Exploration of Chlorophyll a Fluorescence and Plant Gas Exchange Parameters as Indicators of Drought Tolerance in Perennial Ryegrass



 ,
, 
 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants, Growth Conditions, and Experiment Design
2.2. The Chlorophyll Fluorescence and the Gas Exchange Measurements
2.3. Statistical Analysis
3. Results
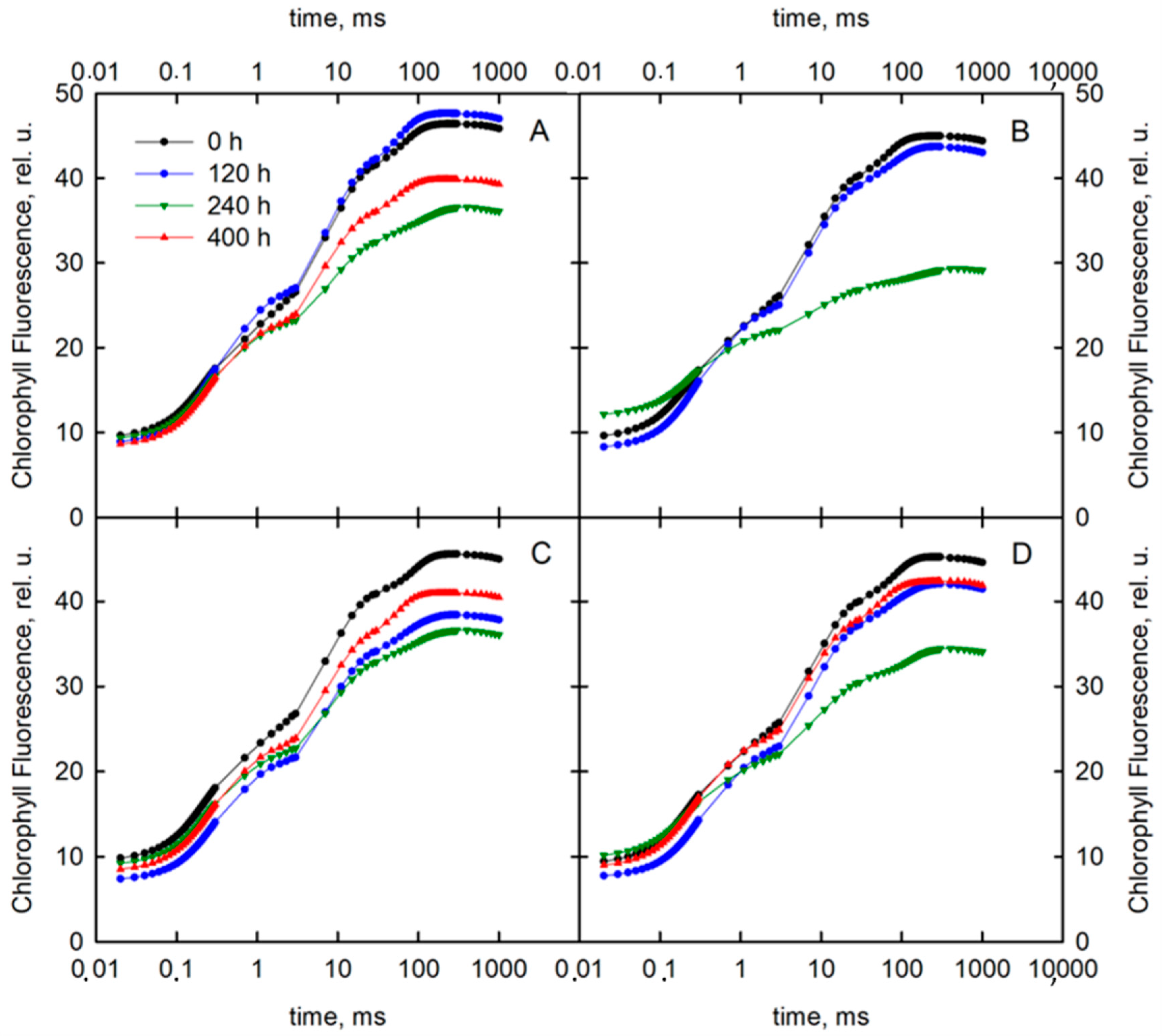
3.1. Prompt Chlorophyll a Fluorescence
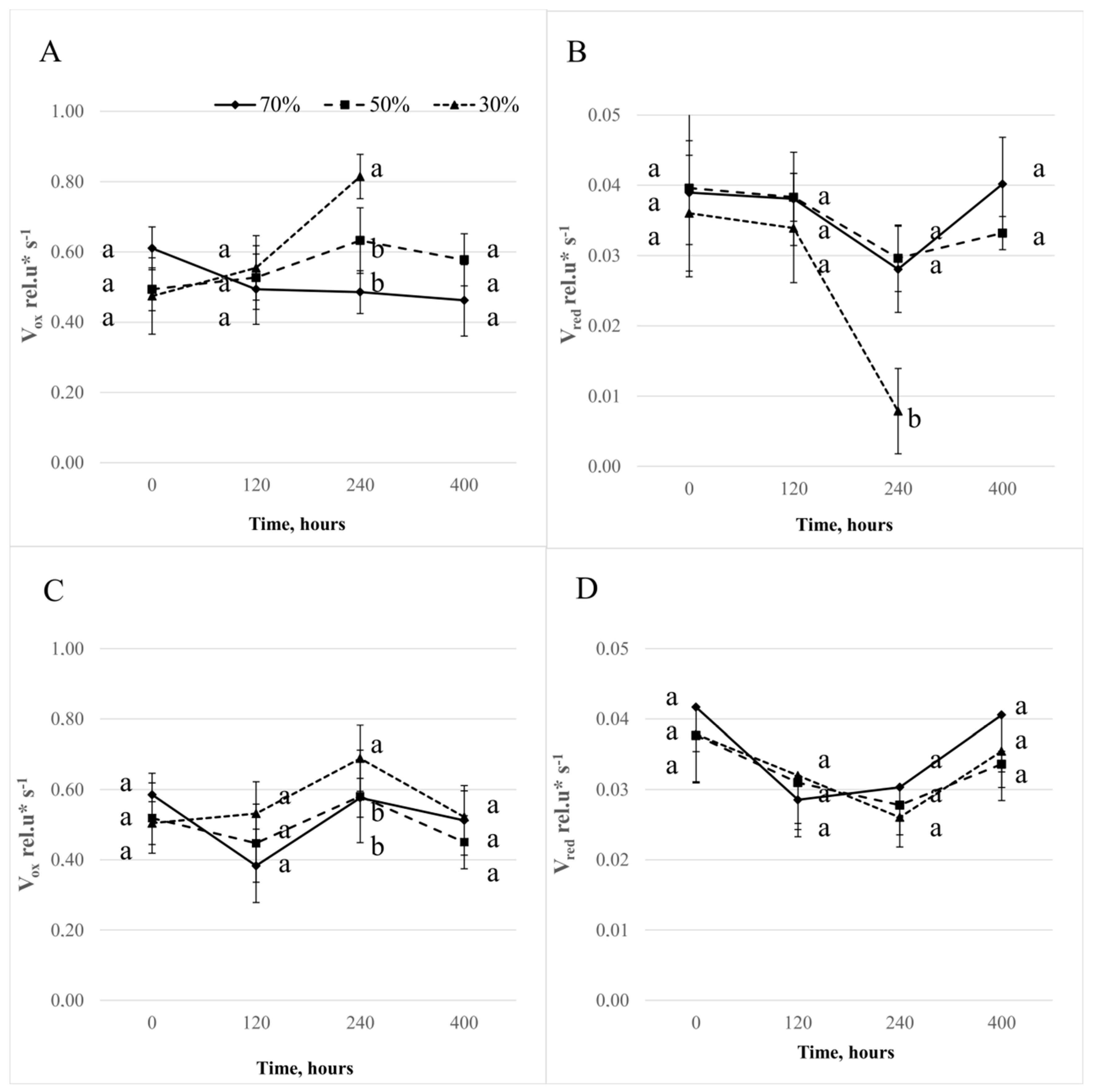
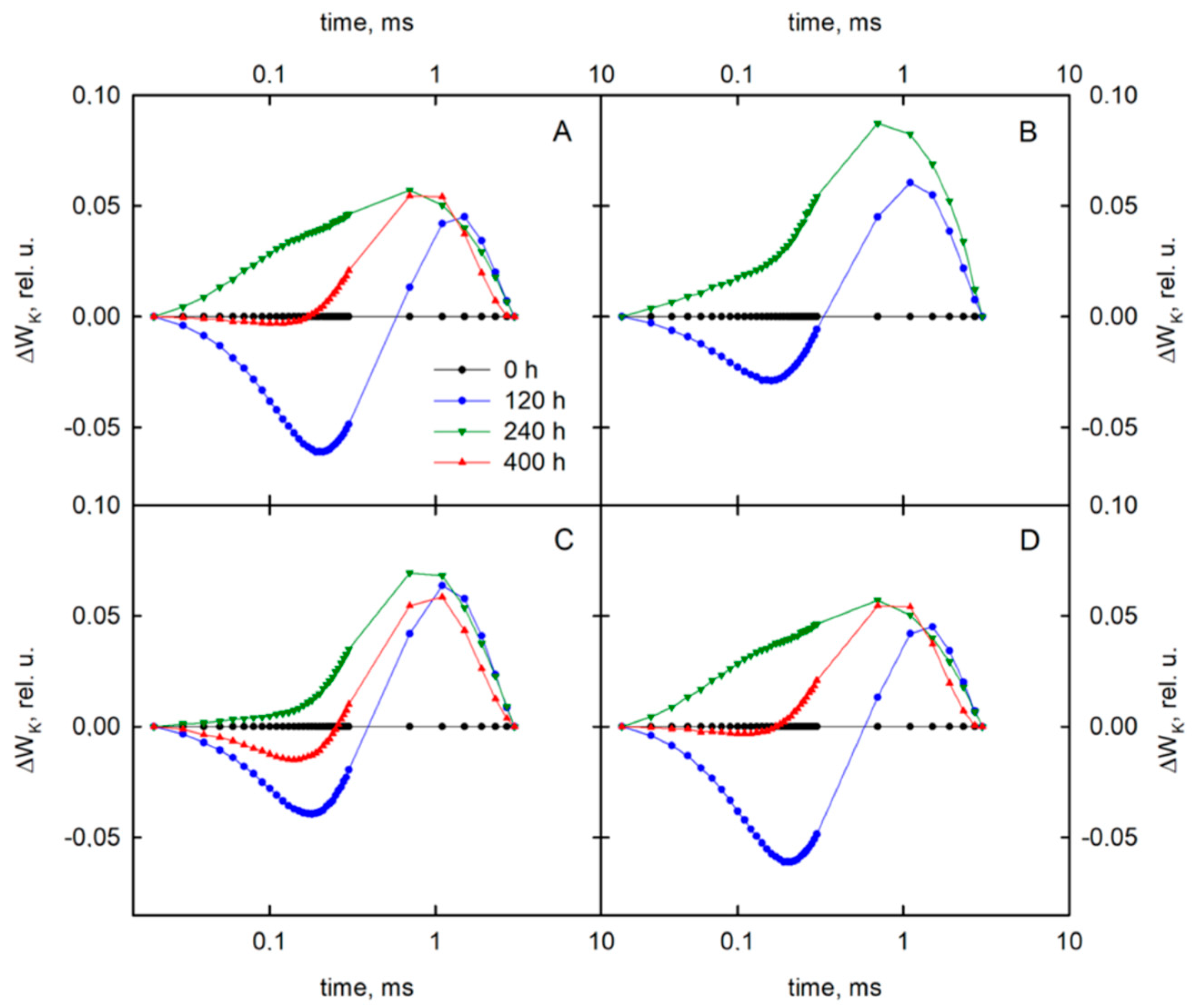
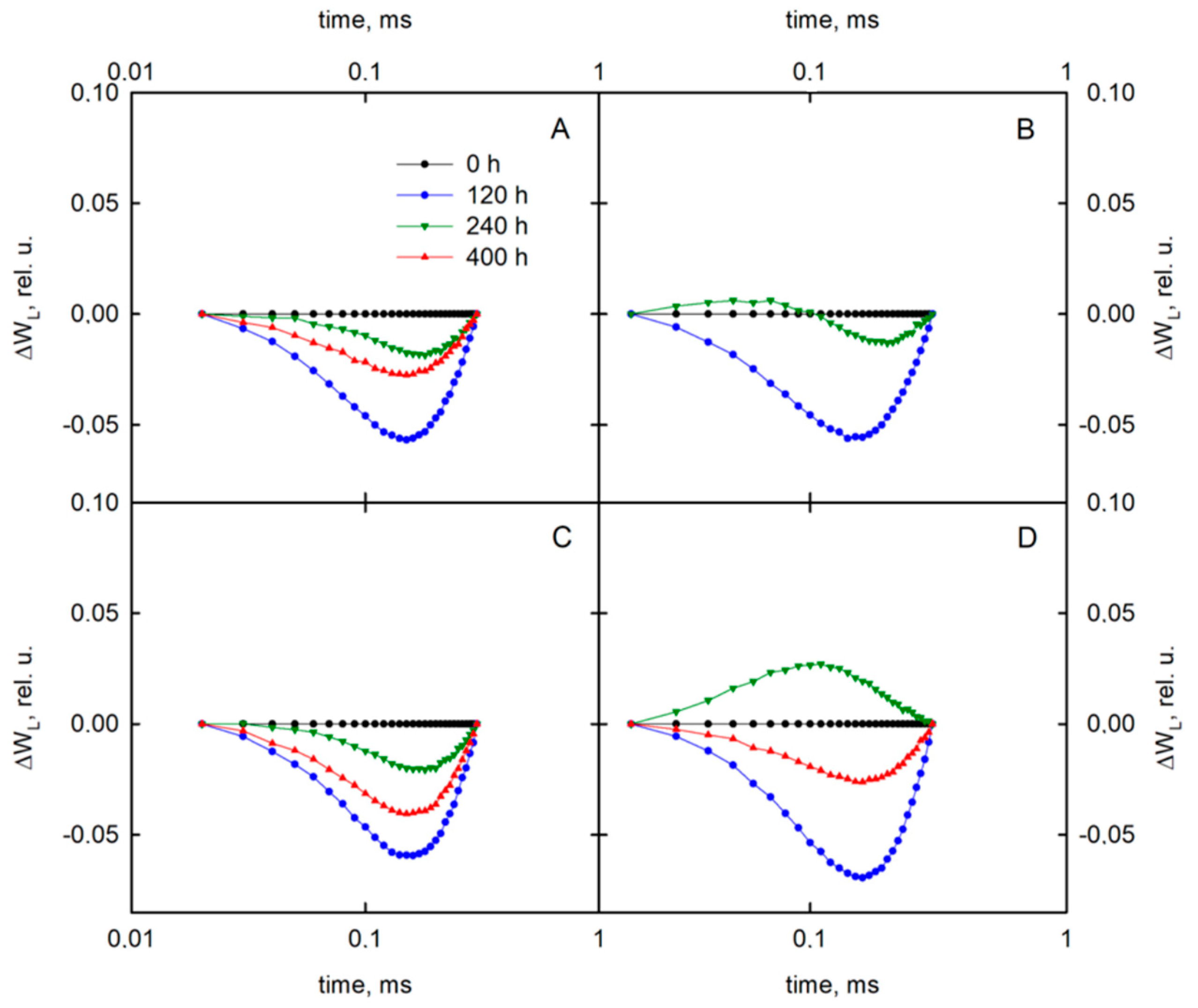
3.2. Delayed Chlorophyll a Fluorescence
3.3. The Modulated Light Reflection Signal Measured at 820 nm
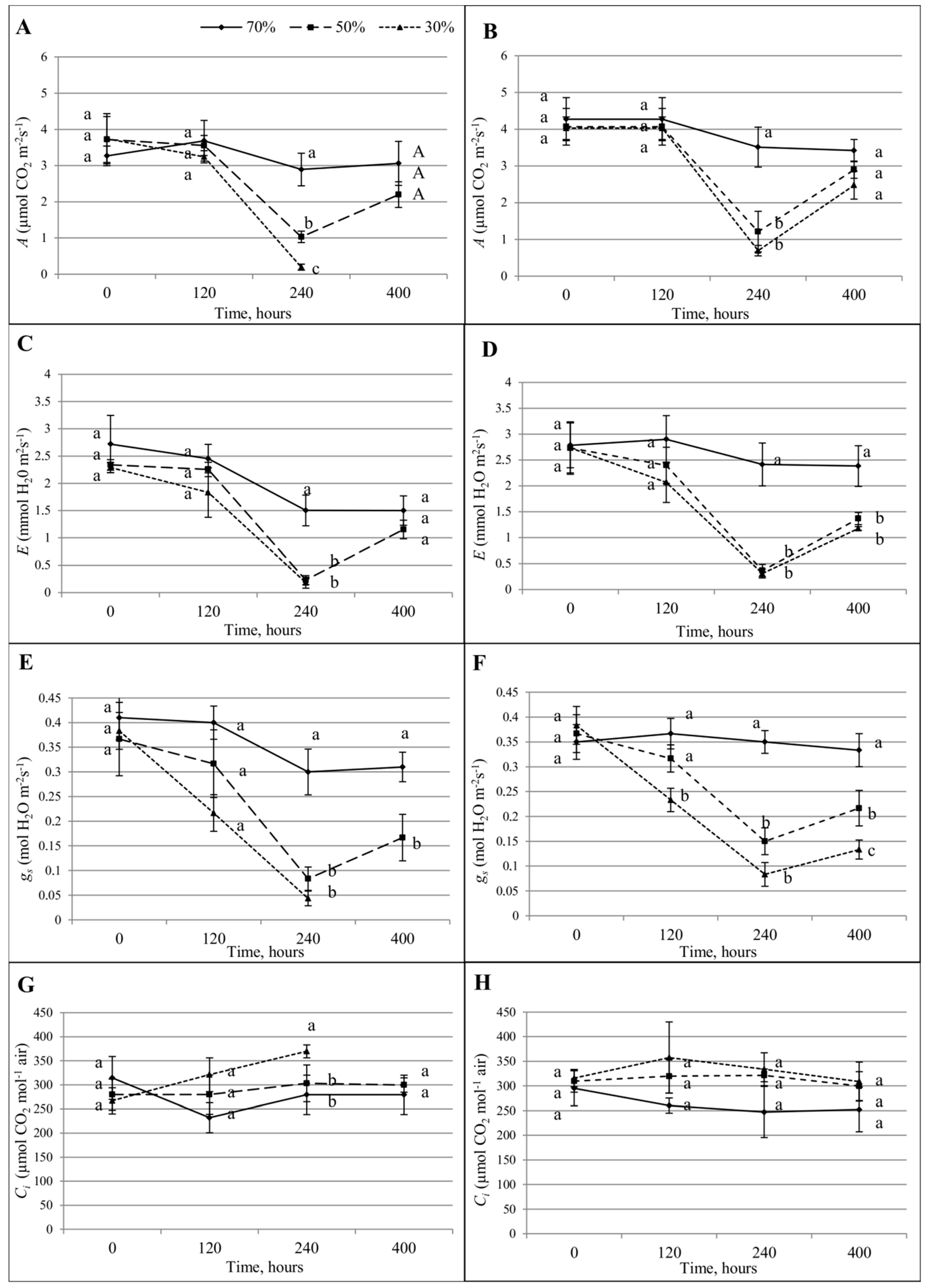
3.4. Gas Exchange Analyses
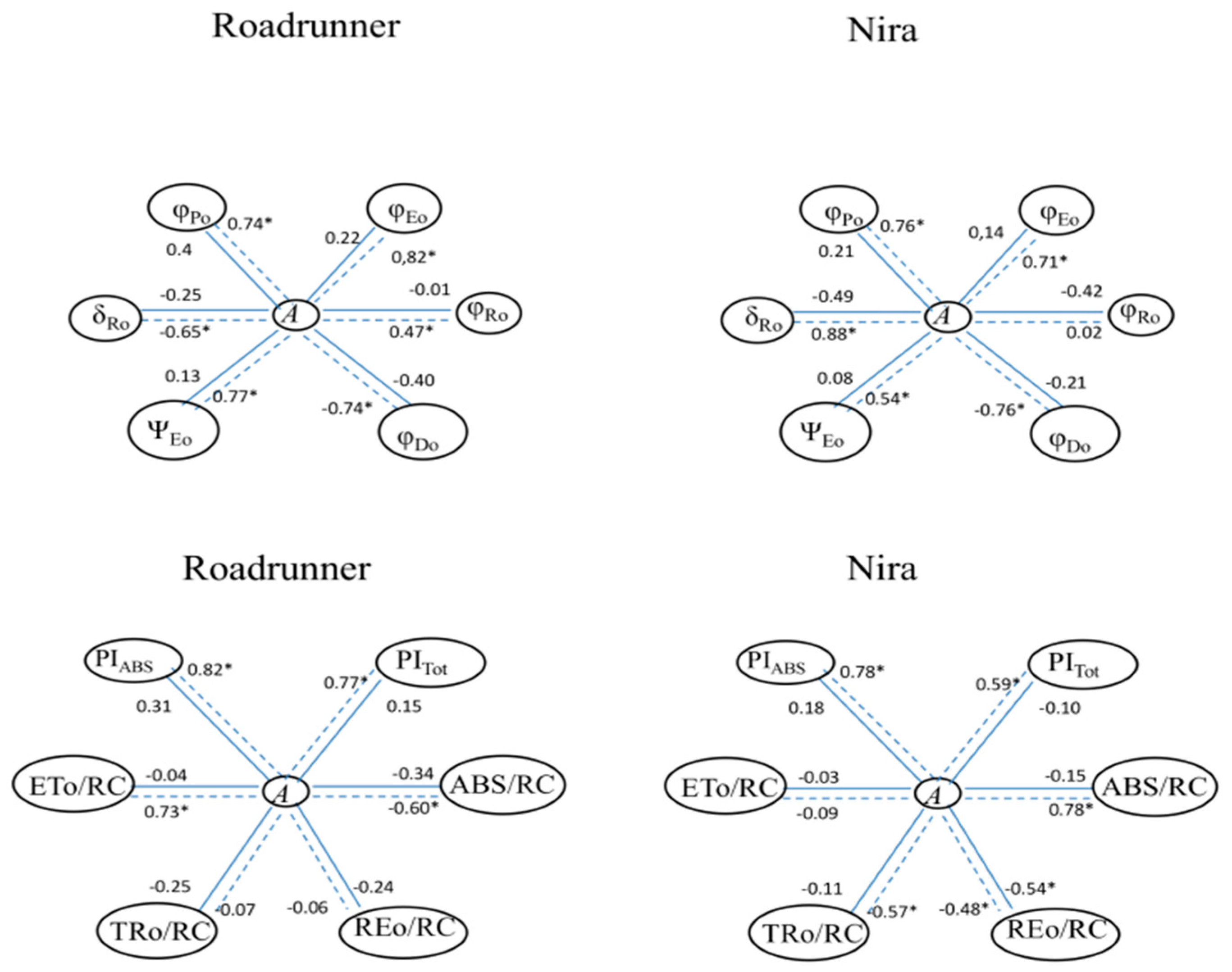
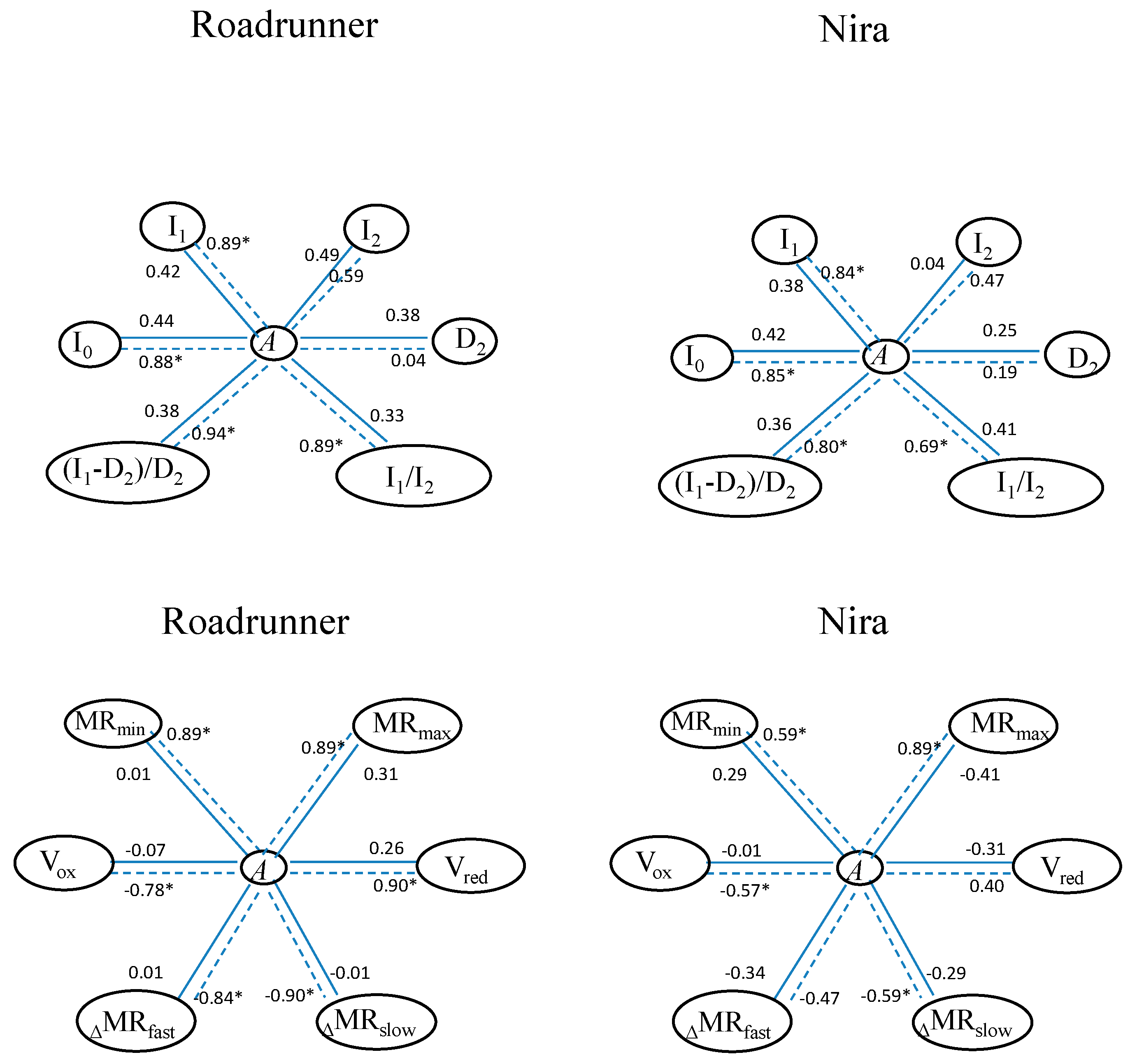
3.5. Relationships among ChlF Parameters and Gas Exchange
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A




References
- Radziemska, M.; Gusiatin, Z.M.; Bilgin, A.A. Potential of using immobilizing agents in aided phytostabilization on simulated contamination of soil with lead. Ecol. Eng. 2017, 102, 490–500. [Google Scholar] [CrossRef]
- Radziemska, M.; Bilgin, A.; Vaverková, M.D. Application of mineral–based amendments for enhancing phytostabilization in Lolium perenne L. cultivation. Clean-Soil Air Water 2018, 46. [Google Scholar] [CrossRef]
- Bęś, A.; Warmiński, K.; Adomas, B. Long-term responses of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) to the contamination of light soils with diesel oil. Environ. Sci. Pollut. Res. 2019, in press. [Google Scholar] [CrossRef]
- Sung, C.Y. Mitigating surface urban heat island by a tree protection policy: A case study of The Woodland, Texas, USA. Urban For. Urban Green. 2013, 12, 474–480. [Google Scholar] [CrossRef]
- Hidalgo, J.; Masson, V.; Baklanov, A.; Pigeon, G.; Gimeno, L. Advances in urbanclimate modeling. Ann. N. Y. Acad. Sci. 2008, 1146, 354–374. [Google Scholar] [CrossRef]
- Fusaro, L.; Salvatori, E.; Mereu, S.; Marando, F.; Scassellati, E.; Abbate, G.; Manes, F. Urban and peri-urban forests in the metropolitan area of Rome: Ecophysiological response of Quercus ilex L. in two greeninfrastructures in an ecosystem services perspective. Urban For. Urban Green. 2015, 14, 1147–1156. [Google Scholar] [CrossRef]
- Nosalewicz, A.; Siecińska, J.; Kondracka, K.; Nosalewicz, M. The functioning of Festuca arundinacea and Lolium perenne under drought is improved to a different extend by the previous exposure to water deficit. Environ. Exp. Bot. 2018, 156, 271–278. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 2002. [CrossRef]
- Zivcak, M.; Kalaji, H.M.; Shao, H.-B.; Olsovska, K.; Brestic, M. Photosynthetic proton and electron transport in wheat leaves under prolonged moderate drought stress. J. Photoch. Photobiol. B 2014, 137, 107–115. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Rastogi, A.; Živčák, M.; Brestic, M.; Daszkowska-Golec, A.; Sitko, K.; Alsharafa, K.Y.; Lotfi, R.; Stypiński, P.; Samborska, I.A.; et al. Prompt chlorophyll fluorescence as a tool for crop phenotyping: An example of barley landraces exposed to various abiotic stress factors. Photosynthetica 2018, 56, 953–961. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Račková, L.; Paganová, V.; Swoczyna, T.; Rusinkowski, S.; Sitko, K. Can chlorophyll-a fluorescence parameters be used as bio-indicators to distinguish between drought and salinity stress in Tilia cordata Mill? Environ. Exp. Bot. 2018, 152, 149–157. [Google Scholar] [CrossRef]
- Kosmala, A.; Perlikowski, D.; Pawłowicz, I.; Rapacz, M. Changes in the chloroplast proteome following water deficit and subsequent watering in a high and a low drought tolerant genotype of Festuca arundinacea. J. Exp. Bot. 2012, 63, 6161–6172. [Google Scholar] [CrossRef]
- Perlikowski, D.; Kiersznowska, S.; Sawikowska, A.; Krajewski, P.; Rapacz, M.; Eckhardt, Ä.; Kosmala, A. Remodeling of leaf cellular glycerolipid composition under drought and re-hydration conditions in grasses from the Lolium-Festuca complex. Front. Plant Sci. 2016, 7, 1027. [Google Scholar] [CrossRef]
- Digrado, A.; Bachy, A.; Mozaffar, A.; Schoon, N.; Bussotti, F.; Amelynck, C.; Dalcq, A.-C.; Fauconnier, M.-L.; Aubinet, M.; Heinesch, B.; et al. Long-term measurements of chlorophyll a fluorescence using the JIP-test show that combined abiotic stresses influence the photosynthetic performance of the perennial ryegrass (Lolium perenne) in a managed temperate grassland. Physiol. Plant 2017, 161, 355–371. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Pawluśkiewicz, B.; Kalaji, H.M.; Baczewska, A.H. The effect of light availability on leaf area index, biomass production and plant species composition of park grasslands in Warsaw. Plant Soil Environ. 2013, 59, 543–548. [Google Scholar] [CrossRef]
- Common Catalogue of Varieties of Agricultural Plant Species. 36th complete edition. (2017/C 433/01). Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3AC2017%2F433%2F01 (accessed on 6 June 2019).
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence: A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef]
- Ouni, Y.; Mateos-Naranjo, E.; Abdelly, C.; Lakhdar, A. Interactive effect of salinity and zinc stress on growth and photosynthetic responses of the perennial grass, Polypogon monspeliensis. Ecol. Eng. 2016, 95, 171–179. [Google Scholar] [CrossRef]
- Pollastrini, M.; Holland, V.; Brüggemann, W.; Bruelheide, H.; Dânilâ, I.; Jaroszewicz, B.; Valladares, F.; Bussotti, F. Taxonomic and ecological relevance of the chlorophyll a fluorescence signature of tree species in mixed European forests. New Phytol. 2016, 212, 51–65. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Pawluśkiewicz, B.; Baczewska, A.H.; Kalaji, H.M.; Oglęcki, P. Chlorophyll a fluorescence of perennial ryegrass (Lolium perenne L.) varieties under long term exposure to shade. Zemdirbyste-Agriculture 2015, 102, 305–312. [Google Scholar] [CrossRef]
- Strasser, B.; Strasser, R. Measuring fast fluorescence transients to address environmental questions: The JIP-test. In Photosynthesis: From Light to Biosphere; Garab, G., Ed.; Kluwer: Dordrecht, Netherlands, 1995; Volume 5, pp. 977–980. [Google Scholar]
- Kalaji, H.M.; Bosa, K.; Kościelniak, J.; Żuk-Gołaszewska, K. Effects of salt stress on photosystem II efficiency and CO2 assimilation of two Syrian barley landraces. Environ. Exp. Bot. 2011, 73, 64–72. [Google Scholar] [CrossRef]
- Stirbet, A. Govindjee On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B Biol. 2011, 104, 236–257. [Google Scholar] [CrossRef] [PubMed]
- Oukarroum, A.; Madidi, S.E.; Schansker, G.; Strasser, R.J. Probing the responses of barley cultivars (Hordeum vulgare L.) by chlorophyll a fluorescence OLKJIP under drought stress and re-watering. Eviron. Exp. Bot. 2007, 60, 438–446. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Baczewska, A.H.; Pawluśkiewicz, B.; Paunov, M.; Alexantrov, V.; Goltsev, V.; Kalaji, H.M. Prompt chlorophyll a fluorescence as a rapid tool for diagnostic changes in PSII structure inhibited by salt stress in Perennial ryegrass. J. Photochem. Photobiol. B Biol. 2016, 157, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.-L.; Song, J.-F.; Wen, J.; Zhang, J.; Wei, J.-H. Effects of drought stress on fluorescence characteristics of photosystem II in leaves of Plectranthus scutellarioides. Photosynthetica 2016, 54, 414–421. [Google Scholar] [CrossRef]
- Goltsev, V.; Zaharieva, I.; Lambrev, P.; Yordanov, I.; Strasser, R. Simultaneous analysis of prompt and delayed chlorophyll a fluorescence in leaves during the induction period of dark to light adaptation. J. Theor. Biol. 2003, 225, 171–183. [Google Scholar] [CrossRef]
- Zaharieva, I.; Goltsev, V. Advances on Photosystem II investigation by measurement of delayed chlorophyll fluorescence by a phosphoroscopic method. Photochem. Photobiol. 2003, 77, 292–298. [Google Scholar] [CrossRef]
- Guo, Y.; Tan, J. A kinetic model structure for delayed fluorescence from plants. BioSystems 2009, 95, 98–103. [Google Scholar] [CrossRef]
- Oukarroum, A.; Goltsev, V.; Strasser, R.J. Temperature Effects on Pea Plants Probed by Simultaneous Measurements of the Kinetics of Prompt Fluorescence, Delayed Fluorescence and Modulated 820 nm Reflection. PLoS ONE 2013, 8, e59433. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Kalaji, H.M.; Baczewska, A.H.; Pawluśkiewicz, B.; Mastalerczuk, G.; Borawska-Jarmułowicz, B.; Paunov, M.; Goltsev, V. Delayed chlorophyll a fluorescence, MR 820, and gas exchange changes in perennial ryegrass under salt stress. J. Lumin. 2017, 183, 322–333. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Qiang, S.; Goltsev, V. Simultaneous in vivo recording of prompt and delayed fluorescence and 820-nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. Biochim. Biophys. Acta Bioenerget. 2010, 1797, 1313–1326. [Google Scholar] [CrossRef]
- Schansker, G.; Srivastava, A.; Govindjee; Strasser, R.J. Characterization of the 820-nm transmission signal paralleling the chlorophyll a fluorescence rise (OJIP) in pea leaves. Funct. Plant Biol. 2003, 30, 785–796. [Google Scholar] [CrossRef] [Green Version]
- Goltsev, V.; Zaharieva, I.; Chernev, P.; Strasser, R. Delayed fluorescence in photosynthesis. Photosynth. Res. 2009, 101, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Van Heerden, P.D.R.; Swanepoel, J.W.; Kruger, G.H.J. Modulation of photosynthesis by drought in two desert scrub species exhibiting C3-mode CO2 assimilation. Environ. Exp. Bot. 2009, 61, 124–136. [Google Scholar] [CrossRef]
- Kocheva, K.V.; Busheva, M.C.; Georgiev, G.I.; Lambrev, P.H.; Goltsev, V.N. Influence of short-term osmotic stress on the photosynthetic activity of barley seedlings. Biol. Plant. 2005, 49, 145–148. [Google Scholar] [CrossRef]
- Lawson, T.; Oxborough, K.; Morison, J.L.; Baker, N.R. The responses of guard and mesophyll cell photosynthesis to CO2, O2, light, and water stress in a range of species are similar. J. Exp. Bot. 2003, 54, 1743–1752. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.X.; Wang, G.X. Effects of Drought Stress on Leaf Gas Exchange and Chlorophyll Fluorescence of Glycyrrhiza uralensis. Rus. J. Ecol. 2014, 46, 532–538. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Zhang, G.C.; Gu, S.Y.; Xia, J.B.; Zhao, J.K. Critical responses of photosynthetic efficiency of goldspur apple tree to soil water variation in semiarid loess hilly area. Photosynthetica 2010, 48, 589–595. [Google Scholar] [CrossRef]
- Lang, Y.; Wang, M.; Xia, J.; Zhao, Q. Effects of soil drought stress on photosynthetic gas exchange traits and chlorophyll fluorescence in Forsythia suspensa. For. Res. 2018, 29, 45–53. [Google Scholar] [CrossRef]
- Ohashi, Y.; Nakayama, N.; Saneokal, H.; Fujita, K. Effects of drought stress on photosynthetic gas exchange, chlorophyll fluorescence and stem diameter of soybean plants. Biol. Plant. 2006, 50, 138–141. [Google Scholar] [CrossRef]
- Pukacki, P.M.; Kamińska-Rożek, E. Effect of drought stress on chlorophyll a fluorescence and electrical admittance of shoots in Norway spruce seedlings. Trees 2005, 19, 539–544. [Google Scholar] [CrossRef]
- Guo, P.; Baum, M.; Varshney, R.K.; Graner, A.; Grando, S.; Ceccarelli, S. QTLs for chlorophyll and chlorophyll fluorescence parameters in barley under post-flowering drought. Euphytica 2008, 163, 203–214. [Google Scholar] [CrossRef]
- Longenberger, P.S.; Smith, C.W.; Duke, S.E.; McMichael, B.L. Evaluation of chlorophyll fluorescence as a tool for the identification of drought tolerance in upland cotton. Euphytica 2009, 66, 25–33. [Google Scholar] [CrossRef]
- Hura, T.; Grzesiak, S.; Hura, K.; Thiemt, E.; Tokarz, K.; Wędzony, M. Physiological and biochemical tools useful in drought-tolerance detection in genotypes of winter triticale: Accumulation of ferulic acid correlates with drought tolerance. Ann. Bot. 2007, 100, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Razavi, F.; Pollet, B.; Steppe, K.; Labeke, M.C. Chlorophyll fluorescence as a tool for evaluation of drought stress in strawberry. Photosynthetica 2008, 46, 631–633. [Google Scholar] [CrossRef]
- Perlikowski, D.; Kosmala, A.; Rapacz, M.; Kościelniak, J.; Pawłowicz, I.; Zwierzykowski, Z. Influence of short-term drought conditions and subsequent re-watering on the physiology and proteome of Lolium multiflorum/Festuca arundinacea introgression forms with contrasting levels of tolerance to long-term drought. Plant Biol. 2014, 16, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Slabbert, R.M.; Krüger, G.H.J. Assessment of changes in photosystem II structure and function as affected by water deficit in Amaranthus hypochondriacus L. and Amaranthus hybridus L. Plant Physiol. Biochem. 2011, 49, 978–984. [Google Scholar] [CrossRef]
- Oukarroum, A.; Bussotti, F.; Goltsev, V.; Kalaji, H.M. Correlation between reactive oxygen species production and photochemistry of photosystems I and II in Lemna gibba L. plants under salt stress. Environ. Exp. Bot. 2015, 109, 80–88. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant. Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef]
- Gao, J.; Li, P.; Ma, F.; Goltsev, V. Photosynthetic performance during leaf expansion in Malus micromalus probed by chlorophyll a fluorescence and modulated 820 nm reflection. J. Photochem. Photobiol. B Biol. 2014, 137, 144–150. [Google Scholar] [CrossRef]
- Oukarroum, A.; Strasser, R.J.; Schansker, G. Heat stress and the photosynthetic electron transport chain of the lichen Parmelina tiliacea (Hoffm.) Ach. in the dry and the wet state: Differences and similarities with the heat stress response of higher plants. Photosynth. Res. 2012, 111, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Hamdani, S.; Qu, M.; Xin, C.-P.; Li, M.; Chu, C.; Govindjee; Zhu, X.-G. Variations between the photosynthetic properties of elite and landrace Chinese rice cultivars revealed by simultaneous measurements of 820 nm transmission signal and chlorophyll a fluorescence induction. J. Plant Physiol. 2015, 177, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Goltsev, V.; Zaharieva, I.; Chernev, P.; Kouzmanova, M.; Kalaji, H.M.; Yordanov, I.; Krasteva, V.; Alexandrov, V.; Stefanov, D.; Allakhverdiev, S.I.; et al. Drought-induced modifications of photosynthetic electron transport in intact leaves: Analysis and use of neural networks as a tool for a rapid non-invasive estimation. Biochim. Biophys. Acta 2012, 1817, 1490–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvatori, E.; Fusaro, L.; Gottardini, E.; Pollastrini, M.; Goltsev, V.; Strasser, R.J.; Bussotti, F. Plant stress analysis: Application of prompt, delayed chlorophyll fluorescence and 820 nm modulated reflectance. Insights from independent experiments. Plant Physiol. Biochem. 2014, 85, 105–113. [Google Scholar] [CrossRef]
- Zhou, R.; Kan, X.; Chen, J.; Hua, H.; Li, Y.; Ren, J.; Feng, K.; Liu, H.; Deng, D.; Yin, Z. Drought-induced changes in photosynthetic electron transport in maize probed by prompt fluorescence, delayed fluorescence, P700 and cyclic electron flow signals. Environ. Exp. Bot. 2019, 158, 51–62. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VJ = (FJ − F0)/(FM − F0) | Relative variable fluorescence at the J-step |
| φPo = 1 − F0/FM | Maximum quantum yield of primary photochemistry (at t = 0) |
| φEo = (1 − F0/FM)(1 − VJ) | Quantum yield of electron transport (at t = 0) |
| φRo = (1 − F0/FM)(1 − VI) | Quantum yield of reduction of end electron acceptors at the PSI acceptor side (reaction center-RE) |
| φDo = F0/FM | Quantum yield (at t = 0) of energy dissipation |
| ΨEo = 1 − VJ | Probability (at t = 0) that a trapped exciton moves an electron into the electron transport chain beyond Q A − |
| δRo = (1 − VI)/(1 − VJ) | Efficiency/probability with which an electron from the intersystem electron carriers moves to reduce end electron acceptors at the PSI acceptor side (RE) |
| t(FM) | Time (in m/s) to reach the maximal fluorescence intensity FM |
| PIABS = γRC/(1 − γRC) × φPo/(1 − φPo) × ΨEo/(1 − ΨEo) | Performance index (potential) for energy conservation from exciton to the reduction of intersystem electron acceptors |
| PItotal = PIABS × δRo/(1 − δRo) | Performance index (potential) for energy conservation from exciton to the reduction of PSI end acceptors |
| ABS/RC = (1 − γRC)/γRC | Absorption flux (of antenna Chls) per RC |
| M0 | Approximated initial slope (in ms−1) of the fluorescence transient V = f(t) |
| TR0/RC = M0(1/VJ) | Trapping flux (leading to QA reduction) per RC |
| ET0/RC = M0(1/VJ)Ψ0 | Electron transport flux (further than Q A−) per RC |
| RE0/RC = M0(1/VJ)(1 − VI) | Electron flux reducing end electron acceptors at the PSI acceptor side per RC |
| DI0/RC = (ABS/RC − TR0/RC) | Dissipated energy flux per RC (at t = 0) |
| RC/CS0 = φPo (VJ/M0) F0 | Density of RCs (QA reducing PSII reaction centers) |
| I1 and I2 | Maximum of the DF induction curve |
| D2 | Minimum of the DF induction curve |
| MR0 | Modulated 820-nm reflection intensity at Time “0” |
| MRmin | Minimum of modulated 820-nm reflection intensity |
| MRmax | Maximum of modulated 820-nm reflection intensity |
| ΔMRfast | Fast phase (oxidation) of reflection intensity = MR0 − MRmin |
| ΔMRslow | Slow phase (reduction) of reflection intensity = MRmax − MRmin |
| A | CO2 assimilation (µmol CO2 m−2·s−1) |
| E | H20 transpiration rate (mmol H2O m−2·s−1) |
| Gs | Stomatal conductance (mol H2O m−2·s−1) |
| Ci | Sub-stomatal CO2 concentration (µmol CO2 mol−1 air) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dąbrowski, P.; Baczewska-Dąbrowska, A.H.; Kalaji, H.M.; Goltsev, V.; Paunov, M.; Rapacz, M.; Wójcik-Jagła, M.; Pawluśkiewicz, B.; Bąba, W.; Brestic, M. Exploration of Chlorophyll a Fluorescence and Plant Gas Exchange Parameters as Indicators of Drought Tolerance in Perennial Ryegrass. Sensors 2019, 19, 2736. https://doi.org/10.3390/s19122736
Dąbrowski P, Baczewska-Dąbrowska AH, Kalaji HM, Goltsev V, Paunov M, Rapacz M, Wójcik-Jagła M, Pawluśkiewicz B, Bąba W, Brestic M. Exploration of Chlorophyll a Fluorescence and Plant Gas Exchange Parameters as Indicators of Drought Tolerance in Perennial Ryegrass. Sensors. 2019; 19(12):2736. https://doi.org/10.3390/s19122736
Chicago/Turabian StyleDąbrowski, Piotr, Aneta H. Baczewska-Dąbrowska, Hazem M. Kalaji, Vasilij Goltsev, Momchil Paunov, Marcin Rapacz, Magdalena Wójcik-Jagła, Bogumiła Pawluśkiewicz, Wojciech Bąba, and Marian Brestic. 2019. "Exploration of Chlorophyll a Fluorescence and Plant Gas Exchange Parameters as Indicators of Drought Tolerance in Perennial Ryegrass" Sensors 19, no. 12: 2736. https://doi.org/10.3390/s19122736
APA StyleDąbrowski, P., Baczewska-Dąbrowska, A. H., Kalaji, H. M., Goltsev, V., Paunov, M., Rapacz, M., Wójcik-Jagła, M., Pawluśkiewicz, B., Bąba, W., & Brestic, M. (2019). Exploration of Chlorophyll a Fluorescence and Plant Gas Exchange Parameters as Indicators of Drought Tolerance in Perennial Ryegrass. Sensors, 19(12), 2736. https://doi.org/10.3390/s19122736