
Anatomical and Metabolome Features of Haloxylon aphyllum and Haloxylon persicum Elucidate the Resilience against Gall-Forming Insects

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Anatomical Structure of Intact and Damaged H. aphyllum and H. persicum
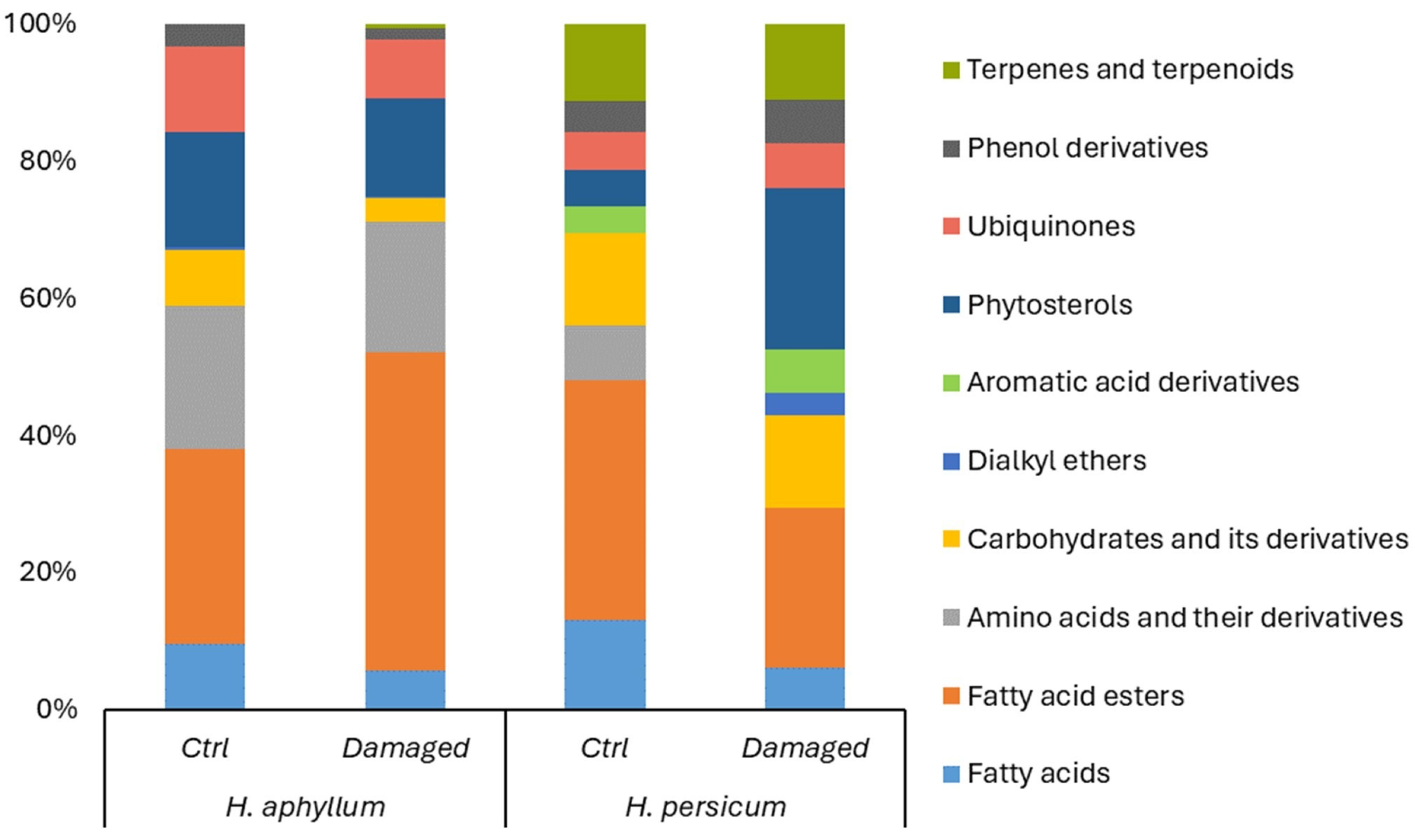
2.2. Metabolomic Structure of Intact and Damaged H. aphyllum and H. persicum
3. Discussion
3.1. Alterations in the Anatomical Structure of Intact and Damaged H. aphyllum and H. persicum
3.2. Alterations in the Metabolomic Structure of Intact and Damaged H. aphyllum and H. persicum
3.2.1. Fatty Acids and Fatty Acid Esters
3.2.2. Dialkyl Ethers
3.2.3. Amino Acids and Their Derivatives
3.2.4. Carbohydrates and Their Derivatives
3.2.5. Aromatic Acid Derivatives
3.2.6. Phytosterols
3.2.7. γ-Tocopherol (Vitamin E)
3.2.8. Phenols
3.2.9. Terpenoids
4. Materials and Methods
4.1. Research Objects and Plant Collection Sites
4.2. Anatomical Structure Analysis
4.3. Metabolomic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mukanov, B.M.; Uteshkaliev, M.D.; Mambetov, B.T. Sandy Deserts of Kazakhstan and Their Forest Reclamation Development; Publishing House “World of Printing”: Kokshetau, Kazakhstan, 2017; p. 548. [Google Scholar]
- Vibe, Y.P.; Fedotova, Z.A.; Krekova, Y.A. Saxaul Plantations of the Kyzylkum: Methodology, Gall Formers and Issues of Selection Assessment. In Practical Guide; Print Centr: Kostanaj, Kazakhstan, 2022; p. 108. [Google Scholar]
- Ministry of Ecology, Geology and Natural Resources of the Republic of Kazakhstan, Committee of Forestry and Wildlife, Republican State Treasury Enterprise “Kazakh Forest Management Enterprise”. Explanatory Note to the Materials of the State Forest Cadaster and the Cadaster of Specially Protected Forest Areas of the Forest Fund of the Republic of Kazakhstan as of January 1, 2023; Almaty, Kazakhstan, 2023; p. 133. Available online: https://www.gov.kz/memleket/entities/ecogeo/documents/details/482988?lang=ru&ysclid=lvgchtsxvi136395407 (accessed on 9 April 2024).
- Fan, L.; Wang, G.; Hu, W.; Pantha, P.; Tran, K.-N.; Zhang, H.; An, L.; Dassanayake, M.; Qiu, Q.-S. Transcriptomic View of Survival during Early Seedling Growth of the Extremophyte Haloxylon ammodendron. Plant Physiol. Biochem. 2018, 132, 475–489. [Google Scholar] [CrossRef] [PubMed]
- Purayil, F.T.; Rajashekar, B.; Kurup, S.S.; Cheruth, A.J.; Subramaniam, S.; Tawfik, N.H.; Amiri, K.M. Transcriptome Profiling of Haloxylon persicum (Bunge Ex Boiss and Buhse) an Endangered Plant Species under PEG-Induced Drought Stress. Genes 2020, 11, 640. [Google Scholar] [CrossRef] [PubMed]
- Shuyskaya, E.V.; Li, E.V.; Rahmankulova, Z.F.; Kuznetsova, N.A.; Toderich, K.N.; Voronin, P.Y. Morphophysiological Adaptation Aspects of Different Haloxylon aphyllum (Chenopodiaceae) Genotypes along a Salinity Gradient. Russ. J. Ecol. 2014, 45, 181–187. [Google Scholar] [CrossRef]
- Arabzadeh, N. The Effect of Drought Stress on Soluble Carbohydrates (Sugars) in Two Species of Haloxylon persicum and Haloxylon aphyllum. Asian J. Plant Sci. 2012, 11, 44–51. [Google Scholar] [CrossRef]
- Suo, Z.; Jia, Z.; Lu, Q.; Pan, B.; Jin, X.; Xu, G.; Peng, X.; Sun, H.; Tao, Y. Distinguishing Haloxylon Persicum and H. ammodendron (Haloxylon Bunge, Amaranthaceae) Using DNA Marker. AASRI Procedia 2012, 1, 305–310. [Google Scholar] [CrossRef]
- Buras, A.; Wucherer, W.; Zerbe, S.; Noviskiy, Z.; Muchitdinov, N.; Shimshikov, B.; Zverev, N.; Schmidt, S.; Wilmking, M.; Thevs, N. Allometric Variability of Haloxylon Species in Central Asia. For. Ecol. Manag. 2012, 274, 1–9. [Google Scholar] [CrossRef]
- Krekova, Y.A.; Kerdyashkin, A.V.; Vibe, E.P. Ecological and phytocoenotic assessment of the state of saxaul forests of Northern Kyzylkums. Environ. Eng. 2024, 1, 130–136. [Google Scholar] [CrossRef]
- Kurochkina, L.Y. Vegetation of the Sandy Deserts of Kazakhstan. Vegetation cover of Kazakhstan; Science: Alma-Ata, Kazakhstan, 1966; Volume 1, pp. 191–582. [Google Scholar]
- Kurochkina, L.Y. Psammophilous Vegetation of the Deserts of Kazakhstan; Science: Alma-Ata, Kazakhstan, 1978. [Google Scholar]
- Mamaev, B.M. Gall-Forming Insects of Saxaul Forests. In Insects as Components of the Biogeocenosis of Saxaul Forest; Science: Moscow, Russia, 1975; pp. 61–82. [Google Scholar]
- Kaplin, V.G. Complexes of Arthropod Animals Living in Plant Tissues of Sandy Deserts; Ylym: Ashgabat, Turkmenistan, 1981; p. 376. [Google Scholar]
- Fedotova, Z.A. Phytophagous Gall Midges (Diptera, Cecidomyiidae) of the Deserts and Mountains of Kazakhstan; Samara State Agricultural Academy: Samara, Russia, 2000; p. 804. [Google Scholar]
- Li, F.-L.; Li, T.; Su, J.; Yang, S.; Wang, P.-L.; Zhang, J.-P. Temporal and Spatial Differences in Gall Induction on Haloxylon by Aceria haloxylonis (Acari: Eriophyidae) in the Gurbantünggüt Desert. Syst. Appl. Acarol. 2016, 21, 1670–1680. [Google Scholar] [CrossRef]
- Skuhrava, M.; Karimpour, Y.; Sadeghi, H.; Gol, A.; Joghataie, M. Gall Midges (Diptera: Cecidomyiidae) of Iran-Annotated List and Zoogeographical Analysis. Acta Soc. Zool. Bohemicae 2014, 78, 269–301. [Google Scholar]
- Saji, A.; Sakkir, S.; Al Dhaheri, S.S. Galling Insects Associated with Haloxylon spp. (Bunge) in Abu Dhabi, United Arab Emirates. Int. J. 2013, 5, 289–295. [Google Scholar] [CrossRef]
- Slepyan, E. Specific Characteristics of Gall-Forming and Teratogenic Processes on the Assimilational Shoots of Haloxylon aphyllum and H. persicum Chenopodiaceae Referat. Bot Zhur 1958, 43, 1595–1607. [Google Scholar]
- Mohammadi, M. Phenological Relation of the Pests Associated with Saxaul Plant in Abardeg Area. Iran. J. Range Desert Res. 2003, 10, 17–38. [Google Scholar]
- Xue, X.-F.; Zhang, J.-P.; Li, F.-L.; Hong, X.-Y. A New Eriophyoid Mite Species (Acari: Eriophyidae) Infesting Haloxylon ammodendron and H. persicum (Chenopodiaceae) in Xinjiang Uigur Autonomous Region, Northwest China. Syst. Appl. Acarol. 2012, 17, 202–209. [Google Scholar] [CrossRef]
- Li, F.; Wu, X.; Wang, P.; Liang, H.; Wang, X.; Wang, L.; Zhang, J. The Occurrence Regularity of Psyllid in Haloxylon spp. and Its Influencing Factors. Shengtai XuebaoActa Ecol. Sin. 2012, 32, 2311–2319. [Google Scholar]
- Sinadsky, Y.V. Dendrophilous Insects of the Deserts of Central Asia and Kazakhstan and Measures to Combat Them; Science: Moscow, Russia, 1968; p. 127. [Google Scholar]
- Loginova, M. Review of the Genus Caillardia Bergevin (Homoptera, Aphalaridae) with Descriptions of New Species. Tr. Zool. Instituta 1978, 71, 6–22. [Google Scholar]
- Loginova, M.M.; Parfentiev, V.J. Species of the Genus Caillardia Bergevin (Homoptera, Psyllidae) Injurious to Haloxylon. Entomol Obozr 1956, 35, 377–396. [Google Scholar]
- Zhao, Q.; Jiang, L.-L.; Guo, J.; Zhang, D.-K.; Hu, H.-Y. Differences in Gall Induction of Flower-like Galls on Haloxylon by Psyllids (Hemiptera: Aphalaridae), and the Emergence of Corresponding Parasitoids. Insects 2021, 12, 861. [Google Scholar] [CrossRef]
- Nurmuratov, T. Insects and Rodents Inhabited in the Deserts of South-Eastern Kazakhstan; Publishing House «Konzhyk»: Almaty, Kazakhstan, 1998; p. 288. [Google Scholar]
- Liu, X.; Ma, D.; Zhang, Z.; Wang, S.; Du, S.; Deng, X.; Yin, L. Plant Lipid Remodeling in Response to Abiotic Stresses. Environ. Exp. Bot. 2019, 165, 174–184. [Google Scholar] [CrossRef]
- Toderich, K.N.; Terletskaya, N.V.; Zorbekova, A.N.; Saidova, L.T.; Ashimuly, K.; Mamirova, A.; Shuyskaya, E.V. Abiotic Stresses Utilisation for Altering the Natural Antioxidant Biosynthesis in Chenopodium quinoa L. Russ. J. Plant Physiol. 2023, 70, 155. [Google Scholar] [CrossRef]
- Terletskaya, N.V.; Korbozova, N.K.; Grazhdannikov, A.E.; Seitimova, G.A.; Meduntseva, N.D.; Kudrina, N.O. Accumulation of Secondary Metabolites of Rhodiola semenovii Boriss. In Situ in the Dynamics of Growth and Development. Metabolites 2022, 12, 622. [Google Scholar] [CrossRef]
- Gätjens-Boniche, O. The Mechanism of Plant Gall Induction by Insects: Revealing Clues, Facts, and Consequences in a Cross-Kingdom Complex Interaction. Rev. Biol. Trop. 2019, 67, 1359–1382. [Google Scholar] [CrossRef]
- Peters, K.; Worrich, A.; Weinhold, A.; Alka, O.; Balcke, G.; Birkemeyer, C.; Bruelheide, H.; Calf, O.W.; Dietz, S.; Dührkop, K.; et al. Current Challenges in Plant Eco-Metabolomics. Int. J. Mol. Sci. 2018, 19, 1385. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Parida, A.K. Metabolomics and Network Analysis Reveal the Potential Metabolites and Biological Pathways Involved in Salinity Tolerance of the Halophyte Salvadora persica. Environ. Exp. Bot. 2018, 148, 85–99. [Google Scholar] [CrossRef]
- Rodríguez-Calcerrada, J.; Rodrigues, A.M.; Perdiguero, P.; António, C.; Atkin, O.K.; Li, M.; Collada, C.; Gil, L. A Molecular Approach to Drought-Induced Reduction in Leaf CO2 Exchange in Drought-Resistant Quercus ilex. Physiol. Plant. 2018, 162, 394–408. [Google Scholar] [CrossRef]
- Schiffmann, C.; Hansen, R.; Baumann, S.; Kublik, A.; Nielsen, P.H.; Adrian, L.; von Bergen, M.; Jehmlich, N.; Seifert, J. Comparison of Targeted Peptide Quantification Assays for Reductive Dehalogenases by Selective Reaction Monitoring (SRM) and Precursor Reaction Monitoring (PRM). Anal. Bioanal. Chem. 2014, 406, 283–291. [Google Scholar] [CrossRef]
- Shih, M.-L.; Morgan, J.A. Metabolic Flux Analysis of Secondary Metabolism in Plants. Metab. Eng. Commun. 2020, 10, e00123. [Google Scholar] [CrossRef]
- Alghanem, S. Ecological and Botanical Diversity in Haloxylon Persicum Community at Al-Qassim Region in Kingdom of Saudi Arabia. Am. J. Environ. Prot. 2018, 6, 43–49. [Google Scholar] [CrossRef]
- Zhaglovskaya, A.; Aidosova, S.; Akhtayeva, N.; Mamurova, A.; Yesimova, D. Anatomical and Morphological Stem Features of Two Haloxylon Species (Chenopodiaceae Vent.) of Drought Stress, Kazakhstan. Biosci. Biotechnol. Res. Asia 2015, 12, 1965–1974. [Google Scholar] [CrossRef]
- Wang, T.-T.; Chu, G.-M.; Jiang, P.; Niu, P.-X.; Wang, M. Effects of Sand Burial and Seed Size on Seed Germination, Seedling Emergence and Seedling Biomass of Anabasis aphylla. Pak. J. Bot. 2017, 49, 391–396. [Google Scholar]
- Xu, G.; Yu, D.; Xie, J.; Tang, L.; Li, Y. What Makes Haloxylon persicum Grow on Sand Dunes While H. ammodendron Grows on Interdune Lowlands: A Proof from Reciprocal Transplant Experiments. J. Arid Land 2014, 6, 581–591. [Google Scholar] [CrossRef]
- Terletskaya, N.V.; Erbay, M.; Zorbekova, A.N.; Prokofieva, M.Y.; Saidova, L.T.; Mamirova, A. Influence of Osmotic, Salt, and Combined Stress on Morphophysiological Parameters of Chenopodium quinoa Photosynthetic Organs. Agriculture 2022, 13, 1. [Google Scholar] [CrossRef]
- Ruan, X.; Wang, Q.; Xu, N.; Li, J.; Huang, J. Physio-Ecological Response of Haloxylon persicum Photosynthetic Shoots to Drought Stress. Sci. Silvae Sin. 2005, 41, 28–32. [Google Scholar] [CrossRef]
- Anvari, S.M.; Mehdikhani, H.; Shahriari, A.; Nouri, G. Effect of Salinity Stress on 7 Species of Range Plants in Germination Stage. Iran. J. Range Desert Res. 2009, 16, 262–273. [Google Scholar]
- Guo, R.; Shi, L.; Jiao, Y.; Li, M.; Zhong, X.; Gu, F.; Liu, Q.; Xia, X.; Li, H. Metabolic Responses to Drought Stress in the Tissues of Drought-Tolerant and Drought-Sensitive Wheat Genotype Seedlings. AoB Plants 2018, 10, ply016. [Google Scholar] [CrossRef]
- Fàbregas, N.; Fernie, A.R. The Metabolic Response to Drought. J. Exp. Bot. 2019, 70, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Wang, Y.; Sun, H. Targeted and Untargeted Metabolomics Reveals Deep Analysis of Drought Stress Responses in Needles and Roots of Pinus taeda Seedlings. Front. Plant Sci. 2023, 13, 1031466. [Google Scholar] [CrossRef]
- Pires, M.V.; Pereira Júnior, A.A.; Medeiros, D.B.; Daloso, D.M.; Pham, P.A.; Barros, K.A.; Engqvist, M.K.M.; Florian, A.; Krahnert, I.; Maurino, V.G.; et al. The Influence of Alternative Pathways of Respiration That Utilize Branched-Chain Amino Acids Following Water Shortage in Arabidopsis. Plant Cell Environ. 2016, 39, 1304–1319. [Google Scholar] [CrossRef]
- Xu, C.; Wei, L.; Huang, S.; Yang, C.; Wang, Y.; Yuan, H.; Xu, Q.; Zhang, W.; Wang, M.; Zeng, X.; et al. Drought Resistance in Qingke Involves a Reprogramming of the Phenylpropanoid Pathway and UDP-Glucosyltransferase Regulation of Abiotic Stress Tolerance Targeting Flavonoid Biosynthesis. J. Agric. Food Chem. 2021, 69, 3992–4005. [Google Scholar] [CrossRef] [PubMed]
- Mescher, M.C.; De Moraes, C.M. Role of Plant Sensory Perception in Plant–Animal Interactions. J. Exp. Bot. 2015, 66, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Lv, G. Metabolomic Analysis of the Response of Haloxylon ammodendron and Haloxylon persicum to Drought. Int. J. Mol. Sci. 2023, 24, 9099. [Google Scholar] [CrossRef]
- Martinière, A.; Gayral, P.; Hawes, C.; Runions, J. Building Bridges: Formin1 of Arabidopsis Forms a Connection between the Cell Wall and the Actin Cytoskeleton. Plant J. 2011, 66, 354–365. [Google Scholar] [CrossRef] [PubMed]
- He, M.; He, C.-Q.; Ding, N.-Z. Abiotic Stresses: General Defenses of Land Plants and Chances for Engineering Multistress Tolerance. Front. Plant Sci. 2018, 9, 1771. [Google Scholar] [CrossRef]
- Kaur, H.; Mukherjee, S.; Baluska, F.; Bhatla, S.C. Regulatory Roles of Serotonin and Melatonin in Abiotic Stress Tolerance in Plants. Plant Signal. Behav. 2015, 10, e1049788. [Google Scholar] [CrossRef]
- Gupta, P.; De, B. Metabolomics Analysis of Rice Responses to Salinity Stress Revealed Elevation of Serotonin, and Gentisic Acid Levels in Leaves of Tolerant Varieties. Plant Signal. Behav. 2017, 12, e1335845. [Google Scholar] [CrossRef] [PubMed]
- Tola, A.J.; Jaballi, A.; Germain, H.; Missihoun, T.D. Recent Development on Plant Aldehyde Dehydrogenase Enzymes and Their Functions in Plant Development and Stress Signaling. Genes 2021, 12, 51. [Google Scholar] [CrossRef]
- Farmer, E.E.; Davoine, C. Reactive Electrophile Species. Curr. Opin. Plant Biol. 2007, 10, 380–386. [Google Scholar] [CrossRef]
- Abubacker, M.N.; Deepalakshmi, T.; Sathya, C. Isolation and Identification of Biolarvicide from Soursop (Annona muricata Linn.) Aqueous Leaf Extract to Mosquito (Aedes aegypti Linn) Larvae. Biolife 2014, 2, 579–585. [Google Scholar]
- Chintem, W.D.G. Larvicidal Potential of the Leaf Exctract of Datura stramonium and Occimum gratissimum against Culex quinquefasciatus Mosquito Species. MSc; Ahmadu Bello University: Zaria, Nigeria, 2013. [Google Scholar]
- Rahuman, A.A.; Gopalakrishnan, G.; Ghouse, B.S.; Arumugam, S.; Himalayan, B. Effect of Feronia Limonia on Mosquito Larvae. Fitoterapia 2000, 71, 553–555. [Google Scholar] [CrossRef]
- Al-Harbi, N.A.; Al Attar, N.M.; Hikal, D.M.; Mohamed, S.E.; Abdel Latef, A.A.H.; Ibrahim, A.A.; Abdein, M.A. Evaluation of Insecticidal Effects of Plants Essential Oils Extracted from Basil, Black Seeds and Lavender against Sitophilus oryzae. Plants 2021, 10, 829. [Google Scholar] [CrossRef]
- Kim, B.-R.; Kim, H.M.; Jin, C.H.; Kang, S.-Y.; Kim, J.-B.; Jeon, Y.G.; Park, K.Y.; Lee, I.-S.; Han, A.-R. Composition and Antioxidant Activities of Volatile Organic Compounds in Radiation-Bred Coreopsis Cultivars. Plants 2020, 9, 717. [Google Scholar] [CrossRef]
- Tangavelou, A.; Viswanathan, M.; Balakrishna, K.; Patra, A. Phytochemical Analysis in the Leaves of Chamaecrista nigricans (Leguminosae). Pharm. Anal. Acta 2018, 9, 582. [Google Scholar] [CrossRef]
- Nyalo, P.; Omwenga, G.; Ngugi, M. Quantitative Phytochemical Profile and In Vitro Antioxidant Properties of Ethyl Acetate Extracts of Xerophyta spekei (Baker) and Grewia tembensis (Fresen). J. Evid. Based Integr. Med. 2023, 28, 2515690X231165096. [Google Scholar] [CrossRef]
- Georgieva, D.N.; Rypniewski, W.; Gabdoulkhakov, A.; Genov, N.; Betzel, C. Asp49 Phospholipase A2–Elaidoylamide Complex: A New Mode of Inhibition. Biochem. Biophys. Res. Commun. 2004, 319, 1314–1321. [Google Scholar] [CrossRef]
- Mohamad, O.A.A.; Li, L.; Ma, J.-B.; Hatab, S.; Xu, L.; Guo, J.-W.; Rasulov, B.A.; Liu, Y.-H.; Hedlund, B.P.; Li, W.-J. Evaluation of the Antimicrobial Activity of Endophytic Bacterial Populations from Chinese Traditional Medicinal Plant Licorice and Characterization of the Bioactive Secondary Metabolites Produced by Bacillus atrophaeus Against Verticillium dahliae. Front. Microbiol. 2018, 9, 924. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Wang, W.; Zu, Y.; Reichling, J.; Suschke, U. Composition and Antimicrobial Activity Study on Essential Oil from the Aerial Parts of Chinese Licorice (Glycyrrhiza uralensis Fisch.). J. Indian Chem. Soc. 2009, 86, 1214–1217. [Google Scholar]
- Tienda-Parrilla, M.; López-Hidalgo, C.; Guerrero-Sanchez, V.M.; Infantes-González, Á.; Valderrama-Fernández, R.; Castillejo, M.-Á.; Jorrín-Novo, J.V.; Rey, M.-D. Untargeted MS-Based Metabolomics Analysis of the Responses to Drought Stress in Quercus ilex L. Leaf Seedlings and the Identification of Putative Compounds Related to Tolerance. Forests 2022, 13, 551. [Google Scholar] [CrossRef]
- Oren, A. Formation and Breakdown of Glycine Betaine and Trimethylamine in Hypersaline Environments. Antonie Van Leeuwenhoek 1990, 58, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Bashir, A.; Hoffmann, T.; Smits, S.H.J.; Bremer, E. Dimethylglycine Provides Salt and Temperature Stress Protection to Bacillus subtilis. Appl. Environ. Microbiol. 2014, 80, 2773–2785. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.H.H.; Murata, N. Glycinebetaine Protects Plants against Abiotic Stress: Mechanisms and Biotechnological Applications. Plant Cell Environ. 2011, 34, 1–20. [Google Scholar] [CrossRef]
- Lanzinger, A.; Frank, T.; Reichenberger, G.; Herz, M.; Engel, K.-H. Metabolite Profiling of Barley Grain Subjected to Induced Drought Stress: Responses of Free Amino Acids in Differently Adapted Cultivars. J. Agric. Food Chem. 2015, 63, 4252–4261. [Google Scholar] [CrossRef]
- Munns, R.; Greenway, H.; Delane, R.; Gibbs, J. Ion Concentration and Carbohydrate Status of the Elongating Leaf Tissue Hordeum vulgare Growing at High External NaCl: II. Cause of the Growth Reduction. J. Exp. Bot. 1982, 33, 574–583. [Google Scholar] [CrossRef]
- Hall, A.J.; Milthorpe, F.L. Assimilate Source-Sink Relationships in Capsicum Annuum L. III. The Effects of Fruit Excision on Photosynthesis and Leaf and Stem Carbohydrates. Funct. Plant Biol. 1978, 5, 1–13. [Google Scholar] [CrossRef]
- Deryabin, A.N.; Trunova, T.I. Colligative Effects of Solutions of Low Molecular Weight Sugars and Their Role in Plants during Hypothermia. Biol. Bull. 2022, 48, S29–S37. [Google Scholar] [CrossRef]
- Latif, M.T.; ChanSiew, M.; Mohd Hanif, N.; Srithawirat, T. Levoglucosan as an Indicator of Biomass Burning from Selected Tropical Plants. Environ. Asia 2012, 5, 22–27. [Google Scholar]
- Noge, K.; Tamogami, S. Herbivore-Induced Phenylacetonitrile Is Biosynthesized from de Novo-Synthesized l-Phenylalanine in the Giant Knotweed, Fallopia Sachalinensis. FEBS Lett. 2013, 587, 1811–1817. [Google Scholar] [CrossRef]
- Seidelmann, K.; Weinert, H.; Ferenz, H.-J. Wings and Legs Are Production Sites for the Desert Locust Courtship-Inhibition Pheromone, Phenylacetonitrile. J. Insect Physiol. 2003, 49, 1125–1133. [Google Scholar] [CrossRef]
- Asano, Y.; Kato, Y. Z-Phenylacetaldoxime Degradation by a Novel Aldoxime Dehydratase from Bacillus sp. Strain OxB-1. FEMS Microbiol. Lett. 1998, 158, 185–190. [Google Scholar] [CrossRef]
- Helms, A.M.; De Moraes, C.M.; Mescher, M.C.; Tooker, J.F. The Volatile Emission of Eurosta solidaginis Primes Herbivore-Induced Volatile Production in Solidago altissima and Does Not Directly Deter Insect Feeding. BMC Plant Biol. 2014, 14, 173. [Google Scholar] [CrossRef]
- Helms, A.M.; De Moraes, C.M.; Tooker, J.F.; Mescher, M.C. Exposure of Solidago altissima Plants to Volatile Emissions of an Insect Antagonist (Eurosta solidaginis) Deters Subsequent Herbivory. Proc. Natl. Acad. Sci. USA 2013, 110, 199–204. [Google Scholar] [CrossRef]
- Ferrer, A.; Altabella, T.; Arró, M.; Boronat, A. Emerging Roles for Conjugated Sterols in Plants. Prog. Lipid Res. 2017, 67, 27–37. [Google Scholar] [CrossRef]
- Babu, S.; Jayaraman, S. An Update on β-Sitosterol: A Potential Herbal Nutraceutical for Diabetic Management. Biomed. Pharmacother. 2020, 131, 110702. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under Stressful Environments: An Overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Terletskaya, N.V.; Seitimova, G.A.; Kudrina, N.O.; Meduntseva, N.D.; Ashimuly, K. The Reactions of Photosynthetic Capacity and Plant Metabolites of Sedum hybridum L. in Response to Mild and Moderate Abiotic Stresses. Plants 2022, 11, 828. [Google Scholar] [CrossRef] [PubMed]
- Shorthouse, J.D.; Rohfritsch, O. Biology of Insect-Induced Galls; Oxford University Press: Oxford, UK, 1992; ISBN 978-0-19-506716-3. [Google Scholar]
- Huang, M.-Y.; Huang, W.-D.; Chou, H.-M.; Chen, C.-C.; Chen, P.-J.; Chang, Y.-T.; Yang, C.-M. Structural, Biochemical, and Physiological Characterization of Photosynthesis in Leaf-Derived Cup-Shaped Galls on Litsea acuminata. BMC Plant Biol. 2015, 15, 61. [Google Scholar] [CrossRef] [PubMed]
- Tooker, J.F.; De Moraes, C.M. Gall Insects and Indirect Plant Defenses: A Case of Active Manipulation? Plant Signal. Behav. 2008, 3, 503–504. [Google Scholar] [CrossRef] [PubMed]
- Oates, C.N.; Külheim, C.; Myburg, A.A.; Slippers, B.; Naidoo, S. The Transcriptome and Terpene Profile of Eucalyptus grandis Reveals Mechanisms of Defense Against the Insect Pest, Leptocybe invasa. Plant Cell Physiol. 2015, 56, 1418–1428. [Google Scholar] [CrossRef] [PubMed]
- Kot, I.; Jakubczyk, A.; Karaś, M.; Złotek, U. Biochemical Responses Induced in Galls of Three Cynipidae Species in Oak Trees. Bull. Entomol. Res. 2018, 108, 494–500. [Google Scholar] [CrossRef]
- Chen, H.; Liu, J.; Cui, K.; Lu, Q.; Wang, C.; Wu, H.; Yang, Z.; Ding, W.; Shao, S.; Wang, H.; et al. Molecular Mechanisms of Tannin Accumulation in Rhus Galls and Genes Involved in Plant-Insect Interactions. Sci. Rep. 2018, 8, 9841. [Google Scholar] [CrossRef]
- Li, X.Q.; Liu, Y.Z.; Guo, W.F.; Solanki, M.K.; Yang, Z.D.; Xiang, Y.; Ma, Z.C.; Wen, Y.G. The Gall Wasp Leptocybe invasa (Hymenoptera: Eulophidae) Stimulates Different Chemical and Phytohormone Responses in Two Eucalyptus Varieties That Vary in Susceptibility to Galling. Tree Physiol. 2017, 37, 1208–1217. [Google Scholar] [CrossRef]
- Nyman, T.; Julkunen-Tiitto, R. Manipulation of the Phenolic Chemistry of Willows by Gall-Inducing Sawflies. Proc. Natl. Acad. Sci. USA 2000, 97, 13184–13187. [Google Scholar] [CrossRef]
- Agudelo, I.; Cogoi, L.; Filip, R.; Kuzmanich, N.; Wagner, M.L.; Ricco, R.A. Anatomy, Histochemistry, and Comparative Analysis of Hydroxycinnamic Derivatives in Healthy Leaves and Galls Induced by Baccharopelma spp. (Hemiptera: Psyllidae) in Baccharis Spicata (Lam) Baill (Asteraceae). Biochem. Syst. Ecol. 2018, 77, 22–30. [Google Scholar] [CrossRef]
- Ananthakrishnan, T.N. Insect Gall Systems: Patterns, Processes and Adaptive Diversity. Curr. Sci. 1998, 75, 672–676. [Google Scholar]
- Platonov, V.V.; Khadartsev, A.A.; Sukhikh, G.T.; Dunaeva, I.V.; Volochaeva, M.V. Chromato-Mass-Spectrometry of the Ethanol Extract of Roots of Glycyrrhiza glabra L. (Family Fabaceae). Bull. New Med. Technol. Electron. Ed. 2020, 14, 137–142. [Google Scholar] [CrossRef]
- Michaelakis, A.; Papachristos, D.; Kimbaris, A.; Koliopoulos, G.; Giatropoulos, A.; Polissiou, M.G. Citrus Essential Oils and Four Enantiomeric Pinenes against Culex pipiens (Diptera: Culicidae). Parasitol. Res. 2009, 105, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Lardeux, F.; Depickère, S.; Duchon, S.; Chavez, T. Insecticide Resistance of Triatoma infestans (Hemiptera, Reduviidae) Vector of Chagas Disease in Bolivia. Trop. Med. Int. Health 2010, 15, 1037–1048. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. Botanical Insecticides, Deterrents, and Repellents in Modern Agriculture and an Increasingly Regulated World. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Hilker, M.; Meiners, T. Plants and Insect Eggs: How Do They Affect Each Other? Phytochemistry 2011, 72, 1612–1623. [Google Scholar] [CrossRef]
- Dambolena, J.S.; Zunino, M.P.; Herrera, J.M.; Pizzolitto, R.P.; Areco, V.A.; Zygadlo, J.A. Terpenes: Natural Products for Controlling Insects of Importance to Human Health—A Structure-Activity Relationship Study. Psyche J. Entomol. 2016, 2016, e4595823. [Google Scholar] [CrossRef]
- Naidoo, S.; Christie, N.; Acosta, J.J.; Mphahlele, M.M.; Payn, K.G.; Myburg, A.A.; Külheim, C. Terpenes Associated with Resistance against the Gall Wasp, Leptocybe invasa, in Eucalyptus grandis. Plant Cell Environ. 2018, 41, 1840–1851. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Yeh, S. Isoprene Emission from Plants. Annu. Rev. Plant Biol. 2001, 52, 407–436. [Google Scholar] [CrossRef]
- Velikova, V.; Edreva, A.; Loreto, F. Endogenous Isoprene Protects Phragmites australis Leaves against Singlet Oxygen. Physiol. Plant. 2004, 122, 219–225. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Drenaggi, E.; Desneux, N.; Maggi, F. Phytol, (E)-Nerolidol and Spathulenol from Stevia rebaudiana Leaf Essential Oil as Effective and Eco-Friendly Botanical Insecticides against Metopolophium dirhodum. Ind. Crops Prod. 2020, 155, 112844. [Google Scholar] [CrossRef]
- Gliszczyńska, A.; Dancewicz, K.; Gabryś, B.; Świtalska, M.; Wietrzyk, J.; Maciejewska, G. Synthesis of Novel Phytol-Derived γ-Butyrolactones and Evaluation of Their Biological Activity. Sci. Rep. 2021, 11, 4262. [Google Scholar] [CrossRef] [PubMed]
- Babu, S.; Tharan, B.; Dhanasekaran, S.; Thushimenan, S.; Kovendan, K.; Alagarmalai, J. Chemical Compositions, Antifeedant and Larvicidal Activity of Pongamia pinnata (L.) against Polyphagous Field Pest, Spodoptera litura. Int. J. Zool. Investig. 2016, 2, 48–57. [Google Scholar]
- Chauhan, N.; Kumar, P.; Mishra, S.; Verma, S.; Malik, A.; Sharma, S. Insecticidal Activity of Jatropha curcas Extracts against Housefly, Musca domestica. Environ. Sci. Pollut. Res. 2015, 22, 14793–14800. [Google Scholar] [CrossRef] [PubMed]
- Okonkwo, C.O.; Moses, S.E. Insecticidal Potentials and Chemical Composition of Ethanol Extracts from the Leaves of Acanthus montanus on Selected Insect Pests. J. Chem. Pharm. Res. 2017, 9, 111–116. [Google Scholar]
- Martínez de Murguía Fernández, L.; Puig Algora, G.; Bajona Roig, M.; Bacchini, G. Effectiveness and Tolerability of a Squalane and Dimethicone-Based Treatment for Head Lice. Parasitol. Res. 2021, 120, 1883–1890. [Google Scholar] [CrossRef] [PubMed]
- Rattan, R.S. Mechanism of Action of Insecticidal Secondary Metabolites of Plant Origin. Crop Prot. 2010, 29, 913–920. [Google Scholar] [CrossRef]
- Panda, A.; Rangani, J.; Parida, A.K. Unraveling Salt Responsive Metabolites and Metabolic Pathways Using Non-Targeted Metabolomics Approach and Elucidation of Salt Tolerance Mechanisms in the Xero-Halophyte Haloxylon salicornicum. Plant Physiol. Biochem. 2021, 158, 284–296. [Google Scholar] [CrossRef]
- Ahmed, Z.; Khan, M.F.; Rana, H. Toxicological Effects of Haloxylon recurvum Bunge Ex Boiss (Khar Boti) Whole Plant Extract and Novel Insecticide Chlorantraniliprole against Maize Weevil, Sitophilus zeamais Motschulsky. Punjab Univ. J. Zool. 2017, 32, 159–166. [Google Scholar]
- Abdel-Farid, I.B.; Massoud, M.S.; Al-Enazy, Y.; Abdel Latef, A.A.H.; Jahangir, M.; Gomaa, N.H. Allelopathic Potential of Haloxylon persicum against Wheat and Black Mustard with Special Reference to Its Phytochemical Composition and Antioxidant Activity. Agronomy 2021, 11, 244. [Google Scholar] [CrossRef]
- Liu, R.; Feng, X.; Li, C.; Ma, J.; Wang, Y.; Li, Y. The Importance of Stem Photosynthesis for Two Desert Shrubs Across Different Groundwater Depths. Front. Plant Sci. 2022, 13, 804786. [Google Scholar] [CrossRef] [PubMed]
- Marikovskij, P.I. New Gall Midges (Diptera, Itonididae) on Saksaul. Entomol. Obozr. 1955, 34, 298–312. [Google Scholar]
- Barykina, R.; Veselova, T.; Devyatov, A.; Dzhalilova, K.K.; Ilyina, G.; Chubatova, N. Guide on Botanical Microtechique. Basics Methods; Moscow University Press: Moscow, Russia, 2004; p. 312. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite Classes | H. aphyllum | H. persicum | ||
|---|---|---|---|---|
| Ctrl | Damaged | Ctrl | Damaged | |
| Fatty acids | ||||
| Hexadecanoic acid | 9.73 ± 0.68 | 5.72 ± 0.74 * | 5.46 ± 0.42 | - |
| 9,12-Octadecadienoic acid (Z,Z)- | - | - | 1.94 ± 0.53 | - |
| 9,12,15-Octadecatrienoic acid, (Z,Z,Z)- | - | - | 4.54 ± 0.23 | 5.44 ± 0.71 |
| Fatty acid esters | ||||
| Hexadecanoic acid, methyl | 4.89 ± 0.47 | 1.92 ± 0.28 * | 2.81 ± 0.29 | 4.39 ± 0.87 * |
| Hexadecanoic acid, octadecyl | 1.65 ± 0.17 | - | - | - |
| 8-Octadecenoic acid, methyl | - | 0.73 ± 0.13 | - | - |
| 9-Octadecenoic acid, methyl (E)- | 1.67 ± 0.14 | - | 2.09 ± 0.31 | 2.84 ± 0.80 |
| 12-Octadecadienoic acid (Z,Z)-, methyl | - | 1.61 ± 0.25 * | - | - |
| 9,12-Octadecadienoic acid (Z,Z)-, methyl | 3.36 ± 0.19 | - | 3.21 ± 0.24 | 4.56 ± 0.55 * |
| 9,12,15-Octadecatrienoic acid, methyl, (Z,Z,Z)- | 5.79 ± 0.18 | 2.62 ± 0.34 * | - | 5.53 ± 0.76 * |
| 9,12,15-Octadecatrienoic acid, ethyl, (Z,Z,Z)- | 2.43 ± 0.04 | 1.03 ± 0.03 * | - | 1.55 ± 0.10 * |
| 9,12,15-Octadecatrienoic acid, 2,3-dihydroxypropyl, (Z,Z,Z)- | 8.92 ± 0.16 | 4.70 ± 0.57 * | - | - |
| Olean-12-en-28-oic acid, 3-oxo-, methyl | - | - | 17.0 ± 0.89 | - |
| Olean-12-en-28-oic acid, 2,3,23-trihydroxy-, methyl, (2α,3β,4α)- | - | 10.1 ± 1.52 * | - | - |
| Olean-12-en-28-oic acid, 3-(acetyloxy)-, methyl, (3β)- | - | 25.1 ± 1.55 * | 6.63 ± 0.59 | - |
| Linoleic acid ethyl | - | - | - | 1.56 ± 0.40 * |
| Dialkyl ethers | ||||
| Ethanol, 2-(9-octadecenyloxy)-, (Z)- | 0.44 ± 0.13 | 0.40 ± 0.09 | - | - |
| Ethanol, 2-(9,12-octadecadienyloxy)-, (Z,Z)- | - | - | - | 3.02 ± 0.29 * |
| Amino acids and their derivatives | ||||
| N,N-Dimethylglycine | 21.2 ± 3.60 | 18.8 ± 3.64 | 7.21 ± 0.15 | - |
| Carbohydrates and derivatives | ||||
| Sucrose | 8.25 ± 0.35 | 3.41 ± 0.34* | 1.92 ± 0.18 | 2.62 ± 0.12 * |
| β-D-Glucopyranose, 1,6-anhydro- (levoglucosan) | - | - | 10.5 ± 0.85 | 9.26 ± 0.79 |
| Aromatic acid derivatives | ||||
| Benzeneacetonitrile, 4-methoxy- | - | - | - | 1.47 ± 0.11 * |
| Benzeneacetonitrile, 4-hydroxy- | - | - | 3.57 ± 0.28 | 3.99 ± 0.25 |
| Phytosterols | ||||
| γ-Sitosterol | 16.9 ± 1.82 | 14.1 ± 0.17* | 4.72 ± 0.23 | 7.19 ± 0.12 * |
| β-Sitosterol acetate | - | - | - | 4.70 ± 0.58 * |
| Stigmasterol | - | - | - | 8.81 ± 0.98 * |
| γ-tocopherol (vitamin E) | ||||
| γ-tocopherol (vitamin E) | 12.7 ± 0.95 | 8.55 ± 0.64 * | 4.99 ± 0.43 | 5.80 ± 0.32 |
| Phenol derivatives | ||||
| 2-Methoxy-4-vinylphenol | 3.31 ± 0.37 | 1.67 ± 0.07 * | 4.18 ± 0.16 | 5.56 ± 0.36 * |
| Terpenoids | ||||
| Squalene | - | 0.89 ± 0.09 * | 2.63 ± 0.13 | 3.21 ± 0.20 * |
| Phytol | - | - | 6.85 ± 0.54 | 6.68 ± 0.46 |
| Phytol, acetate | - | - | 0.92 ± 0.06 | - * |
| Collection Site | Taxation Profile | Damage | |||
|---|---|---|---|---|---|
| GPS | Location | Height, m | Stem Diameter, cm | Crown Diameter, m | |
| H. aphyllum | |||||
| N 44°29′40″ E 62°22′59″ | Kyzylorda region, Kazalinsky district, 57 km southeast of the village Azhar | 1.2 | 2.9 | 1.15 | yes |
| 1.3 | 3.2 | 1.65 | no | ||
| 1.5 | 4.0 | 1.85 | no | ||
| 1.5 | 4.2 | 1.80 | no | ||
| 1.6 | 3.8 | 1.45 | yes | ||
| 1.7 | 5.5 | 1.75 | yes | ||
| H. persicum | |||||
| N 44°55′32″ E 62°15′15″ | Kyzylorda region, Kazalinsky district, 89 km south of Kazalinsk | 1.3 | 3.5 | 1.35 | yes |
| 1.3 | 5.7 | 1.80 | yes | ||
| 1.3 | 7.1 | 1.85 | no | ||
| 1.4 | 3.4 | 1.45 | no | ||
| 1.4 | 3.8 | 1.10 | yes | ||
| 1.6 | 4.4 | 1.25 | no | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terletskaya, N.V.; Mamirova, A.; Ashimuly, K.; Vibe, Y.P.; Krekova, Y.A. Anatomical and Metabolome Features of Haloxylon aphyllum and Haloxylon persicum Elucidate the Resilience against Gall-Forming Insects. Int. J. Mol. Sci. 2024, 25, 4738. https://doi.org/10.3390/ijms25094738
Terletskaya NV, Mamirova A, Ashimuly K, Vibe YP, Krekova YA. Anatomical and Metabolome Features of Haloxylon aphyllum and Haloxylon persicum Elucidate the Resilience against Gall-Forming Insects. International Journal of Molecular Sciences. 2024; 25(9):4738. https://doi.org/10.3390/ijms25094738
Chicago/Turabian StyleTerletskaya, Nina V., Aigerim Mamirova, Kazhybek Ashimuly, Yekaterina P. Vibe, and Yana A. Krekova. 2024. "Anatomical and Metabolome Features of Haloxylon aphyllum and Haloxylon persicum Elucidate the Resilience against Gall-Forming Insects" International Journal of Molecular Sciences 25, no. 9: 4738. https://doi.org/10.3390/ijms25094738
APA StyleTerletskaya, N. V., Mamirova, A., Ashimuly, K., Vibe, Y. P., & Krekova, Y. A. (2024). Anatomical and Metabolome Features of Haloxylon aphyllum and Haloxylon persicum Elucidate the Resilience against Gall-Forming Insects. International Journal of Molecular Sciences, 25(9), 4738. https://doi.org/10.3390/ijms25094738