Hsieh, Y.W.; Peng, C.T. Therapeutic mechanism of combined oral chelation therapy to maximize efficacy of iron removal in transfusion-dependent thalassemia major—A pilot study. Expert Rev. Hematol. 2019, 12, 265–272. [Google Scholar] [CrossRef]
- Parakh, N.; Chandra, J.; Sharma, S.; Dhingra, B.; Jain, R.; Mahto, D. Efficacy and Safety of Combined Oral Chelation With Deferiprone and Deferasirox in Children With β-Thalassemia Major: An Experience From North India. J. Pediatr. Hematol. Oncol. 2017, 39, 209–213. [Google Scholar] [CrossRef]
- Eghbali, A.; Shokri, P.; Afzal, R.R.; Bagheri, B. A 1-year randomized trial of deferasirox alone versus deferasirox and deferoxamine combination for the treatment of iron overload in thalassemia major. Transfus. Apher. Sci. 2019, 58, 429–433. [Google Scholar] [CrossRef]
- Anonymous. Exjade (Deferasirox) Tablets for Oral Suspension. Prescribing Information. (Novartis Pharmaceutical Corporation USA (NDA 21–882). October 2006; pp. 1–12. Changes to the Warnings and Adverse Reactions Sections. Available online: http://www.fda.gov/ohrms/dockets/ac/05/briefing/2005-4177B1_02_b.pdf (accessed on 1 December 2015).
- Anonymous. Exjade (Deferasirox) Tablets for Oral Suspension (Highlights of Prescribing Information. Novartis Pharmaceutical Corp. USA (T2011-106). August 2011; pp. 1–16). Available online: http://www.accessdata.fda.gov/drugsatfda_docs/label/2010/021882s010lbl.pdf (accessed on 1 December 2015).
- Kontoghiorghes, G.J. A record of 1320 suspect, deferasirox-related, patient deaths reported in 2009: Insufficient toxicity testing, low efficacy and lack of transparency may endanger the lives of iron loaded patients. Hemoglobin 2011, 35, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J. A record number of fatalities in many categories of patients treated with deferasirox: Loopholes in regulatory and marketing procedures undermine patient safety and misguide public funds? Expert Opin. Drug Saf. 2013, 12, 605–609. [Google Scholar] [CrossRef]
- Badeli, H.; Baghersalimi, A.; Eslami, S.; Saadat, F.; Rad, A.H.; Basavand, R.; Papkiadeh, S.R.; Darbandi, B.; Kooti, W.; Peluso, I. Early Kidney Damage Markers after Deferasirox Treatment in Patients with Thalassemia Major: A Case-Control Study. Oxidative Med. Cell. Longev. 2019, 2019, 5461617. [Google Scholar] [CrossRef] [PubMed]
- Boelaert, J.R.; Fenves, A.Z.; Coburn, J.W. Deferoxamine therapy and mucormycosis in dialysis patients: Report of an international registry. Am. J. Kidney Dis. 1991, 18, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Baath, J.S.; Lam, W.C.; Kirby, M.; Chun, A. Deferoxamine-related ocular toxicity: Incidence and outcome in a pediatric population. Retina 2008, 28, 894–899. [Google Scholar] [CrossRef]
- Levine, J.E.; Cohen, A.; MacQueen, M.; Martin, M.; Giardina, P.J. Sensorimotor neurotoxicity associated with high-dose deferoxamine treatment. J. Pediatr. Hematol. Oncol. 1997, 19, 139–141. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.B.; Rosenwasser, L.J.; Bessette, J.A.; Beer, D.J.; Rocklin, R.E. Rapid desensitisation for desferrioxamine anaphylactic reaction. Lancet 1981, 317, 1059. [Google Scholar] [CrossRef]
- Robins-Browne, R.M.; Pipic, J.K. Desferrioxamine and systemic yersiniosis. Lancet 1983, 322, 1372. [Google Scholar] [CrossRef] [PubMed]
- Chan, G.C.; Ng, D.M.; Fong, D.Y.; Ha, S.Y.; Lau, Y.L. Comparison of subcutaneous infusion needles for transfusion-dependent thalassemia patients by the intrapersonal cross-over assessment model. Am. J. Hematol. 2004, 76, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.R.; Galanello, R.; Piga, A.; Dipalma, A.; Vullo, C.; Tricta, F. Safety profile of the oral iron chelator deferiprone: A multicentre study. Br. J. Haematol. 2000, 108, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Boddaert, N.; Le Quan Sang, K.H.; Rötig, A.; Leroy-Willig, A.; Gallet, S.; Brunelle, F.; Sidi, D.; Thalabard, J.C.; Munnich, A.; Cabantchik, Z.I. Selective iron chelation in Friedreich ataxia: Biologic and clinical implications. Blood 2007, 110, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Abbruzzese, G.; Cossu, G.; Balocco, M.; Marchese, R.; Murgia, D.; Melis, M.; Galanello, R.; Barella, S.; Matta, G.; Ruffinengo, U.; et al. A pilot trial of deferiprone for neurodegeneration with brain iron accumulation. Haematologica 2011, 96, 1708–1711. [Google Scholar] [CrossRef] [PubMed]
- Martin-Bastida, A.; Ward, R.J.; Newbould, R.; Piccini, P.; Sharp, D.; Kabba, C.; Patel, M.C.; Spino, M.; Connelly, J.; Tricta, F.; et al. Brain iron chelation by deferiprone in a phase 2 randomised double-blinded placebo controlled clinical trial in Parkinson’s disease. Sci. Rep. 2017, 7, 1398. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Barr, J.; Baillod, R.A. Studies of aluminium mobilization in renal dialysis patients using the oral chelator 1,2-dimethyl-3-hydroxypyrid-4-one. Arzneimittelforschung 1994, 44, 522–526. [Google Scholar] [PubMed]
- Romano, N.; Baiardi, G.; Pinto, V.M.; Quintino, S.; Gianesin, B.; Sasso, R.; Diociasi, A.; Mattioli, F.; Marchese, R.; Abbruzzese, G.; et al. Long-Term Neuroradiological and Clinical Evaluation of NBIA Patients Treated with a Deferiprone Based Iron-Chelation Therapy. J. Clin. Med. 2022, 11, 4524. [Google Scholar] [CrossRef] [PubMed]
- Rajapurkar, M.M.; Hegde, U.; Bhattacharya, A.; Alam, M.G.; Shah, S.V. Effect of deferiprone, an oral iron chelator, in diabetic and non-diabetic glomerular disease. Toxicol. Mech. Methods 2013, 23, 5–10. [Google Scholar] [CrossRef]
- Mohanty, D.; Ghosh, K.; Pathare, A.V.; Karnad, D. Deferiprone (L1) as an adjuvant therapy for Plasmodium falciparum malaria. Indian J. Med. Res. 2002, 115, 17–21. [Google Scholar]
- Saxena, D.; Spino, M.; Tricta, F.; Connelly, J.; Cracchiolo, B.M.; Hanauske, A.R.; D’Alliessi Gandolfi, D.; Mathews, M.B.; Karn, J.; Holland, B.; et al. Drug-Based Lead Discovery: The Novel Ablative Antiretroviral Profile of Deferiprone in HIV-1-Infected Cells and in HIV-Infected Treatment-Naive Subjects of a Double-Blind, Placebo-Controlled, Randomized Exploratory Trial. PLoS ONE 2016, 11, e0154842. [Google Scholar] [CrossRef]
- Vreugdenhil, G.; Swaak, A.J.; Kontoghiorghes, G.J.; van Eijk, H.G. Efficacy and safety of oral iron chelator L1 in anaemic rheumatoid arthritis patients. Lancet 1989, 334, 1398–1399. [Google Scholar] [CrossRef] [PubMed]
- Devos, D.; Labreuche, J.; Rascol, O.; Corvol, J.C.; Duhamel, A.; Guyon Delannoy, P.; Poewe, W.; Compta, Y.; Pavese, N.; Růžička, E.; et al. Trial of Deferiprone in Parkinson’s Disease. N. Engl. J. Med. 2022, 387, 2045–2055. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J. The Vital Role Played by Deferiprone in the Transition of Thalassaemia from a Fatal to a Chronic Disease and Challenges in Its Repurposing for Use in Non-Iron-Loaded Diseases. Pharmaceuticals 2023, 16, 1016. [Google Scholar] [CrossRef] [PubMed]
- Petry, N.; Egli, I.; Zeder, C.; Walczyk, T.; Hurrell, R. Polyphenols and phytic acid contribute to the low iron bioavailability from common beans in young women. J. Nutr. 2010, 140, 1977–1982. [Google Scholar] [CrossRef] [PubMed]
- Iyengar, V.; Pullakhandam, R.; Nair, K.M. Dietary ligands as determinants of iron-zinc interactions at the absorptive enterocyte. J. Food Sci. 2010, 75, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Sotelo, A.; González-Osnaya, L.; Sánchez-Chinchillas, A.; Trejo, A. Role of oxate, phytate, tannins and cooking on iron bioavailability from foods commonly consumed in Mexico. Int. J. Food Sci. Nutr. 2010, 61, 29–39. [Google Scholar] [CrossRef]
- Jaramillo, Á.; Briones, L.; Andrews, M.; Arredondo, M.; Olivares, M.; Brito, A.; Pizarro, F. Effect of phytic acid, tannic acid and pectin on fasting iron bioavailability both in the presence and absence of calcium. J. Trace Elem. Med. Biol. 2015, 30, 112–117. [Google Scholar] [CrossRef]
- Thompson, K.H.; Barta, C.A.; Orvig, C. Metal complexes of maltol and close analogues in medicinal inorganic chemistry. Chem. Soc. Rev. 2006, 35, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Barve, A.; Kumbhar, A.; Bhat, M.; Joshi, B.; Butcher, R.; Sonawane, U.; Joshi, R. Mixed-ligand copper(II) maltolate complexes: Synthesis, characterization, DNA binding and cleavage, and cytotoxicity. Inorg. Chem. 2009, 48, 9120–9132. [Google Scholar] [CrossRef] [PubMed]
- Kosuru, R.Y.; Roy, A.; Das, S.K.; Bera, S. Gallic Acid and Gallates in Human Health and Disease: Do Mitochondria Hold the Key to Success? Mol. Nutr. Food Res. 2018, 62, 1700699. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.A.; Perkins, A.C.; Walton, P.H. 99mTc-labelled human serum transferrin for tumour imaging: An in vitro and in vivo study of the complex. Nucl. Med. Commun. 2004, 25, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Tsopelas, C. A study of radiogallium aqueous chemistry: In vitro and in vivo characterisation of (67) Ga-hydrolysed-stannous fluoride particles. J. Label. Compd. Radiopharm. 2016, 59, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Dahlqvist, G.E.; Jamar, F.; Zech, F.; Geubel, A.P. In-111 transferrin scintigraphy in cirrhosis with hypoalbuminemia: Evidence for protein-losing enteropathy in a small group of selected cases. Scand. J. Gastroenterol. 2012, 47, 1247–1252. [Google Scholar] [CrossRef] [PubMed]
- Sava, G.; Pacor, S.; Bregant, F.; Ceschia, V. Metal complexes of ruthenium: A potential class of selective anticancer drugs. Anticancer. Res. 1991, 11, 1103–1107. [Google Scholar] [PubMed]
- Doroshow, J.H. Dexrazoxane for the prevention of cardiac toxicity and treatment of extravasation injury from the anthracycline antibiotics. Curr. Pharm. Biotechnol. 2012, 13, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, P.; Barootkoob, B.; ElHashash, A.; Nair, A. Efficacy of Dexrazoxane in Cardiac Protection in Pediatric Patients Treated With Anthracyclines. Cureus 2023, 15, e37308. [Google Scholar] [CrossRef] [PubMed]
- Mody, H.; Vaidya, T.R.; Ait-Oudhia, S. In vitro to clinical translational pharmacokinetic/pharmacodynamic modeling of doxorubicin (DOX) and dexrazoxane (DEX) interactions: Safety assessment and optimization. Sci. Rep. 2023, 13, 3100. [Google Scholar] [CrossRef]
- de Baat, E.C.; van Dalen, E.C.; Mulder, R.L.; Hudson, M.M.; Ehrhardt, M.J.; Engels, F.K.; Feijen, E.A.M.; Grotenhuis, H.B.; Leerink, J.M.; Kapusta, L.; et al. Primary cardioprotection with dexrazoxane in patients with childhood cancer who are expected to receive anthracyclines: Recommendations from the International Late Effects of Childhood Cancer Guideline Harmonization Group. Lancet Child Adolesc. Health 2022, 6, 885–894. [Google Scholar] [CrossRef]
- Niewerth, M.; Kunze, D.; Seibold, M.; Schaller, M.; Korting, H.C.; Hube, B. Ciclopirox olamine treatment affects the expression pattern of Candida albicans genes encoding virulence factors, iron metabolism proteins, and drug resistance factors. Antimicrob. Agents Chemother. 2003, 47, 1805–1817. [Google Scholar] [CrossRef]
- Shen, T.; Huang, S. Repositioning the Old Fungicide Ciclopirox for New Medical Uses. Curr. Pharm. Des. 2016, 22, 4443–4450. [Google Scholar] [CrossRef] [PubMed]
- Minden, M.D.; Hogge, D.E.; Weir, S.J.; Kasper, J.; Webster, D.A.; Patton, L.; Jitkova, Y.; Hurren, R.; Gronda, M.; Goard, C.A.; et al. Oral ciclopirox olamine displays biological activity in a phase I study in patients with advanced hematologic malignancies. Am. J. Hematol. 2014, 89, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Born, T.; Kontoghiorghe, C.N.; Spyrou, A.; Kolnagou, A.; Kontoghiorghes, G.J. EDTA chelation reappraisal following new clinical trials and regular use in millions of patients: Review of preliminary findings and risk/benefit assessment. Toxicol. Mech. Methods 2013, 23, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Konstantinou, E.; Pashalidis, I.; Kolnagou, A.; Kontoghiorghes, G.J. Interactions of hydroxycarbamide (hydroxyurea) with iron and copper: Implications on toxicity and therapeutic strategies. Hemoglobin 2011, 35, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Lavelle, D.; Engel, J.D.; Saunthararajah, Y. Fetal Hemoglobin Induction by Epigenetic Drugs. Semin. Hematol. 2018, 55, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Ehsani, M.A.; Hedayati-Asl, A.A.; Bagheri, A.; Zeinali, S.; Rashidi, A. Hydroxyurea-induced hematological response in transfusion-independent beta-thalassemia intermedia: Case series and review of literature. Pediatr. Hematol. Oncol. 2009, 26, 560–565. [Google Scholar] [CrossRef]
- Fraser, D.I.; Liu, K.T.; Reid, B.J.; Hawkins, E.; Sevier, A.; Pyle, M.; Robinson, J.W.; Ouellette, P.H.; Ballantyne, J.S. Widespread Natural Occurrence of Hydroxyurea in Animals. PLoS ONE 2015, 10, e0142890. [Google Scholar] [CrossRef]
- Shah, S.; Sheth, R.; Shah, K.; Patel, K. Safety and effectiveness of thalidomide and hydroxyurea combination in β-thalassaemia intermedia and major: A retrospective pilot study. Br. J. Haematol. 2020, 188, e18–e21. [Google Scholar] [CrossRef]
- Pinto, V.M.; Romano, N.; Balocco, M.; Carrara, P.; Lamagna, M.; Quintino, S.; Castaldi, A.; Forni, G.L. Reduction of extramedullary erythropoiesis and amelioration of anemia in a β-thalassemia patient treated with thalidomide. Am. J. Hematol. 2023, 99, 463–464. [Google Scholar] [CrossRef]
- Jain, M.; Chakrabarti, P.; Dolai, T.K.; Ghosh, P.; Mandal, P.K.; Baul, S.N.; De, R. Comparison of efficacy and safety of thalidomide vs. hydroxyurea in patients with Hb E-β thalassemia—A pilot study from a tertiary care Centre of India. Blood Cells Mol. Dis. 2021, 88, 102544. [Google Scholar] [CrossRef]
- Garbowski, M.W.; Ugidos, M.; Risueño, A.; Shetty, J.K.; Schwickart, M.; Hermine, O.; Porter, J.B.; Thakurta, A.; Vodala, S. Luspatercept stimulates erythropoiesis, increases iron utilization, and redistributes body iron in transfusion-dependent thalassemia. Am. J. Hematol. 2024, 99, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Denton, C.C.; Vodala, S.; Veluswamy, S.; Hofstra, T.C.; Coates, T.D.; Wood, J.C. Splenic iron decreases without change in volume or liver parameters during luspatercept therapy. Blood 2023, 142, 1932–1934. [Google Scholar] [CrossRef] [PubMed]
- Platzbecker, U.; Della Porta, M.G.; Santini, V.; Zeidan, A.M.; Komrokji, R.S.; Shortt, J.; Valcarcel, D.; Jonasova, A.; Dimicoli-Salazar, S.; Tiong, I.S.; et al. Efficacy and safety of luspatercept versus epoetin alfa in erythropoiesis-stimulating agent-naive, transfusion-dependent, lower-risk myelodysplastic syndromes (COMMANDS): Interim analysis of a phase 3, open-label, randomised controlled trial. Lancet 2023, 402, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Longo, F.; Motta, I.; Pinto, V.; Piolatto, A.; Ricchi, P.; Tartaglione, I.; Origa, R. Treating Thalassemia Patients with Luspatercept: An Expert Opinion Based on Current Evidence. J. Clin. Med. 2023, 12, 2584. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Tuffour, A.; Hao, G.; Peprah, F.A.; Huang, A.; Zhou, Y.; Zhang, H. Distinctive modulation of hepcidin in cancer and its therapeutic relevance. Front. Oncol. 2023, 13, 1141603. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhao, J.; Feng, H. Targeting iron metabolism in osteosarcoma. Discov. Oncol. 2023, 14, 31. [Google Scholar] [CrossRef]
- Scheers, N.; Sandberg, A.S. Iron Transport through Ferroportin Is Induced by Intracellular Ascorbate and Involves IRP2 and HIF2α. Nutrients 2014, 6, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Cajander, S.; Kox, M.; Scicluna, B.P.; Weigand, M.A.; Mora, R.A.; Flohé, S.B.; Martin-Loeches, I.; Lachmann, G.; Girardis, M.; Garcia-Salido, A.; et al. Profiling the dysregulated immune response in sepsis: Overcoming challenges to achieve the goal of precision medicine. Lancet Respir. Med. 2023, 12, 305–322. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Fetta, S.; Kontoghiorghe, C.N. The need for a multi-level drug targeting strategy to curb the COVID-19 pandemic. Front. Biosci. 2021, 26, 1723–1736. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Barr, J.; Nortey, P.; Sheppard, L. Selection of a new generation of orally active alpha-ketohydroxypyridine iron chelators intended for use in the treatment of iron overload. Am. J. Hematol. 1993, 42, 340–349. [Google Scholar] [CrossRef]
- Lestón Vázquez, M.; Vilaplana-Carnerero, C.; Gomez-Lumbreras, A.; Prat-Vallverdu, O.; Marsal, J.R.; Vedia Urgell, C.; Giner-Soriano, M.; Morros, R. Drug exposure during pregnancy in primary care: An algorithm and observational study from SIDIAP database, Catalunya, Spain. BMJ Open 2023, 13, e071335. [Google Scholar] [CrossRef] [PubMed]
- Bellanti, F.; Del Vecchio, G.C.; Putti, M.C.; Cosmi, C.; Fotzi, I.; Bakshi, S.D.; Danhof, M.; Della Pasqua, O. Model-Based Optimisation of Deferoxamine Chelation Therapy. Pharm. Res. 2016, 33, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Ferih, K.; Elsayed, B.; Elshoeibi, A.M.; Elsabagh, A.A.; Elhadary, M.; Soliman, A.; Abdalgayoom, M.; Yassin, M. Applications of Artificial Intelligence in Thalassemia: A Comprehensive Review. Diagnostics 2023, 13, 1551. [Google Scholar] [CrossRef] [PubMed]
- Mayer, B.; Kringel, D.; Lötsch, J. Artificial intelligence and machine learning in clinical pharmacological research. Expert Rev. Clin. Pharmacol. 2024, 17, 79–91. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Specific ferric iron–chelator complex drugs Ferric iron tri-maltol. Ferric iron bis-glycinate chelate. |
| Ferric iron complex drug formulations Ferric fumarate, ferric polymaltose, iron dextran, ferric iron sucrose, ferric gluconate, ferric saccharate. |
| Ferric iron complex intravenous drug formulations Ferric iron sucrose, ferric gluconate, ferric carboxymaltose, iron isomaltoside-1000, ferumoxytol, iron dextran (low-molecular-weight forms). |
| Ferrous iron complex drug formulations Ferrous sulphate, ferrous ascorbate, ferrous fumarate, ferrous gluconate, ferroglycine sulphate. |
| Dietary iron complexes Haem (mostly in meat and blood food products). Non-haem iron complexes (mostly in vegetarian food products). Inorganic and organic iron complexes as food supplements and in food fortification. |
| Iron-containing proteins (iron complex prosthetic group composition—function) Haemoglobin (haem—oxygen transport). Myoglobin (haem—oxygen storage). Cytochromes (haem—electron transport; respiration). Cytochrome P450 (haem—drug detoxification). Ribonucleotide reductase (amino acid–iron complex—DNA synthesis). Proline hydroxylase (amino acid–iron complex—collagen synthesis). Phenylalanine hydroxylase (amino acid–iron complex—Tyrosine synthesis, deficit associated with phenylketonuria). Tryptophan 2,3-dioxygenage (haem—degradation of tryptophan). Homogentisic acid 2,3-dioxygenase (amino acid–iron complex—degradation of homogentisate in the tyrosine catabolic pathway, deficit associated with alkaptonuria). Peroxidases (haem—decomposition of hydroperoxides and use of peroxides for oxidation of diverse products, e.g., glutathione, iodide and other biomolecules). Catalase (haem—decomposition of hydrogen peroxide). Lipoxygenase (amino acid–iron complex—HPETE and leukotriene synthesis). Cyclooxygenase (haem and amino acid–iron complex—Prostaglandin and thromboxane synthesis). Adrenodoxin (iron–sulphur complex—electron transport; oxidation/reduction). Aconitase (iron–sulphur complex—Tricarboxylic acid cycle). Succinate dehydrogenase (iron–sulphur complex—Tricarboxylic acid cycle). NADH dehydrogenase (iron–sulphur complex—electron transport; respiration). Xanthine oxidase (iron–sulphur complex—conversion of xanthine to uric acid). Aldehyde oxidase (iron–sulphur complex—metabolism of aldehydes). Transferrin (amino acid–iron complex—iron transport in plasma). Lactoferrin (amino acids–iron complex—iron binding in milk and other secretions). Ferritin (Oxohydroxide, phosphate iron complex—iron storage). Haemosiderin (Oxohydroxide, phosphate iron complex—iron storage). |
| Iron-chelating drugs for the treatment of iron overload Deferiprone, deferoxamine, deferasirox [11,149,150,151,152,153]. |
| Drugs with iron-chelation capacity used in other diseases Hydroxyurea (anticancer), doxorubicin (anticancer), tetracyclines (antibacterial), ciclopirox (antifungal), EDTA (alternative medicine), DTPA (radioactive metal decontamination) [294,295,296,297,298,299,300,301,302,303,304,305,306]. |
| Iron-chelating pro-drugs used in other diseases Dexrazoxane (doxorubicin and other anthracycline toxicity), aspirin (anti-inflammatory) [302,303,304,305]. |
| Iron-chelating proteins Transferrin (blood), lactoferrin (milk and other secretions; neutrophils) [11,91,92,93,94,95,150,151,152,153]. |
| Examples of endogenous low-molecular-weight chelators Phosphates: Pyridoxal phosphate, thiamine pyrophosphate, ribonucleoside and deoxyribonucleoside phosphates, phytic acid (IP6), pyrophosphate, ATP, ADP, AMP, etc. Amino acids: Aspartic acid, glutamic acid, histidine, cysteine, tyrosine, etc. Carboxylic acids: Citric acid, aconitic acid, oxaloacetic acid, etc. Mono- and disaccharides: Fructose, glucose, lactose, etc. Fatty acids and phosphoglycerides. Other naturally occurring chelators: Catecholamines, pteridines, purines, spermine, spermidine, glutathione, folic acid, etc. [11,20,96]. |
| Examples of exogenous dietary low-molecular-weight chelators Polyphenols and other phytochelators: Gallic acid, caffeic acid, quercetin, ellagic acid, curcumin, catechin, maltol, ascorbic acid, etc. [90,176,177,178,179,180]. |
| Vitamins Ascorbic acid, lipoic acid, riboflavin [168,169,170,171,172,173,174,175]. |
| Examples of microbial siderophores Enterobacactin, mycobactin, aspergillic acid, etc. [149,150,181,182,183]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kontoghiorghes, G.J. The Importance and Essentiality of Natural and Synthetic Chelators in Medicine: Increased Prospects for the Effective Treatment of Iron Overload and Iron Deficiency. Int. J. Mol. Sci. 2024, 25, 4654. https://doi.org/10.3390/ijms25094654
Kontoghiorghes GJ. The Importance and Essentiality of Natural and Synthetic Chelators in Medicine: Increased Prospects for the Effective Treatment of Iron Overload and Iron Deficiency. International Journal of Molecular Sciences. 2024; 25(9):4654. https://doi.org/10.3390/ijms25094654
Chicago/Turabian StyleKontoghiorghes, George J. 2024. "The Importance and Essentiality of Natural and Synthetic Chelators in Medicine: Increased Prospects for the Effective Treatment of Iron Overload and Iron Deficiency" International Journal of Molecular Sciences 25, no. 9: 4654. https://doi.org/10.3390/ijms25094654
APA StyleKontoghiorghes, G. J. (2024). The Importance and Essentiality of Natural and Synthetic Chelators in Medicine: Increased Prospects for the Effective Treatment of Iron Overload and Iron Deficiency. International Journal of Molecular Sciences, 25(9), 4654. https://doi.org/10.3390/ijms25094654