
Human Thalamic Somatosensory Nucleus (Ventral Caudal, Vc) as a Locus for Stimulation by INPUTS from Tactile, Noxious and Thermal Sensors on an Active Prosthesis

 ,
, 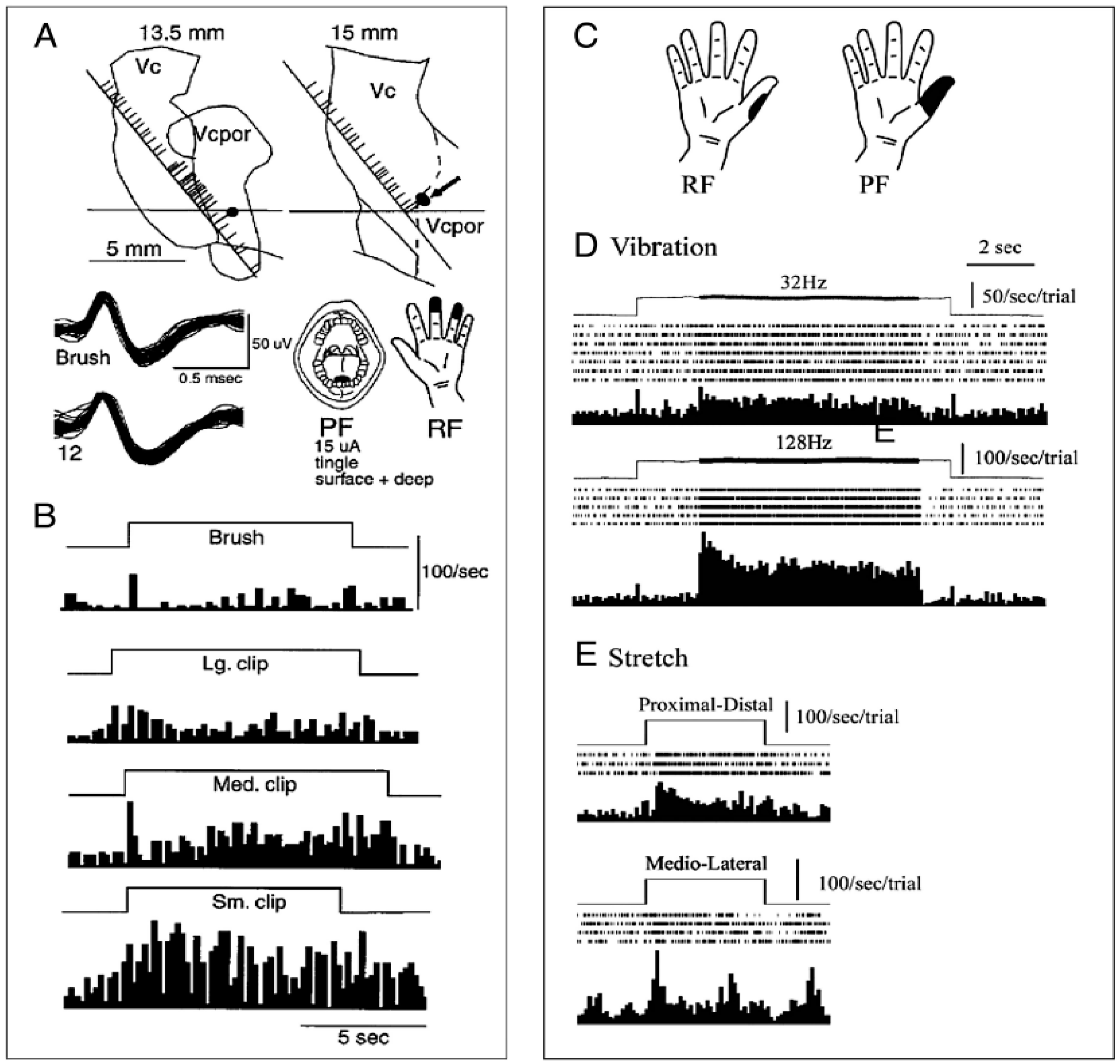{kind=link}
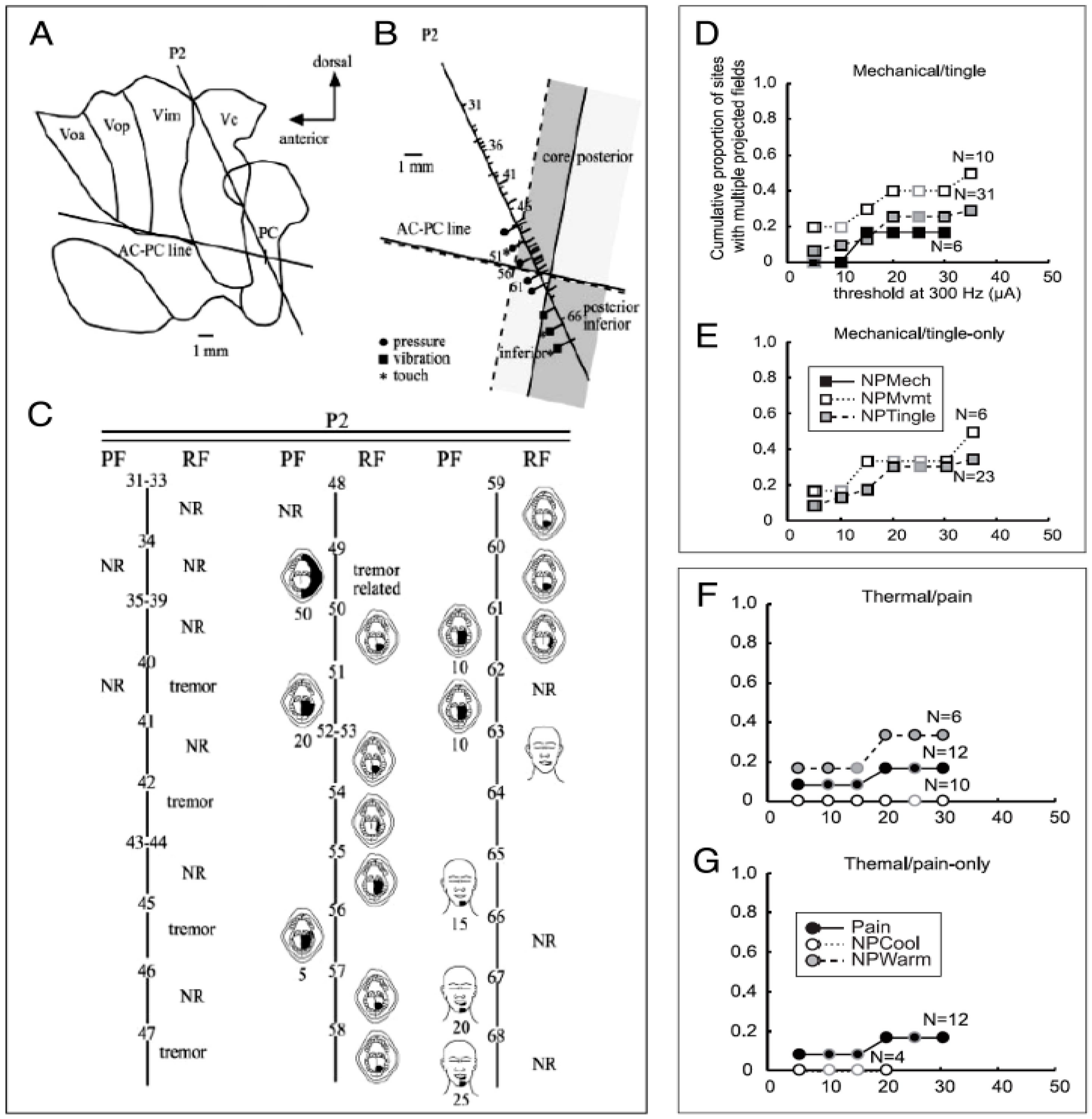{kind=link}
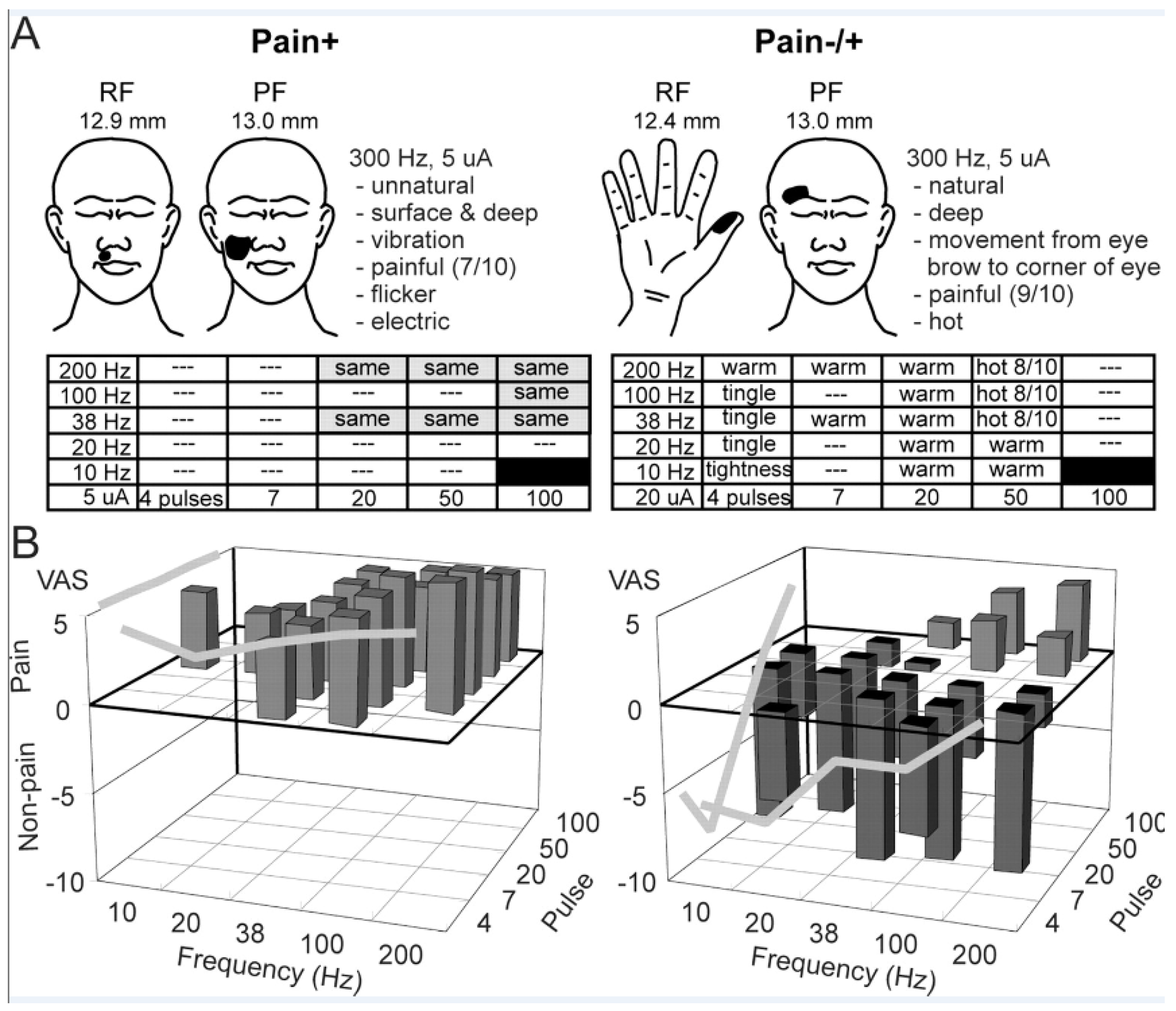{kind=link}
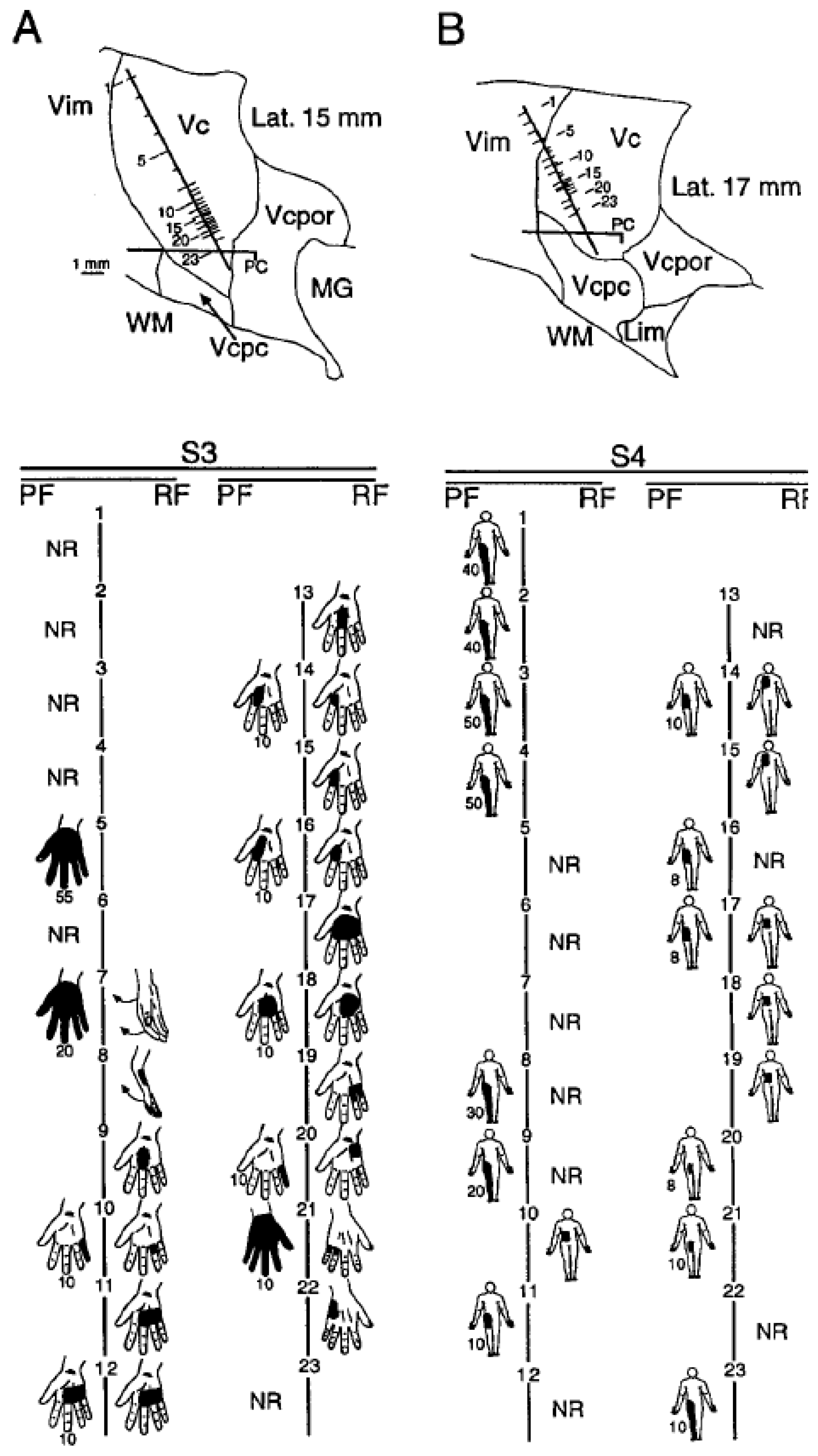{kind=link}
Abstract
:1. Introduction
2. Neuronal Responses and Microstimulation Evoked Sensation Related to Cutaneous Stimuli
3. Thalamic Vc Activity Related to Thermal or Painful Stimuli
4. Segmentation by Modality and Somatotopy
5. Patterned TMIS Should Encode Discrete Natural Sensations
6. Somatotopy of RFs and PFs Following Amputation or Spinal Cord Transection
7. Thalamic Function in Primates with Major Injuries to the Somatic Sensory System
8. Implications for Forebrain Loci for TMIS by Signals from Sensors
9. Segmentation within Vc
10. Stability of Thalamic Representations after a Major Injury
11. Pain Evoked by TMIS of Thalamus or Cortex
12. Conclusions
Acknowledgments
Conflicts of Interest
References
- Sinclair, R.J.; Burton, H. Neuronal activity in the primary somatosensory cortex in monkeys (Macaca mulatta) during active touch of textured surface gratings: Responses to groove width, applied force, and velocity of motion. J. Neurophysiol. 1991, 66, 153–169. [Google Scholar] [PubMed]
- Vega-Bermudez, F.; Johnson, K.O.; Hsiao, S.S. Human tactile pattern recognition: Active versus passive touch, velocity effects, and patterns of confusion. J. Neurophysiol. 1991, 65, 531–546. [Google Scholar] [PubMed]
- Iriki, A.; Tanaka, M.; Iwamura, Y. Coding of modified body schema during tool use by macaque postcentral neurons. Neuroreport 1996, 7, 2325–2330. [Google Scholar] [PubMed]
- Farne, A.; Iriki, A.; Ladavas, E. Shaping multisensory action-space with tools: Evidence from patients with cross-modal extinction. Neuropsychologia 2005, 43, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Higgins, S.T.; Morris, E.K. Generality of free-operant avoidance conditioning to human behavior. Psychol. Bull. 1984, 96, 247–272. [Google Scholar] [CrossRef] [PubMed]
- Torebjork, E.; Vallbo, A.B.; Ochoa, J. Intraneural Microstimulation in Man: Its relation to specificity of tactile sensations. Brain 1987, 110, 1509–1529. [Google Scholar] [CrossRef] [PubMed]
- Lenz, F.A.; Casey, K.L.; Jones, E.G.; Willis, W.D., Jr. The Human Pain System: Experimental and Clinical Perspectives; Cambridge University Press: New York, NY, USA, 2010; pp. 1–797. [Google Scholar]
- Vlaeyen, J.W.; Crombez, G.; Linton, S.J. The fear-avoidance model of pain. Pain 2016, 157, 1588–1589. [Google Scholar] [CrossRef] [PubMed]
- Bensmaia, S.J. Biological and bionic hands: Natural neural coding and artificial perception. Philos. Trans. R. Soc. Lond. B 2015, 370, 20140209. [Google Scholar] [CrossRef] [PubMed]
- Delhaye, B.P.; Saal, H.P.; Bensmaia, S.J. Key considerations in designing a somatosensory neuroprosthesis. J. Physiol. Paris 2016. [Google Scholar] [CrossRef] [PubMed]
- Ackerley, R.; Kavounoudias, A. The role of tactile afference in shaping motor behaviour and implications for prosthetic innovation. Neuropsychologia 2015, 79, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Wurtz, R.H.; Sommer, M.A. Identifying corollary discharges for movement in the primate brain. Prog. Brain Res. 2004, 144, 47–60. [Google Scholar] [PubMed]
- Sommer, M.A.; Wurtz, R.H. Influence of the thalamus on spatial visual processing in frontal cortex. Nature 2006, 444, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Anderson, W.S.; Weiss, N.; Lawson, H.C.; Ohara, S.; Rowland, L.; Lenz, F.A. Demonstration of motor imagery movement and phantom movement-related neuronal activity in human thalamus. Neuroreport 2011, 22, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Anderson, W.S.; Lenz, F.A. Review of motor and phantom-related imagery. Neuroreport 2011, 22, 939–942. [Google Scholar] [CrossRef] [PubMed]
- Mulvey, M.R.; Fawkner, H.J.; Radford, H.E.; Johnson, M.I. Perceptual embodiment of prosthetic limbs by transcutaneous electrical nerve stimulation. Neuromodulation 2012, 15, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Mabuchi, K. An artificial arm/hand system with a haptic sensory function using electric stimulation of peripheral sensory nerve fibers. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2013, 2013, 3130–3133. [Google Scholar] [PubMed]
- Alam, M.; Rodrigues, W.; Pham, B.N.; Thakor, N.V. Brain-machine interface facilitated neurorehabilitation via spinal stimulation after spinal cord injury: Recent progress and future perspectives. Brain Res. 2016, 1646, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Semework, M. Microstimulation: Principles, Techniques, and Approaches to Somatosensory Neuroprosthesis. Crit Rev. Biomed. Eng. 2015, 43, 61–95. [Google Scholar] [CrossRef] [PubMed]
- Heming, E.A.; Choo, R.; Davies, J.N.; Kiss, Z.H. Designing a thalamic somatosensory neural prosthesis: Consistency and persistence of percepts evoked by electrical stimulation. IEEE Trans. Neural Syst. Rehabil. Eng. 2011, 19, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Hirai, T.; Jones, E.G. A new parcellation of the human thalamus on the basis of histochemical staining. Brain Res. Rev. 1989, 14, 1–34. [Google Scholar] [CrossRef]
- Schaltenbrand, G.; Wahren, W. An Atlas for Stereotaxy of the Human Brain; Thieme-Stratton: New York, NY, USA, 1982; pp. 1–99. [Google Scholar]
- Holsapple, J.W.; Preston, J.B.; Strick, P.L. The origin of thalamic inputs to the “hand” representation in the primary motor cortex. J. Neurosci. 1991, 11, 2644–2654. [Google Scholar] [PubMed]
- Lenz, F.A.; Kwan, H.C.; Dostrovsky, J.O.; Tasker, R.R.; Murphy, J.T.; Lenz, Y.E. Single unit analysis of the human ventral thalamic nuclear group: Activity correlated with movement. Brain 1990, 113, 1795–1821. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Kim, J.H.; Anderson, W.S.; Lenz, F.A. Neurosurgical treatment of tremor. In Youman’s Neurological Surgery; Winn, R., Ed.; Saunders: Philadelphia, PA, USA, 2009; pp. 932–937. [Google Scholar]
- Lenz, F.A. Ablative Procedures. In Handbook of Neurology: Section on Parkinson’s Disease and Related Disorders; Kller, W.C., Melamed, E., Eds.; Elsevier: Amsterdam, The Netherlands, 2006; pp. 241–260. [Google Scholar]
- Lenz, F.A. Neurosurgical Treatment of Pain. In Handbook of Neurology: Section on Pain; Jensen, T.S., Cervero, F., Eds.; Elsevier: Amsterdam, The Netherlands, 2006; pp. 869–886. [Google Scholar]
- Benabid, A.L.; Wallace, B.; Mitrofanis, J.; Xia, C.; Piallat, B.; Fraix, V.; Batir, A.; Krack, P.; Pollak, P.; Berger, F. Therapeutic electrical stimulation of the central nervous system. C. R. Biol. 2005, 328, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Cruccu, G.; Aziz, T.Z.; Garcia-Larrea, L.; Hansson, P.; Jensen, T.S.; Lefaucheur, J.P.; Simpson, B.A.; Taylor, R.S. EFNS guidelines on neurostimulation therapy for neuropathic pain. Eur. J. Neurol. 2007, 14, 952–970. [Google Scholar] [CrossRef] [PubMed]
- Hopfner, F.; Haubenberger, D.; Galpern, W.R.; Gwinn, K.; Van’t Veer, A.; White, S.; Bhatia, K.; Adler, C.H.; Eidelberg, D.; Ondo, W.; et al. Knowledge gaps and research recommendations for essential tremor. Parkinsonism. Relat. Disord. 2016, 33, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Elble, R.J. Essential tremor is a monosymptomatic disorder. Mov. Disord. 2002, 17, 633–637. [Google Scholar] [CrossRef] [PubMed]
- Lenz, F.A.; Dostrovsky, J.O.; Kwan, H.C.; Tasker, R.R.; Yamashiro, K.; Murphy, J.T. Methods for microstimulation and recording of single neurons and evoked potentials in the human central nervous system. J. Neurosurg. 1988, 68, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Greenspan, J.D.; Coghill, R.H.; Ohara, S.; Lenz, F.A. Lesions limited to the human thalamic principal somatosensory nucleus (ventral caudal) are associated with loss of cold sensations and central pain. J. Neurosci. 2007, 27, 4995–5004. [Google Scholar] [CrossRef] [PubMed]
- Lenz, F.A.; Byl, N.N. Reorganization in the cutaneous core of the human thalamic principal somatic sensory nucleus (Ventral caudal) in patients with dystonia. J. Neurophysiol. 1999, 82, 3204–3212. [Google Scholar] [PubMed]
- Lenz, F.A.; Dostrovsky, J.O.; Tasker, R.R.; Yamashiro, K.; Kwan, H.C.; Murphy, J.T. Single-unit analysis of the human ventral thalamic nuclear group: Somatosensory responses. J. Neurophysiol. 1988, 59, 299–316. [Google Scholar] [PubMed]
- Lenz, F.A.; Kwan, H.C.; Martin, R.; Tasker, R.R.; Richardson, R.T.; Dostrovsky, J.O. Characteristics of somatotopic organization and spontaneous neuronal activity in the region of the thalamic principal sensory nucleus in patients with spinal cord transection. J. Neurophysiol. 1994, 72, 1570–1587. [Google Scholar] [PubMed]
- Ohara, S.; Weiss, N.; Lenz, F.A. Microstimulation in the region of the human thalamic principal somatic sensory nucleus evokes sensations like those of mechanical stimulation and movement. J. Neurophysiol. 2004, 91, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Weiss, N.; Ohara, S.; Johnson, K.O.; Lenz, F.A. The human thalamic somatic sensory nucleus [ventral caudal (Vc)] shows neuronal mechanoreceptor-like responses to optimal stimuli for peripheral mechanoreceptors. J. Neurophysiol. 2009, 101, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Schmid, A.C.; Chien, J.H.; Greenspan, J.D.; Garonzik, I.; Weiss, N.; Ohara, S.; Lenz, F.A. Neuronal Responses to Tactile Stimuli and Tactile Sensations evoked by Microstimulation in the Human Thalamic Principle Somatic Sensory Nucleus - Ventral Caudal. J. Neurophysiol. 2016, 215, 2421–2433. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.I.; Ohara, S.; Dougherty, P.M.; Lenz, F.A. Pain and temperature encoding in the human thalamic somatic sensory nucleus (Ventral caudal): Inhibition-related bursting evoked by somatic stimuli. J. Neurophysiol. 2005, 94, 1676–1687. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.I.; Antezanna, D.; Dougherty, P.M.; Lenz, F.A. Responses of neurons in the region of the thalamic somatosensory nucleus to mechanical and thermal stimuli graded into the painful range. J. Comp. Neurol. 1999, 410, 541–555. [Google Scholar] [CrossRef]
- Johnson, K.O.; Yoshioka, T.; Vega-Bermudez, F. Tactile functions of mechanoreceptive afferents innervating the hand. J. Clin. Neurophysiol. 2000, 17, 539–558. [Google Scholar] [CrossRef] [PubMed]
- Johansson, R.S.; Vallbo, A.B. Skin mechanoreceptors in the human hand: An inference of some population properties. In Sensory Functions of the Skin in Primates; Zotterman, Y., Ed.; Pergamon Press: Oxford, UK, 1976; pp. 171–184. [Google Scholar]
- Johansson, R.S.; Landstrom, U.; Lundstrom, R. Responses of mechanoreceptive afferent units in the glabrous skin of the human hand to sinusoidal skin displacements. Brain Res. 1982, 244, 17–25. [Google Scholar] [CrossRef]
- Zhang, H.Q.; Murray, G.M.; Coleman, G.M.; Turman, A.B.; Zhang, S.P.; Rowe, M.J. Functional characteristics of the parallel SI- and SII-projecting neurons of the thalamic ventral posterior nucleus in the marmoset. J. Neurophysiol. 2001, 85, 1805–1822. [Google Scholar] [PubMed]
- Warren, S.; Kelahan, A.M.; Pubols, B.H. The somatosensory thalamus of the raccoon: Properties of single neurons responsive to light mechanical stimulation in the forepaw. J. Neurosci. 1986, 6, 308–317. [Google Scholar] [PubMed]
- Willis, W.D.; Trevino, D.L.; Coulter, J.D.; Maunz, R.A. Responses of primate spinothalamic tract neurons to natural stimulation of hindlimb. J. Neurophysiol. 1974, 37, 358–372. [Google Scholar] [PubMed]
- Willis, W.D. The Pain System; Karger: Basel, Switzerland, 1985. [Google Scholar]
- Bushnell, M.C.; Duncan, G.H.; Tremblay, N. Thalamic VPM nucleus in the behaving monkey. I. Multimodal and discriminative properties of thermosensitive neurons. J. Neurophysiol. 1993, 69, 739–752. [Google Scholar] [PubMed]
- Chung, J.M.; Surmeier, D.J.; Lee, K.H.; Sorkin, L.; Honda, N.; Tsong, Y.; Willis, W.D. Classification of primate spinothalamic and somatosensory thalamic neurons based on cluster analysis. J. Neurophysiol. 1986, 56, 308–327. [Google Scholar] [PubMed]
- Adriaensen, H.; Gybels, J.; Handwerker, H.O.; Van Hees, J. Nociceptor discharges and sensations due to prolonged noxious mechanical stimulation—A paradox. Hum. Neurobiol. 1984, 3, 53–58. [Google Scholar] [CrossRef]
- Ferrington, D.G.; Downie, J.W.; Willis, W.D. Primate nucleus gracilis neurons: Responses to innocuous and noxious stimuli. J. Neurophysiol. 1988, 59, 886–907. [Google Scholar] [PubMed]
- Jones, E.G.; Friedman, D.P.; Hendry, S.H. Thalamic basis of place- and modality-specific columns in monkey somatosensory cortex: A correlative anatomical and physiological study. J. Neurophysiol. 1982, 48, 545–568. [Google Scholar] [PubMed]
- Kaas, J.H.; Nelson, R.J.; Sur, M.; Dykes, R.W.; Merzenich, M.M. The somatotopic organization of the ventroposterior thalamus of the squirrel monkey, Saimiri sciureus. J. Comp. Neurol. 1984, 226, 111–140. [Google Scholar] [CrossRef] [PubMed]
- Rausell, E.; Bae, C.S.; Vinuela, A.; Huntley, G.W.; Jones, E.G. Calbindin and parvalbumin cells in monkey VPL thalamic nucleus: Distribution, laminar cortical projections, and relations to spinothalamic terminations. J. Neurosci. 1992, 12, 4088–4111. [Google Scholar] [PubMed]
- Hirai, T.; Schwark, H.D.; Yen, C.T.; Honda, C.N.; Jones, E.G. Morphology of Physiologically Characterized Medial Lemniscal Axons Terminating in Cat Ventral Posterior Thalamic Nucleus. J. Neurophysiol. 1988, 60, 1439–1459. [Google Scholar] [PubMed]
- Rausell, E.; Jones, E.G. Chemically distinct compartments of the thalamic VPM nucleus in monkeys relay principal and spinal trigeminal pathways to different layers of the somatosensory cortex. J. Neurosci. 1991, 11, 226–237. [Google Scholar] [PubMed]
- Tremblay, N.; Bushnell, M.C.; Duncan, G.H. Thalamic VPM nucleus in the behaving monkey: II. response to air puff stimulation during discrimination and attention tasks. J. Neurophysiol. 1993, 69, 753–763. [Google Scholar] [PubMed]
- Morrow, T.J.; Casey, K.L. State-related modulation of thalamic somatosensory responses in the awake monkey. J. Neurophysiol. 1992, 67, 305–317. [Google Scholar] [PubMed]
- Landry, P.; Deschenes, M. Intracortical arborizations and receptive fields of identified ventrobasal thalamocortical afferents to the primary somatic sensory cortex in the cat. J. Comp. Neurol. 1981, 199, 345–371. [Google Scholar] [CrossRef] [PubMed]
- Ranck, J.B. Which elements are excited in electrical stimulation of mammalian central nervous system: A review. Brain Res. 1981, 98, 417–440. [Google Scholar] [CrossRef]
- Mehler, W.R. The anatomy of the so-called “pain tract” in man: An analysis of the course and distribution of the ascending fibers of the fasciculus anterolateralis. In Basic Research in Paraplegia; French, J.D., Porter, R.W., Eds.; Thomas: Springfield, IL, USA, 1962; pp. 26–55. [Google Scholar]
- Mehler, W.R.; Feferman, M.E.; Nauta, W.H.J. Ascending axon degeneration following anterolateral cordotomy. An experimental study in the monkey. Brain 1960, 83, 718–750. [Google Scholar] [CrossRef] [PubMed]
- Apkarian, A.V.; Shi, T. Squirrel monkey lateral thalamus. I. somatic nociresponsive neurons and their relation to spinothalamic terminals. J. Neurosci. 1994, 14, 6779–6795. [Google Scholar] [PubMed]
- Willis, W.D.; Zhang, X.; Honda, C.N.; Giesler, G.J. Projections from the marginal zone and deep dorsal horn to the ventrobasal nuclei of the primate thalamus. Pain 2001, 92, 267–276. [Google Scholar] [CrossRef]
- Casey, K.L.; Morrow, T.J. Nociceptive neurons in the ventral posterior thalamus of the awake squirrel monkey: Observations on identification, modulation, and drug effects. In Thalamus and Pain; Besson, J.-M., Guilbaud, G., Peschanski, M., Eds.; Elsevier Science Publishers B.V.: New York, NY, USA, 1987; pp. 211–257. [Google Scholar]
- Davis, K.D.; Lozano, A.M.; Manduch, M.; Tasker, R.R.; Kiss, Z.H.; Dostrovsky, J.O. Thalamic relay site for cold perception in humans. J. Neurophysiol. 1999, 81, 1970–1973. [Google Scholar] [PubMed]
- Ohara, S.; Lenz, F.A. Medial lateral extent of thermal and pain sensations evoked by microstimulation in somatic sensory nuclei of human thalamus. J. Neurophysiol. 2003, 90, 2367–2377. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.D. Retrograde analyses of spinothalamic projections in the macaque monkey: Input to ventral posterior nuclei. J. Comp. Neurol. 2006, 499, 965–978. [Google Scholar] [CrossRef] [PubMed]
- Lenz, F.A.; Ohara, S.; Gracely, R.H.; Dougherty, P.M.; Patel, S.H. Pain encoding in the human forebrain: Binary and analog exteroceptive channels. J. Neurosci. 2004, 24, 6540–6544. [Google Scholar] [CrossRef] [PubMed]
- Morison, R.S.; Dempsey, E.W. A study of thalamo-cortical relations. Am. J. Physiol. 1941, 135, 281–292. [Google Scholar]
- Steriade, M. Grouping of brain rhythms in corticothalamic systems. Neuroscience 2006, 137, 1087–1106. [Google Scholar] [CrossRef] [PubMed]
- Steriade, M.; Jones, E.G.; McCormick, D.A. Thalamus Organization and Function; Elsevier: Amsterdam, The Netherlands, 1997; Volume 1. [Google Scholar]
- Ramcharan, E.J.; Cox, C.L.; Zhan, X.J.; Sherman, S.M.; Gnadt, J.W. Cellular mechanisms underlying activity patterns in the monkey thalamus during visual behavior. J. Neurophysiol. 2000, 84, 1982–1987. [Google Scholar] [PubMed]
- Martinez-Conde, S.; Macknik, S.L.; Hubel, D.H. The function of bursts of spikes during visual fixation in the awake primate lateral geniculate nucleus and primary visual cortex. Proc. Natl. Acad. Sci. USA 2002, 99, 13920–13925. [Google Scholar] [CrossRef] [PubMed]
- Jahnsen, H.; Llinas, R. Electrophysiological properties of guinea-pig thalamic neurones: An in vitro study. J. Physiol. 1984, 349, 205–226. [Google Scholar] [CrossRef] [PubMed]
- Destexhe, A.; Sejnowski, T.J. Thalamcortical Assemblies; Oxford University Press: New York, NY, USA, 2001; pp. 1–443. [Google Scholar]
- Jones, E.G. Distribution patterns of individual medial lemniscal axons in the ventrobasal complex of the monkey thalamus. J. Comp. Neurol. 1983, 215, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Apkarian, A.V.; Shi, T.; Bruggemann, J.; Airapetian, L.R. Segregation of nociceptive and non-nociceptive networks in the squirrel monkey somatosensory thalamus. J. Neurophysiol. 2002, 84, 484–494. [Google Scholar]
- Jones, E.G.; Friedman, D.P. Projection pattern of functional components of thalamic ventrobasal complex on monkey somatosensory cortex. J. Neurophysiol. 1982, 48, 521–544. [Google Scholar] [PubMed]
- Lenz, F.A.; Garonzik, I.M.; Zirh, T.A.; Dougherty, P.M. Neuronal activity in the region of the thalamic principal sensory nucleus (ventralis caudalis) in patients with pain following amputations. Neuroscience 1998, 86, 1065–1081. [Google Scholar] [CrossRef]
- Lenz, F.A.; Gracely, R.H.; Baker, F.H.; Richardson, R.T.; Dougherty, P.M. Reorganization of sensory modalities evoked by microstimulation in region of the thalamic principal sensory nucleus in patients with pain due to nervous system injury. J. Comp. Neurol. 1998, 399, 125–138. [Google Scholar] [CrossRef]
- Jensen, T.S.; Rasmussen, P. Phantom pain and related phenomena after amputation. In Textbook of Pain; Wall, P.D., Melzack, R., Eds.; Churchill Livingstone: New York, NY, USA, 1994; pp. 651–665. [Google Scholar]
- Jensen, T.S.; Krebs, B.; Nielsen, J.; Rasmussen, P. Non-painful phantom limb phenomena in amputees: Incidence, clinical characteristics and temporal course. Acta Neurol. Scand. 1984, 70, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.D.; Kiss, Z.H.; Luo, L.; Tasker, R.R.; Lozano, A.M.; Dostrovsky, J.O. Phantom sensations generated by thalamic microstimulation. Nature 1998, 391, 385–387. [Google Scholar] [CrossRef] [PubMed]
- Woods, T.M.; Cusick, C.G.; Pons, T.P.; Taub, E.; Jones, E.G. Progressive transneuronal changes in the brainstem and thalamus after long-term dorsal rhizotomies in adult macaque monkeys. J. Neurosci. 2000, 20, 3884–3899. [Google Scholar] [PubMed]
- Blomqvist, A.; Ericson, A.C.; Craig, A.D.; Broman, J. Evidence for glutamate as a neurotransmitter in spinothalamic tract terminals in the posterior region of owl monkeys. Exp. Brain Res. 1996, 108, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, P.M.; Li, J.; Lenz, F.A.; Rowland, L.; Mittman, S. Evidence that excitatory amino acids mediate afferent input to the primate somatosensory thalamus. Brain Res. 1996, 278, 267–273. [Google Scholar] [CrossRef]
- Hassler, R.; Reichert, T. Klinische und anatomische Befunde bei stereotaktischen Schmerzoperationen im Thalamus. Arch. Psychiat. Nerverkr. 1959, 200, 93–122. [Google Scholar] [CrossRef]
- Dostrovsky, J.O.; Wells, F.E.B.; Tasker, R.R. Pain evoked by stimulation in human thalamus. In International Symposium on Processing Nociceptive Information; Sjigenaga, Y., Ed.; Elsevier: Amsterdam, The Netherlands, 1991; pp. 115–120. [Google Scholar]
- Lenz, F.A.; Seike, M.; Richardson, R.T.; Lin, Y.C.; Baker, F.H.; Khoja, I.; Jaeger, C.J.; Gracely, R.H. Thermal and pain sensations evoked by microstimulation in the area of human ventrocaudal nucleus. J. Neurophysiol. 1993, 70, 200–212. [Google Scholar] [PubMed]
- Halliday, A.M.; Logue, V. Painful sensations evoked by electrical stimulation in the thalamus. In Neurophysiology Studied in Man; Somjen, G.G., Ed.; Excerpta Medica: Amsterdam, The Netherlands, 1972; pp. 221–230. [Google Scholar]
- Edgar, R.E.; Best, L.G.; Quail, P.A.; Obert, A.D. Computer-assisted DREZ microcoagulation: Posttraumatic spinal deafferentation pain. J. Spinal Disord. 1993, 6, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Finnerup, N.B.; Johannesen, I.L.; Fuglsang-Frederiksen, A.; Bach, F.W.; Jensen, T.S. Sensory function in spinal cord injury patients with and without central pain. Brain 2003, 126, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Finnerup, N.B.; Johannesen, I.L.; Bach, F.W.; Jensen, T.S. Sensory function above lesion level in spinal cord injury patients with and without pain. Somatosens. Mot. Res. 2003, 20, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Milhorat, T.H.; Kotzen, R.M.; Mu, H.T.; Capocelli, A.L.; Milhorat, R.H. Dysesthetic pain in patients with syringomyelia. Neurosurgery 1996, 38, 940–946. [Google Scholar] [CrossRef] [PubMed]
- Finnerup, N.B.; Sorensen, L.; Biering-Sorensen, F.; Johannesen, I.L.; Jensen, T.S. Segmental hypersensitivity and spinothalamic function in spinal cord injury pain. Exp. Neurol. 2007, 207, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Davidoff, G.; Roth, E. Clinical characteristics of central (dysesthetic) pain syndrome in spinal cord injury patients. In Pain and Central Nervous System Diseases: The Central Pain Syndromes; Casey, K.L., Ed.; Raven Press: New York, NY, USA, 1991; pp. 77–83. [Google Scholar]
- Radhakrishnan, V.; Tsoukatos, J.; Davis, K.D.; Tasker, R.R.; Lozano, A.M.; Dostrovsky, J.O. A comparison of the burst activity of lateral thalamic neurons in chronic pain and non-pain patients. Pain 1999, 80, 567–575. [Google Scholar] [CrossRef]
- Lu, S.M.; Guido, W.; Sherman, S.M. Effects of membrane voltage on receptive field properties of lateral geniculate neurons in the cat: Contributions of the low-threshold Ca2+ conductance. J. Neurophysiol. 1992, 68, 2185–2198. [Google Scholar] [PubMed]
- Llinas, R.; Nicholson, C. Electrophysiological Properties of Dendrites and somata in Alligator Purkinje Cells. J. Neurophysiol. 1971, 34, 532–550. [Google Scholar] [PubMed]
- Vierck, C.J.; Greenspan, J.D.; Ritz, L.A. Long-term changes in purposive and reflexive responses to nociceptive stimulation following anterolateral chordotomy. J. Neurosci. 1990, 10, 2077–2095. [Google Scholar] [PubMed]
- Beric, A.; Dimitrijevic, M.R.; Lindblom, U. Central dysesthesia syndrome in spinal cord injury patients. Pain 1988, 34, 109–116. [Google Scholar] [CrossRef]
- Ralston, D.D.; Dougherty, P.M.; Lenz, F.A.; Weng, H.R.; Vierck, C.J.; Ralston, H.J. Plasticity of the inhibitory circuitry and neuronal responses in the primate somatosensory thalamus following lesions of the dorsal column and spinothalamic pathways. Prog. Pain. Res. Manag. 2000, 16, 427–434. [Google Scholar]
- Weng, H.R.; Lee, J.-I.; Lenz, F.A.; Vierck, C.J.; Rowland, L.H.; Dougherty, P.M. Functional plasticity in primate somatosensory thalamus following chronic lesion of the ventral lateral spinal cord. Neuroscience 2000, 101, 393–401. [Google Scholar] [CrossRef]
- Vierck, C.J. Can mechanisms of central pain syndromes be investigated in animal models? In Pain and Central Nervous System Disease: The Central Pain Syndromes; Casey, K.L., Ed.; Raven Press: New York, NY, USA, 1991; pp. 129–141. [Google Scholar]
- Patel, S.; Ohara, S.; Dougherty, P.M.; Gracely, R.H.; Lenz, F.A. Psychophysical elements of place and modality specificity in the thalamic somatic sensory nucleus (ventral caudal, vc) of awake humans. J. Neurophysiol. 2006, 95, 646–659. [Google Scholar] [CrossRef] [PubMed]
- Hobson, J.A.; Steriade, M. Neuronal basis of behavioral state control. In Handbook of Physiology - The Nervous System IV; Bloom, F.E., Ed.; Williams & Wilkins: Baltimore, MD, USA, 1986; pp. 701–823. [Google Scholar]
- Zirh, A.T.; Lenz, F.A.; Reich, S.G.; Dougherty, P.M. Patterns of bursting occurring in thalamic cells during parkinsonian tremor. Neuroscience 1997, 83, 107–121. [Google Scholar] [CrossRef]
- Kiss, Z.H.; Tsoukatos, J.; Tasker, R.R.; Davis, K.D.; Dostrovsky, J.O. Sleeping cells in the human thalamus. Stereotact. Funct. Neurosurg. 1995, 65, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.D.; Kiss, Z.H.; Tasker, R.R.; Dostrovsky, J.O. Thalamic stimulation-evoked sensations in chronic pain patients and nonpain (movement disorder) patients. J. Neurophysiol. 1996, 75, 1026–1037. [Google Scholar] [PubMed]
- Erickson, T.C.; Bleckwenn, W.J.; Woolsey, C.N. Observations on the post central gyrus in relation to pain. Trans. Am. Neurol. Assoc. 1952, 56, 57–59. [Google Scholar] [PubMed]
- Lewin, W.; Phillips, C.G. Observations on Partial Removal of the Post-Central Gyrus for Pain. J. Neurol. Neurosurg. Psychiatry 1952, 15, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Kenshalo, D.R., Jr.; Willis, W.D. The role of the cerebral cortex in pain sensation. In Cerebral Cortex, Vol 9 Normal and Altered States of Function; Peters, A., Jones, E.G., Eds.; Plenum Press: New York, NY, USA; London, UK, 1991; pp. 153–212. [Google Scholar]
- Tomlinson, T.; Miller, I. Multi-electrode stimulation in somatosensory area 2 induces a natural sensation of limb movement. Neuromodulation 2014, 17, e114. [Google Scholar]
- Lenz, F.A.; Dougherty, P.M. Neurons in the human thalamic somatosensory nucleus (Ventralis caudalis) respond to innocuous cool and mechanical stimuli. J. Neurophysiol. 1998, 79, 2227–2230. [Google Scholar] [PubMed]
- Jones, E.G. The anatomy of sensory relay functions in the thalamus. Prog. Brain Res. 1991, 87, 29–51. [Google Scholar] [PubMed]
- Rausell, E.; Jones, E.G. Histochemical and immunocytochemical compartments of the thalamic VPM nucleus in monkeys and their relationship to the representational map. J. Neurosci. 1990, 11, 210–225. [Google Scholar]
- Song, W.; Semework, M. Tactile representation in somatosensory thalamus (VPL) and cortex (S1) of awake primate and the plasticity induced by VPL neuroprosthetic stimulation. Brain Res. 2015, 1625, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Hugdahl, K.; Rosen, G.; Ersland, L.; Lundervold, A.; Smievoll, A.I.; Barndon, R.; Thomsen, T. Common pathways in mental imagery and pain perception: An fMRI study of a subject with an amputated arm. Scand. J. Psychol. 2001, 42, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Pare, M.; Behets, C.; Cornu, O. Paucity of presumptive ruffini corpuscles in the index finger pad of humans. J. Comp. Neurol. 2003, 345, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Henshaw, H.; Ferguson, M.A. Efficacy of individual computer-based auditory training for people with hearing loss: A systematic review of the evidence. PLoS ONE 2013, 8, e62836. [Google Scholar] [CrossRef] [PubMed]
- Kral, A.; Yusuf, P.A.; Land, R. Higher-order auditory areas in congenital deafness: Top-down interactions and corticocortical decoupling. Hear. Res. 2017, 343, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.F.; Asghari, A.; Egorov, D.D.; Rutkowski, S.B.; Siddall, P.J.; Soden, R.J.; Ruff, R. Impact of spinal cord injury on self-perceived pre- and postmorbid cognitive, emotional and physical functioning. Spinal Cord 2007, 45, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Nikolajsen, L.; Ilkjaer, S.; Jensen, T.S. Relationship between mechanical sensitivity and postamputation pain: A prospective study. Eur. J. Pain 2000, 4, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Treede, R.D.; Rief, W.; Barke, A.; Aziz, Q.; Bennett, M.I.; Benoliel, R.; Cohen, M.; Evers, S.; Finnerup, N.B.; First, M.B.; et al. A classification of chronic pain for ICD-11. Pain 2015, 156, 1003–1007. [Google Scholar] [CrossRef] [PubMed]




© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chien, J.H.; Korzeniewska, A.; Colloca, L.; Campbell, C.; Dougherty, P.; Lenz, F. Human Thalamic Somatosensory Nucleus (Ventral Caudal, Vc) as a Locus for Stimulation by INPUTS from Tactile, Noxious and Thermal Sensors on an Active Prosthesis. Sensors 2017, 17, 1197. https://doi.org/10.3390/s17061197
Chien JH, Korzeniewska A, Colloca L, Campbell C, Dougherty P, Lenz F. Human Thalamic Somatosensory Nucleus (Ventral Caudal, Vc) as a Locus for Stimulation by INPUTS from Tactile, Noxious and Thermal Sensors on an Active Prosthesis. Sensors. 2017; 17(6):1197. https://doi.org/10.3390/s17061197
Chicago/Turabian StyleChien, Jui Hong, Anna Korzeniewska, Luana Colloca, Claudia Campbell, Patrick Dougherty, and Frederick Lenz. 2017. "Human Thalamic Somatosensory Nucleus (Ventral Caudal, Vc) as a Locus for Stimulation by INPUTS from Tactile, Noxious and Thermal Sensors on an Active Prosthesis" Sensors 17, no. 6: 1197. https://doi.org/10.3390/s17061197
APA StyleChien, J. H., Korzeniewska, A., Colloca, L., Campbell, C., Dougherty, P., & Lenz, F. (2017). Human Thalamic Somatosensory Nucleus (Ventral Caudal, Vc) as a Locus for Stimulation by INPUTS from Tactile, Noxious and Thermal Sensors on an Active Prosthesis. Sensors, 17(6), 1197. https://doi.org/10.3390/s17061197