
Examination of the Complex Molecular Landscape in Obesity and Type 2 Diabetes



Abstract
:1. Introduction
2. Results
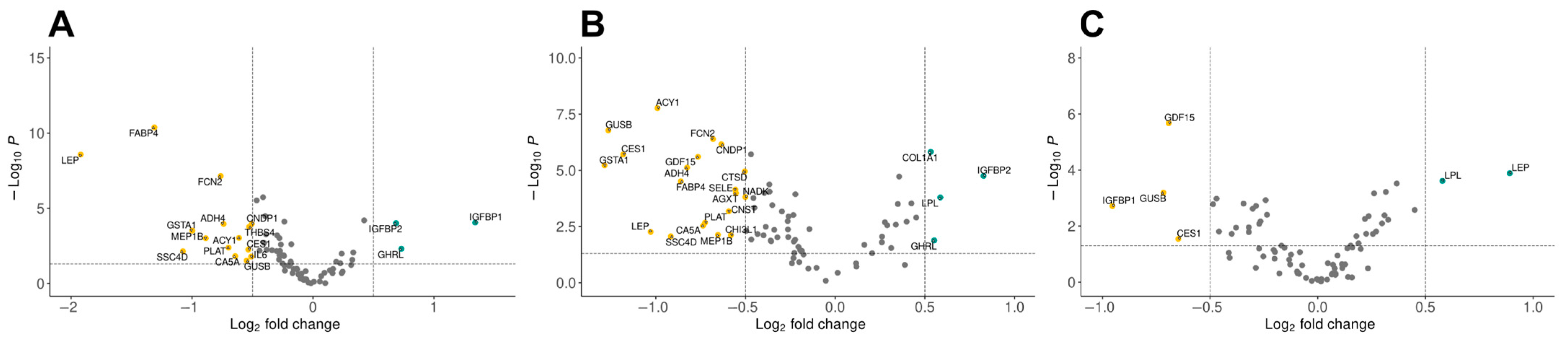
2.1. Differential Expression Analysis Reveals Distinct Protein Profiles in Obesity and Type 2 Diabetes
2.2. Differentially Expressed Proteins Are Responsible for Various Functions
2.3. Olink Data Correlates with Metabolomics and Clinical Data
2.4. Cardiometabolic Proteins Interact with Each Other
3. Discussion
4. Materials and Methods
4.1. Study Subjects and Sample Collection
4.2. Analysis of Cardiometabolic Proteins with Proximity Extension Assay
4.3. Data Analysis
4.3.1. Pathway Enrichment Analysis
4.3.2. Statistical Analysis
4.3.3. Correlation Analysis
4.3.4. Network Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kopelman, P.G. Obesity as a Medical Problem. Nature 2000, 404, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Stumvoll, M.; Goldstein, B.J.; Van Haeften, T.W. Type 2 Diabetes: Principles of Pathogenesis and Therapy. Lancet 2005, 365, 1333–1346. [Google Scholar] [CrossRef]
- Guh, D.P.; Zhang, W.; Bansback, N.; Amarsi, Z.; Birmingham, C.L.; Anis, A.H. The Incidence of Co-Morbidities Related to Obesity and Overweight: A Systematic Review and Meta-Analysis. BMC Public Health 2009, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Dettmer, K.; Aronov, P.A.; Hammock, B.D. Mass Spectrometry-based Metabolomics. Mass Spectrom. Rev. 2007, 26, 51–78. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O. Metabolomics—The Link between Genotypes and Phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Saccenti, E.; Hoefsloot, H.C.J.; Smilde, A.K.; Westerhuis, J.A.; Hendriks, M.M.W.B. Reflections on Univariate and Multivariate Analysis of Metabolomics Data. Metabolomics 2014, 10, 361–374. [Google Scholar] [CrossRef]
- Wik, L.; Nordberg, N.; Broberg, J.; Björkesten, J.; Assarsson, E.; Henriksson, S.; Grundberg, I.; Pettersson, E.; Westerberg, C.; Liljeroth, E.; et al. Proximity Extension Assay in Combination with Next-Generation Sequencing for High-Throughput Proteome-Wide Analysis. Mol. Cell. Proteom. 2021, 20, 100168. [Google Scholar] [CrossRef]
- Vavruch, C.; Nowak, C.; Feldreich, T.; Östgren, C.J.; Sundström, J.; Söderberg, S.; Lind, L.; Nyström, F.; Ärnlöv, J. Using Proximity Extension Proteomics Assay to Discover Novel Biomarkers Associated with Circulating Leptin Levels in Patients with Type 2 Diabetes. Sci. Rep. 2020, 10, 13097. [Google Scholar] [CrossRef]
- Kozakova, M.; Morizzo, C.; Goncalves, I.; Natali, A.; Nilsson, J.; Palombo, C. Cardiovascular Organ Damage in Type 2 Diabetes Mellitus: The Role of Lipids and Inflammation. Cardiovasc. Diabetol. 2019, 18, 61. [Google Scholar] [CrossRef]
- Assarsson, E.; Lundberg, M.; Holmquist, G.; Björkesten, J.; Bucht Thorsen, S.; Ekman, D.; Eriksson, A.; Rennel Dickens, E.; Ohlsson, S.; Edfeldt, G.; et al. Homogenous 96-Plex PEA Immunoassay Exhibiting High Sensitivity, Specificity, and Excellent Scalability. PLoS ONE 2014, 9, e95192. [Google Scholar] [CrossRef]
- Nokhoijav, E.; Guba, A.; Kumar, A.; Kunkli, B.; Kalló, G.; Káplár, M.; Somodi, S.; Garai, I.; Csutak, A.; Tóth, N.; et al. Metabolomic Analysis of Serum and Tear Samples from Patients with Obesity and Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2022, 23, 4534. [Google Scholar] [CrossRef]
- Franks, P.W.; Pearson, E.; Florez, J.C. Gene-Environment and Gene-Treatment Interactions in Type 2 Diabetes. Diabetes Care 2013, 36, 1413–1421. [Google Scholar] [CrossRef] [PubMed]
- La Sala, L.; Pontiroli, A.E. Prevention of Diabetes and Cardiovascular Disease in Obesity. Int. J. Mol. Sci. 2020, 21, 8178. [Google Scholar] [CrossRef] [PubMed]
- Geyer, P.E.; Holdt, L.M.; Teupser, D.; Mann, M. Revisiting Biomarker Discovery by Plasma Proteomics. Mol. Syst. Biol. 2017, 13, 942. [Google Scholar] [CrossRef]
- Cronjé, H.T.; Mi, M.Y.; Austin, T.R.; Biggs, M.L.; Siscovick, D.S.; Lemaitre, R.N.; Psaty, B.M.; Tracy, R.P.; Djoussé, L.; Kizer, J.R.; et al. Plasma Proteomic Risk Markers of Incident Type 2 Diabetes Reflect Physiologically Distinct Components of Glucose-Insulin Homeostasis. Diabetes 2023, 72, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, M.; Lewis, G.D.; Ericson, U.; Orho-Melander, M.; Hedblad, B.; Engström, G.; Östling, G.; Clish, C.; Wang, T.J.; Gerszten, R.E.; et al. A Diabetes-Predictive Amino Acid Score and Future Cardiovascular Disease. Eur. Heart J. 2013, 34, 1982–1989. [Google Scholar] [CrossRef]
- Yeung, D.C.Y.; Xu, A.; Cheung, C.W.S.; Wat, N.M.S.; Yau, M.H.; Fong, C.H.Y.; Chau, M.T.; Lam, K.S.L. Serum Adipocyte Fatty Acid-Binding Protein Levels Were Independently Associated with Carotid Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1796–1802. [Google Scholar] [CrossRef]
- Sookoian, S.; Castaño, G.O.; Scian, R.; Fernández Gianotti, T.; Dopazo, H.; Rohr, C.; Gaj, G.; San Martino, J.; Sevic, I.; Flichman, D.; et al. Serum Aminotransferases in Nonalcoholic Fatty Liver Disease Are a Signature of Liver Metabolic Perturbations at the Amino Acid and Krebs Cycle Level. Am. J. Clin. Nutr. 2016, 103, 422–434. [Google Scholar] [CrossRef]
- Geyer, P.E.; Kulak, N.A.; Pichler, G.; Holdt, L.M.; Teupser, D.; Mann, M. Plasma Proteome Profiling to Assess Human Health and Disease. Cell Syst. 2016, 2, 185–195. [Google Scholar] [CrossRef]
- Kotronen, A.; Peltonen, M.; Hakkarainen, A.; Sevastianova, K.; Bergholm, R.; Johansson, L.M.; Lundbom, N.; Rissanen, A.; Ridderstråle, M.; Groop, L.; et al. Prediction of Non-Alcoholic Fatty Liver Disease and Liver Fat Using Metabolic and Genetic Factors. Gastroenterology 2009, 137, 865–872. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The Human Metabolome Database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef] [PubMed]
- Floegel, A.; Stefan, N.; Yu, Z.; Mühlenbruch, K.; Drogan, D.; Joost, H.-G.; Fritsche, A.; Häring, H.-U.; Hrabě De Angelis, M.; Peters, A.; et al. Identification of Serum Metabolites Associated with Risk of Type 2 Diabetes Using a Targeted Metabolomic Approach. Diabetes 2013, 62, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite Profiles and the Risk of Developing Diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Rhee, E.P.; Larson, M.G.; Lewis, G.D.; McCabe, E.L.; Shen, D.; Palma, M.J.; Roberts, L.D.; Dejam, A.; Souza, A.L.; et al. Metabolite Profiling Identifies Pathways Associated with Metabolic Risk in Humans. Circulation 2012, 125, 2222–2231. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter 1 | Control | Obesity | Type 2 Diabetes |
|---|---|---|---|
| (N = 32) | (N = 44) | (N = 54) | |
| Age, years | 49.25 ± 10.13 | 51.93 ± 9.61 | 50.72 ± 7.94 |
| Sex (number) | Male (19) | Male (20) | Male (32) |
| Female (13) | Female (24) | Female (22) | |
| Body mass index (BMI) | 25.33 ± 2.08 | 37.8 ± 5.82 | 33.78 ± 5.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vadadokhau, U.; Varga, I.; Káplár, M.; Emri, M.; Csősz, É. Examination of the Complex Molecular Landscape in Obesity and Type 2 Diabetes. Int. J. Mol. Sci. 2024, 25, 4781. https://doi.org/10.3390/ijms25094781
Vadadokhau U, Varga I, Káplár M, Emri M, Csősz É. Examination of the Complex Molecular Landscape in Obesity and Type 2 Diabetes. International Journal of Molecular Sciences. 2024; 25(9):4781. https://doi.org/10.3390/ijms25094781
Chicago/Turabian StyleVadadokhau, Uladzislau, Imre Varga, Miklós Káplár, Miklós Emri, and Éva Csősz. 2024. "Examination of the Complex Molecular Landscape in Obesity and Type 2 Diabetes" International Journal of Molecular Sciences 25, no. 9: 4781. https://doi.org/10.3390/ijms25094781
APA StyleVadadokhau, U., Varga, I., Káplár, M., Emri, M., & Csősz, É. (2024). Examination of the Complex Molecular Landscape in Obesity and Type 2 Diabetes. International Journal of Molecular Sciences, 25(9), 4781. https://doi.org/10.3390/ijms25094781