
How the Ethylene Biosynthesis Pathway of Semi-Halophytes Is Modified with Prolonged Salinity Stress Occurrence?

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
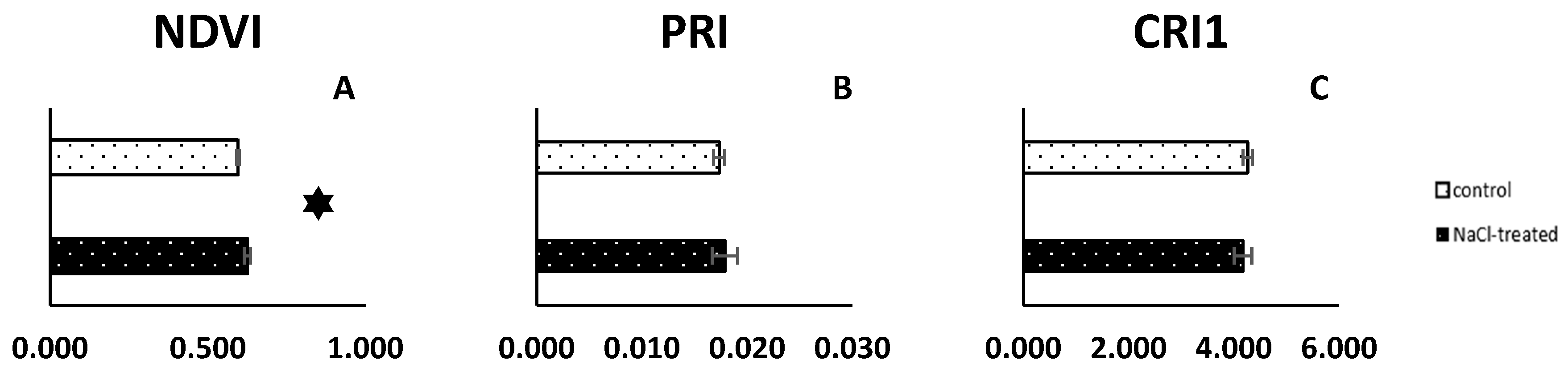
2.1. Reflectance Parameters Results
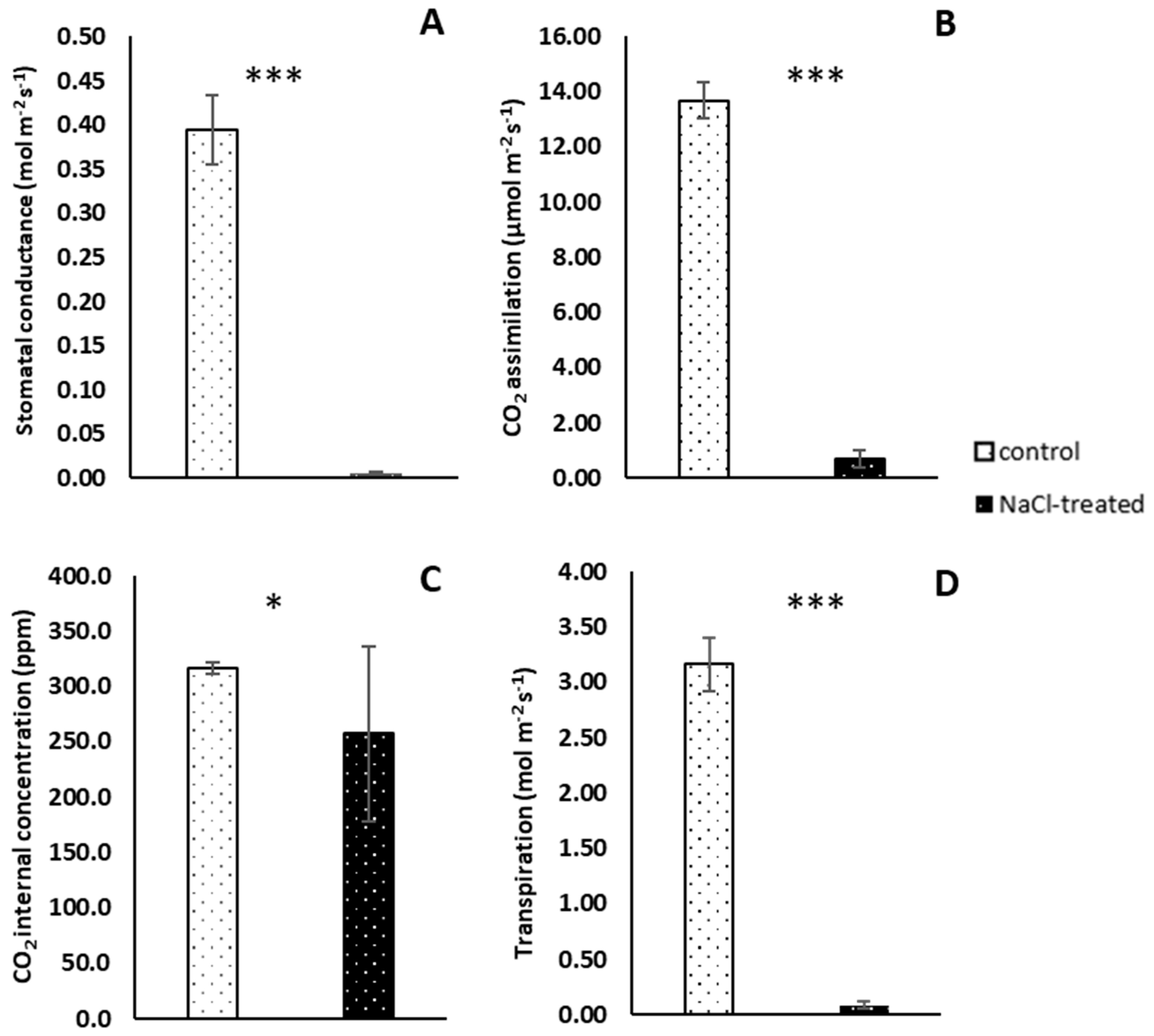
2.2. Gaseous Exchange
2.3. Genome-Wide Identification of Expressed Genes in M. crystallinum under Salinity Stress
2.4. Gene Expression Analysis
2.5. ACC Content Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Cultivation
4.2. Reflectance Measurement
4.3. Gas Exchange Parameters Measurement
4.4. RNA Isolation
4.5. RNA Sequencing
4.6. Bioinformatics Analysis
4.7. qPCR
4.8. HPLC-MS Analysis of ACC Content
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Riyazuddin, R.; Verma, R.; Singh, K.; Nisha, N.; Keisham, M.; Bhati, K.K.; Kim, S.T.; Gupta, R. Ethylene: A master regulator of salinity stress tolerance in plants. Biomolecules 2020, 10, 959. [Google Scholar] [CrossRef] [PubMed]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef] [PubMed]
- Mager, W.H.; de Boer, A.H.; Siderius, M.H.; Voss, H.P. Cellular responses to oxidative and osmotic stress. Cell Stress Chaperones 2000, 5, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Hellebusi, J.A. Osmoregulation. Annu. Rev. Plant Physiol. 1976, 27, 485–505. [Google Scholar] [CrossRef]
- Ozturk, M.; Turkyilmaz Unal, B.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and Its Actions during the Drought Stress in Plants. Physiol. Plant. 2021, 172, 1321–1335. [Google Scholar] [CrossRef]
- Libik-Konieczny, M.; Kuźniak, E.; Surówka, E.; Ślesak, I.; Nosek, M.; Miszalski, Z. Crassulacean acid metabolism and its role in plant acclimatization to abiotic stresses and defence against pathogens. In Progress in Botany; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–30. [Google Scholar]
- Gonçalves, A.Z.; Latansio, S.; Detmann, K.C.; Marabesi, M.A.; Neto, A.A.C.; Aidar, M.P.M.; DaMatta, F.M.; Mercier, H. What does the RuBisCO activity tell us about a C(3)-CAM plant? Plant Physiol. Biochem. 2020, 147, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q.; Tan, B.; Kelley, T.M.; Tian, J.; Chen, S. Physiological Changes in Mesembryanthemum crystallinum During the C3 to CAM Transition Induced by Salt Stress. Front. Plant Sci. 2020, 11, 283. [Google Scholar] [CrossRef]
- Wakamatsu, A.; Mori, I.C.; Matsuura, T.; Taniwaki, Y.; Ishii, R.; Yoshida, R. Possible roles for phytohormones in controlling the stomatal behavior of Mesembryanthemum crystallinum during the salt-induced transition from C3 to Crassulacean acid metabolism. J. Plant Physiol. 2021, 262, 153448. [Google Scholar] [CrossRef]
- Taybi, T.; Cushman, J.C. Abscisic acid signaling and protein synthesis requirements for phosphoenolpyruvate carboxylase transcript induction in the common ice plant. J. Plant Physiol. 2002, 159, 1235–1243. [Google Scholar] [CrossRef]
- Großkinsky, D.K.; Tafner, R.; Moreno, M.V.; Stenglein, S.A.; de Salamone, I.E.G.; Nelson, L.M.; Novák, O.; Strnad, M.; van der Graaff, E.; Roitsch, T. Cytokinin production by Pseudomonas fluorescens G20-18 determines biocontrol activity against Pseudomonas syringae in Arabidopsis. Sci. Rep. 2016, 6, 23310. [Google Scholar] [CrossRef]
- Wilkinson, S.; Davies, W.J. Drought, ozone, ABA and ethylene: New insights from cell to plant to community. Plant Cell Environ. 2010, 33, 510–525. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 2nd ed.; Palgrave Press: London, UK, 2012. [Google Scholar]
- Xu, J.; Zhang, S. Ethylene biosynthesis and regulation in plants. In Ethylene in Plants; Wen, C.K., Ed.; Springer: Dordrecht, The Netherlands, 2015; pp. 1–25. [Google Scholar]
- Sang-Dong, Y.; Younghee, C.; Jen, S. Emerging connections in the ethylene signaling network. Trend. Plant Sci. 2009, 14, 270–279. [Google Scholar]
- De Wild, H.; Otma, E.C.; Peppelenbos, W. Carbon dioxide action on ethylene biosynthesis of preclimacteric and climacteric pear fruit. J. Exp. Bot. 2013, 54, 1537–1544. [Google Scholar] [CrossRef]
- Guo, H.; Sun, Y.; Li, Y.; Zhang, W.; Ge, F. Elevated CO2 decreases the response of the ethylene signalling pathway in Medicago truncatula and increases the abundance of the pea aphid. New Phytol. 2014, 201, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Kazan, K. Diverse roles of jasmonates and ethylene in abiotic stress tolerance. Trends Plant Sci. 2015, 20, 219–229. [Google Scholar] [CrossRef] [PubMed]
- AbdElgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R.; Asard, H.; Abuelsoud, W. High salinity induces different oxidative stress and antioxidant responses in maize seedlings organs. Front. Plant Sci. 2016, 7, 276. [Google Scholar] [CrossRef]
- Xu, L.; Xiang, G.; Sun, Q.; Ni, Y.; Jin, Z.; Gao, Z.; Yao, Y. Melatonin enhances salinity tolerance by promoting MYB108A-mediated ethylene biosynthesis in grapevines. Hortic. Res. 2019, 6, 114. [Google Scholar] [CrossRef]
- Freitas, V.S.; de Souza Miranda, R.; Costa, J.H.; de Oliveira, D.F.; de Oliveira Paula, S.; de Castro Miguel, E.; Freire, R.S.; Prisco, J.T.; Gomes-Filho, E. Ethylene triggers salinity tolerance in maize genotypes by modulating polyamine catabolism enzymes associated with H2O2 production. Environ. Exp. Bot. 2018, 145, 75–86. [Google Scholar] [CrossRef]
- Gharbi, E.; Martínez, J.P.; Benahmed, H.; Lepoint, G.; Vanpee, B.; Quinet, M.; Lutts, S. Inhibition of ethylene synthesis reduces salinity-tolerance in tomato wild relative species Solanum chilense. J. Plant Physiol. 2017, 210, 24–37. [Google Scholar] [CrossRef]
- Yang, C.; Ma, B.; He, S.J.; Xiong, Q.; Duan, K.X.; Yin, C.C.; Chen, H.; Lu, X.; Chen, S.H.; Zhang, J.H. MAOHUZI6/ETHYLENE INSENSITIVE3-LIKE1 and ETHYLENE INSENSITIVE3-LIKE2 regulate ethylene response of roots and coleoptiles and negatively affect salinity tolerance in rice. Plant Physiol. 2015, 169, 148–165. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; He, S.; Gong, W.; Sun, J.; Pan, Z.; Xu, F.; Lu, Y.; Du, X. Comprehensive analysis of differentially expressed genes and transcriptional regulation induced by salt stress in two contrasting cotton genotypes. BMC Genom. 2014, 15, 760. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.L.; Cao, W.H.; Cao, Y.R.; Liu, J.; Hao, Y.J.; Zhang, J.S.; Chen, S.Y. Roles of ethylene receptor NTHK1 domains in plant growth, stress response and protein phosphorylation. FEBS Lett. 2006, 580, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.H.; Liu, J.; He, X.J.; Mu, R.L.; Zhou, H.L.; Chen, S.Y.; Zhang, J.S. Modulation of ethylene responses affects plant salinity-stress responses. Plant Physiol. 2007, 143, 707–719. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Liu, J.; Lei, G.; Liu, Y.F.; Li, Z.G.; Tao, J.J.; Hao, Y.J.; Cao, Y.R.; Lin, Q.; Zhang, W.K.; et al. Effects of tobacco Ethylene receptor mutations on receptor kinase activity, plant growth and stress responses. Plant Cell Physiol. 2009, 50, 1636–1650. [Google Scholar] [CrossRef] [PubMed]
- Lei, G.; Shen, M.; Li, Z.G.; Zhang, B.; Duan, K.X.; Wang, N.; Cao, Y.R.; Zhang, W.K.; Ma, B.; Ling, H.Q.; et al. EIN2 regulates salinity stress response and interacts with a MA3 domain-containing protein ECIP1 in Arabidopsis. Plant Cell Environ. 2011, 34, 1678–1692. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Chen, H.; Ma, B.; Zhang, W.; Chen, S.; Zhang, J. The role of ethylene in plants under salinity stress. Front. Plant Sci. 2015, 6, 1059. [Google Scholar] [CrossRef]
- Kholodova, V.; Volkov, K.; Kuznetsov, V. Adaptation of the common ice plant to high copper and zinc concentrations and their potential using for phytoremediation. Russ. J. Plant Physiol. 2005, 52, 748–757. [Google Scholar] [CrossRef]
- Amari, T.; Ghnaya, T.; Debez, A.; Taamali, M.; Ben Youssef, N.; Lucchini, G.; Sacchi, G.A.; Abdelly, C. Comparative Ni tolerance and accumulation potentials between Mesembryanthemum crystallinum (halophyte) and Brassica juncea: Metal accumulation, nutrient status and photosynthetic activity. J. Plant Physiol. 2014, 171, 1634–1644. [Google Scholar] [CrossRef]
- Śliwa-Cebula, M.; Kaszycki, P.; Kaczmarczyk, A.; Nosek, M.; Lis-Krzyścin, A.; Miszalski, Z. The Common Ice Plant (Mesembryanthemum crystallinum L.)–Phytoremediation Potential for Cadmium and Chromate-Contaminated Soils. Plants 2020, 9, 1230. [Google Scholar] [CrossRef] [PubMed]
- Niewiadomska, E.; Bilger, W.; Gruca, M.; Mulisch, M.; Miszalski, Z.; Krupinska, K. CAM-related changes in chloroplastic metabolism of Mesembryanthemum crystallinum L. Planta 2011, 233, 275–285. [Google Scholar] [CrossRef]
- Winter, K.; Holtum, J.A.M. Facultative crassulacean acid metabolism (CAM) plants: Powerful tools for unravelling the functional elements of CAM photosynthesis. J. Exp. Bot. 2014, 65, 3425–3441. [Google Scholar] [CrossRef] [PubMed]
- Winter, K. Ecophysiology of constitutive and facultative CAM photosynthesis. J. Exp. Bot. 2019, 70, 6495–6508. [Google Scholar] [CrossRef] [PubMed]
- Nosek, M.; Gawrońska, K.; Rozpądek, P.; Szechyńska-Hebda, M.; Kornaś, A.; Miszalski, Z. Withdrawal from functional Crassulacean acid metabolism (CAM) is accompanied by changes in both gene expression and activity of antioxidative enzymes. J. Plant Physiol. 2018, 229, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Nosek, M.; Gawrońska, K.; Rozpądek, P.; Sujkowska-Rybkowska, M.; Miszalski, Z.; Kornaś, A. At the edges of photosynthetic metabolic plasticity-on the rapidity and extent of changes accompanying salinity stress-induced CAM photosynthesis withdrawal. Int. J. Mol. Sci. 2021, 22, 8426. [Google Scholar] [CrossRef] [PubMed]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W.; Harlan, J.C. Monitoring the Vernal Advancement of Natural Vegetation; Final report; NASA/GCSFC: Greenbelt, MD, USA, 1974. [Google Scholar]
- Gamon, J.A.; Peñuelas, J.; Field, C.B. A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency. Remote Sens. Environ. 1992, 41, 35–44. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Winter, K.; Gademann, R. Daily changes in CO2 and water vapor exchange, chlorophyll fluorescence, and leaf water relations in the halophyte Mesembryanthemum crystallinum during the induction of crassulacean acid metabolism in response to high NaCl salinity. Plant Physiol. 1991, 95, 768–776. [Google Scholar] [CrossRef]
- Szarek, S.R.; Ting, I.P. Photosynthetic Efficiency of CAM Plants in Relation to C3 and C4 Plants; Springer: Dordrecht, The Netherlands, 1975; pp. 289–297. [Google Scholar]
- Bhagwat, A.S. Photosynthetic carbon assimilation of C3, C4, and CAM pathways. In Handbook of Photosynthesis, 2nd ed.; Pessarakli, M., Ed.; Taylor and Francis Group: Tucson, AZ, USA, 2005; pp. 367–389. [Google Scholar]
- Wingler, A.; Lea, P.J.; Quick, W.P.; Leegood, R.C. Photorespiration: Metabolic pathways and their role in stress protection. Philos. Trans. R. Soc. Lond. Ser. B 2000, 355, 1517–1529. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.K.; Shabala, S. Mechanisms of plant responses and adaptation to soil salinity. Innovation 2020, 1, 100017. [Google Scholar] [PubMed]
- Volkov, V.; Wang, B.; Dominy, P.J.; Fricke, W.; Amtmann, A. Thellungiella halophila, a salt tolerant relative of Arabidopsis thaliana, possesses effective mechanisms to discriminate between potassium and sodium. Plant Cell Environ. 2004, 27, 1–14. [Google Scholar] [CrossRef]
- Percey, W.J.; Shabala, L.; Wu, Q.; Su, N.; Breadmore, M.C.; Guijt, R.M.; Bose, J.; Shabala, S. Potassium retention in leaf mesophyll as an element of salinity tissue tolerance in halophytes. Plant Physiol. Biochem. 2016, 109, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Cadenas, A.; Tadeo, F.R.; Primo-Millo, E.; Talón, M. Involvement of abscisic acid and ethylene in the responses of citrus seedlings to salinity shock. Physiol. Plant. 1998, 103, 475–484. [Google Scholar] [CrossRef]
- Mühlenbock, P.; Szechyńska-Hebda, M.; Płaszczyca, M.; Baudo, M.; Mateo, A.; Mullineaux, P.M.; Parker, J.E.; Karpińska, B.; Karpiński, S. Chloroplast signaling and LESION SIMULATING DISEASE1 regulate crosstalk between light acclimation and immunity in Arabidopsis. Plant Cell. 2008, 20, 2339–2356. [Google Scholar] [CrossRef] [PubMed]
- Arraes, F.B.M.; Beneventi, M.A.; de Sa, M.E.L.; Paixao, J.F.R.; Albuquerque, E.V.S.; Marin, S.R.R.; Purgatto, E.; Nepomuceno, A.L.; Grossi-de-Sa, M.F. Implications of ethylene biosynthesis and signaling in soybean drought stress tolerance. BMC Plant Biol. 2015, 15, 213. [Google Scholar] [CrossRef] [PubMed]
- Park, S.S.-Y.; Fung, P.; Nishimura, N.; Jensen, D.R.; Fujii, H.; Zhao, Y.; Lumba, S.; Santiago, J.; Rodrigues, A.; Chow, T.F.; et al. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL Family of START proteins. Science 2009, 324, 1068–1069. [Google Scholar] [CrossRef] [PubMed]
- Osakabe, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S.P. ABA control of plant macroelement membrane transport systems in response to water deficit and high salinity. New Phytol. 2014, 202, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Munné-Bosch, S. Ethylene response factors: A key regulatory hub in hormone and stress signaling. Plant Physiol. 2015, 169, 32–41. [Google Scholar] [CrossRef]
- Huang, J.; Huang, Z.; Zhou, X.; Xia, C.; Imran, M.; Wang, S.; Xu, C.; Zha, M.; Liu, Y.; Zhang, C. Tissue-specific transcriptomic profiling of Plantago major provides insights for the involvement of vasculature in phosphate deficiency responses. Mol. Genet. Genom. 2019, 294, 159–175. [Google Scholar] [CrossRef]
- Lin, R.C.; Park, H.J.; Wang, H.Y. Role of Arabidopsis RAP2.4 in regulating light- and ethylene-mediated developmental processes and drought stress tolerance. Mol. Plant. 2008, 1, 42–57. [Google Scholar] [CrossRef] [PubMed]
- Papdi, C.; Perez-Salamo, I.; Joseph, M.P.; Giuntoli, B.; Bogre, L.; Koncz, C.; Szabados, L. The low oxygen, oxidative and osmotic stress responses synergistically act through the ethylene response factor VII genes RAP2.12, RAP2.2 and RAP2.3. Plant J. 2015, 82, 772–784. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Zhang, Q.; Liu, Q.; Zhai, H.; Zhao, N.; He, S. An AP2/ERF gene, IbRAP2-12, from sweetpotato is involved in salinity and drought tolerance in transgenic Arabidopsis. J. Plant Sci. 2019, 281, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, E.-J.; Cheng, M.-C.; Lin, T.-P. Functional characterization of an abiotic stress-inducible transcription factor AtERF53 in Arabidopsis thaliana. Plant Mol. Biol. 2013, 82, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Resnick, J.S.; Wen, C.K.; Shockey, J.A.; Chang, C. REVERSION-TO-ETHYLENE SENSITIVITY1, a conserved gene that regulates ethylene receptor function in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 7917–7922. [Google Scholar] [CrossRef]
- Li, S.; Zhou, X.; Chen, L.; Huang, W.; Yu, D. Functional characterization of Arabidopsis thaliana WRKY39 in heat stress. Mol. Cells 2010, 29, 475–483. [Google Scholar] [CrossRef]
- Argueso, C.T.; Hansen, M.; Kieber, J.J. Regulation of ethylene biosynthesis. J. Plant Growth Regul. 2007, 26, 92–105. [Google Scholar] [CrossRef]
- Lin, Z.; Zhong, S.; Grierson, D. Recent advances in ethylene research. J. Exp. Bot. 2009, 60, 3311–3336. [Google Scholar] [CrossRef]
- Available online: https://www.arabidopsis.org/index.jsp (accessed on 27 February 2024).
- Jiang, C.; Belfield, E.J.; Cao, Y.; Smith, J.A.; Harberd, N.P. An Arabidopsis soil-salinity-tolerance mutation confers ethylene-mediated enhancement of sodium/potassium homeostasis. Plant Cell 2013, 25, 3535–3552. [Google Scholar] [CrossRef]
- Wang, N.N.; Shih, M.C.; Li, N. The GUS reporter-aided analysis of the promoter activities of Arabidopsis ACC synthase genes AtACS4, AtACS5, and AtACS7 induced by hormones and stresses. J. Exp. Bot. 2005, 56, 909–920. [Google Scholar] [CrossRef]
- Achard, P.; Cheng, H.; De Grauwe, L.; Decat, J.; Schoutteten, H.; Moritz, T.; Van Der Straeten, D.; Peng, J.; Harberd, N.P. Integration of plant responses to environmentally activated phytohormonal signals. Science 2006, 311, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.H.; Liu, J.; Zhou, Q.Y.; Cao, Y.R.; Zheng, S.F.; Du, B.X.; Zhang, J.S.; Chen, S.Y. Expression of tobacco ethylene receptor NTHK1 alters plant responses to salt stress. Plant Cell Environ. 2006, 29, 1210–1219. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Li, Z.; Wen, X.; Li, W.; Shi, H.; Yang, L.; Zhu, H.; Gou, H. Salt induced stabilization of EIN3/EIL1 confers salinity tolerance by deterring ROS accumulation in Arabidopsis. PLoS Genet. 2014, 10, e1004664. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Wang, Z.; Song, X.; Xu, J.; Jiang, C.; Zhao, Y.; Ma, C.; Zhang, H. Transcriptomic profiling revealed an important role of cell wall remodelling and ethylene signalling pathway during salt acclimation in Arabidopsis. Plant Mol. Biol. 2014, 86, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Ellouzi, H.; Hamed, K.B.; Hernandez, I.; Cela, J.; Muller, M.; Magne, C.; Abdelly, C.; Munne-Bosch, S. A comparative study of the early osmotic, ionic, redox and hormonal signaling response in leaves and roots of two halophytes and a glycophyte to salinity. Planta 2014, 240, 1299–1317. [Google Scholar] [CrossRef]
- Kukreja, S.; Nandwal, A.; Kumar, N.; Sharma, S.; Unvi, V.; Sharma, P. Plant water status, H2O2 scavenging enzymes, ethylene evolution and membrane integrity of Cicer arietinum roots as affected by salinity. Biol. Plant. 2005, 49, 305–308. [Google Scholar] [CrossRef]
- Chen, D.; Ma, X.; Li, C.; Zhang, W.; Xia, G.; Wang, M. A wheat aminocyclopropane-1-carboxylate oxidase gene, TaACO1, negatively regulates salinity stress in Arabidopsis thaliana. Plant Cell Rep. 2014, 33, 1815–1827. [Google Scholar] [CrossRef]
- Miszalski, Z.; Kornas, A.; Rozpądek, P.; Fischer-Schliebs, E.; Lüttge, U. Independent fluctuations of malate and citrate in the CAM species Clusia hilariana Schltdl. under low light and high light in relation to photoprotection. J. Plant Physiol. 2015, 170, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Andrew, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 29th May 2022).
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolau, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-Seq: Reference generation and analysis with Trinity. Nat. Protocs. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Mykles, D.L. A comparison of resources for the annotation of a de novo assembled transcriptome in the molting gland (Y-Organ) of the blackback land crab, Gecarcinus lateralis. Integr. Comp. Biol. 2016, 56, 1103–1112. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 6, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, 7. [Google Scholar] [CrossRef]
- Alexa, A.; Rahnenfuhrer, J. topGO: Enrichment Analysis for Gene Ontology, R Package 2.26.0; R Project for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Munné-Bosch, S. Rapid and sensitive hormonal profiling of complex plant samples by liquid chromatography coupled to electrospray ionization tandem mass spectrometry. Plant Met. 2011, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Annotated Species | Description | Unigene ID | log2 Fold Change NaCl-Treated vs. Control | p Value |
|---|---|---|---|---|---|
| Ethylene (ET)-related genes (ERGs) | |||||
| ACO | A. deliciosa | 1-aminocyclopropane-1-carboxylate oxidase | TRINITY_DN78175 | 3.0 | 2 × 10−24 |
| TRINITY_DN69902 | 2.8 | 4 × 10−19 | |||
| ACO5 | A. thaliana | 1-aminocyclopropane-1-carboxylate oxidase | TRINITY_DN31732 | −1.5 | 5 × 10−4 |
| ETR | P. persica | Ethylene receptor | TRINITY_DN29696 | 1.7 | 4 × 10−4 |
| RTE1 | A. thaliana | Protein reversion-to-ethylene sensitivity1 | TRINITY_DN30452 | 2.2 | 1 × 10−6 |
| RAP2.12 | A. thaliana | Ethylene-responsive transcription factor RAP2-12 | TRINITY_DN11475 | 1.8 | 1 × 10−12 |
| TRINITY_DN18663 | 2.4 | 3 × 10−7 | |||
| RAP2.3 | A. thaliana | Ethylene-responsive transcription factor RAP2-3 | TRINITY_DN19854 | 1.5 | 2 × 10−8 |
| RAP2.4 | A. thaliana | Ethylene-responsive transcription factor RAP2-4 | TRINITY_DN50762 | 1.3 | 4 × 10−4 |
| ERF53 | A. thaliana | Ethylene-responsive transcription factor ERF053 | TRINITY_DN24801 | −2.0 | 9 × 10−10 |
| ERF61 | A. thaliana | Ethylene-responsive transcription factor ERF061 | TRINITY_DN79349 | 2.9 | 4 × 10−12 |
| ERF80 | A. thaliana | Ethylene-responsive transcription factor 9 | TRINITY_DN32188 | 2.0 | 8 × 10−4 |
| AP2L1 | A. thaliana | AP2-like ethylene-responsive transcription factor At1g16060 | TRINITY_DN25103 | 1.2 | 7 × 10−5 |
| RAV2 | A. thaliana | AP2/ERF and B3 domain-containing transcription repressor RAV2 | TRINITY_DN25847 | −4.4 | 7 × 10−13 |
| WRKY23 | A. thaliana | WRKY transcription factor 23 | TRINITY_DN33449 | 1.4 | 4 × 10−6 |
| Abscisic acid (ABA) catabolic process (GO:0046345) | |||||
| PPCK1 | A. thaliana | Phosphoenolpyruvate carboxylase kinase 1 | TRINITY_DN29388 | 4.5 | 7 × 10−17 |
| TRINITY_DN25641 | 3.7 | 1 × 10−7 | |||
| ATHB7 | A. thaliana | Homeobox-leucine zipper protein ATHB-7 | TRINITY_DN28056 | 2.6 | 1 × 10−17 |
| TRINITY_DN23684 | 2.6 | 7 × 10−15 | |||
| TRINITY_DN89546 | 2.4 | 1 × 10−17 | |||
| TRINITY_DN29717 | 2.2 | 3 × 10−26 | |||
| BEL1 | A. thaliana | Homeobox protein BEL1 homolog | TRINITY_DN18515 | 2.1 | 2 × 10−13 |
| BELH1 | A. thaliana | BEL1-like homeodomain protein 1 | TRINITY_DN27069 | 1.9 | 4 × 10−21 |
| CAR4 | A. thaliana | Protein C2-domain ABA-related 4 | TRINITY_DN24769 | 3.7 | 3 × 10−5 |
| TRINITY_DN17184 | 2.7 | 4 × 10−12 | |||
| P2C24 | A. thaliana | Probable protein phosphatase 2C 24 | TRINITY_DN25846 | 3.9 | 3 × 10−20 |
| P2C37 | A. thaliana | Protein phosphatase 2C 37 | TRINITY_DN2812 | 1.2 | 5 × 10−8 |
| P2C56 | A. thaliana | Protein phosphatase 2C 56 | TRINITY_DN30187 | 2.0 | 8 × 10−11 |
| ALFC2 | P. sativum | Fructose-bisphosphate aldolase 2 | TRINITY_DN12822 | 2.9 | 2 × 10−6 |
| PIP22 | A. thaliana | Aquaporin PIP2-2 | TRINITY_DN16293 | 2.4 | 1 × 10−8 |
| ERD7 | A. thaliana | Protein early-responsive to dehydration 7 | TRINITY_DN11051 | 2.6 | 3 × 10−8 |
| AFP2 | A. thaliana | Ninja-family protein AFP2 | TRINITY_DN37779 | 3.5 | 8 × 10−6 |
| AMO | A. thaliana | Primary amine oxidase | TRINITY_DN32642 | 2.8 | 5 × 10−4 |
| TRINITY_DN33051 | 3.5 | 8 × 10−13 | |||
| SAPK2 | O. sativa | Serine/threonine-protein kinase SAPK2 | TRINITY_DN39476 | 2.9 | 2 × 10−12 |
| SRK2I | A. thaliana | Serine/threonine-protein kinase SRK2I | TRINITY_DN37625 | 1.2 | 2 × 10−6 |
| CIPK1 | A. thaliana | CBL-interacting serine/threonine-protein kinase 1 | TRINITY_DN29316 | 1.2 | 2 × 10−5 |
| Y1141 | A. thaliana | G-type lectin S-receptor-like serine/threonine-protein kinase At1g11410 | TRINITY_DN40172 | 1.6 | 2 × 10−14 |
| CRK | D. carota | CDPK-related protein kinase | TRINITY_DN40076 | 1.6 | 2 × 10−7 |
| CDPKO | O. sativa | Calcium-dependent protein kinase 24 | TRINITY_DN32679 | 1.0 | 1 × 10−14 |
| RMR41 | A. thaliana | Remorin 4.1 | TRINITY_DN21179 | 1.1 | 3 × 10−5 |
| AP2L1 | A. thaliana | AP2-like ethylene-responsive transcription factor At1g16060 | TRINITY_DN25103 | 1.3 | 7 × 10−5 |
| CRPM4 | A. thaliana | Cold-regulated 413 plasma membrane protein 4 | TRINITY_DN33710 | 1.7 | 7 × 10−5 |
| RGLG2 | A. thaliana | E3 ubiquitin-protein ligase RGLG2 | TRINITY_DN33838 | 2.9 | 5 × 10−5 |
| GBLPA | A. thaliana | Receptor for activated C kinase 1A | TRINITY_DN3696 | 1.8 | 2 × 10−5 |
| HHP1 | A. thaliana | Heptahelical transmembrane protein 1 | TRINITY_DN40228 | 2.4 | 2 × 10−13 |
| ECP44 | D. carota | Phosphoprotein ECPP44 | TRINITY_DN50318 | 1.8 | 1 × 10−4 |
| ANXD4 | A. thaliana | Annexin D4 | TRINITY_DN59321 | 1.1 | 6 × 10−5 |
| GOLS2 | A. thaliana | Galactinol synthase 2 | TRINITY_DN63076 | 3.6 | 4 × 10−6 |
| SRM1 | A. thaliana | Transcription factor SRM1 | TRINITY_DN1724 | 1.3 | 6 × 10−8 |
| MYB88 | A. thaliana | Transcription factor MYB88 | TRINITY_DN28071 | 1.3 | 1 × 10−4 |
| MYBS3 | O. sativa | Transcription factor MYBS3 | TRINITY_DN9971 | 1.2 | 2 × 10−5 |
| MY102 | A. thaliana | Transcription factor MYB102 | TRINITY_DN36820 | 1.1 | 5 × 10−4 |
| NAP2 | S.lycopersicum | NAC domain-containing protein 2 | TRINITY_DN23251 | 1.6 | 1 × 10−6 |
| NAC2 | A. thaliana | NAC domain-containing protein 2 | TRINITY_DN2528 | 1.5 | 8 × 10−6 |
| AL7B4 | A. thaliana | Aldehyde dehydrogenase family 7 member B4 | TRINITY_DN49632 | 1.3 | 1 × 10−9 |
| TRINITY_DN23905 | 1.2 | 7 × 10−4 | |||
| CP29B | A. thaliana | RNA-binding protein CP29B | TRINITY_DN42372 | −1.6 | 9 × 10−4 |
| Abscisic acid catabolic process (GO:0046345), (+)-abscisic acid 8′-hydroxylase activity (GO:0010295) | |||||
| ABAH2 | A. thaliana | Abscisic acid 8′-hydroxylase 2 | TRINITY_DN51717 | −2.6 | 2 × 10−17 |
| TRINITY_DN36601 | −2.3 | 9 × 10−25 | |||
| ABAH4 | A. thaliana | Abscisic acid 8′-hydroxylase 4 | TRINITY_DN87577 | −7.4 | 2 × 10−18 |
| Nitric oxide biosynthetic process (GO:0006809) | |||||
| NOS | A. thaliana | Nitric oxide synthase | TRINITY_DN39531 | 1.8 | 1 × 10−4 |
| AMO | A. thaliana | Primary amine oxidase | TRINITY_DN32642 | 2.8 | 5 × 10−4 |
| TRINITY_DN33051 | 3.5 | 8 × 10−13 | |||
| Regulation of stomatal movement (GO:0010119) | |||||
| BCA1 | A. thaliana | Beta carbonic anhydrase 1 | TRINITY_DN1383 | 2.2 | 4 × 10−15 |
| TRINITY_DN16509 | 2.3 | 1 × 10−7 | |||
| BCA2 | A. thaliana | Beta carbonic anhydrase 2 | TRINITY_DN16509 | 2.6 | 9 × 10−5 |
| TRINITY_DN1383 | 2.7 | 5 × 10−28 | |||
| CAH2 | F. linearis | Carbonic anhydrase 2 | TRINITY_DN38373 | 1.6 | 1 × 10−5 |
| P2C37 | A. thaliana | Protein phosphatase 2C 37 | TRINITY_DN2812 | 1.2 | 5 × 10−8 |
| P2C56 | A. thaliana | Protein phosphatase 2C 56 | TRINITY_DN30187 | 2.0 | 8 × 10−11 |
| SRK2I | A. thaliana | Serine/threonine-protein kinase SRK2I | TRINITY_DN39476 | 2.9 | 2 × 10−12 |
| ZIFL1 | A. thaliana | Protein zinc induced facilitator-like 1 | TRINITY_DN54306 | 2.4 | 2 × 10−12 |
| TRINITY_DN38574 | 2.1 | 8 × 10−12 | |||
| AKT1 | A. thaliana | Potassium channel AKT1 | TRINITY_DN28154 | 2.1 | 1 × 10−5 |
| NHX2 | A. thaliana | Sodium/hydrogen exchanger 2 | TRINITY_DN68370 | 1.4 | 1 × 10−20 |
| MYB61 | A. thaliana | Transcription factor MYB61 | TRINITY_DN31421 | 2.1 | 3 × 10−8 |
| Reductive pentose-phosphate cycle (GO:0019253), ribulose-bisphosphate carboxylase activity (GO:0016984) | |||||
| RBS1 | M. crystallinum | Ribulose bisphosphate carboxylase small chain 1 | TRINITY_DN71513 | −2.1 | 1 × 10−9 |
| RBS3 | M. crystallinum | Ribulose bisphosphate carboxylase small chain 3 | TRINITY_DN28102 | −1.2 | 2 × 10−3 |
| TRINITY_DN39730 | −1.5 | 1 × 10−3 | |||
| RBS4 | M. crystallinum | Ribulose bisphosphate carboxylase small chain 4 | TRINITY_DN24229 | −3.2 | 5 × 10−7 |
| RBS5 | M. crystallinum | Ribulose bisphosphate carboxylase small chain 5 | TRINITY_DN16432 | −4.0 | 4 × 10−17 |
| RBS6 | M. crystallinum | Ribulose bisphosphate carboxylase small chain 6 | TRINITY_DN90082 | −1.9 | 9 × 10−13 |
| RAF2 | A. thaliana | Rubisco accumulation factor 1.2, chloroplastic | TRINITY_DN4605 | −1.1 | 3 × 10−20 |
| Reductive pentose-phosphate cycle (GO:0019253) | |||||
| KPPR | M. crystallinum | Phosphoribulokinase | TRINITY_DN40068 | −1.6 | 6 × 10−7 |
| TRINITY_DN78943 | −1.3 | 4 × 10−7 | |||
| TRINITY_DN42604 | −1.2 | 3 × 10−5 | |||
| G3PB | A. thaliana | Glyceraldehyde-3-phosphate dehydrogenase GAPB | TRINITY_DN43620 | −1.7 | 1 × 10−3 |
| S17P | T. aestivum | Sedoheptulose-1,7-bisphosphatase | TRINITY_DN88837 | −1.2 | 2 × 10−5 |
| Glycine catabolic process (GO:0006546) | |||||
| GCSH | F. anomala | Glycine cleavage system H protein, mitochondrial | TRINITY_DN54747 | −2.1 | 4 × 10−10 |
| M. crystallinum | TRINITY_DN70038 | −1.8 | 3 × 10−12 | ||
| GCST | M. crystallinum | Aminomethyltransferase, mitochondrial | TRINITY_DN22073 | −1.7 | 2 × 10−6 |
| TRINITY_DN78482 | −1.3 | 4 × 10−14 | |||
| GCSP | S. tuberosum | Glycine dehydrogenase (decarboxylating), mitochondrial | TRINITY_DN92007 | −2.1 | 1 × 10−6 |
| TRINITY_DN95999 | −1.9 | 5 × 10−9 | |||
| Chlorophyll binding (GO:0016168), chloroplast thylakoid membrane (GO:0009535), photosystem I (GO:0009522), photosystem II (GO:0009523), photosynthesis, light harvesting (GO:0009765) | |||||
| CB2A | S. oleracea | Chlorophyll a-b binding protein | TRINITY_DN60950 | −3.3 | 6 × 10−4 |
| TRINITY_DN38282 | −3.0 | 2 × 10−3 | |||
| TRINITY_DN4800 | −4.2 | 6 × 10−5 | |||
| CB2D | S. lycopersicum | Chlorophyll a-b binding protein 1D | TRINITY_DN22522 | −4.6 | 5 × 10−12 |
| TRINITY_DN30304 | −2.5 | 1 × 10−5 | |||
| TRINITY_DN40637 | −3.7 | 2 × 10−4 | |||
| TRINITY_DN68714 | −4.1 | 2 × 10−4 | |||
| TRINITY_DN80965 | −2.3 | 4 × 10−4 | |||
| TRINITY_DN30304 | −2.8 | 2 × 10−3 | |||
| TRINITY_DN11494 | −2.9 | 3 × 10−3 | |||
| CB4C | A. thaliana | Chlorophyll a-b binding protein CP29.3 | TRINITY_DN42750 | −1.6 | 7 × 10−4 |
| CB21 | S. latifolia | Chlorophyll a-b binding protein | TRINITY_DN38282 | −2.7 | 5 × 10−24 |
| TRINITY_DN51262 | −2.6 | 6 × 10−20 | |||
| R. sativus | Chlorophyll a-b binding of LHCII type 1 protein | TRINITY_DN63494 | −3.0 | 2 × 10−5 | |
| TRINITY_DN69269 | −1.8 | 2 × 10−3 | |||
| CB23 | N. tabacum | Chlorophyll a-b binding protein 36 | TRINITY_DN59686 | −2.9 | 3 × 10−7 |
| TRINITY_DN38282 | −2.6 | 3 × 10−19 | |||
| TRINITY_DN88301 | −2.4 | 2 × 10−10 | |||
| CB27 | N. tabacum | Chlorophyll a-b binding protein 7 | TRINITY_DN27874 | −3.0 | 2 × 10−3 |
| TRINITY_DN28870 | −3.8 | 4 × 10−6 | |||
| TRINITY_DN30304 | −3.1 | 9 × 10−4 | |||
| Photosystem II (GO:0009523), chloroplast thylakoid membrane (GO:0009535), photosynthesis, light harvesting (GO:0009765) | |||||
| CP29B | A. thaliana | RNA-binding protein CP29B | TRINITY_DN90726 | −3.1 | 2 × 10−12 |
| TRINITY_DN42372 | −1.6 | 9 × 10−4 | |||
| PSBQ1 | A. thaliana | Oxygen-evolving enhancer protein 3-1 | TRINITY_DN17369 | −1.6 | 3 × 10−6 |
| Photosystem I reaction centre (GO:0009538), photosystem I (GO:0009522), chloroplast thylakoid membrane (GO:0009535) | |||||
| PSAD | S. oleracea | Photosystem I reaction centre subunit II | TRINITY_DN59085 | −1.1 | 6 × 10−4 |
| PSAH | O. sativa | Photosystem I reaction centre subunit VI | TRINITY_DN14879 | −1.6 | 4 × 10−6 |
| S. oleracea | TRINITY_DN53439 | −1.4 | 6 × 10−5 | ||
| Chloroplast thylakoid membrane (GO:0009535) | |||||
| PTAC5 | A. thaliana | Protein disulphide isomerase pTAC5 | TRINITY_DN27918 | −1.3 | 1 × 10−19 |
| PTA16 | A. thaliana | Protein plastid transcriptionally active 16 | TRINITY_DN37868 | −1.3 | 8 × 10−8 |
| CHL | A. thaliana | Chloroplastic lipocalin | TRINITY_DN16390 | −1.1 | 3 × 10−8 |
| CG160 | A. thaliana | Protein conserved in the green lineage 160 | TRINITY_DN24793 | −2.2 | 3 × 10−19 |
| CAO | A. thaliana | Chlorophyllide a oxygenase | TRINITY_DN21455 | −1.7 | 6 × 10−5 |
| TRINITY_DN31549 | −2.5 | 3 × 10−4 | |||
| O. sativa | TRINITY_DN84817 | −2.5 | 3 × 10−7 | ||
| DNJA6 | A. thaliana | Chaperone protein dnaJ A6 | TRINITY_DN32102 | −1.1 | 2 × 10−14 |
| TL29 | A. thaliana | Thylakoid lumenal 29 kDa protein | TRINITY_DN35144 | −1.7 | 3 × 10−9 |
| STR4 | A. thaliana | Rhodanese-like domain-containing protein 4 | TRINITY_DN20986 | −2.5 | 3 × 10−4 |
| STR9 | A. thaliana | Rhodanese-like domain-containing protein 9 | TRINITY_DN59649 | −2.5 | 3 × 10−7 |
| ABA2 | S. oleracea | Zeaxanthin epoxidase | TRINITY_DN87513 | −1.8 | 2 × 10−5 |
| CUT1A | A. thaliana | Protein curvature thylakoid 1A | TRINITY_DN32471 | −1.3 | 6 × 10−4 |
| CRR3 | A. thaliana | Probable NAD(P)H dehydrogenase subunit CRR3 | TRINITY_DN50182 | −1.2 | 1 × 10−8 |
| NDF5 | A. thaliana | Protein NDH-dependent cyclic electron flow 5 | TRINITY_DN61095 | −2.1 | 4 × 10−22 |
| NDHK | E. globus | NAD(P)H-quinone oxidoreductase | TRINITY_DN6819 | −1.2 | 2 × 10−13 |
| Protochlorophyllide reductase activity (GO:0016630) | |||||
| PORA | A. thaliana | Protochlorophyllide reductase A | TRINITY_DN13370 | −2.7 | 4 × 10−7 |
| C. sativus | Protochlorophyllide reductase | TRINITY_DN26265 | −1.7 | 7 × 10−4 | |
| TRINITY_DN68683 | −1.7 | 2 × 10−12 | |||
| PORB | H. vulgare | Protochlorophyllide reductase B | TRINITY_DN40740 | −1.7 | 4 × 10−10 |
| Starch binding (GO:2001070) | |||||
| DPE2 | A. thaliana | 4-alpha-glucanotransferase DPE2 | TRINITY_DN29808 | −3.3 | 1 × 10−7 |
| DSP4 | C. sativa | Phosphoglucan phosphatase DSP4, amyloplastic | TRINITY_DN33943 | −4.3 | 1 × 10−20 |
| Plant-type vacuole (GO:0000325) | |||||
| PTR2 | A. thaliana | Protein NRT1/ PTR family 8.3 | TRINITY_DN38391 | −2.0 | 2 × 10−8 |
| CAX3 | A. thaliana | Vacuolar cation/proton exchanger 3 | TRINITY_DN38001 | −1.1 | 9 × 10−15 |
| TRINITY_DN40895 | −1.5 | 3 × 10−11 | |||
| TPC1 | A. thaliana | Two pore calcium channel protein 1 | TRINITY_DN6810 | −1.5 | 4 × 10−6 |
| TIP11 | A. thaliana | Aquaporin TIP1-1 | TRINITY_DN35041 | −1.3 | 5 × 10−5 |
| TIP21 | A. thaliana | Aquaporin TIP2-1 | TRINITY_DN40936 | −3.4 | 1 × 10−14 |
| CEP1 | A. thaliana | KDEL-tailed cysteine endopeptidase CEP1 | TRINITY_DN11687 | −2.2 | 3 × 10−33 |
| TRINITY_DN28528 | −1.9 | 2 × 10−7 | |||
| TRINITY_DN29270 | −2.0 | 3 × 10−25 | |||
| AA5GT | D. caryophyllus | Cyanidin 3-O-glucoside 5-O-glucosyltransferase (acyl-glucose) | TRINITY_DN39241 | −3.5 | 3 × 10−9 |
| TRINITY_DN39972 | −2.4 | 1 × 10−8 | |||
| AB3C | A. thaliana | ABC transporter C family member 3 | TRINITY_DN40021 | −1.1 | 7 × 10−5 |
| AB8C | A. thaliana | ABC transporter C family member 8 | TRINITY_DN40157 | −5.1 | 6 × 10−49 |
| NRT25 | A. thaliana | High affinity nitrate transporter 2.5 | TRINITY_DN77992 | −2.1 | 1 × 10−4 |
| ALMTC | A. thaliana | Aluminium-activated malate transporter 12 | TRINITY_DN33933 | −1.4 | 3 × 10−21 |
| ALMT2 | A. thaliana | Aluminium-activated malate transporter 2 | TRINITY_DN21972 | −1.4 | 4 × 10−6 |
| ERDL6 | A. thaliana | Sugar transporter ERD6-like 6 | TRINITY_DN56467 | −2.1 | 1 × 10−3 |
| NCL | A. thaliana | Sodium/calcium exchanger NCL | TRINITY_DN63539 | −3.3 | 1 × 10−12 |
| RNHX1 | A. thaliana | Putative ribonuclease H protein At1g65750 | TRINITY_DN15507 | −3.0 | 1 × 10−10 |
| CYSEP | V. mungo | Vignain | TRINITY_DN33759 | −1.9 | 2 × 10−6 |
| OCT3 | A. thaliana | Organic cation/carnitine transporter 3 | TRINITY_DN35062 | −3.9 | 8 × 10−16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gieniec, M.; Miszalski, Z.; Rozpądek, P.; Jędrzejczyk, R.J.; Czernicka, M.; Nosek, M. How the Ethylene Biosynthesis Pathway of Semi-Halophytes Is Modified with Prolonged Salinity Stress Occurrence? Int. J. Mol. Sci. 2024, 25, 4777. https://doi.org/10.3390/ijms25094777
Gieniec M, Miszalski Z, Rozpądek P, Jędrzejczyk RJ, Czernicka M, Nosek M. How the Ethylene Biosynthesis Pathway of Semi-Halophytes Is Modified with Prolonged Salinity Stress Occurrence? International Journal of Molecular Sciences. 2024; 25(9):4777. https://doi.org/10.3390/ijms25094777
Chicago/Turabian StyleGieniec, Miron, Zbigniew Miszalski, Piotr Rozpądek, Roman J. Jędrzejczyk, Małgorzata Czernicka, and Michał Nosek. 2024. "How the Ethylene Biosynthesis Pathway of Semi-Halophytes Is Modified with Prolonged Salinity Stress Occurrence?" International Journal of Molecular Sciences 25, no. 9: 4777. https://doi.org/10.3390/ijms25094777
APA StyleGieniec, M., Miszalski, Z., Rozpądek, P., Jędrzejczyk, R. J., Czernicka, M., & Nosek, M. (2024). How the Ethylene Biosynthesis Pathway of Semi-Halophytes Is Modified with Prolonged Salinity Stress Occurrence? International Journal of Molecular Sciences, 25(9), 4777. https://doi.org/10.3390/ijms25094777