
CX3CL1 (Fractalkine)-CX3CR1 Axis in Inflammation-Induced Angiogenesis and Tumorigenesis


Abstract
:1. Introduction
1.1. Angiogenesis
1.2. Chemokines
2. Chemokine CX3CL1 (Fractalkine, FKN)
2.1. CX3CL1 Structure
2.2. CX3CL1 Function
2.2.1. Membrane-Bound CX3CL1 (mFKN) as an Adhesion Molecule
2.2.2. Soluble CX3CL1 (sFKN) as a Chemoattractant
3. CX3CR1—The Sole Fractalkine Receptor
3.1. CX3CR1 Structure
3.2. FKN Signaling via CX3CR1
3.2.1. Conformational Rearrangements following FKN Binding
3.2.2. Main FKN/CX3CR1 Signaling Pathways
4. FKN/CX3CR1 Axis and Inflammation
4.1. FKN/CX3CR1 Axis and Inflammation-Induced Angiogenesis
4.1.1. Interdependence of Inflammation and Angiogenesis
4.1.2. Pro-Angiogenic Effects of FKN/CX3CR1 Signaling on the Inflammatory Response
5. FKN/CX3CR1 Axis and Tumorigenesis
5.1. Hypoxia and Angiogenesis in the Tumor Microenvironment (TME)
5.2. FKN in the TME
5.2.1. FKN/CX3CR1 Signaling May Promote Tumorigenesis
5.2.2. FKN/CX3CR1 Signaling May Be a Good Prognostic Factor in Cancer
5.2.3. Possible Reasons for the Contradictory Results of FKN/CX3CR1 Signaling in Cancer
6. Concluding Remarks
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Å | Ångström, unit of length, equal to 10−10 m, or 0.1 nm |
| AC | adenylyl cyclase |
| ACKRs | atypical chemokine receptors |
| ADAM10 | a disintegrin and metalloproteinase domain-containing protein 10 |
| ADAM17 | a disintegrin and metalloproteinase domain-containing protein 17, also known as tumor necrosis factor alpha (TNF-α) converting enzyme (TACE) |
| ADCC | antibody-dependent cellular cytotoxicity |
| Akt | protein kinase B |
| ALS | amyotrophic lateral sclerosis |
| AMI | acute myocardial infarction |
| AP | angina pectoris |
| AP2 | adapter protein 2 |
| APC | antigen presenting cells |
| BCL2 | B-cell lymphoma 2 gene |
| BCL-xL | B-cell lymphoma extra-large gene |
| BM | bone marrow |
| BMSCs | bone marrow-derived mesenchymal stem cells |
| cAMP | cyclic adenosine monophosphate |
| cCKRs | conventional chemokine receptors |
| CCL26 | chemokine eotaxin-3 |
| CCR2 | C-C chemokine receptor 2 |
| CCR5 | C-C chemokine receptor type 5 |
| CD | chemokine domain |
| CD106 | cluster of differentiation 106, an adhesion molecule |
| CD40 | cluster of differentiation 40 also known as—tumor necrosis factor receptor superfamily member 5 (TNFRSF5) |
| CD40L | cluster of differentiation 40 (CD40) ligand |
| CNS | central nervous system |
| CREB | cyclic adenosine monophosphate(cAMP)/Ca2+ response element binding protein |
| Cryo-EM | cryo-electron microscopy |
| CTCs | circulating tumor cells |
| CTS | cathepsin S |
| CX3CL1 | chemokine (C-X3-C motif) ligand 1, also known as fractalkine (FKN) |
| CX3CL1.35 | chemokine, US28-engineered fractalkine |
| CX3CR1 | high-affinity fractalkine (FKN) receptor or chemokine (C-X3-C motif) ligand 1 (CX3CL1) receptor, also known as G protein-coupled receptor 13 (GPR13), previously known as V28 |
| DC | dendritic cell |
| DVT | deep vein thrombosis |
| ECM | extracellular matrix |
| EC | endothelial cell |
| ECL1–ECL3 | three extracellular loops within G protein-coupled receptor (GPCR) |
| EGFR | epidermal growth factor receptor, a member of the family closely related to receptor tyrosine kinases ErbB-1 (EGFR) and ErbB-2 (HER2/neu) |
| EMT | epithelial-to-mesenchymal transition |
| eNOS | endothelial nitric oxide synthase |
| EPCs | endothelial progenitor cells |
| ERK | extracellular signal-regulated kinase |
| FAK | focal adhesion kinase |
| FGR | fetal growth restriction |
| FKN | fractalkine also known as chemokine (C-X3-C motif) ligand 1 (CX3CL1) |
| FLSs | fibroblast-like synoviocytes |
| FOXO | member of the class O of forkhead box transcription factors |
| Gα, Gβ, Gγ | subunits of the heterotrimeric G proteins (G protein complex) |
| Gαi | activated Gα subunit of the G protein complex |
| GDP | guanosine diphosphate |
| GPCR | G protein-coupled receptor |
| GPR domain | G protein regulatory domain-containing specific protein |
| GTP | guanosine triphosphate |
| HCC | hepatocellular carcinoma |
| HIF | hypoxia inducible factor |
| HIF-1α | hypoxia inducible factor 1 alpha |
| HIMECs | human intestinal microvascular endothelial cell |
| IBD | inflammatory bowel disease |
| ICL1–ICL3 | three intracellular loops within G protein-coupled receptor (GPCR) |
| IDD | intervertebral disc degeneration |
| IKK | IkappaBeta (Iκβ) kinase |
| IL-1, IL-1β, IL-6, IL-33 | interleukin-1, -1β, -6 and -33, respectively |
| IP3 | inositol 1,4,5-trisphosphate |
| ITGA5 | integrin alpha 5 also known as anti-CD49e antigen |
| JAK | Janus kinase |
| JNK | cJun NH(2)-terminal kinase |
| (KLRG1)+ NK cells | (killer cell lectin-like receptor subfamily G member 1)+ natural killer cells, a subset considered terminally differentiated |
| LPS | lipopolysaccharide |
| MACEs | major adverse cardiovascular events |
| MAPK | mitogen-activated protein kinase |
| MC | mast cells |
| MCP-1 | monocyte chemoattractant protein-1 |
| MEK | mitogen-activated protein kinase kinase |
| MEKK | mitogen-activated protein kinase kinase (MEK) kinase |
| mFKN | membrane-bound form of fractalkine |
| MLN | mesenteric lymph node |
| MM | multiple myeloma |
| MMP-2 | matrix metalloprotease-2 |
| MVO | microvascular obstruction |
| NAFLD | nonalcoholic fatty liver disease |
| NASH | nonalcoholic steatohepatitis |
| NCBI | National Centre for Biotechnology Information |
| NF-κβ | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NK cell | natural killer cell |
| NO | nitric oxide |
| NRP-1, NRP-2 | neuropilin 1, neuropilin 2 |
| NTx | N-terminal telopeptide of type I collagen |
| OA | osteoarthritis |
| OCPs | osteoclast precursors |
| P38 | mitogen-activated protein kinases |
| PAH | pulmonary arterial hypertension |
| PBHSC | peripheral blood-derived hematopoietic stem cells |
| PD | Parkinson’s disease |
| PDK1 | phosphoinositide-dependent kinase 1 |
| PE | pre-eclampsia |
| PECAMN-1 | platelet-endothelial adhesion molecule-1 also known as CD31 |
| pGlu | pyroglutamate |
| PI3K | phosphoinositide 3-kinase |
| PIP2 | phosphatidylinositol 4,5-biphosphate |
| PKC | protein kinase C |
| PLC | phospholipase C |
| QC | glutaminyl cyclase |
| RA | rheumatoid arthritis |
| Raf | Raf kinases |
| Ras | Ras kinases |
| RIC8 | non-receptor guanine nucleotide exchange factor for Gα subunits (also known as synembryn) |
| RMSD | root mean square deviation |
| RTKs | receptor tyrosine kinases |
| sFKN | soluble form of fractalkine |
| SNARE | vesicle-associated v-soluble N-ethylmaleimide-sensitive factor attachment protein receptor |
| SP | signal peptide |
| Src | steroid receptor coactivator |
| STAT | signal transducer and activator of transcription protein |
| STEMI | ST(segment)-elevation myocardial infarction |
| STX13 | protein syntaxin 13 |
| TACE | tumor necrosis factor alpha (TNF-α) converting enzyme, also known as a disintegrin and metalloproteinase domain-containing protein 17 |
| TGF-α | transforming growth factor alpha |
| TIE-2 | tyrosine kinase with immunoglobulin-like and EGF-like domains 2 |
| TIL | tumor-infiltrating lymphocytes |
| TIMER | Tumor IMmune Estimation Resource database |
| TM1–TM7 | seven hydrophobic α-helical transmembrane segments or domains within a G protein-coupled receptor (GPCR) |
| TME | tumor microenvironment |
| TNFα | tumor necrosis factor alpha |
| TNFRSF5 | tumor necrosis factor receptor superfamily member 5 also known as cluster of differentiation 40 (CD40) |
| TRACP-5b | tartrate-resistant acid phosphatase 5b |
| US28-engineered FKN | chemokine CX3CL1.35 |
| VAMP3 | vesicle-associated membrane protein 3 |
| VCAM-1 | vascular cell adhesion molecule 1 also known as vascular cell adhesion protein 1 |
| VE-cadherin | Vascular endothelial cadherin, also known as CD144 |
| VEGF | vascular endothelial growth factor |
| VEGFR | vascular endothelial growth factor (VEGF) receptor |
| VEGFR-1, VEGFR-2 | vascular endothelial growth factor (VEGF) receptor type 1 and 2, respectively |
| VEGF-A | vascular endothelial growth factor (VEGF) isoform A |
| V/EVTI | vascular/extravascular tissue index |
| VPF | vascular permeability factor |
| VSMCs | vascular smooth muscle cells |
References
- Adair, T.H.; Montani, J.P. Angiogenesis; Morgan & Claypool Life Sciences: San Rafael, CA, USA, 2010; Chapter 1; Overview of Angiogenesis. Available online: https://www.ncbi.nlm.nih.gov/books/NBK53238 (accessed on 15 March 2024).
- Szewczyk, G.; Maciejewski, T.M.; Szukiewicz, D. Current progress in the inflammatory background of angiogenesis in gynecological cancers. Inflamm. Res. 2019, 68, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Yoder, M.C. Human endothelial progenitor cells. Cold Spring Harb. Perspect. Med. 2012, 2, a006692. [Google Scholar] [CrossRef] [PubMed]
- Marçola, M.; Rodrigues, C.E. Endothelial progenitor cells in tumor angiogenesis: Another brick in the wall. Stem. Cells Int. 2015, 2015, 832649. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.Y.; Cheung, C. Origins and functional differences of blood endothelial cells. Semin. Cell Dev. Biol. 2024, 155 Pt C, 23–29. [Google Scholar] [CrossRef]
- Fujisawa, T.; Tura-Ceide, O.; Hunter, A.; Mitchell, A.; Vesey, A.; Medine, C.; Gallogly, S.; Hadoke, P.W.F.; Keith, C.; Sproul, A.; et al. Endothelial Progenitor Cells Do Not Originate from the Bone Marrow. Circulation 2019, 140, 1524–1526. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Solomonidis, E.G.; Meloni, M.; Taylor, R.S.; Duffin, R.; Dobie, R.; Magalhaes, M.S.; Henderson, B.E.P.; Louwe, P.A.; D’Amico, G.; et al. Single-cell transcriptome analyses reveal novel targets modulating cardiac neovascularization by resident endothelial cells following myocardial infarction. Eur. Heart J. 2019, 40, 2507–2520. [Google Scholar] [CrossRef]
- Ackermann, M.; Houdek, J.P.; Gibney, B.C.; Ysasi, A.; Wagner, W.; Belle, J.; Schittny, J.C.; Enzmann, F.; Tsuda, A.; Mentzer, S.J.; et al. Sprouting and intussusceptive angiogenesis in postpneumonectomy lung growth: Mechanisms of alveolar neovascularization. Angiogenesis 2014, 17, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Hickey, M.M.; Simon, M.C. Regulation of angiogenesis by hypoxia and hypoxia-inducible factors. Curr. Top. Dev. Biol. 2006, 76, 217–257. [Google Scholar] [CrossRef]
- Lin, S.; Chai, Y.; Zheng, X.; Xu, X. The role of HIF in angiogenesis, lymphangiogenesis, and tumor microenvironment in urological cancers. Mol. Biol. Rep. 2023, 51, 14. [Google Scholar] [CrossRef]
- Yuan, X.; Ruan, W.; Bobrow, B.; Carmeliet, P.; Eltzschig, H.K. Targeting hypoxia-inducible factors: Therapeutic opportunities and challenges. Nat. Rev. Drug Discov. 2023, 23, 175–200. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, Y.; Yang, Y.; Dang, Y.; Guo, Z.; Zhuang, Q.; Zheng, X.; Wang, F.; Cheng, N.; Liu, X.; et al. Hypoxia induces hepatocellular carcinoma metastasis via the HIF-1α/METTL16/lnc-CSMD1-7/RBFOX2 axis. iScience 2023, 26, 108495. [Google Scholar] [CrossRef] [PubMed]
- Laderoute, K.R.; Amin, K.; Calaoagan, J.M.; Knapp, M.; Le, T.; Orduna, J.; Foretz, M.; Viollet, B. 5′-AMP-activated protein kinase (AMPK) is induced by low-oxygen and glucose deprivation conditions found in solid-tumor microenvironments. Mol. Cell Biol. 2006, 26, 5336–5347. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Luo, S.; Fan, D.; Guo, X.; Ma, S. The role of vascular endothelial cells in tumor metastasis. Acta Histochem. 2023, 125, 152070. [Google Scholar] [CrossRef] [PubMed]
- Ebeling, S.; Kowalczyk, A.; Perez-Vazquez, D.; Mattiola, I. Regulation of tumor angiogenesis by the crosstalk between innate immunity and endothelial cells. Front. Oncol. 2023, 13, 1171794. [Google Scholar] [CrossRef] [PubMed]
- Bisht, M.; Dhasmana, D.C.; Bist, S.S. Angiogenesis: Future of pharmacological modulation. Indian J. Pharmacol. 2010, 42, 2–8. [Google Scholar] [CrossRef]
- Wong, B.W.; Marsch, E.; Treps, L.; Baes, M.; Carmeliet, P. Endothelial cell metabolism in health and disease: Impact of hypoxia. EMBO J. 2017, 36, 2187–2203. [Google Scholar] [CrossRef] [PubMed]
- Senger, D.R.; Davis, G.E. Angiogenesis. Cold Spring Harb. Perspect. Biol. 2011, 3, a005090. [Google Scholar] [CrossRef] [PubMed]
- Nan, W.; He, Y.; Wang, S.; Zhang, Y. Molecular mechanism of VE-cadherin in regulating endothelial cell behaviour during angiogenesis. Front. Physiol. 2023, 14, 1234104. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.H.; Choi, O.; Taylor-Weiner, H.; Fuhrmann, A.; Karpiak, J.V.; Almutairi, A.; Engler, A.J. Haptotaxis is cell type specific and limited by substrate adhesiveness. Cell Mol. Bioeng. 2015, 8, 530–542. [Google Scholar] [CrossRef]
- Kazerounian, S.; Lawler, J. Integration of pro- and anti-angiogenic signals by endothelial cells. J. Cell Commun. Signal. 2018, 12, 171–179. [Google Scholar] [CrossRef]
- Geindreau, M.; Bruchard, M.; Vegran, F. Role of Cytokines and Chemokines in Angiogenesis in a Tumor Context. Cancers 2022, 14, 2446. [Google Scholar] [CrossRef] [PubMed]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor angiogenesis: Causes, consequences, challenges and opportunities. Cell Mol. Life Sci. 2020, 77, 1745–1770. [Google Scholar] [CrossRef] [PubMed]
- Balogh, E.; Biniecka, M.; Fearon, U.; Veale, D.J.; Szekanecz, Z. Angiogenesis in Inflammatory Arthritis. Isr. Med. Assoc. J. 2019, 21, 345–352. [Google Scholar] [PubMed]
- Lu, E.; Li, C.; Wang, J.; Zhang, C. Inflammation and angiogenesis in the corpus luteum. J. Obstet. Gynaecol. Res. 2019, 45, 1967–1974. [Google Scholar] [CrossRef] [PubMed]
- Corliss, B.A.; Azimi, M.S.; Munson, J.M.; Peirce, S.M.; Murfee, W.L. Macrophages: An Inflammatory Link Between Angiogenesis and Lymphangiogenesis. Microcirculation 2016, 23, 95–121. [Google Scholar] [CrossRef]
- Walsh, D.A.; Pearson, C.I. Angiogenesis in the pathogenesis of inflammatory joint and lung diseases. Arthritis Res. 2001, 3, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Tsoupras, A.; Lordan, R.; Zabetakis, I. Inflammation, not Cholesterol, Is a Cause of Chronic Disease. Nutrients 2018, 10, 604. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, U.; Augustin, H.G. Angiopoietins: A link between angiogenesis and inflammation. Trends Immunol. 2006, 27, 552–558. [Google Scholar] [CrossRef]
- Jeong, J.H.; Ojha, U.; Lee, Y.M. Pathological angiogenesis and inflammation in tissues. Arch. Pharm. Res. 2021, 44, 1–15. [Google Scholar] [CrossRef]
- Nagy, J.A.; Benjamin, L.; Zeng, H.; Dvorak, A.M.; Dvorak, H.F. Vascular permeability, vascular hyperpermeability and angiogenesis. Angiogenesis 2008, 11, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Mehrad, B.; Keane, M.P.; Strieter, R.M. Chemokines as mediators of angiogenesis. Thromb. Haemost. 2007, 97, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.C.; Mayo, K.H. Chemokines from a Structural Perspective. Int. J. Mol. Sci. 2017, 18, 2088. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.E.; Nibbs, R.J.B. A guide to chemokines and their receptors. FEBS J. 2018, 285, 2944–2971. [Google Scholar] [CrossRef] [PubMed]
- Kufareva, I.; Salanga, C.L.; Handel, T.M. Chemokine and chemokine receptor structure and interactions: Implications for therapeutic strategies. Immunol. Cell Biol. 2015, 93, 372–383. [Google Scholar] [CrossRef] [PubMed]
- Legler, D.F.; Thelen, M. New insights in chemokine signaling. F1000Research 2018, 7, 95. [Google Scholar] [CrossRef]
- Stone, M.J.; Hayward, J.A.; Huang, C.; EHuma, Z.; Sanchez, J. Mechanisms of Regulation of the Chemokine-Receptor Network. Int. J. Mol. Sci. 2017, 18, 342. [Google Scholar] [CrossRef]
- Portella, L.; Bello, A.M.; Scala, S. CXCL12 Signaling in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2021, 1302, 51–70. [Google Scholar] [CrossRef]
- Nazari, A.; Khorramdelazad, H.; Hassanshahi, G. Biological/pathological functions of the CXCL12/CXCR4/CXCR7 axes in the pathogenesis of bladder cancer. Int. J. Clin. Oncol. 2017, 22, 991–1000. [Google Scholar] [CrossRef]
- Shakir, M.; Tang, D.; Zeh, H.J.; Tang, S.W.; Anderson, C.J.; Bahary, N.; Lotze, M.T. The chemokine receptors CXCR4/CXCR7 and their primary heterodimeric ligands CXCL12 and CXCL12/high mobility group box 1 in pancreatic cancer growth and development: Finding flow. Pancreas 2015, 44, 528–534. [Google Scholar] [CrossRef]
- Imai, T.; Hieshima, K.; Haskell, C.; Baba, M.; Nagira, M.; Nishimura, M.; Kakizaki, M.; Takagi, S.; Nomiyama, H.; Schall, T.J.; et al. Identification and molecular characterization of fractalkine receptor CX3CR1, which mediates both leukocyte migration and adhesion. Cell 1997, 91, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Wojdasiewicz, P.; Poniatowski, L.A.; Kotela, A.; Deszczyński, J.; Kotela, I.; Szukiewicz, D. The chemokine CX3CL1 (fractalkine) and its receptor CX3CR1: Occurrence and potential role in osteoarthritis. Arch. Immunol. Ther. Exp. 2014, 62, 395–403. [Google Scholar] [CrossRef]
- Loh, S.X.; Ekinci, Y.; Spray, L.; Jeyalan, V.; Olin, T.; Richardson, G.; Austin, D.; Alkhalil, M.; Spyridopoulos, I. Fractalkine Signalling (CX3CL1/CX3CR1 Axis) as an Emerging Target in Coronary Artery Disease. J. Clin. Med. 2023, 12, 4821. [Google Scholar] [CrossRef]
- Imaizumi, T.; Yoshida, H.; Satoh, K. Regulation of CX3CL1/fractalkine expression in endothelial cells. J. Atheroscler. Thromb. 2004, 11, 15–21. [Google Scholar] [CrossRef]
- Kiefer, F.; Siekmann, A.F. The role of chemokines and their receptors in angiogenesis. Cell Mol. Life Sci. 2011, 68, 2811–2830. [Google Scholar] [CrossRef]
- Strieter, R.M.; Burdick, M.D.; Gomperts, B.N.; Belperio, J.A.; Keane, M.P. CXC chemokines in angiogenesis. Cytokine Growth Factor Rev. 2005, 16, 593–609. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.A.; Hippe, A.; Buhren, B.A.; Müller, A.; Homey, B. Chemokines in tumor-associated angiogenesis. Biol. Chem. 2009, 390, 1213–1223. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Yang, W.; Sun, A.; Wei, Z.; Lin, Q. The Role of CXC Chemokines in Cancer Progression. Cancers 2022, 15, 167. [Google Scholar] [CrossRef]
- Bazan, J.F.; Bacon, K.B.; Hardiman, G.; Wang, W.; Soo, K.; Rossi, D.; Greaves, D.R.; Zlotnik, A.; Schall, T.J. A new class of membrane-bound chemokine with a CX3C motif. Nature 1997, 385, 640–644. [Google Scholar] [CrossRef]
- Pan, Y.; Lloyd, C.; Zhou, H.; Dolich, S.; Deeds, J.; Gonzalo, J.A.; Vath, J.; Gosselin, M.; Ma, J.; Dussault, B.; et al. Neurotactin, a membrane-anchored chemokine upregulated in brain inflammation. Nature 1997, 387, 611–617, Erratum in Nature 1997, 389, 100. [Google Scholar] [CrossRef]
- Hortsch, M.; Patel, N.H.; Bieber, A.J.; Traquina, Z.R.; Goodman, C.S. Drosophila neurotactin, a surface glycoprotein with homology to serine esterases, is dynamically expressed during embryogenesis. Development 1990, 110, 1327–1340. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Q.; Cheng, K.; Ming, Y. CX3CL1/CX3CR1 Axis, as the Therapeutic Potential in Renal Diseases: Friend or Foe? Curr. Gene Ther. 2017, 17, 442–452. [Google Scholar] [CrossRef]
- D’Haese, J.G.; Demir, I.E.; Friess, H.; Ceyhan, G.O. Fractalkine/CX3CR1: Why a single chemokine-receptor duo bears a major and unique therapeutic potential. Expert Opin. Ther. Targets 2010, 14, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.Y.; Peng, P.W.; Renn, T.Y.; Lee, C.J.; Chang, T.M.; Wei, A.I.; Liu, J.F. CX3CL1 induces cell migration and invasion through ICAM-1 expression in oral squamous cell carcinoma cells. J. Cell Mol. Med. 2023, 27, 1509–1522. [Google Scholar] [CrossRef] [PubMed]
- White, G.E.; Greaves, D.R. Fractalkine: A survivor’s guide: Chemokines as antiapoptotic mediators. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Lucas, A.D.; Chadwick, N.; Warren, B.F.; Jewell, D.P.; Gordon, S.; Powrie, F.; Greaves, D.R. The transmembrane form of the CX3CL1 chemokine fractalkine is expressed predominantly by epithelial cells in vivo. Am. J. Pathol. 2001, 158, 855–866. [Google Scholar] [CrossRef]
- Jones, B.A.; Beamer, M.; Ahmed, S. Fractalkine/CX3CL1: A potential new target for inflammatory diseases. Mol. Interv. 2010, 10, 263–270. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, S.A.; Gasparini, F.; Mir, A.K.; Dev, K.K. Fractalkine shedding is mediated by p38 and the ADAM10 protease under pro-inflammatory conditions in human astrocytes. J. Neuroinflammation 2016, 13, 189. [Google Scholar] [CrossRef] [PubMed]
- Hundhausen, C.; Misztela, D.; Berkhout, T.A.; Broadway, N.; Saftig, P.; Reiss, K.; Hartmann, D.; Fahrenholz, F.; Postina, R.; Matthews, V.; et al. The disintegrin-like metalloproteinase ADAM10 is involved in constitutive cleavage of CX3CL1 (fractalkine) and regulates CX3CL1-mediated cell-cell adhesion. Blood 2003, 102, 1186–1195. [Google Scholar] [CrossRef]
- Poniatowski, Ł.A.; Wojdasiewicz, P.; Krawczyk, M.; Szukiewicz, D.; Gasik, R.; Kubaszewski, Ł.; Kurkowska-Jastrzębska, I. Analysis of the Role of CX3CL1 (Fractalkine) and Its Receptor CX3CR1 in Traumatic Brain and Spinal Cord Injury: Insight into Recent Advances in Actions of Neurochemokine Agents. Mol. Neurobiol. 2017, 54, 2167–2188. [Google Scholar] [CrossRef]
- Jones, B.A.; Riegsecker, S.; Rahman, A.; Beamer, M.; Aboualaiwi, W.; Khuder, S.A.; Ahmed, S. Role of ADAM-17, p38 MAPK, cathepsins, and the proteasome pathway in the synthesis and shedding of fractalkine/CX₃ CL1 in rheumatoid arthritis. Arthritis Rheum. 2013, 65, 2814–2825. [Google Scholar] [CrossRef] [PubMed]
- Bourd-Boittin, K.; Basset, L.; Bonnier, D.; L’helgoualc’h, A.; Samson, M.; Théret, N. CX3CL1/fractalkine shedding by human hepatic stellate cells: Contribution to chronic inflammation in the liver. J. Cell Mol. Med. 2009, 13, 1526–1535. [Google Scholar] [CrossRef] [PubMed]
- Uchida, M.; Ito, T.; Nakamura, T.; Igarashi, H.; Oono, T.; Fujimori, N.; Kawabe, K.; Suzuki, K.; Jensen, R.T.; Takayanagi, R. ERK pathway and sheddases play an essential role in ethanol-induced CX3CL1 release in pancreatic stellate cells. Lab Investig. 2013, 93, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Lu, X. Structure and Function of Ligand CX3CL1 and its Receptor CX3CR1 in Cancer. Curr. Med. Chem. 2022, 29, 6228–6246. [Google Scholar] [CrossRef]
- Iemmolo, M.; Ghersi, G.; Bivona, G. The Cytokine CX3CL1 and ADAMs/MMPs in Concerted Cross-Talk Influencing Neurodegenerative Diseases. Int. J. Mol. Sci. 2023, 24, 8026. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.L.; Mangnall, D.; Bird, N.C.; Blair-Zajdel, M.E.; Bunning, R.A. Effects of pro-inflammatory cytokines on the production of soluble fractalkine and ADAM17 by HepG2 cells. J. Gastrointestin. Liver Dis. 2010, 19, 265–271. [Google Scholar]
- Fonović, U.P.; Jevnikar, Z.; Kos, J. Cathepsin S generates soluble CX3CL1 (fractalkine) in vascular smooth muscle cells. Biol. Chem. 2013, 394, 1349–1352. [Google Scholar] [CrossRef]
- Hundhausen, C.; Schulte, A.; Schulz, B.; Andrzejewski, M.G.; Schwarz, N.; von Hundelshausen, P.; Winter, U.; Paliga, K.; Reiss, K.; Saftig, P.; et al. Regulated shedding of transmembrane chemokines by the disintegrin and metalloproteinase 10 facilitates detachment of adherent leukocytes. J. Immunol. 2007, 178, 8064–8072. [Google Scholar] [CrossRef]
- Scarselli, M.; Donaldson, J.G. Constitutive internalization of G protein-coupled receptors and G proteins via clathrin-independent endocytosis. J. Biol. Chem. 2009, 284, 3577–3585. [Google Scholar] [CrossRef]
- Hattermann, K.; Gebhardt, H.; Krossa, S.; Ludwig, A.; Lucius, R.; Held-Feindt, J.; Mentlein, R. Transmembrane chemokines act as receptors in a novel mechanism termed inverse signaling. eLife 2016, 5, e10820. [Google Scholar] [CrossRef]
- Shimaoka, T.; Nakayama, T.; Fukumoto, N.; Kume, N.; Takahashi, S.; Yamaguchi, J.; Minami, M.; Hayashida, K.; Kita, T.; Ohsumi, J.; et al. Cell surface-anchored SR-PSOX/CXC chemokine ligand 16 mediates firm adhesion of CXC chemokine receptor 6-expressing cells. J. Leukoc. Biol. 2004, 75, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Ostuni, M.A.; Hermand, P.; Saindoy, E.; Guillou, N.; Guellec, J.; Coens, A.; Hattab, C.; Desuzinges-Mandon, E.; Jawhari, A.; Iatmanen-Harbi, S.; et al. CX3CL1 homo-oligomerization drives cell-to-cell adherence. Sci. Rep. 2020, 10, 9069. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Lee, Y.; Song, J.; Lee, J.; Chang, S.Y. Tissue-specific Role of CX3CR1 Expressing Immune Cells and Their Relationships with Human Disease. Immune. Netw. 2018, 18, e5. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Zhuge, F.; Ni, L.; Nagata, N.; Yamashita, T.; Mukaida, N.; Kaneko, S.; Ota, T.; Nagashimada, M. CX3CL1/CX3CR1 interaction protects against lipotoxicity-induced nonalcoholic steatohepatitis by regulating macrophage migration and M1/M2 status. Metabolism 2022, 136, 155272. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Dong, L.; Li, J.; Chen, Y.; Yao, Y.; Zeng, R.; Shushakova, N.; Haller, H.; Xu, G.; Rong, S. C-X3-C motif chemokine ligand 1/receptor 1 regulates the M1 polarization and chemotaxis of macrophages after hypoxia/reoxygenation injury. Chronic. Dis. Transl. Med. 2021, 7, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Cormican, S.; Griffin, M.D. Fractalkine (CX3CL1) and Its Receptor CX3CR1: A Promising Therapeutic Target in Chronic Kidney Disease? Front. Immunol. 2021, 12, 664202. [Google Scholar] [CrossRef] [PubMed]
- Kerfoot, S.M.; Lord, S.E.; Bell, R.B.; Gill, V.; Robbins, S.M.; Kubes, P. Human fractalkine mediates leukocyte adhesion but not capture under physiological shear conditions; a mechanism for selective monocyte recruitment. Eur. J. Immunol. 2003, 33, 729–739. [Google Scholar] [CrossRef]
- Goda, S.; Imai, T.; Yoshie, O.; Yoneda, O.; Inoue, H.; Nagano, Y.; Okazaki, T.; Imai, H.; Bloom, E.T.; Domae, N.; et al. CX3C-chemokine, fractalkine-enhanced adhesion of THP-1 cells to endothelial cells through integrin-dependent and -independent mechanisms. J. Immunol. 2000, 164, 4313–4320. [Google Scholar] [CrossRef] [PubMed]
- Flierl, U.; Bauersachs, J.; Schäfer, A. Modulation of platelet and monocyte function by the chemokine fractalkine (CX3 CL1) in cardiovascular disease. Eur. J. Clin. Investig. 2015, 45, 624–633. [Google Scholar] [CrossRef]
- Umehara, H.; Imai, T. Role of fractalkine in leukocyte adhesion and migration and in vascular injury. Drug News Perspect. 2001, 14, 460–464. [Google Scholar] [CrossRef]
- Hermand, P.; Pincet, F.; Carvalho, S.; Ansanay, H.; Trinquet, E.; Daoudi, M.; Combadière, C.; Deterre, P. Functional adhesiveness of the CX3CL1 chemokine requires its aggregation. Role of the transmembrane domain. J. Biol. Chem. 2008, 283, 30225–30234. [Google Scholar] [CrossRef] [PubMed]
- Ostuni, M.A.; Guellec, J.; Hermand, P.; Durand, P.; Combadière, C.; Pincet, F.; Deterre, P. CX3CL1, a chemokine finely tuned to adhesion: Critical roles of the stalk glycosylation and the membrane domain. Biol. Open. 2014, 3, 1173–1182. [Google Scholar] [CrossRef]
- Huang, Y.W.; Su, P.; Liu, G.Y.; Crow, M.R.; Chaukos, D.; Yan, H.; Robinson, L.A. Constitutive endocytosis of the chemokine CX3CL1 prevents its degradation by cell surface metalloproteases. J. Biol. Chem. 2009, 284, 29644–29653. [Google Scholar] [CrossRef]
- Wong, H.S.; Jaumouillé, V.; Heit, B.; Doodnauth, S.A.; Patel, S.; Huang, Y.W.; Grinstein, S.; Robinson, L.A. Cytoskeletal confinement of CX3CL1 limits its susceptibility to proteolytic cleavage by ADAM10. Mol. Biol. Cell 2014, 25, 3884–3899. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.Y.; Kulasingam, V.; Alexander, R.T.; Touret, N.; Fong, A.M.; Patel, D.D.; Robinson, L.A. Recycling of the membrane-anchored chemokine, CX3CL1. J. Biol. Chem. 2005, 280, 19858–19866. [Google Scholar] [CrossRef]
- Miller, A.F.; Falke, J.J. Chemotaxis receptors and signaling. Adv. Protein Chem. 2004, 68, 393–444. [Google Scholar] [CrossRef] [PubMed]
- Conroy, M.J.; Lysaght, J. CX3CL1 Signaling in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1231, 1–12. [Google Scholar] [CrossRef]
- Zlotnik, A.; Yoshie, O. The chemokine superfamily revisited. Immunity 2012, 36, 705–716. [Google Scholar] [CrossRef]
- Richmond, A. Chemokine research moves on. Exp. Cell Res. 2011, 317, 553–555. [Google Scholar] [CrossRef]
- Mortier, A.; Van Damme, J.; Proost, P. Overview of the mechanisms regulating chemokine activity and availability. Immunol. Lett. 2012, 145, 2–9. [Google Scholar] [CrossRef]
- Johnston, B.; Butcher, E.C. Chemokines in rapid leukocyte adhesion triggering and migration. Semin. Immunol. 2002, 14, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Nanki, T.; Imai, T.; Nagasaka, K.; Urasaki, Y.; Nonomura, Y.; Taniguchi, K.; Hayashida, K.; Hasegawa, J.; Yoshie, O.; Miyasaka, N. Migration of CX3CR1-positive T cells producing type 1 cytokines and cytotoxic molecules into the synovium of patients with rheumatoid arthritis. Arthritis Rheum. 2002, 46, 2878–2883. [Google Scholar] [CrossRef] [PubMed]
- Hamann, I.; Unterwalder, N.; Cardona, A.E.; Meisel, C.; Zipp, F.; Ransohoff, R.M.; Infante-Duarte, C. Analyses of phenotypic and functional characteristics of CX3CR1-expressing natural killer cells. Immunology 2011, 133, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Mionnet, C.; Buatois, V.; Kanda, A.; Milcent, V.; Fleury, S.; Lair, D.; Langelot, M.; Lacoeuille, Y.; Hessel, E.; Coffman, R.; et al. CX3CR1 is required for airway inflammation by promoting T helper cell survival and maintenance in inflamed lung. Nat. Med. 2010, 16, 1305–1312. [Google Scholar] [CrossRef] [PubMed]
- Ancuta, P.; Rao, R.; Moses, A.; Mehle, A.; Shaw, S.K.; Luscinskas, F.W.; Gabuzda, D. Fractalkine preferentially mediates arrest and migration of CD16+ monocytes. J. Exp. Med. 2003, 197, 1701–1707. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, E.J.; Fitzhugh, D.J.; Tkaczyk, C.; Gilfillan, A.M.; Sassetti, C.; Metcalfe, D.D.; Hwang, S.T. Mast cells migrate, but do not degranulate, in response to fractalkine, a membrane-bound chemokine expressed constitutively in diverse cells of the skin. Eur. J. Immunol. 2000, 30, 2355–2361. [Google Scholar] [CrossRef] [PubMed]
- Beck, G.C.; Ludwig, F.; Schulte, J.; van Ackern, K.; van der Woude, F.J.; Yard, B.A. Fractalkine is not a major chemoattractant for the migration of neutrophils across microvascular endothelium. Scand J. Immunol. 2003, 58, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.D.; Kurtz, S.L.; Rigel, N.W.; Gunn, B.M.; Taft-Benz, S.; Morrison, J.P.; Fong, A.M.; Patel, D.D.; Braunstein, M.; Kawula, T.H. The impact of chemokine receptor CX3CR1 deficiency during respiratory infections with Mycobacterium tuberculosis or Francisella tularensis. Clin. Exp. Immunol. 2009, 156, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Volin, M.V.; Huynh, N.; Klosowska, K.; Reyes, R.D.; Woods, J.M. Fractalkine-induced endothelial cell migration requires MAP kinase signaling. Pathobiology 2010, 77, 7–16. [Google Scholar] [CrossRef]
- Liu, J.F.; Tsao, Y.T.; Hou, C.H. Fractalkine/CX3CL1 induced intercellular adhesion molecule-1-dependent tumor metastasis through the CX3CR1/PI3K/Akt/NF-κB pathway in human osteosarcoma. Oncotarget 2016, 8, 54136–54148. [Google Scholar] [CrossRef]
- Tang, J.; Xiao, L.; Cui, R.; Li, D.; Zheng, X.; Zhu, L.; Sun, H.; Pan, Y.; Du, Y.; Yu, X. CX3CL1 increases invasiveness and metastasis by promoting epithelial-to-mesenchymal transition through the TACE/TGF-α/EGFR pathway in hypoxic androgen-independent prostate cancer cells. Oncol. Rep. 2016, 35, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Garin, A.; Pellet, P.; Deterre, P.; Debré, P.; Combadière, C. Cloning and functional characterization of the human fractalkine receptor promoter regions. Biochem. J. 2002, 368 Pt 3, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Raport, C.J.; Schweickart, V.L.; Eddy, R.L., Jr.; Shows, T.B.; Gray, P.W. The orphan G-protein-coupled receptor-encoding gene V28 is closely related to genes for chemokine receptors and is expressed in lymphoid and neural tissues. Gene 1995, 163, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Combadiere, C.; Gao, J.; Tiffany, H.L.; Murphy, P.M. Gene cloning, RNA distribution, and functional expression of mCX3CR1, a mouse chemotactic receptor for the CX3C chemokine fractalkine. Biochem. Biophys. Res. Commun. 1998, 253, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Q.; Ou, J.; Zhang, S.; Ming, Y. Crosstalk between the CX3CL1/CX3CR1 Axis and Inflammatory Signaling Pathways in Tissue Injury. Curr. Protein Pept. Sci. 2019, 20, 844–854. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, N.; Pruessmeyer, J.; Hess, F.M.; Dreymueller, D.; Pantaler, E.; Koelsch, A.; Windoffer, R.; Voss, M.; Sarabi, A.; Weber, C.; et al. Requirements for leukocyte transmigration via the transmembrane chemokine CX3CL1. Cell Mol. Life Sci. 2010, 67, 4233–4248. [Google Scholar] [CrossRef]
- Raucci, R.; Costantini, S.; Castello, G.; Colonna, G. An overview of the sequence features of N- and C-terminal segments of the human chemokine receptors. Cytokine 2014, 70, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Szpakowska, M.; Perez Bercoff, D.; Chevigné, A. Closing the ring: A fourth extracellular loop in chemokine receptors. Sci. Signal. 2014, 7, pe21. [Google Scholar] [CrossRef]
- Nomiyama, H.; Yoshie, O. Functional roles of evolutionary conserved motifs and residues in vertebrate chemokine receptors. J. Leukoc. Biol. 2015, 97, 39–47. [Google Scholar] [CrossRef]
- Mafi, A.; Kim, S.K.; Goddard, W.A., 3rd. The mechanism for ligand activation of the GPCR-G protein complex. Proc. Natl. Acad. Sci. USA 2022, 119, e2110085119. [Google Scholar] [CrossRef]
- Tardáguila, M.; Mañes, S. The complex role of chemokines in cancer: The case of the CX3CL1/CX3CR1 axis. In Oncology Theory & Practice, 1st ed.; iConcept Press Ltd.: Madrid, Spain, 2014; Chapter 8. [Google Scholar]
- Chaudhri, A.; Bu, X.; Wang, Y.; Gomez, M.; Torchia, J.A.; Hua, P.; Hung, S.H.; Davies, M.A.; Lizee, G.A.; von Andrian, U.; et al. The CX3CL1-CX3CR1 chemokine axis can contribute to tumor immune evasion and blockade with a novel CX3CR1 monoclonal antibody enhances response to anti-PD-1 immunotherapy. Front. Immunol. 2023, 14, 1237715. [Google Scholar] [CrossRef] [PubMed]
- Goode-Romero, G.; Dominguez, L. Computational study of the conformational ensemble of CX3C chemokine receptor 1 (CX3CR1) and its interactions with antagonist and agonist ligands. J. Mol. Graph. Model. 2022, 117, 108278. [Google Scholar] [CrossRef] [PubMed]
- Mizoue, L.S.; Bazan, J.F.; Johnson, E.C.; Handel, T.M. Solution structure and dynamics of the CX3C chemokine domain of fractalkine and its interaction with an N-terminal fragment of CX3CR1. Biochemistry 1999, 38, 1402–1414. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, T.; Watanabe, Y.; Oiso, N.; Higuchi, T.; Shigeta, A.; Mizuguchi, N.; Katou, F.; Hashimoto, K.; Kawada, A.; Yoshie, O. Eotaxin-3/CC chemokine ligand 26 is a functional ligand for CX3CR1. J. Immunol. 2010, 185, 6472–6479. [Google Scholar] [CrossRef] [PubMed]
- Faure, S.; Meyer, L.; Costagliola, D.; Vaneensberghe, C.; Genin, E.; Autran, B.; Delfraissy, J.F.; McDermott, D.H.; Murphy, P.M.; Debré, P.; et al. Rapid progression to AIDS in HIV+ individuals with a structural variant of the chemokine receptor CX3CR1. Science 2000, 287, 2274–2277. [Google Scholar] [CrossRef] [PubMed]
- Niess, J.H.; Brand, S.; Gu, X.; Landsman, L.; Jung, S.; McCormick, B.A.; Vyas, J.M.; Boes, M.; Ploegh, H.L.; Fox, J.G.; et al. CX3CR1-mediated dendritic cell access to the intestinal lumen and bacterial clearance. Science 2005, 307, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Ludeman, J.P.; Stone, M.J. The structural role of receptor tyrosine sulfation in chemokine recognition. Br. J. Pharmacol. 2014, 171, 1167–1179. [Google Scholar] [CrossRef] [PubMed]
- Villaseca, S.; Romero, G.; Ruiz, M.J.; Pérez, C.; Leal, J.I.; Tovar, L.M.; Torrejón, M. Gαi protein subunit: A step toward understanding its non-canonical mechanisms. Front. Cell Dev. Biol. 2022, 10, 941870. [Google Scholar] [CrossRef]
- Lu, M.; Zhao, W.; Han, S.; Lin, X.; Xu, T.; Tan, Q.; Wang, M.; Yi, C.; Chu, X.; Yang, W.; et al. Activation of the human chemokine receptor CX3CR1 regulated by cholesterol. Sci. Adv. 2022, 8, eabn8048. [Google Scholar] [CrossRef]
- Laganà, M.; Schlecht-Louf, G.; Bachelerie, F. The G Protein-Coupled Receptor Kinases (GRKs) in Chemokine Receptor-Mediated Immune Cell Migration: From Molecular Cues to Physiopathology. Cells 2021, 10, 75. [Google Scholar] [CrossRef]
- Niessner, A.; Marculescu, R.; Haschemi, A.; Endler, G.; Zorn, G.; Weyand, C.M.; Maurer, G.; Mannhalter, C.; Wojta, J.; Wagner, O.; et al. Opposite effects of CX3CR1 receptor polymorphisms V249I and T280M on the development of acute coronary syndrome. A possible implication of fractalkine in inflammatory activation. Thromb. Haemost. 2005, 93, 949–954. [Google Scholar] [CrossRef]
- Wu, J.; Yin, R.X.; Lin, Q.Z.; Guo, T.; Shi, G.Y.; Sun, J.Q.; Shen, S.W.; Li, Q. Two polymorphisms in the Fractalkine receptor CX3CR1 gene influence the development of atherosclerosis: A meta-analysis. Dis. Markers 2014, 2014, 913678. [Google Scholar] [CrossRef]
- Chamera, K.; Szuster-Głuszczak, M.; Basta-Kaim, A. Shedding light on the role of CX3CR1 in the pathogenesis of schizophrenia. Pharmacol. Rep. 2021, 73, 1063–1078. [Google Scholar] [CrossRef]
- Sakai, M.; Takeuchi, H.; Yu, Z.; Kikuchi, Y.; Ono, C.; Takahashi, Y.; Ito, F.; Matsuoka, H.; Tanabe, O.; Yasuda, J.; et al. Polymorphisms in the microglial marker molecule CX3CR1 affect the blood volume of the human brain. Psychiatry Clin. Neurosci. 2018, 72, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Imai, T.; Yasuda, N. Therapeutic intervention of inflammatory/immune diseases by inhibition of the fractalkine (CX3CL1)-CX3CR1 pathway. Inflamm. Regen. 2016, 36, 9. [Google Scholar] [CrossRef]
- Mirzadegan, T.; Benkö, G.; Filipek, S.; Palczewski, K. Sequence analyses of G-protein-coupled receptors: Similarities to rhodopsin. Biochemistry 2003, 42, 2759–2767. [Google Scholar] [CrossRef] [PubMed]
- Joost, P.; Methner, A. Phylogenetic analysis of 277 human G-protein-coupled receptors as a tool for the prediction of orphan receptor ligands. Genome Biol. 2002, 3, research0063. [Google Scholar] [CrossRef]
- Zhou, X.E.; Melcher, K.; Xu, H.E. Structure and activation of rhodopsin. Acta Pharmacol. Sin. 2012, 33, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, R.J. Historical review: A brief history and personal retrospective of seven-transmembrane receptors. Trends Pharmacol. Sci. 2004, 25, 413–422. [Google Scholar] [CrossRef]
- Vischer, H.F.; Hulshof, J.W.; de Esch, I.J.; Smit, M.J.; Leurs, R. Virus-encoded G-protein-coupled receptors: Constitutively active (dys)regulators of cell function and their potential as drug target. In Ernst Schering Foundation Symposium Proceedings; Springer: Berlin/Heidelberg, Germany, 2006; pp. 187–209. [Google Scholar] [CrossRef]
- Burg, J.S.; Ingram, J.R.; Venkatakrishnan, A.J.; Jude, K.M.; Dukkipati, A.; Feinberg, E.N.; Angelini, A.; Waghray, D.; Dror, R.O.; Ploegh, H.L.; et al. Structural biology. Structural basis for chemokine recognition and activation of a viral G protein-coupled receptor. Science 2015, 347, 1113–1117. [Google Scholar] [CrossRef]
- Hjortø, G.M.; Kiilerich-Pedersen, K.; Selmeczi, D.; Kledal, T.N.; Larsen, N.B. Human cytomegalovirus chemokine receptor US28 induces migration of cells on a CX3CL1-presenting surface. J. Gen. Virol. 2013, 94 Pt 5, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Pierce, K.L.; Premont, R.T.; Lefkowitz, R.J. Seven-transmembrane receptors. Nat. Rev. Mol. Cell Biol. 2002, 3, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Palczewski, K. G protein-coupled receptor rhodopsin. Annu. Rev. Biochem. 2006, 75, 743–767. [Google Scholar] [CrossRef] [PubMed]
- Gustavsson, M.; Zheng, Y.; Handel, T.M. Production of Chemokine/Chemokine Receptor Complexes for Structural Biophysical Studies. Methods Enzymol. 2016, 570, 233–260. [Google Scholar] [CrossRef] [PubMed]
- Miles, T.F.; Spiess, K.; Jude, K.M.; Tsutsumi, N.; Burg, J.S.; Ingram, J.R.; Waghray, D.; Hjorto, G.M.; Larsen, O.; Ploegh, H.L.; et al. Viral GPCR US28 can signal in response to chemokine agonists of nearly unlimited structural degeneracy. eLife 2018, 7, e35850. [Google Scholar] [CrossRef]
- Tsutsumi, N.; Maeda, S.; Qu, Q.; Vögele, M.; Jude, K.M.; Suomivuori, C.M.; Panova, O.; Waghray, D.; Kato, H.E.; Velasco, A.; et al. Atypical structural snapshots of human cytomegalovirus GPCR interactions with host G proteins. Sci. Adv. 2022, 8, eabl5442. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Yang, D.; Wu, M.; Guo, Y.; Guo, W.; Zhong, L.; Cai, X.; Dai, A.; Jang, W.; Shakhnovich, E.I.; et al. Common activation mechanism of class A GPCRs. eLife 2019, 8, e50279. [Google Scholar] [CrossRef] [PubMed]
- Syrovatkina, V.; Alegre, K.O.; Dey, R.; Huang, X.Y. Regulation, Signaling, and Physiological Functions of G-Proteins. J. Mol. Biol. 2016, 428, 3850–3868. [Google Scholar] [CrossRef]
- Lagerström, M.C.; Schiöth, H.B. Structural diversity of G protein-coupled receptors and significance for drug discovery. Nat. Rev. Drug Discov. 2008, 7, 339–357, Erratum in Nat. Rev. Drug Discov. 2008, 7, 542. [Google Scholar] [CrossRef]
- Cancellieri, C.; Vacchini, A.; Locati, M.; Bonecchi, R.; Borroni, E.M. Atypical chemokine receptors: From silence to sound. Biochem. Soc. Trans. 2013, 41, 231–236. [Google Scholar] [CrossRef]
- Wingler, L.M.; Lefkowitz, R.J. Conformational Basis of G Protein-Coupled Receptor Signaling Versatility. Trends Cell Biol. 2020, 30, 736–747. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, C.; Zürn, A.; Bünemann, M.; Lohse, M.J. Conformational changes in G-protein-coupled receptors-the quest for functionally selective conformations is open. Br. J. Pharmacol. 2008, 153 (Suppl. S1), S358–S366. [Google Scholar] [CrossRef] [PubMed]
- Hamza, A.; Samad, A.; Parray, Z.A.; Ara, S.; Ahmed, A.; Almajhdi, F.N.; Hussain, T.; Islam, A.; Parveen, S. Mutation in the CX3C Motif of G Protein Disrupts Its Interaction with Heparan Sulfate: A Calorimetric, Spectroscopic, and Molecular Docking Study. Int. J. Mol. Sci. 2022, 23, 1950. [Google Scholar] [CrossRef] [PubMed]
- Darbandi-Tehrani, K.; Hermand, P.; Carvalho, S.; Dorgham, K.; Couvineau, A.; Lacapère, J.J.; Combadière, C.; Deterre, P. Subtle conformational changes between CX3CR1 genetic variants as revealed by resonance energy transfer assays. FASEB J. 2010, 24, 4585–4598. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, K.; Fujita, Y.; Kawabata, T.; Kimura, H.; Iwayama, Y.; Inada, T.; Okahisa, Y.; Egawa, J.; Usami, M.; Kushima, I.; et al. Rare genetic variants in CX3CR1 and their contribution to the increased risk of schizophrenia and autism spectrum disorders. Transl. Psychiatry 2017, 7, e1184. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Latha, K.; Manyam, G.; Yang, Y.; Rao, A.; Rao, G. Role of CX3CR1 signaling in malignant transformation of gliomas. Neuro. Oncol. 2020, 22, 1463–1473. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Fuentes, S.; Salgado-Aguayo, A.; Arratia-Quijada, J.; Gorocica-Rosete, P. Regulation and biological functions of the CX3CL1-CX3CR1 axis and its relevance in solid cancer: A mini-review. J. Cancer 2021, 12, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Kehlen, A.; Haegele, M.; Böhme, L.; Cynis, H.; Hoffmann, T.; Demuth, H.U. N-terminal pyroglutamate formation in CX3CL1 is essential for its full biologic activity. Biosci. Rep. 2017, 37, BSR20170712. [Google Scholar] [CrossRef] [PubMed]
- Maciejewski-Lenoir, D.; Chen, S.; Feng, L.; Maki, R.; Bacon, K.B. Characterization of fractalkine in rat brain cells: Migratory and activation signals for CX3CR-1-expressing microglia. J. Immunol. 1999, 163, 1628–1635. [Google Scholar] [CrossRef]
- Pallandre, J.R.; Krzewski, K.; Bedel, R.; Ryffel, B.; Caignard, A.; Rohrlich, P.S.; Pivot, X.; Tiberghien, P.; Zitvogel, L.; Strominger, J.L.; et al. Dendritic cell and natural killer cell cross-talk: A pivotal role of CX3CL1 in NK cytoskeleton organization and activation. Blood 2008, 112, 4420–4424. [Google Scholar] [CrossRef]
- Foussat, A.; Coulomb-L’Hermine, A.; Gosling, J.; Krzysiek, R.; Durand-Gasselin, I.; Schall, T.; Balian, A.; Richard, Y.; Galanaud, P.; Emilie, D. Fractalkine receptor expression by T lymphocyte subpopulations and in vivo production of fractalkine in human. Eur. J. Immunol. 2000, 30, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Chidambaram, H.; Das, R.; Chinnathambi, S. Interaction of Tau with the chemokine receptor, CX3CR1 and its effect on microglial activation, migration and proliferation. Cell Biosci. 2020, 10, 109. [Google Scholar] [CrossRef]
- Chen, Y.; Green, S.R.; Almazan, F.; Quehenberger, O. The amino terminus and the third extracellular loop of CX3CR1 contain determinants critical for distinct receptor functions. Mol. Pharmacol. 2006, 69, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Kharche, S.; Joshi, M.; Chattopadhyay, A.; Sengupta, D. Conformational plasticity and dynamic interactions of the N-terminal domain of the chemokine receptor CXCR1. PLoS Comput. Biol. 2021, 17, e1008593. [Google Scholar]
- Srivastava, D.; Gakhar, L.; Artemyev, N.O. Structural underpinnings of Ric8A function as a G-protein α-subunit chaperone and guanine-nucleotide exchange factor. Nat. Commun. 2019, 10, 3084. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.J.; Inchausti, R.; Eaton, C.J.; Krystofova, S.; Borkovich, K.A. RIC8 is a guanine-nucleotide exchange factor for Galpha subunits that regulates growth and development in Neurospora crassa. Genetics 2011, 189, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Weis, W.I.; Kobilka, B.K. The Molecular Basis of G Protein-Coupled Receptor Activation. Annu. Rev. Biochem. 2018, 87, 897–919. [Google Scholar] [CrossRef] [PubMed]
- Arnoux, I.; Audinat, E. Fractalkine Signaling and Microglia Functions in the Developing Brain. Neural Plast. 2015, 2015, 689404. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Morinaga, H.; Kim, J.J.; Lagakos, W.; Taylor, S.; Keshwani, M.; Perkins, G.; Dong, H.; Kayali, A.G.; Sweet, I.R.; et al. The fractalkine/CX3CR1 system regulates β cell function and insulin secretion. Cell 2013, 153, 413–425. [Google Scholar] [CrossRef]
- Wang, A.; Yang, T.; Zhang, L.; Jia, L.; Wu, Q.; Yao, S.; Xu, J.; Yang, H. IP3-Mediated Calcium Signaling Is Involved in the Mechanism of Fractalkine-Induced Hyperalgesia Response. Med. Sci. Monit. 2018, 24, 8804–8811. [Google Scholar] [CrossRef]
- Wojdasiewicz, P.; Turczyn, P.; Dobies-Krzesniak, B.; Frasunska, J.; Tarnacka, B. Role of CX3CL1/CX3CR1 Signaling Axis Activity in Osteoporosis. Mediat. Inflamm. 2019, 2019, 7570452. [Google Scholar] [CrossRef]
- Lyons, A.; Lynch, A.M.; Downer, E.J.; Hanley, R.; O’Sullivan, J.B.; Smith, A.; Lynch, M.A. Fractalkine-induced activation of the phosphatidylinositol-3 kinase pathway attentuates microglial activation in vivo and in vitro. J. Neurochem. 2009, 110, 1547–1556. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Song, K.H.; Ha, H. Fractalkine increases mesangial cell proliferation through reactive oxygen species and mitogen-activated protein kinases. Transplant. Proc. 2012, 44, 1026–1028. [Google Scholar] [CrossRef] [PubMed]
- Garton, K.J.; Gough, P.J.; Blobel, C.P.; Murphy, G.; Greaves, D.R.; Dempsey, P.J.; Raines, E.W. Tumor necrosis factor-alpha-converting enzyme (ADAM17) mediates the cleavage and shedding of fractalkine (CX3CL1). J. Biol. Chem. 2001, 276, 37993–38001. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Namkoong, S.; Kim, Y.M.; Kim, C.K.; Lee, H.; Ha, K.S.; Chung, H.T.; Kwon, Y.G.; Kim, Y.M. Fractalkine stimulates angiogenesis by activating the Raf-1/MEK/ERK- and PI3K/Akt/eNOS-dependent signal pathways. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H2836-46. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.K.; Srivastava, R.K.; Shankar, S. Inhibition of PI3K/AKT and MAPK/ERK pathways causes activation of FOXO transcription factor, leading to cell cycle arrest and apoptosis in pancreatic cancer. J. Mol. Signal. 2010, 5, 10. [Google Scholar] [CrossRef]
- Wang, H.; Cai, J.; Du, S.; Guo, Z.; Xin, B.; Wang, J.; Wei, W.; Shen, X. Fractalkine/CX3CR1 induces apoptosis resistance and proliferation through the activation of the AKT/NF-κB cascade in pancreatic cancer cells. Cell Biochem. Funct. 2017, 35, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Landsman, L.; Bar-On, L.; Zernecke, A.; Kim, K.W.; Krauthgamer, R.; Shagdarsuren, E.; Lira, S.A.; Weissman, I.L.; Weber, C.; Jung, S. CX3CR1 is required for monocyte homeostasis and atherogenesis by promoting cell survival. Blood 2009, 113, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.S.; Khuri, F.R.; Herbst, R.S. Epidermal growth factor receptor biology (IMC-C225). Curr. Opin. Oncol. 2001, 13, 506–513. [Google Scholar] [CrossRef]
- Cai, Z.; Zhang, H.; Liu, J.; Berezov, A.; Murali, R.; Wang, Q.; Greene, M.I. Targeting erbB receptors. Semin. Cell Dev. Biol. 2010, 21, 961–966. [Google Scholar] [CrossRef]
- Ledonne, A.; Mercuri, N.B. Insights on the Functional Interaction between Group 1 Metabotropic Glutamate Receptors (mGluRI) and ErbB Receptors. Int. J. Mol. Sci. 2020, 21, 7913. [Google Scholar] [CrossRef] [PubMed]
- Oda, K.; Matsuoka, Y.; Funahashi, A.; Kitano, H. A comprehensive pathway map of epidermal growth factor receptor signaling. Mol. Syst. Biol. 2005, 1, 2005.0010. [Google Scholar] [CrossRef]
- Thomas, S.M.; Bhola, N.E.; Zhang, Q.; Contrucci, S.C.; Wentzel, A.L.; Freilino, M.L.; Gooding, W.E.; Siegfried, J.M.; Chan, D.C.; Grandis, J.R. Cross-talk between G protein-coupled receptor and epidermal growth factor receptor signaling pathways contributes to growth and invasion of head and neck squamous cell carcinoma. Cancer Res. 2006, 66, 11831–11839. [Google Scholar] [CrossRef] [PubMed]
- Cantor, A.J.; Shah, N.H.; Kuriyan, J. Deep mutational analysis reveals functional trade-offs in the sequences of EGFR autophosphorylation sites. Proc. Natl. Acad. Sci. USA 2018, 115, E7303–E7312. [Google Scholar] [CrossRef] [PubMed]
- Jurišić, V.; Obradovic, J.; Pavlović, S.; Djordjevic, N. Epidermal Growth Factor Receptor Gene in Non-Small-Cell Lung Cancer: The Importance of Promoter Polymorphism Investigation. Anal. Cell Pathol. 2018, 2018, 6192187. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Berezov, A.; Wang, Q.; Zhang, G.; Drebin, J.; Murali, R.; Greene, M.I. ErbB receptors: From oncogenes to targeted cancer therapies. J. Clin. Investig. 2007, 117, 2051–2058. [Google Scholar] [CrossRef] [PubMed]
- Tardáguila, M.; Mira, E.; García-Cabezas, M.A.; Feijoo, A.M.; Quintela-Fandino, M.; Azcoitia, I.; Lira, S.A.; Mañes, S. CX3CL1 promotes breast cancer via transactivation of the EGF pathway. Cancer Res. 2013, 73, 4461–4473. [Google Scholar] [CrossRef] [PubMed]
- Bai, Q.; Wang, J.; Zhou, X. EGFR exon20 insertion mutations in non-small cell lung cancer: Clinical implications and recent advances in targeted therapies. Cancer Treat. Rev. 2023, 120, 102605. [Google Scholar] [CrossRef] [PubMed]
- Gschwind, A.; Zwick, E.; Prenzel, N.; Leserer, M.; Ullrich, A. Cell communication networks: Epidermal growth factor receptor transactivation as the paradigm for interreceptor signal transmission. Oncogene 2001, 20, 1594–1600. [Google Scholar] [CrossRef]
- Burch, M.L.; Osman, N.; Getachew, R.; Al-Aryahi, S.; Poronnik, P.; Zheng, W.; Hill, M.A.; Little, P.J. G protein coupled receptor transactivation: Extending the paradigm to include serine/threonine kinase receptors. Int. J. Biochem. Cell Biol. 2012, 44, 722–727. [Google Scholar] [CrossRef]
- Chandrasekar, B.; Mummidi, S.; Perla, R.P.; Bysani, S.; Dulin, N.O.; Liu, F.; Melby, P.C. Fractalkine (CX3CL1) stimulated by nuclear factor kappaB (NF-kappaB)-dependent inflammatory signals induces aortic smooth muscle cell proliferation through an autocrine pathway. Biochem. J. 2003, 373 Pt 2, 547–558. [Google Scholar] [CrossRef]
- Szukiewicz, D.; Wojciechowska, M.; Bilska, A.; Stangret, A.; Szewczyk, G.; Mittal, T.K.; Watroba, M.; Kochanowski, J. Aspirin Action in Endothelial Cells: Different Patterns of Response Between Chemokine CX3CL1/CX3CR1 and TNF-α/TNFR1 Signaling Pathways. Cardiovasc. Drugs Ther. 2015, 29, 219–229. [Google Scholar] [CrossRef]
- Zwijnenburg, A.J.; Pokharel, J.; Varnaitė, R.; Zheng, W.; Hoffer, E.; Shryki, I.; Comet, N.R.; Ehrström, M.; Gredmark-Russ, S.; Eidsmo, L.; et al. Graded expression of the chemokine receptor CX3CR1 marks differentiation states of human and murine T cells and enables cross-species interpretation. Immunity 2023, 56, 1955–1974.e10. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.; Zhang, J.; Dai, S.; Wang, C.; Chen, Z.; Zhang, S.; Xu, J.; Qin, X.; Liu, F. CX3CR1 deficiency exacerbates immune-mediated hepatitis by increasing NF-κB-mediated cytokine production in macrophage and T cell. Exp. Biol. Med. 2023, 248, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Wojdasiewicz, P.; Poniatowski, Ł.A.; Kotela, A.; Skoda, M.; Pyzlak, M.; Stangret, A.; Kotela, I.; Szukiewicz, D. Comparative Analysis of the Occurrence and Role of CX3CL1 (Fractalkine) and Its Receptor CX3CR1 in Hemophilic Arthropathy and Osteoarthritis. J. Immunol. Res. 2020, 2020, 2932696, Erratum in J. Immunol. Res. 2020, 2020, 7179283. [Google Scholar] [CrossRef]
- White, G.E.; Tan, T.C.; John, A.E.; Whatling, C.; McPheat, W.L.; Greaves, D.R. Fractalkine has anti-apoptotic and proliferative effects on human vascular smooth muscle cells via epidermal growth factor receptor signalling. Cardiovasc. Res. 2010, 85, 825–835. [Google Scholar] [CrossRef]
- White, G.E.; McNeill, E.; Channon, K.M.; Greaves, D.R. Fractalkine promotes human monocyte survival via a reduction in oxidative stress. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2554–2562. [Google Scholar] [CrossRef] [PubMed]
- Raei Sadigh, A.; Darabi, M.; Salmassi, A.; Hamdi, K.; Farzadi, L.; Ghasemzadeh, A.; Fattahi, A.; Nouri, M. Fractalkine and apoptotic/anti-apoptotic markers in granulosa cells of women with polycystic ovarian syndrome. Mol. Biol. Rep. 2020, 47, 3593–3603. [Google Scholar] [CrossRef]
- Sheridan, G.K.; Murphy, K.J. Neuron-glia crosstalk in health and disease: Fractalkine and CX3CR1 take centre stage. Open Biol. 2013, 3, 130181. [Google Scholar] [CrossRef]
- Pawelec, P.; Ziemka-Nalecz, M.; Sypecka, J.; Zalewska, T. The Impact of the CX3CL1/CX3CR1 Axis in Neurological Disorders. Cells 2020, 9, 2277. [Google Scholar] [CrossRef]
- Paolicelli, R.C.; Bisht, K.; Tremblay, M.È. Fractalkine regulation of microglial physiology and consequences on the brain and behavior. Front. Cell Neurosci. 2014, 8, 129. [Google Scholar] [CrossRef]
- Camacho-Hernández, N.P.; Peña-Ortega, F. Fractalkine/CX3CR1-Dependent Modulation of Synaptic and Network Plasticity in Health and Disease. Neural Plast. 2023, 2023, 4637073. [Google Scholar] [CrossRef]
- Mizuno, T.; Kawanokuchi, J.; Numata, K.; Suzumura, A. Production and neuroprotective functions of fractalkine in the central nervous system. Brain Res. 2003, 979, 65–70. [Google Scholar] [CrossRef]
- Mecca, C.; Giambanco, I.; Donato, R.; Arcuri, C. Microglia and Aging: The Role of the TREM2-DAP12 and CX3CL1-CX3CR1 Axes. Int. J. Mol. Sci. 2018, 19, 318. [Google Scholar] [CrossRef]
- Subbarayan, M.S.; Joly-Amado, A.; Bickford, P.C.; Nash, K.R. CX3CL1/CX3CR1 signaling targets for the treatment of neurodegenerative diseases. Pharmacol. Ther. 2022, 231, 107989. [Google Scholar] [CrossRef]
- Cipriani, R.; Villa, P.; Chece, G.; Lauro, C.; Paladini, A.; Micotti, E.; Perego, C.; De Simoni, M.G.; Fredholm, B.B.; Eusebi, F.; et al. CX3CL1 is neuroprotective in permanent focal cerebral ischemia in rodents. J. Neurosci. 2011, 31, 16327–16335. [Google Scholar] [CrossRef]
- Luo, P.; Chu, S.F.; Zhang, Z.; Xia, C.Y.; Chen, N.H. Fractalkine/CX3CR1 is involved in the cross-talk between neuron and glia in neurological diseases. Brain Res. Bull. 2019, 146, 12–21. [Google Scholar] [CrossRef]
- Bivona, G.; Iemmolo, M.; Ghersi, G. CX3CL1 Pathway as a Molecular Target for Treatment Strategies in Alzheimer’s Disease. Int. J. Mol. Sci. 2023, 24, 8230. [Google Scholar] [CrossRef]
- Nash, K.R.; Moran, P.; Finneran, D.J.; Hudson, C.; Robinson, J.; Morgan, D.; Bickford, P.C. Fractalkine over expression suppresses α-synuclein-mediated neurodegeneration. Mol. Ther. 2015, 23, 17–23. [Google Scholar] [CrossRef]
- Juliani, J.; Vassileff, N.; Spiers, J.G. Inflammatory-Mediated Neuron-Glia Communication Modulates ALS Pathophysiology. J. Neurosci. 2021, 41, 1142–1144. [Google Scholar] [CrossRef]
- Lee, S.; Varvel, N.H.; Konerth, M.E.; Xu, G.; Cardona, A.E.; Ransohoff, R.M.; Lamb, B.T. CX3CR1 deficiency alters microglial activation and reduces beta-amyloid deposition in two Alzheimer’s disease mouse models. Am. J. Pathol. 2010, 177, 2549–2562. [Google Scholar] [CrossRef]
- Cho, S.H.; Sun, B.; Zhou, Y.; Kauppinen, T.M.; Halabisky, B.; Wes, P.; Ransohoff, R.M.; Gan, L. CX3CR1 protein signaling modulates microglial activation and protects against plaque-independent cognitive deficits in a mouse model of Alzheimer disease. J. Biol. Chem. 2011, 286, 32713–32722. [Google Scholar] [CrossRef]
- Fuhrmann, M.; Bittner, T.; Jung, C.K.; Burgold, S.; Page, R.M.; Mitteregger, G.; Haass, C.; LaFerla, F.M.; Kretzschmar, H.; Herms, J. Microglial Cx3cr1 knockout prevents neuron loss in a mouse model of Alzheimer’s disease. Nat. Neurosci. 2010, 13, 411–413. [Google Scholar] [CrossRef]
- Eugenín, J.; Eugenín-von Bernhardi, L.; von Bernhardi, R. Age-dependent changes on fractalkine forms and their contribution to neurodegenerative diseases. Front. Mol. Neurosci. 2023, 16, 1249320. [Google Scholar] [CrossRef]
- Stratoulias, V.; Venero, J.L.; Tremblay, M.È.; Joseph, B. Microglial subtypes: Diversity within the microglial community. EMBO J. 2019, 38, e101997. [Google Scholar] [CrossRef]
- Jacquelin, S.; Licata, F.; Dorgham, K.; Hermand, P.; Poupel, L.; Guyon, E.; Deterre, P.; Hume, D.A.; Combadière, C.; Boissonnas, A. CX3CR1 reduces Ly6Chigh-monocyte motility within and release from the bone marrow after chemotherapy in mice. Blood 2013, 122, 674–683. [Google Scholar] [CrossRef]
- Hoshino, A.; Ueha, S.; Hanada, S.; Imai, T.; Ito, M.; Yamamoto, K.; Matsushima, K.; Yamaguchi, A.; Iimura, T. Roles of chemokine receptor CX3CR1 in maintaining murine bone homeostasis through the regulation of both osteoblasts and osteoclasts. J. Cell Sci. 2013, 126 Pt 4, 1032–1045. [Google Scholar] [CrossRef]
- Kuboi, Y.; Kuroda, Y.; Ohkuro, M.; Motoi, S.; Tomimori, Y.; Yasuda, H.; Yasuda, N.; Imai, T.; Matsuo, K. The Fractalkine-CX3CR1 Axis Regulates Non-inflammatory Osteoclastogenesis by Enhancing Precursor Cell Survival. JBMR Plus 2022, 6, e10680. [Google Scholar] [CrossRef]
- Yano, R.; Yamamura, M.; Sunahori, K.; Takasugi, K.; Yamana, J.; Kawashima, M.; Makino, H. Recruitment of CD16+ monocytes into synovial tissues is mediated by fractalkine and CX3CR1 in rheumatoid arthritis patients. Acta Med. Okayama 2007, 61, 89–98. [Google Scholar] [CrossRef]
- Marchica, V.; Toscani, D.; Corcione, A.; Bolzoni, M.; Storti, P.; Vescovini, R.; Ferretti, E.; Dalla Palma, B.; Vicario, E.; Accardi, F.; et al. Bone Marrow CX3CL1/Fractalkine is a New Player of the Pro-Angiogenic Microenvironment in Multiple Myeloma Patients. Cancers 2019, 11, 321. [Google Scholar] [CrossRef]
- Sciumè, G.; De Angelis, G.; Benigni, G.; Ponzetta, A.; Morrone, S.; Santoni, A.; Bernardini, G. CX3CR1 expression defines 2 KLRG1+ mouse NK-cell subsets with distinct functional properties and positioning in the bone marrow. Blood 2011, 117, 4467–4475. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, J.; Zhou, Z.; Zhou, H.; Wang, Y.; Gong, Z.; Zhu, J. Fractalkine promotes chemotaxis of bone marrow-derived mesenchymal stem cells towards ischemic brain lesions through Jak2 signaling and cytoskeletal reorganization. FEBS J. 2015, 282, 891–903. [Google Scholar] [CrossRef]
- Teupser, D.; Pavlides, S.; Tan, M.; Gutierrez-Ramos, J.C.; Kolbeck, R.; Breslow, J.L. Major reduction of atherosclerosis in fractalkine (CX3CL1)-deficient mice is at the brachiocephalic artery, not the aortic root. Proc. Natl. Acad. Sci. USA 2004, 101, 17795–17800. [Google Scholar] [CrossRef]
- Ma, J.; Luo, J.; Sun, Y.; Zhao, Z. Cytokines associated with immune response in atherosclerosis. Am. J. Transl. Res. 2022, 14, 6424–6444. [Google Scholar]
- Riopel, M.; Vassallo, M.; Ehinger, E.; Pattison, J.; Bowden, K.; Winkels, H.; Wilson, M.; de Jong, R.; Patel, S.; Balakrishna, D.; et al. CX3CL1-Fc treatment prevents atherosclerosis in Ldlr KO mice. Mol. Metab. 2019, 20, 89–101. [Google Scholar] [CrossRef]
- Liu, H.; Jiang, D. Fractalkine/CX3CR1 and atherosclerosis. Clin. Chim. Acta 2011, 412, 1180–1186. [Google Scholar] [CrossRef]
- Elliott, M.R.; Koster, K.M.; Murphy, P.S. Efferocytosis Signaling in the Regulation of Macrophage Inflammatory Responses. J. Immunol. 2017, 198, 1387–1394. [Google Scholar] [CrossRef]
- Lucas, A.D.; Bursill, C.; Guzik, T.J.; Sadowski, J.; Channon, K.M.; Greaves, D.R. Smooth muscle cells in human atherosclerotic plaques express the fractalkine receptor CX3CR1 and undergo chemotaxis to the CX3C chemokine fractalkine (CX3CL1). Circulation 2003, 108, 2498–2504. [Google Scholar] [CrossRef]
- Harman, J.L.; Jørgensen, H.F. The role of smooth muscle cells in plaque stability: Therapeutic targeting potential. Br. J. Pharmacol. 2019, 176, 3741–3753. [Google Scholar] [CrossRef]
- Apostolakis, S.; Spandidos, D. Chemokines and atherosclerosis: Focus on the CX3CL1/CX3CR1 pathway. Acta Pharmacol. Sin. 2013, 34, 1251–1256. [Google Scholar] [CrossRef]
- Skoda, M.; Stangret, A.; Szukiewicz, D. Fractalkine and placental growth factor: A duet of inflammation and angiogenesis in cardiovascular disorders. Cytokine Growth Factor Rev. 2018, 39, 116–123. [Google Scholar] [CrossRef]
- Noels, H.; Weber, C.; Koenen, R.R. Chemokines as Therapeutic Targets in Cardiovascular Disease. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 583–592. [Google Scholar] [CrossRef]
- Flamant, M.; Mougenot, N.; Balse, E.; Le Fèvre, L.; Atassi, F.; Gautier, E.L.; Le Goff, W.; Keck, M.; Nadaud, S.; Combadière, C.; et al. Early activation of the cardiac CX3CL1/CX3CR1 axis delays β-adrenergic-induced heart failure. Sci. Rep. 2021, 11, 17982. [Google Scholar] [CrossRef]
- Njerve, I.U.; Solheim, S.; Lunde, K.; Hoffmann, P.; Arnesen, H.; Seljeflot, I. Fractalkine levels are elevated early after PCI-treated ST-elevation myocardial infarction; no influence of autologous bone marrow derived stem cell injection. Cytokine 2014, 69, 131–135. [Google Scholar] [CrossRef]
- Yao, K.; Zhang, S.; Lu, H.; Hong, X.; Qian, J.; Sun, A.; Zou, Y.; Ge, J. Changes in fractalkine in patients with ST-elevation myocardial infarction. Coron. Artery Dis. 2015, 26, 516–520. [Google Scholar] [CrossRef]
- Xu, B.; Qian, Y.; Zhao, Y.; Fang, Z.; Tang, K.; Zhou, N.; Li, D.; Wang, J. Prognostic value of fractalkine/CX3CL1 concentration in patients with acute myocardial infarction treated with primary percutaneous coronary intervention. Cytokine 2019, 113, 365–370. [Google Scholar] [CrossRef]
- Boag, S.E.; Das, R.; Shmeleva, E.V.; Bagnall, A.; Egred, M.; Howard, N.; Bennaceur, K.; Zaman, A.; Keavney, B.; Spyridopoulos, I. T lymphocytes and fractalkine contribute to myocardial ischemia/reperfusion injury in patients. J. Clin. Investig. 2015, 125, 3063–3076. [Google Scholar] [CrossRef]
- Furio, E.; García-Fuster, M.J.; Redon, J.; Marques, P.; Ortega, R.; Sanz, M.J.; Piqueras, L. CX3CR1/CX3CL1 Axis Mediates Platelet-Leukocyte Adhesion to Arterial Endothelium in Younger Patients with a History of Idiopathic Deep Vein Thrombosis. Thromb. Haemost. 2018, 118, 562–571. [Google Scholar] [CrossRef]
- Marques, P.; Collado, A.; Martinez-Hervás, S.; Domingo, E.; Benito, E.; Piqueras, L.; Real, J.T.; Ascaso, J.F.; Sanz, M.J. Systemic Inflammation in Metabolic Syndrome: Increased Platelet and Leukocyte Activation, and Key Role of CX3CL1/CX3CR1 and CCL2/CCR2 Axes in Arterial Platelet-Proinflammatory Monocyte Adhesion. J. Clin. Med. 2019, 8, 708. [Google Scholar] [CrossRef]
- Zhang, J.; Patel, J.M. Role of the CX3CL1-CX3CR1 axis in chronic inflammatory lung diseases. Int. J. Clin. Exp. Med. 2010, 3, 233–244. [Google Scholar]
- Balabanian, K.; Foussat, A.; Dorfmüller, P.; Durand-Gasselin, I.; Capel, F.; Bouchet-Delbos, L.; Portier, A.; Marfaing-Koka, A.; Krzysiek, R.; Rimaniol, A.C.; et al. CX(3)C chemokine fractalkine in pulmonary arterial hypertension. Am. J. Respir. Crit. Care Med. 2002, 165, 1419–1425. [Google Scholar] [CrossRef]
- Amsellem, V.; Abid, S.; Poupel, L.; Parpaleix, A.; Rodero, M.; Gary-Bobo, G.; Latiri, M.; Dubois-Rande, J.L.; Lipskaia, L.; Combadiere, C.; et al. Roles for the CX3CL1/CX3CR1 and CCL2/CCR2 Chemokine Systems in Hypoxic Pulmonary Hypertension. Am. J. Respir. Cell Mol. Biol. 2017, 56, 597–608. [Google Scholar] [CrossRef]
- Tsai, W.H.; Chang, S.C.; Lin, Y.C.; Hsu, H.C. CX3CL1(+) Microparticles-Induced MFG-E8 Enhances Apoptotic Cell Clearance by Alveolar Macrophages. Cells 2021, 10, 2583. [Google Scholar] [CrossRef]
- El-Shazly, A.; Berger, P.; Girodet, P.O.; Ousova, O.; Fayon, M.; Vernejoux, J.M.; Marthan, R.; Tunon-de-Lara, J.M. Fraktalkine produced by airway smooth muscle cells contributes to mast cell recruitment in asthma. J. Immunol. 2006, 176, 1860–1868. [Google Scholar] [CrossRef]
- Godwin, M.S.; Jones, M.; Blackburn, J.P.; Yu, Z.; Matalon, S.; Hastie, A.T.; Meyers, D.A.; Steele, C. The chemokine CX3CL1/fractalkine regulates immunopathogenesis during fungal-associated allergic airway inflammation. Am. J. Physiol. Lung Cell Mol. Physiol. 2021, 320, L393–L404. [Google Scholar] [CrossRef]
- Upton, N.; Jackson, D.J.; Nikonova, A.A.; Hingley-Wilson, S.; Khaitov, M.; Del Rosario, A.; Traub, S.; Trujillo-Torralbo, M.B.; Habibi, M.; Elkin, S.L.; et al. Rhinovirus induction of fractalkine (CX3CL1) in airway and peripheral blood mononuclear cells in asthma. PLoS ONE 2017, 12, e0183864. [Google Scholar] [CrossRef]
- Efsen, E.; Grappone, C.; DeFranco, R.M.; Milani, S.; Romanelli, R.G.; Bonacchi, A.; Caligiuri, A.; Failli, P.; Annunziato, F.; Pagliai, G.; et al. Up-regulated expression of fractalkine and its receptor CX3CR1 during liver injury in humans. J. Hepatol. 2002, 37, 39–47. [Google Scholar] [CrossRef]
- Sutti, S.; Locatelli, I.; Bruzzì, S.; Jindal, A.; Vacchiano, M.; Bozzola, C.; Albano, E. CX3CR1-expressing inflammatory dendritic cells contribute to the progression of steatohepatitis. Clin. Sci. 2015, 129, 797–808. [Google Scholar] [CrossRef]
- Nagata, N.; Chen, G.; Xu, L.; Ando, H. An Update on the Chemokine System in the Development of NAFLD. Medicina 2022, 58, 761. [Google Scholar] [CrossRef]
- Aoyama, T.; Inokuchi, S.; Brenner, D.A.; Seki, E. CX3CL1-CX3CR1 interaction prevents carbon tetrachloride-induced liver inflammation and fibrosis in mice. Hepatology 2010, 52, 1390–1400. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, B.J.; Wang, J.Z.; Xie, X.M.; Tong, Q.X. Association of CX3CL1 and CX3CR1 Expression with Liver Fibrosis in a Mouse Model of Schistosomiasis. Curr. Med. Sci. 2020, 40, 1121–1127. [Google Scholar] [CrossRef] [PubMed]
- Karlmark, K.R.; Zimmermann, H.W.; Roderburg, C.; Gassler, N.; Wasmuth, H.E.; Luedde, T.; Trautwein, C.; Tacke, F. The fractalkine receptor CX₃CR1 protects against liver fibrosis by controlling differentiation and survival of infiltrating hepatic monocytes. Hepatology 2010, 52, 1769–1782. [Google Scholar] [CrossRef]
- Wasmuth, H.E.; Zaldivar, M.M.; Berres, M.L.; Werth, A.; Scholten, D.; Hillebrandt, S.; Tacke, F.; Schmitz, P.; Dahl, E.; Wiederholt, T.; et al. The fractalkine receptor CX3CR1 is involved in liver fibrosis due to chronic hepatitis C infection. J. Hepatol. 2008, 48, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Hassan, G.S.; Flores Molina, M.; Shoukry, N.H. The multifaceted role of macrophages during acute liver injury. Front. Immunol. 2023, 14, 1237042. [Google Scholar] [CrossRef]
- Isse, K.; Harada, K.; Zen, Y.; Kamihira, T.; Shimoda, S.; Harada, M.; Nakanuma, Y. Fractalkine and CX3CR1 are involved in the recruitment of intraepithelial lymphocytes of intrahepatic bile ducts. Hepatology 2005, 41, 506–516. [Google Scholar] [CrossRef]
- Shimoda, S.; Harada, K.; Niiro, H.; Taketomi, A.; Maehara, Y.; Tsuneyama, K.; Kikuchi, K.; Nakanuma, Y.; Mackay, I.R.; Gershwin, M.E.; et al. CX3CL1 (fractalkine): A signpost for biliary inflammation in primary biliary cirrhosis. Hepatology 2010, 51, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Joeris, T.; Müller-Luda, K.; Agace, W.W.; Mowat, A.M. Diversity and functions of intestinal mononuclear phagocytes. Mucosal Immunol. 2017, 10, 845–864. [Google Scholar] [CrossRef]
- Bain, C.C.; Mowat, A.M. Intestinal macrophages—Specialised adaptation to a unique environment. Eur. J. Immunol. 2011, 41, 2494–2498. [Google Scholar] [CrossRef]
- Ferretti, E.; Pistoia, V.; Corcione, A. Role of fractalkine/CX3CL1 and its receptor in the pathogenesis of inflammatory and malignant diseases with emphasis on B cell malignancies. Mediat. Inflamm. 2014, 2014, 480941. [Google Scholar] [CrossRef]
- Bain, C.C.; Scott, C.L.; Uronen-Hansson, H.; Gudjonsson, S.; Jansson, O.; Grip, O.; Guilliams, M.; Malissen, B.; Agace, W.W.; Mowat, A.M. Resident and pro-inflammatory macrophages in the colon represent alternative context-dependent fates of the same Ly6Chi monocyte precursors. Mucosal Immunol. 2013, 6, 498–510. [Google Scholar] [CrossRef]
- Bain, C.C.; Bravo-Blas, A.; Scott, C.L.; Perdiguero, E.G.; Geissmann, F.; Henri, S.; Malissen, B.; Osborne, L.C.; Artis, D.; Mowat, A.M. Constant replenishment from circulating monocytes maintains the macrophage pool in the intestine of adult mice. Nat. Immunol. 2014, 15, 929–937. [Google Scholar] [CrossRef]
- Sans, M.; Danese, S.; de la Motte, C.; de Souza, H.S.; Rivera-Reyes, B.M.; West, G.A.; Phillips, M.; Katz, J.A.; Fiocchi, C. Enhanced recruitment of CX3CR1+ T cells by mucosal endothelial cell-derived fractalkine in inflammatory bowel disease. Gastroenterology 2007, 132, 139–153. [Google Scholar] [CrossRef]
- Siwetz, M.; Blaschitz, A.; Kremshofer, J.; Bilic, J.; Desoye, G.; Huppertz, B.; Gauster, M. Metalloprotease dependent release of placenta derived fractalkine. Mediat. Inflamm. 2014, 2014, 839290. [Google Scholar] [CrossRef]
- Siwetz, M.; Sundl, M.; Kolb, D.; Hiden, U.; Herse, F.; Huppertz, B.; Gauster, M. Placental fractalkine mediates adhesion of THP-1 monocytes to villous trophoblast. Histochem. Cell Biol. 2015, 143, 565–574. [Google Scholar] [CrossRef]
- Siwetz, M.; Dieber-Rotheneder, M.; Cervar-Zivkovic, M.; Kummer, D.; Kremshofer, J.; Weiss, G.; Herse, F.; Huppertz, B.; Gauster, M. Placental fractalkine is up-regulated in severe early-onset preeclampsia. Am. J. Pathol. 2015, 185, 1334–1343. [Google Scholar] [CrossRef]
- Vishnyakova, P.; Poltavets, A.; Nikitina, M.; Muminova, K.; Potapova, A.; Vtorushina, V.; Loginova, N.; Midiber, K.; Mikhaleva, L.; Lokhonina, A.; et al. Preeclampsia: Inflammatory signature of decidual cells in early manifestation of disease. Placenta 2021, 104, 277–283. [Google Scholar] [CrossRef]
- Szewczyk, G.; Pyzlak, M.; Pankiewicz, K.; Szczerba, E.; Stangret, A.; Szukiewicz, D.; Skoda, M.; Bierła, J.; Cukrowska, B.; Fijałkowska, A. The potential association between a new angiogenic marker fractalkine and a placental vascularization in preeclampsia. Arch. Gynecol. Obstet. 2021, 304, 365–376. [Google Scholar] [CrossRef]
- Ullah, A.; Zhao, J.; Singla, R.K.; Shen, B. Pathophysiological impact of CXC and CX3CL1 chemokines in preeclampsia and gestational diabetes mellitus. Front. Cell Dev. Biol. 2023, 11, 1272536. [Google Scholar] [CrossRef]
- Szukiewicz, D.; Kochanowski, J.; Pyzlak, M.; Szewczyk, G.; Stangret, A.; Mittal, T.K. Fractalkine (CX3CL1) and its receptor CX3CR1 may contribute to increased angiogenesis in diabetic placenta. Mediat. Inflamm. 2013, 2013, 437576. [Google Scholar] [CrossRef]
- Szukiewicz, D.; Kochanowski, J.; Mittal, T.K.; Pyzlak, M.; Szewczyk, G.; Cendrowski, K. CX3CL1 (fractalkine) and TNFα production by perfused human placental lobules under normoxic and hypoxic conditions in vitro: The importance of CX3CR1 signaling. Inflamm. Res. 2014, 63, 179–189. [Google Scholar] [CrossRef]
- Gokce, S.; Herkiloglu, D.; Cevik, O.; Turan, V. Role of chemokines in early pregnancy loss. Exp. Ther. Med. 2022, 23, 397. [Google Scholar] [CrossRef]
- Tanaka, Y.; Hoshino-Negishi, K.; Kuboi, Y.; Tago, F.; Yasuda, N.; Imai, T. Emerging Role of Fractalkine in the Treatment of Rheumatic Diseases. Immunotargets Ther. 2020, 9, 241–253. [Google Scholar] [CrossRef]
- Koizumi, K.; Saitoh, Y.; Minami, T.; Takeno, N.; Tsuneyama, K.; Miyahara, T.; Nakayama, T.; Sakurai, H.; Takano, Y.; Nishimura, M.; et al. Role of CX3CL1/fractalkine in osteoclast differentiation and bone resorption. J. Immunol. 2009, 183, 7825–7831. [Google Scholar] [CrossRef]
- Guo, Y.N.; Cui, S.J.; Tian, Y.J.; Zhao, N.R.; Zhang, Y.D.; Gan, Y.H.; Zhou, Y.H.; Wang, X.D. Chondrocyte apoptosis in temporomandibular joint osteoarthritis promotes bone resorption by enhancing chemotaxis of osteoclast precursors. Osteoarthr. Cartilage. 2022, 30, 1140–1153. [Google Scholar] [CrossRef]
- Wojdasiewicz, P.; Poniatowski, Ł.A.; Szukiewicz, D. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediat. Inflamm. 2014, 2014, 561459. [Google Scholar] [CrossRef]
- Lu, Z.; Zhang, A.; Dai, Y. CX3CL1 deficiency ameliorates inflammation, apoptosis and accelerates osteogenic differentiation, mineralization in LPS-treated MC3T3-E1 cells via its receptor CX3CR1. Ann. Anat. 2023, 246, 152036. [Google Scholar] [CrossRef]
- Gao, X.W.; Hu, H.L.; Xie, M.H.; Tang, C.X.; Ou, J.; Lu, Z.H. CX3CL1/CX3CR1 axis alleviates inflammation and apoptosis in human nucleus pulpous cells via M2 macrophage polarization. Exp. Ther. Med. 2023, 26, 359. [Google Scholar] [CrossRef]
- Folkman, J. Angiogenesis. Annu. Rev. Med. 2006, 57, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Akbarian, M.; Bertassoni, L.E.; Tayebi, L. Biological aspects in controlling angiogenesis: Current progress. Cell Mol. Life Sci. 2022, 79, 349. [Google Scholar] [CrossRef]
- Szade, A.; Grochot-Przeczek, A.; Florczyk, U.; Jozkowicz, A.; Dulak, J. Cellular and molecular mechanisms of inflammation-induced angiogenesis. IUBMB Life 2015, 67, 145–159. [Google Scholar] [CrossRef]
- Narkar, V.A. Exercise and Ischemia-Activated Pathways in Limb Muscle Angiogenesis and Vascular Regeneration. Methodist Debakey Cardiovasc. J. 2023, 19, 58–68. [Google Scholar] [CrossRef]
- Yin, C.L.; Ma, Y.J. The Regulatory Mechanism of Hypoxia-inducible Factor 1 and its Clinical Significance. Curr. Mol. Pharmacol. 2024, 17, e18761429266116. [Google Scholar] [CrossRef]
- Zhou, W.; Yang, L.; Nie, L.; Lin, H. Unraveling the molecular mechanisms between inflammation and tumor angiogenesis. Am. J. Cancer Res. 2021, 11, 301–317. [Google Scholar]
- Perry, B.N.; Arbiser, J.L. The duality of angiogenesis: Implications for therapy of human disease. J. Investig. Dermatol. 2006, 126, 2160–2166. [Google Scholar] [CrossRef]
- Edgar, L.T.; Park, H.; Crawshaw, J.R.; Osborne, J.M.; Eichmann, A.; Bernabeu, M.O. Traffic Patterns of the Migrating Endothelium: How Force Transmission Regulates Vascular Malformation and Functional Shunting During Angiogenic Remodelling. Front. Cell Dev. Biol. 2022, 10, 840066. [Google Scholar] [CrossRef] [PubMed]
- Silvestre, J.S.; Lévy, B.I.; Tedgui, A. Mechanisms of angiogenesis and remodelling of the microvasculature. Cardiovasc. Res. 2008, 78, 201–202. [Google Scholar] [CrossRef] [PubMed]
- Niklason, L.; Dai, G. Arterial Venous Differentiation for Vascular Bioengineering. Annu. Rev. Biomed. Eng. 2018, 20, 431–447. [Google Scholar] [CrossRef] [PubMed]
- Sarabipour, S.; Kinghorn, K.; Quigley, K.M.; Kovacs-Kasa, A.; Annex, B.H.; Bautch, V.L.; Mac Gabhann, F. Trafficking dynamics of VEGFR1, VEGFR2, and NRP1 in human endothelial cells. PLoS Comput. Biol. 2024, 20, e1011798. [Google Scholar] [CrossRef]
- Colotti, G.; Failla, C.M.; Lacal, P.M.; Ungarelli, M.; Ruffini, F.; Di Micco, P.; Orecchia, A.; Morea, V. Neuropilin-1 is required for endothelial cell adhesion to soluble vascular endothelial growth factor receptor 1. FEBS J. 2022, 289, 183–198. [Google Scholar] [CrossRef]
- Alghamdi, A.A.A.; Benwell, C.J.; Atkinson, S.J.; Lambert, J.; Johnson, R.T.; Robinson, S.D. NRP2 as an Emerging Angiogenic Player; Promoting Endothelial Cell Adhesion and Migration by Regulating Recycling of α5 Integrin. Front. Cell Dev. Biol. 2020, 8, 395. [Google Scholar] [CrossRef]
- Li, T.; Ran, J.; Miao, Z.; Yang, M.; Mou, D.; Jiang, Y.; Xu, X.; Xie, Q.; Jin, K. Deficiency of inflammation-sensing protein neuropilin-2 in myeloid-derived macrophages exacerbates colitis via NF-κB activation. J. Pathol. 2024, 262, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Rahane, D.; Dhingra, T.; Chalavady, G.; Datta, A.; Ghosh, B.; Rana, N.; Borah, A.; Saraf, S.; Bhattacharya, P. Hypoxia and its effect on the cellular system. Cell Biochem. Funct. 2024, 42, e3940. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Lin, H.; Zhao, Y.H.; Ma, J.X.; Li, J.X. Tube Formation Capability and Chemotaxis of Skin Pericytes. Discov. Med. 2024, 36, 308–322. [Google Scholar] [CrossRef]
- Yang, S.; Graham, J.; Kahn, J.W.; Schwartz, E.A.; Gerritsen, M.E. Functional roles for PECAM-1 (CD31) and VE-cadherin (CD144) in tube assembly and lumen formation in three-dimensional collagen gels. Am. J. Pathol. 1999, 155, 887–895. [Google Scholar] [CrossRef]
- Granger, D.N.; Senchenkova, E. Inflammation and the Microcirculation; Morgan & Claypool Life Sciences: San Rafael, CA, USA, 2010; Chapter 6; Angiogenesis. Available online: https://www.ncbi.nlm.nih.gov/books/NBK53377/ (accessed on 11 March 2024).
- Ouarné, M.; Pena, A.; Franco, C.A. From remodeling to quiescence: The transformation of the vascular network. Cells Dev. 2021, 168, 203735. [Google Scholar] [CrossRef]
- Pollina, E.A.; Legesse-Miller, A.; Haley, E.M.; Goodpaster, T.; Randolph-Habecker, J.; Coller, H.A. Regulating the angiogenic balance in tissues. Cell Cycle 2008, 7, 2056–2070. [Google Scholar] [CrossRef]
- Aguilar-Cazares, D.; Chavez-Dominguez, R.; Carlos-Reyes, A.; Lopez-Camarillo, C.; Hernadez de la Cruz, O.N.; Lopez-Gonzalez, J.S. Contribution of Angiogenesis to Inflammation and Cancer. Front. Oncol. 2019, 9, 1399. [Google Scholar] [CrossRef]
- Britzen-Laurent, N.; Weidinger, C.; Stürzl, M. Contribution of Blood Vessel Activation, Remodeling and Barrier Function to Inflammatory Bowel Diseases. Int. J. Mol. Sci. 2023, 24, 5517. [Google Scholar] [CrossRef]
- Reyes, M.E.; Pulgar, V.; Vivallo, C.; Ili, C.G.; Mora-Lagos, B.; Brebi, P. Epigenetic modulation of cytokine expression in gastric cancer: Influence on angiogenesis, metastasis and chemoresistance. Front. Immunol. 2024, 15, 1347530. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.E.; Lee, S.H.; Kim, W.U. Cytokines, Vascular Endothelial Growth Factors, and PlGF in Autoimmunity: Insights from Rheumatoid Arthritis to Multiple Sclerosis. Immune Netw. 2024, 24, e10. [Google Scholar] [CrossRef] [PubMed]
- Vofo, B.N.; Chowers, I. Suppressing Inflammation for the Treatment of Diabetic Retinopathy and Age-Related Macular Degeneration: Dazdotuftide as a Potential New Multitarget Therapeutic Candidate. Biomedicines 2023, 11, 1562. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Liang, H.; Yuan, P.; Wei, Z.; Zhong, P. Brain pericyte biology: From physiopathological mechanisms to potential therapeutic applications in ischemic stroke. Front. Cell Neurosci. 2023, 17, 1267785. [Google Scholar] [CrossRef] [PubMed]
- Bhavsar, P.K.; Sukkar, M.B.; Khorasani, N.; Lee, K.Y.; Chung, K.F. Glucocorticoid suppression of CX3CL1 (fractalkine) by reduced gene promoter recruitment of NF-kappaB. FASEB J. 2008, 22, 1807–1816. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. Regulation of NF-κB by TNF family cytokines. Semin. Immunol. 2014, 26, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Diep, S.; Maddukuri, M.; Yamauchi, S.; Geshow, G.; Delk, N.A. Interleukin-1 and Nuclear Factor Kappa B Signaling Promote Breast Cancer Progression and Treatment Resistance. Cells 2022, 11, 1673. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Ireland, S.J.; Remington, G.; Alvarez, E.; Racke, M.K.; Greenberg, B.; Frohman, E.M.; Monson, N.L. CD40-Mediated NF-κB Activation in B Cells Is Increased in Multiple Sclerosis and Modulated by Therapeutics. J. Immunol. 2016, 197, 4257–4265. [Google Scholar] [CrossRef]
- Seigner, J.; Basilio, J.; Resch, U.; de Martin, R. CD40L and TNF both activate the classical NF-κB pathway, which is not required for the CD40L induced alternative pathway in endothelial cells. Biochem. Biophys. Res. Commun. 2018, 495, 1389–1394. [Google Scholar] [PubMed]
- Yang, C.; Hwang, H.H.; Jeong, S.; Seo, D.; Jeong, Y.; Lee, D.Y.; Lee, K. Inducing angiogenesis with the controlled release of nitric oxide from biodegradable and biocompatible copolymeric nanoparticles. Int. J. Nanomed. 2018, 13, 6517–6530. [Google Scholar] [CrossRef] [PubMed]
- Heydari, P.; Kharaziha, M.; Varshosaz, J.; Kharazi, A.Z.; Javanmard, S.H. Co-release of nitric oxide and L-arginine from poly (β-amino ester)-based adhesive reprogram macrophages for accelerated wound healing and angiogenesis in vitro and in vivo. Biomater. Adv. 2024, 158, 213762. [Google Scholar] [CrossRef]
- Minet, E.; Arnould, T.; Michel, G.; Roland, I.; Mottet, D.; Raes, M.; Remacle, J.; Michiels, C. ERK activation upon hypoxia: Involvement in HIF-1 activation. FEBS Lett. 2000, 468, 53–58. [Google Scholar] [CrossRef]
- Ryu, J.; Lee, C.W.; Hong, K.H.; Shin, J.A.; Lim, S.H.; Park, C.S.; Shim, J.; Nam, K.B.; Choi, K.J.; Kim, Y.H.; et al. Activation of fractalkine/CX3CR1 by vascular endothelial cells induces angiogenesis through VEGF-A/KDR and reverses hindlimb ischaemia. Cardiovasc. Res. 2008, 78, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Lee, J.; Kwak, J.Y.; Noh, K.; Yim, E.; Kim, H.K.; Kim, Y.J.; Broxmeyer, H.E.; Kim, J.A. Fractalkine induces angiogenic potential in CX3CR1-expressing monocytes. J. Leukoc. Biol. 2018, 103, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.L.; Oubaha, M.; Cagnone, G.; Boscher, C.; Kim, J.S.; El Bakkouri, Y.; Zhang, Y.; Chidiac, R.; Corriveau, J.; Delisle, C.; et al. eNOS controls angiogenic sprouting and retinal neovascularization through the regulation of endothelial cell polarity. Cell Mol. Life Sci. 2021, 79, 37. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhang, Y.; Yin, R.; Zhong, W.; Chen, R.; Yan, J. Activating CD137 Signaling Promotes Sprouting Angiogenesis via Increased VEGFA Secretion and the VEGFR2/Akt/eNOS Pathway. Mediat. Inflamm. 2020, 2020, 1649453. [Google Scholar] [CrossRef] [PubMed]
- Zarychta, E.; Ruszkowska-Ciastek, B. Cooperation between Angiogenesis, Vasculogenesis, Chemotaxis, and Coagulation in Breast Cancer Metastases Development: Pathophysiological Point of View. Biomedicines 2022, 10, 300. [Google Scholar] [CrossRef] [PubMed]
- Janiszewska, M.; Primi, M.C.; Izard, T. Cell adhesion in cancer: Beyond the migration of single cells. J. Biol. Chem. 2020, 295, 2495–2505. [Google Scholar] [CrossRef]
- Folkman, J. Role of angiogenesis in tumor growth and metastasis. Semin. Oncol. 2002, 29 (Suppl. S16), 15–18. [Google Scholar] [CrossRef] [PubMed]
- Nishida, N.; Yano, H.; Nishida, T.; Kamura, T.; Kojiro, M. Angiogenesis in cancer. Vasc. Health Risk Manag. 2006, 2, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Tannock, I.F. The relation between cell proliferation and the vascular system in a transplanted mouse mammary tumour. Br. J. Cancer 1968, 22, 258–273. [Google Scholar] [CrossRef]
- Place, T.L.; Domann, F.E.; Case, A.J. Limitations of oxygen delivery to cells in culture: An underappreciated problem in basic and translational research. Free. Radic. Biol. Med. 2017, 113, 311–322, Erratum in Free. Radic. Biol. Med. 2021, 162, 180. [Google Scholar] [CrossRef]
- Folkman, J. Tumor angiogenesis: Therapeutic implications. N. Engl. J. Med. 1971, 285, 1182–1186. [Google Scholar] [CrossRef] [PubMed]
- Welter, M.; Bartha, K.; Rieger, H. Vascular remodelling of an arterio-venous blood vessel network during solid tumour growth. J. Theor. Biol. 2009, 259, 405–422. [Google Scholar] [CrossRef] [PubMed]
- Gerlee, P.; Anderson, A.R. Diffusion-limited tumour growth: Simulations and analysis. Math. Biosci. Eng. 2010, 7, 385–400. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.D.; Seano, G.; Jain, R.K. Normalizing Function of Tumor Vessels: Progress, Opportunities, and Challenges. Annu. Rev. Physiol. 2019, 81, 505–534. [Google Scholar] [CrossRef] [PubMed]
- Lefler, D.S.; Manobianco, S.A.; Bashir, B. Immunotherapy resistance in solid tumors: Mechanisms and potential solutions. Cancer. Biol. Ther. 2024, 25, 2315655. [Google Scholar] [CrossRef] [PubMed]
- Bielenberg, D.R.; Zetter, B.R. The Contribution of Angiogenesis to the Process of Metastasis. Cancer J. 2015, 21, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Schlößer, H.A.; Theurich, S.; Shimabukuro-Vornhagen, A.; Holtick, U.; Stippel, D.L.; von Bergwelt-Baildon, M. Overcoming tumor-mediated immunosuppression. Immunotherapy 2014, 6, 973–988. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Fu, L.H.; Li, C.; Lin, J.; Huang, P. Conquering the Hypoxia Limitation for Photodynamic Therapy. Adv. Mater. 2021, 33, e2103978. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, O.A.A.; Tesen, H.S.; Hany, M.; Sherif, A.; Abdelwahab, M.M.; Elnaggar, M.H. The role of hypoxia on prostate cancer progression and metastasis. Mol. Biol. Rep. 2023, 50, 3873–3884. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, X.; Zhang, W.; Gao, R.; Wei, H.; Yu, C.Y. Current advances in modulating tumor hypoxia for enhanced therapeutic efficacy. Acta Biomater. 2024, 176, 1–27. [Google Scholar] [CrossRef]
- Ma, S.; Pan, X.; Gan, J.; Guo, X.; He, J.; Hu, H.; Wang, Y.; Ning, S.; Zhi, H. DNA methylation heterogeneity attributable to a complex tumor immune microenvironment prompts prognostic risk in glioma. Epigenetics 2024, 19, 2318506. [Google Scholar] [CrossRef] [PubMed]
- Ge, R.; Wang, Z.; Cheng, L. Tumor microenvironment heterogeneity an important mediator of prostate cancer progression and therapeutic resistance. npj Precis. Oncol. 2022, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Wang, A.; Yuan, Y.; Zhu, B.; Long, H. Heterogeneity of the tumor immune microenvironment and its clinical relevance. Exp. Hematol. Oncol. 2022, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Sidibe, A.; Ropraz, P.; Jemelin, S.; Emre, Y.; Poittevin, M.; Pocard, M.; Bradfield, P.F.; Imhof, B.A. Angiogenic factor-driven inflammation promotes extravasation of human proangiogenic monocytes to tumours. Nat. Commun. 2018, 9, 355. [Google Scholar] [CrossRef] [PubMed]
- Richards, D.M.; Hettinger, J.; Feuerer, M. Monocytes and macrophages in cancer: Development and functions. Cancer Microenviron. 2013, 6, 179–191. [Google Scholar] [CrossRef]
- Li, F.; Wang, Z.; Liu, Y.; Li, J. Down-regulation of fractalkine inhibits the in vitro and in vivo angiogenesis of the hepatocellular carcinoma HepG2 cells. Oncol. Rep. 2010, 24, 669–675. [Google Scholar] [PubMed]
- Zheng, J.; Yang, M.; Shao, J.; Miao, Y.; Han, J.; Du, J. Chemokine receptor CX3CR1 contributes to macrophage survival in tumor metastasis. Mol. Cancer 2013, 12, 141. [Google Scholar] [CrossRef]
- Xu, X.; Wang, Y.; Chen, J.; Ma, H.; Shao, Z.; Chen, H.; Jin, G. High expression of CX3CL1/CX3CR1 axis predicts a poor prognosis of pancreatic ductal adenocarcinoma. J. Gastrointest. Surg. 2012, 16, 1493–1498. [Google Scholar] [CrossRef]
- Ren, T.; Chen, Q.; Tian, Z.; Wei, H. Down-regulation of surface fractalkine by RNA interference in B16 melanoma reduced tumor growth in mice. Biochem. Biophys. Res. Commun. 2007, 364, 978–984. [Google Scholar] [CrossRef]
- Shulby, S.A.; Dolloff, N.G.; Stearns, M.E.; Meucci, O.; Fatatis, A. CX3CR1-fractalkine expression regulates cellular mechanisms involved in adhesion, migration, and survival of human prostate cancer cells. Cancer Res. 2004, 64, 4693–4698. [Google Scholar] [CrossRef]
- Goto, Y.; Aoyama, M.; Sekiya, T.; Kakita, H.; Waguri-Nagaya, Y.; Miyazawa, K.; Asai, K.; Goto, S. CXCR4+ CD45- Cells are Niche Forming for Osteoclastogenesis via the SDF-1, CXCL7, and CX3CL1 Signaling Pathways in Bone Marrow. Stem Cells 2016, 34, 2733–2743. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, W.L.; Shimizu, S.; D’Ambrosio, J.A.; Meucci, O.; Fatatis, A. CX3CR1 is expressed by prostate epithelial cells and androgens regulate the levels of CX3CL1/fractalkine in the bone marrow: Potential role in prostate cancer bone tropism. Cancer Res. 2008, 68, 1715–1722. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Liang, Y.; Jiang, L.; Wang, H.; Wang, S.; Dong, J. CX3CL1/fractalkine enhances prostate cancer spinal metastasis by activating the Src/FAK pathway. Int. J. Oncol. 2018, 53, 1544–1556. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Yi, L.; Liu, P.; Jiang, L.; Wang, H.; Hu, A.; Sun, C.; Dong, J. CX3CL1 involves in breast cancer metastasizing to the spine via the Src/FAK signaling pathway. J. Cancer 2018, 9, 3603–3612. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Liang, Y.; Chan, Q.; Jiang, L.; Dong, J. CX3CL1 promotes lung cancer cell migration and invasion via the Src/focal adhesion kinase signaling pathway. Oncol. Rep. 2019, 41, 1911–1917. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Chen, T.; Wang, B.; Zhang, M.; An, H.; Guo, Z.; Yu, Y.; Qin, Z.; Cao, X. Chemoattraction, adhesion and activation of natural killer cells are involved in the antitumor immune response induced by fractalkine/CX3CL1. Immunol. Lett. 2003, 89, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.K. Inference of gene networks using gene expression data with applications. Heliyon 2024, 10, e26065. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Lim, J.; Wang, X.; Liang, F.; Xiao, G. Enhanced construction of gene regulatory networks using hub gene information. BMC Bioinform. 2017, 18, 186. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, C.; Huang, D.; Ge, C.; Chen, L.; Fu, J.; Du, J. Identification and prognostic analysis of candidate biomarkers for lung metastasis in colorectal cancer. Medicine 2024, 103, e37484. [Google Scholar] [CrossRef]
- Yue, Y.; Zhang, Q.; Sun, Z. CX3CR1 Acts as a Protective Biomarker in the Tumor Microenvironment of Colorectal Cancer. Front. Immunol. 2022, 12, 758040. [Google Scholar] [CrossRef]
- Erreni, M.; Siddiqui, I.; Marelli, G.; Grizzi, F.; Bianchi, P.; Morone, D.; Marchesi, F.; Celesti, G.; Pesce, S.; Doni, A.; et al. The Fractalkine-Receptor Axis Improves Human Colorectal Cancer Prognosis by Limiting Tumor Metastatic Dissemination. J. Immunol. 2016, 196, 902–914. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, T.F.; Kuhn, S.; Stange, C.; Heithorst, N.; Schilling, D.; Jelsma, J.; Sievert, W.; Seitz, S.; Stangl, S.; Hapfelmeier, A.; et al. The Chemokine CX3CL1 Improves Trastuzumab Efficacy in HER2 Low-Expressing Cancer In Vitro and In Vivo. Cancer Immunol. Res. 2021, 9, 779–789. [Google Scholar] [CrossRef]
- Wege, A.K.; Dreyer, T.F.; Teoman, A.; Ortmann, O.; Brockhoff, G.; Bronger, H. CX3CL1 Overexpression Prevents the Formation of Lung Metastases in Trastuzumab-Treated MDA-MB-453-Based Humanized Tumor Mice (HTM). Cancers 2021, 13, 2459. [Google Scholar] [CrossRef]
- Kee, J.Y.; Arita, Y.; Shinohara, K.; Ohashi, Y.; Sakurai, H.; Saiki, I.; Koizumi, K. Antitumor immune activity by chemokine CX3CL1 in an orthotopic implantation of lung cancer model in vivo. Mol. Clin. Oncol. 2013, 1, 35–40. [Google Scholar] [CrossRef]
- Liu, J.; Li, Y.; Zhu, X.; Li, Q.; Liang, X.; Xie, J.; Hu, S.; Peng, W.; Li, C. Increased CX3CL1 mRNA expression level is a positive prognostic factor in patients with lung adenocarcinoma. Oncol. Lett. 2019, 17, 4877–4890. [Google Scholar] [CrossRef]
- Matsubara, T.; Ono, T.; Yamanoi, A.; Tachibana, M.; Nagasue, N. Fractalkine-CX3CR1 axis regulates tumor cell cycle and deteriorates prognosis after radical resection for hepatocellular carcinoma. J. Surg. Oncol. 2007, 95, 241–249. [Google Scholar] [CrossRef]
- Hyakudomi, M.; Matsubara, T.; Hyakudomi, R.; Yamamoto, T.; Kinugasa, S.; Yamanoi, A.; Maruyama, R.; Tanaka, T. Increased expression of fractalkine is correlated with a better prognosis and an increased number of both CD8+ T cells and natural killer cells in gastric adenocarcinoma. Ann. Surg. Oncol. 2008, 15, 1775–1782. [Google Scholar] [CrossRef] [PubMed]
- Korbecki, J.; Simińska, D.; Kojder, K.; Grochans, S.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. Fractalkine/CX3CL1 in Neoplastic Processes. Int. J. Mol. Sci. 2020, 21, 3723. [Google Scholar] [CrossRef]
- Shao, D.; Zhou, H.; Yu, H.; Zhu, X. CX3CR1 is a potential biomarker of immune microenvironment and prognosis in epithelial ovarian cancer. Medicine 2024, 103, e36891. [Google Scholar] [CrossRef]
- Yao, X.; Qi, L.; Chen, X.; Du, J.; Zhang, Z.; Liu, S. Expression of CX3CR1 associates with cellular migration, metastasis, and prognosis in human clear cell renal cell carcinoma. Urol. Oncol. 2014, 32, 162–170. [Google Scholar] [CrossRef]
- Sun, C.; Hu, A.; Wang, S.; Tian, B.; Jiang, L.; Liang, Y.; Wang, H.; Dong, J. ADAM17-regulated CX3CL1 expression produced by bone marrow endothelial cells promotes spinal metastasis from hepatocellular carcinoma. Int. J. Oncol. 2020, 57, 249–263. [Google Scholar] [CrossRef] [PubMed]
- Castellana, D.; Zobairi, F.; Martinez, M.C.; Panaro, M.A.; Mitolo, V.; Freyssinet, J.M.; Kunzelmann, C. Membrane microvesicles as actors in the establishment of a favorable prostatic tumoral niche: A role for activated fibroblasts and CX3CL1-CX3CR1 axis. Cancer Res. 2009, 69, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.M., Jr.; Tharakan, A.; Martin, R.K. Targeting ADAM10 in Cancer and Autoimmunity. Front. Immunol. 2020, 11, 499. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Xuan, Z.; Liu, X.; Zheng, M.; Yang, C.; Wang, H. Immunomodulatory role of metalloproteinase ADAM17 in tumor development. Front. Immunol. 2022, 13, 1059376. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.F.; Ingram, K.; Huang, E.J.; Parksong, J.; McKenney, C.; Bever, G.S.; Regot, S. Systematic analysis of the MAPK signaling network reveals MAP3K-driven control of cell fate. Cell Syst. 2022, 13, 885–894.e4. [Google Scholar] [CrossRef]
- Sufyan, M.; Shokat, Z.; Ashfaq, U.A. Artificial intelligence in cancer diagnosis and therapy: Current status and future perspective. Comput. Biol. Med. 2023, 165, 107356. [Google Scholar] [CrossRef]
- Stafie, C.S.; Sufaru, I.G.; Ghiciuc, C.M.; Stafie, I.I.; Sufaru, E.C.; Solomon, S.M.; Hancianu, M. Exploring the Intersection of Artificial Intelligence and Clinical Healthcare: A Multidisciplinary Review. Diagnostics 2023, 13, 1995. [Google Scholar] [CrossRef]
- Penhaskashi, J.; Sekimoto, O.; Chiappelli, F. Permafrost viremia and immune tweening. Bioinformation 2023, 19, 685–691. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
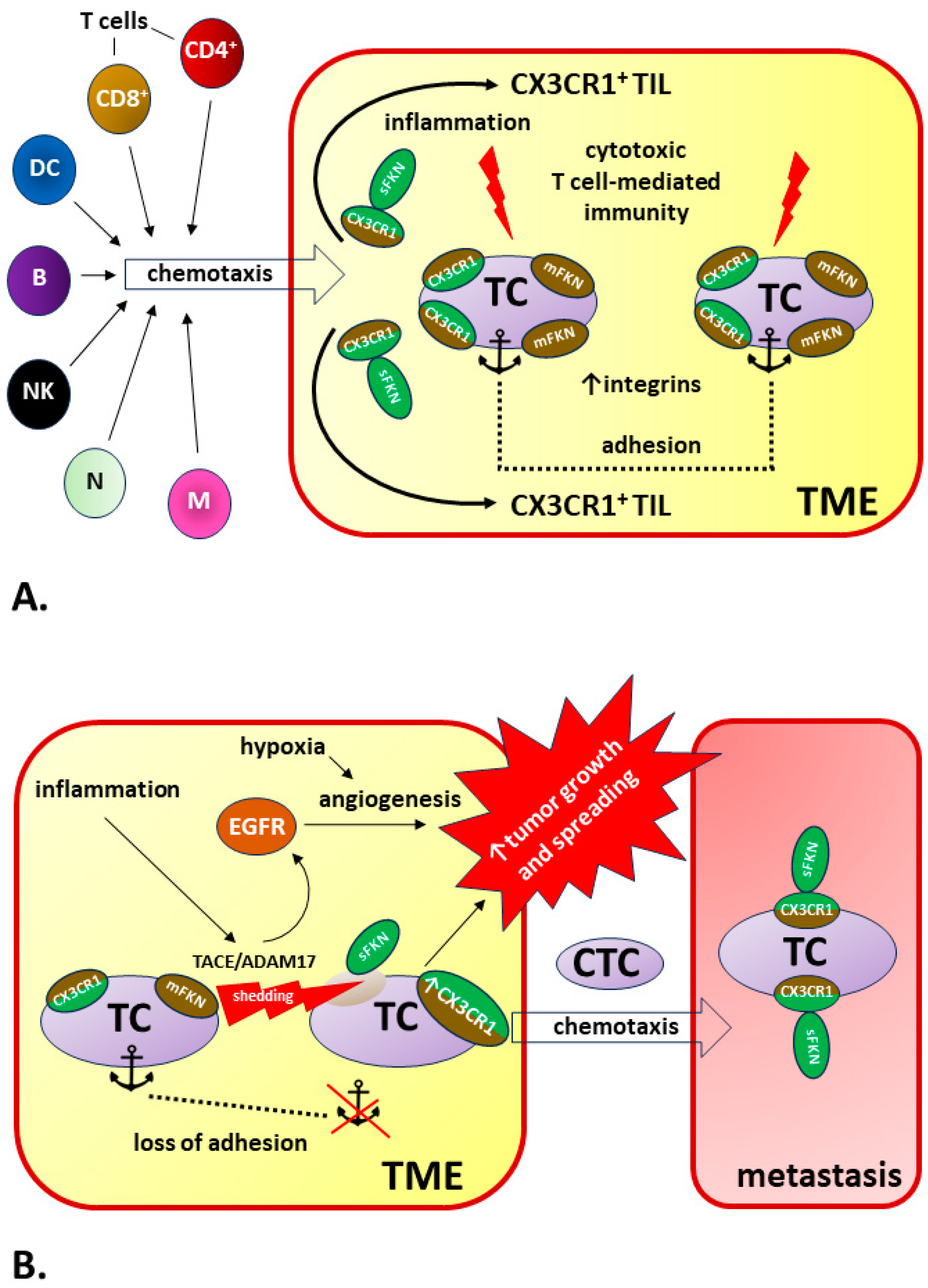{kind=link}
| Location in the Body | Regulation of Biological Processes via FKN/CX3CR1 Axis in Normal and Pathological Conditions | References |
|---|---|---|
| Central nervous system (CNS) | - In brain tissues, FKN is mostly expressed in neurons, while microglia express CX3CR1. - FKN/CX3CR1 signaling enables precise interactions between neurons, microglia, and immune cells. - Due to its key role in microglia–neuron communication, the FKN/CX3CR1 axis regulates a broad spectrum of microglial properties, including microglial cell migration and dynamic surveillance of the brain parenchyma, neuronal integrity and survival, synaptic plasticity, and diverse synaptic functions, as well as neuronal sensitivity to stimuli and excitability via cytokine release modulation, chemotaxis, and phagocytosis. - FKN suppresses lipopolysaccharide (LPS)-induced microglia activation by reducing the production of nitric oxide (NO), interleukin-6 (IL-6), and transforming growth factor alpha (TNF-α), therefore inhibiting neuronal cell death in response to LPS cytotoxicity in the brain tissue. - FKN/CX3CR1 signal disruption is is one of the most important phenomena in the pathomechanisms of CNS-related disorders, especially neurodegenerative diseases and traumatic brain injuries. However, the results of studies on the modulation of inflammation in the CNS by FKN/CX3CR1 are often ambiguous or contradictory. For example, the disruption of FKN signaling is beneficial in limiting the effects of CNS ischemia but detrimental in other neurodegenerative diseases, including Parkinson’s disease (PD) and amyotrophic lateral sclerosis (ALS). Furthermore, the deletion of CX3CR1 in Alzheimer’s disease may, possibly depending on the stage of disease progression, lead to both neuroprotective and detrimental effects. There is also no complete agreement on the importance of the involvement of FKN isoforms in the development of neuropathological processes. - The type of specific response, neurotoxic or neuroprotective, most often depends on the type of damaging factor, the CNS area influencing the regional heterogeneity of microglial cells, and the local concentrations of FKN and CX3CR1. | Sheridan and Murphy 2013 [192] Pawelec et al., 2020 [193] Paolicelli et al., 2014 [194]; Camacho-Hernández and Peña-Ortega 2023 [195] Mizuno et al., 2003 [196]; Lyons et al., 2009 [165]; Mecca et al., 2018 [197] Subbarayan et al., 2022 [198]; Cipriani et al., 2011 [199]; Poniatowski et al., 2017 [61]; Luo et al., 2019 [200]; Bivona et al., 2023 [201]; Nash et al., 2015 [202]; Juliani et al., 2021 [203]; Lee et al., 2010 [204], Cho et al., 2011 [205]; Fuhrmann et al., 2010 [206]; Pawelec et al., 2020 [193]; Eugenín et al., 2023 [207] Sheridan and Murphy 2013 [192]; Stratoulias et al. [208] |
| Bone marrow and immune tissue | - The expression of CX3XR1 increases with the maturation of myeloid cells and shows an inverse correlation with the Ly6C marker and the C-C chemokine receptor 2 (CCR2) in the blood. This may indicate that CX3CR1 limits the motility of Ly6C (high) monocytes within the bone marrow and, thus, controls their release into the blood. - FKN-CX3CL1 axis plays a significant role in an early stage of osteoblast differentiation, possibly through their trans and cis interactions. - FKN regulates mouse osteoclast precursor (OCP) survival and primes OCPs for subsequent osteoclast differentiation. - Autoimmune and inflammatory responses in rheumatoid arthritis (RA) are positively correlated with the concentration of FKN in the serum and synovial fluid. The associated chemotaxis primarily involves the recruitment of CD16+ monocytes into synovial tissues, as CX3CR1 expression in CD16+ monocytes is markedly higher compared to other populations (e.g., CD14+ and CD167−). - Bone marrow (BM) FKN levels are significantly increased in the multiple myeloma (MM) patients and positively correlated with BM microvessel density. - CX3CR1 expression is an additional marker of natural killer (NK) cell differentiation and closely related to their ability to migrate to the central nervous system (CNS) from the periphery. - CX3CR1 is prevalently expressed on killer cell lectin-like receptor subfamily G member 1 (KLRG1)+ NK cells, a subset that is considered terminally differentiated. Therefore, CX3CR1 may represent a marker of a KLRG1(+) NK-cell subset with its own unique properties that can unidirectionally and irreversibly differentiate from the KLRG1(+)/CX3CR1(−) NK cells during a functionally stable period of stay in the bone marrow. - FKN activates the Jak2-Stat5α-ERK1/2 pathway via CX3CR1, thereby triggering integrin-dependent mechanisms of cytoskeleton remodeling to allow chemotactic relocations of bone marrow-derived mesenchymal stem cells (BMSCs) toward an ischemic cerebral lesion. | Jacquelin et al., 2013 [209] Hoshino et al., 2013 [210] Kuboi et al., 2022 [211] Yano et al., 2007 [212] Marchica et al., 2019 [213] Hamman et al., 2011 [94] Sciumè et al., 2011 [214] Zhang et al., 2015 [215] |
| Cardiovascular system | - FKN and CX3CR1 are expressed in atherosclerotic lesions, and FKN is involved in the initiation step of atherosclerotic plaque formation. Monocyte–endothelial cell interactions are partly mediated by the expression of the monocyte CX3CR1 and endothelial cell FKN. The activation of these lymphocytes upon ligand/receptor binding leads to the release of lysis granules that destroy the vascular endothelium. - After endothelial damage, the release of FKN from apoptotic cells results in subsequent recruitment of macrophages, which promotes the removal of apoptotic debris; however, in more advanced stages of atherosclerosis, signaling through the FKN/CX3CR1 axis enhances foam cell formation, promoting the development of atherosclerotic plaques. - CX3CR1 expression on vascular smooth muscle cells (VSMCs) within atherosclerotic plaque causes the functional state of the FKN/CX3CR1 axis to play an important role in plaque stability. Emergency conditions associated with cardiovascular mortality and morbidity are typically caused by the rupture of “vulnerable” atherosclerotic lesions. - FKN promotes the development of atherosclerotic lesions by activating platelets and causing their adhesion to the endothelium. - The early activation of the cardiac FKN/CX3CR1 axis delays and limits β-adrenergic-induced heart failure. - FKN levels are markedly elevated during acute myocardial infarction (AMI) compared to patients with stable angina pectoris (AP), although they do not correlate with infarct size. The inverse pattern in gene expression of CX3CR1 might be here a kind of compensatory mechanism. - In addition to demonstrating a positive correlation of FKN concentration with an increased risk of developing poorer cardiac function after AMI, levels of FKN can also be treated as a prognostic for the risk of developing major adverse cardiovascular events (MACEs) in acute ST-elevation myocardial infarction (STEMI) patients. - The inhibition of the FKN/CX3CR1 interaction has a beneficial effect on the final infarct size after reperfusion, as it reduces the severity of an important complication—ischemia/reperfusion injury. This complication is directly related to the action of CX3CR1+ lymphocytes toward microvascular obstruction (MVO). - Increased risk of deep vein thrombosis (DVT) is positively correlated with increased activity of the FKN/CX3CR1 axis that involves CX3CR1-expressing platelets, then binding to monocytes and CD8+ lymphocytes. - In metabolic syndrome, platelet activation occurs and the percentage of platelet–eosinophil and platelet–lymphocyte aggregates increases, which is accompanied by the upregulation of platelet CX3CR1 expression. FKN-dependent increased adhesion of these aggregates may play a key role in atherogenesis. | Teupser et al., 2004 [216]; Ma et al., 2022 [217]; Riopel et al., 2019 [218]; Liu and Jiang 2011 [219] Elliott et al., 2017 [220]; White et al., 2014 [190]; Landsman et al., 2009 [171] Lucas et al., 2003 [221]; Harman and Jørgensen 2019 [222]; Apostolakis and Spandidos 2013 [223]; Skoda et al., 2018 [224] Noels et al., 2019 [225]; Flierl et al., 2015 [80] Flamant et al., 2021 [226] Njerve et al., 2014 [227]; Yao et al., 2015 [228] Yao et al., 2015 [228]; Xu et al., 2019 [229] Loh et al., 2023 [44]; Boag et al., 2015 [230] Furio et al., 2018 [231] Marques et al., 2019 [232] |
| Respiratory system | - CX3CR1+ leukocyte attachment to the lung vascular endothelium and diapedesis through the glycocalyx, endothelial cell layer and the basement membrane lead to mononuclear cell accumulation in the lung vessel walls and parenchyma. Infiltrated CX3CR1+ immune cells are a source of mediators that induce damage, stimulate proliferation, and/or affect chemoattract inflammatory cells. The result of these cumulative actions is a structural destruction and remodeling in the development of inflammatory lung diseases. - FKN/CX3CR1 signaling may be involved in the pathophysiology of hypoxia-induced pulmonary arterial hypertension (PAH) developing due to chronic inflammation. Both increased FKN concentrations and upregulated CX3CR1 expression cause PAH progression with vascular remodeling and proliferation of pulmonary artery smooth muscle cells. - Soluble FKN chemoattracts and activates CX3CR1+ leukocytes such as CD8+, CD4+, and γδ T lymphocytes; natural killer cells; dendritic cells; and monocytes/macrophages, leading to mononuclear cell influx and accumulation in the lung vessel walls and parenchyma. During the resolution phase of acute lung injury, apoptotic cell-derived CX3CL1 attracts alveolar macrophages transmigration toward apoptotic cells for phagocytosis. - In allergic asthma, CX3CR1 signaling is essential for airway inflammation by promoting T helper cell survival and maintenance in inflamed lung together with chemotaxis recruited mast cells into bronchial mucosa. - FKN is elevated in both bronchoalveolar lavage fluid and sputum from human asthmatics sensitized to fungi, implicating an association with FKN in fungal asthma severity. However, FKN/CX3CR1 axis preserves lung function during fungal-associated allergic airway inflammation through a nonclassical immunoregulatory mechanism. Hence, the knockout of CX3CR1 signaling resulted in a profound impairment in lung function during fungal-associated allergic airway inflammation. - In pulmonary infections, the role of FKN/CX3CR1 axis remains unclear. For example, FKN may be involved in both immunopathological and anti-viral immune responses to rhinovirus infection. | Zhang and Patel 2010 [233] Balabanian et al., 2002 [234]; Amsellem et al., 2017 [235] Tsai et al., 2021 [236] Mionnet et al., 2010 [95]; El-Shazly et al., 2006 [237] Godwin et al., 2021 [238] Upton et al., 2017 [239] |
| Liver | - FKN/CX3CR1 is upregulated during liver damage including chronic inflammatory liver diseases such as chronic hepatitis C, nonalcoholic steatohepatitis (NASH)/nonalcoholic fatty liver disease (NAFLD), and cirrhosis. - The assessment of the impact of increased FKN/CX3CR1 activity on the severity of steatosis, inflammation, and liver fibrosis is still ambiguous. In addition to reports indicating that FKN-CX3CR1 interaction limits inflammatory properties in Kupffer cells/macrophages, resulting in a reduction in liver inflammation intensity and decreased fibrosis, there are also contradictory research data. - FKN/CX3CR1 upregulation was reported in injured bile ducts of primary cirrhosis with its involvement in the recruitment of intraepithelial lymphocytes of intrahepatic bile ducts. Moreover, the correlation between primary biliary cirrhosis and FKN expression is significantly proportional. | Efsen et al., 2002 [240]; Bourd-Boittin et al., 2009 [63]; Sutti et al., 2015 [241]; Nagata et al., 2022 [242] Aoyama et al., 2010 [243]; Zhang et al., 2020 [244]; Ni et al., 2022 [75]; Sutti et al., 2015 [241]; Karlmark et al., 2010 [245]; Wasmuth et al., 2008 [246]; Hassan et al., 2023 [247] Isse et al., 2005 [248]; Shimoda et al., 2010 [249] |
| Gut | - Most macrophages and some dendritic cell (DC) subsets express CX3CR1 in the gut. In resting intestinal mucosa, the role of lamina propria CX3CR1+ macrophage is to pass captured antigen via trans-epithelial dendrites or phagocytosis onto DC for transport to the mesenteric lymph node (MLN) to prime immune responses like lamina propria DC. - The deletion of FKN or CX3CR1 leads to a specific and significant reduction in lamina propria macrophages with reductions in the translocation of bacteria to MLNs and their ability to take up pathogens. Therefore, CX3CR1 may be treated as a specific marker useful for lamina propria macrophages and a key component in sustainment lamina propria macrophage homeostasis. Contradictory, it was demonstrated that CX3CR1 knockout mice show normal numbers of macrophages. - The intestinal microbiome influences the local accumulation of CX3CR1+ phagocytes, and the number of CX3CR1+ cells is reduced in germ-free mouse. - The enhanced recruitment of CX3CR1+ T cells by mucosal human intestinal microvascular endothelial cell (HIMECs)-derived FKN has been demonstrated in inflammatory bowel disease (IBD). | Joeris et al., 2017 [250]; Niess et al., 2005 [118]; Bain and Mowat 2011 [251]; Lee et al., 2018 [74] Ferretti et al., 2014 [252]; Bain et al., 2013 [253] Bain et al., 2014 [254] Sans et al., 2007 [255] |
| Placenta | - Human placenta is a source of FKN, which is expressed in the syncytiotrophoblast and can be released into the maternal vascular compartment (maternal circulation) by constitutive MMP-dependent shedding. - FKN content within the apical microvillous plasma membrane increases significantly in the placenta of full-term pregnancy compared to the first trimester. - FKN/CX3CR1 axis mediates the adhesion of monocytes to the villous trophoblast. - Increased expression and release of placental FKN may be responsible for low-grade systemic inflammatory background and responses in the third trimester of a normal pregnancy. - Placental FKN is upregulated in severe early-onset pre-eclampsia (PE). Significant underdevelopment of placental vascular network with a significantly lowered vascular/extravascular tissue index (V/EVTI) in PE is associated with the dysregulation of the FKN/CX3CR1 system, especially in fetal growth restriction (FGR)-complicated pregnancies. - Increased average FKN content in the diabetic placenta is accompanied by an increase in the density of placental microvessels and a higher expression of CX3CR1 compared to the placenta from a normal pregnancy. Therefore, FKN/CX3CR1 signaling pathway is involved in the pathomechanism of placental microvasculature remodeling during diabetes class C (after White). - Placental hypoxia increases FKN production and upregulates CX3CR1 expression in the placental endothelial cells. Under these conditions, tumor necrosis factor alpha (TNFα) induces FKN, influencing a mechanism of FKN autoregulation via CX3CR1 expression. - Increased FKN concentration, accompanied by a higher mean FKN gene expression level in the tissues of pregnant women with missed abortion, may be responsible for abnormal placental invasion. | Siwetz et al., 2014 [256] Siwetz et al., 2015 [257] Siwetz et al., 2015 [258] Vishnyakova et al., 2021 [259] Szewczyk et al., 2021 [260]; Ullah et al., 2023 [261] Szukiewicz et al., 2013 [262]; Ullah et al., 2023 [261]; Szukiewicz et al., 2014 [263] Gokce et al., 2022 [264] |
| Joint and bone tissue | - The total number of circulating CX3CR1high T cells is increased in the circulation of rheumatoid arthritis (RA) patients. Joint-infiltrated CX3CR1high T cells tightly and strongly adhere to fibroblast-like synoviocytes (FLSs) in the synovium in an FKN-dependent manner. - The FKN/CX3CR1 axis promotes inflammation-free osteoclastogenesis by enhancing precursor cell survival and differentiation. - The apoptosis of chondrocytes during joint osteoarthritis upregulates the FKN-CX3CR1-p38 axis, which results in the enhanced chemotaxis of osteoclast precursors (OCPs) and promotes bone resorption. - The development of osteoarthritis (OA) is largely driven by low-grade local background inflammation based on FKN-mediated chemotaxis. - FKN/CX3CR1 signaling in hemophilia is involved in the pathomechanism of irreversible joint degeneration (hemophilic arthropathy). - Significantly increased concentrations of FKN in human blood serum are accompanied by high concentrations of bone turnover and inflammatory factors in the serum, such as tartrate-resistant acid phosphatase 5b (TRACP-5b), cross-linked N-telopeptides of type I collagen (NTx), and interleukins (IL-1β, IL-6). - FKN knockdown ameliorates inflammation and apoptosis after exposure to LPS and accelerates osteogenic differentiation. These effects related to FKN deficiency can be reversed by increased expression of CX3CR1. - FKN axis signaling alleviates intervertebral disc degeneration (IDD) by reducing inflammation and apoptosis of human nucleus pulposus cells (HNPCs) via macrophages. | Tanaka et al., 2020 [265] Kuboi et al., 2022 [211]; Koizumi et al., 2009 [266] Koizumi et al., 2009 [266]; Guo et al., 2022 [267] Wojdasiewicz et al., 2014 [43,268] Wojdasiewicz et al., 2020 [188] Wojdasiewicz et al., 2019 [164] Lu et al., 2023 [269] Gao et al., 2023 [270] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szukiewicz, D. CX3CL1 (Fractalkine)-CX3CR1 Axis in Inflammation-Induced Angiogenesis and Tumorigenesis. Int. J. Mol. Sci. 2024, 25, 4679. https://doi.org/10.3390/ijms25094679
Szukiewicz D. CX3CL1 (Fractalkine)-CX3CR1 Axis in Inflammation-Induced Angiogenesis and Tumorigenesis. International Journal of Molecular Sciences. 2024; 25(9):4679. https://doi.org/10.3390/ijms25094679
Chicago/Turabian StyleSzukiewicz, Dariusz. 2024. "CX3CL1 (Fractalkine)-CX3CR1 Axis in Inflammation-Induced Angiogenesis and Tumorigenesis" International Journal of Molecular Sciences 25, no. 9: 4679. https://doi.org/10.3390/ijms25094679
APA StyleSzukiewicz, D. (2024). CX3CL1 (Fractalkine)-CX3CR1 Axis in Inflammation-Induced Angiogenesis and Tumorigenesis. International Journal of Molecular Sciences, 25(9), 4679. https://doi.org/10.3390/ijms25094679