Using Airborne Hyperspectral Imaging Spectroscopy to Accurately Monitor Invasive and Expansive Herb Plants: Limitations and Requirements of the Method

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species
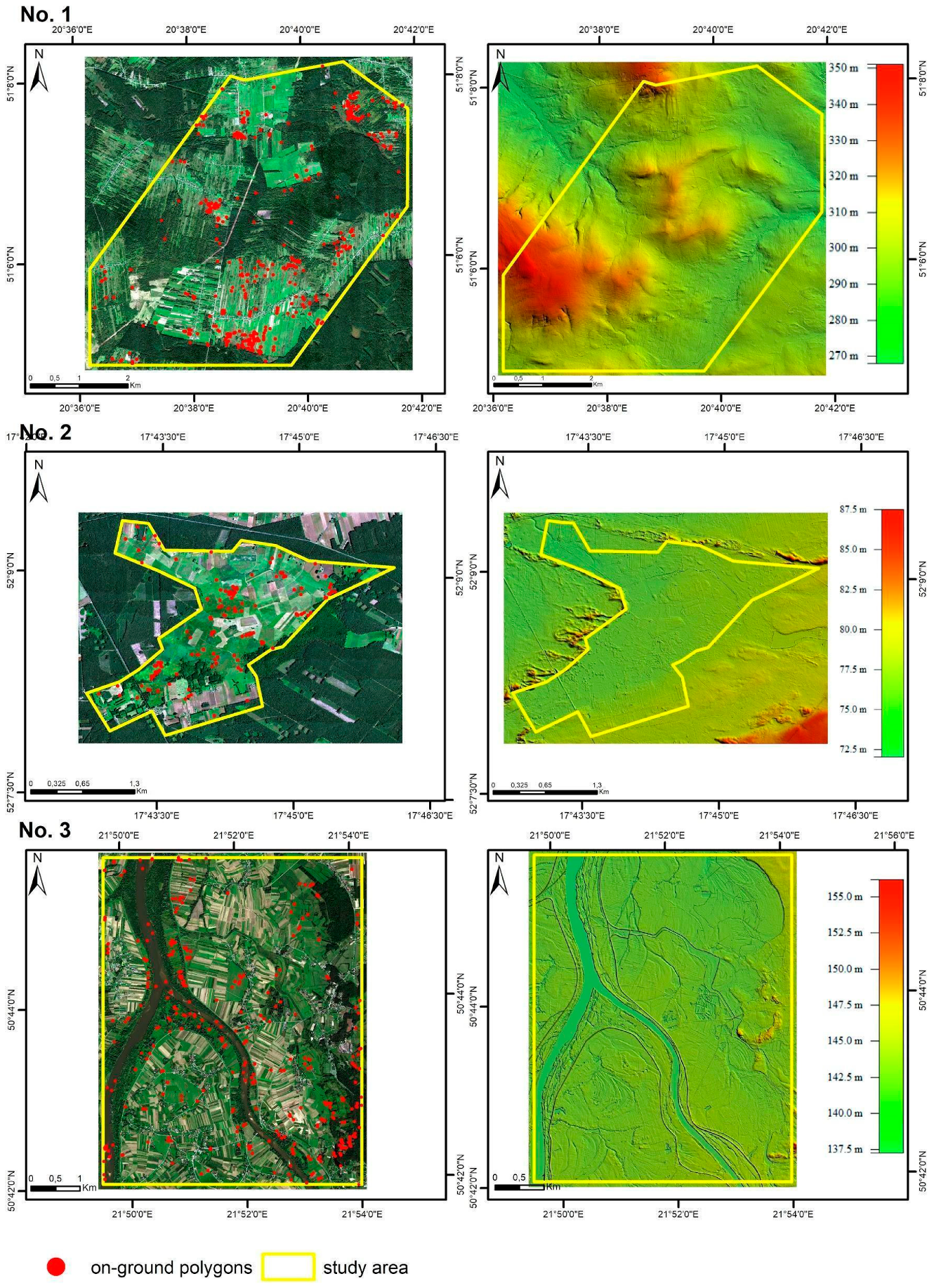
2.2. Study Areas
2.3. Airborne Data Acquisition
2.4. On-Ground Botanical Data for Training and Validation
2.5. Field Mapping
2.6. Random Forest Classification and Accuracy Assessment
3. Results
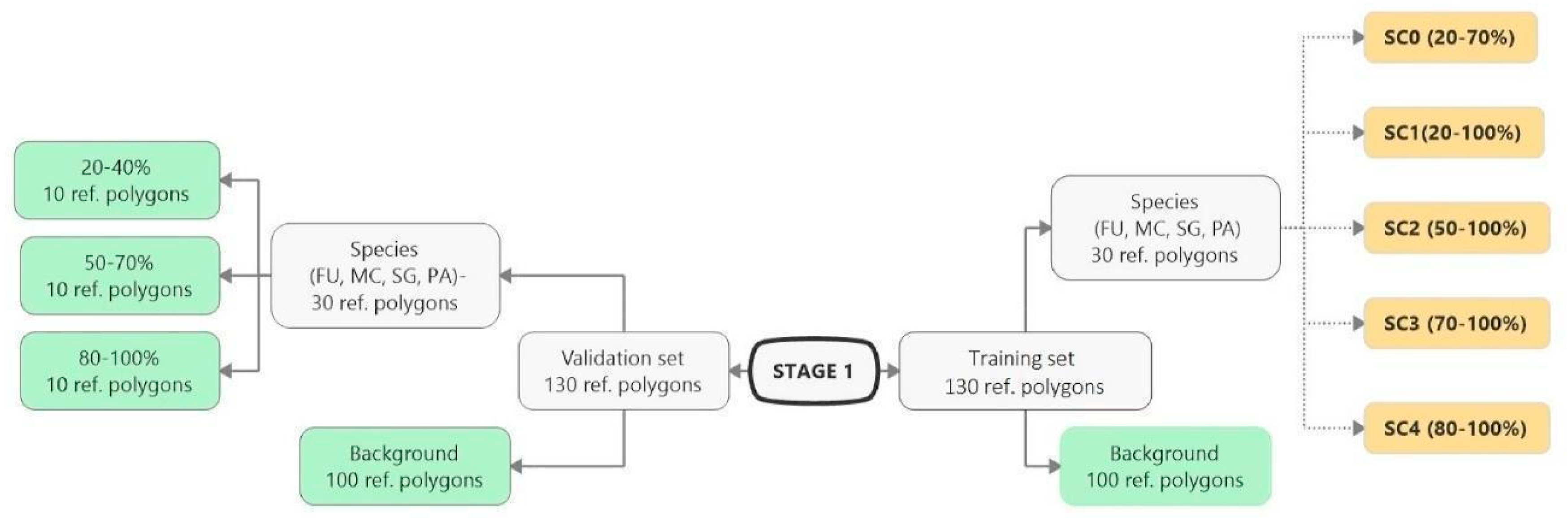
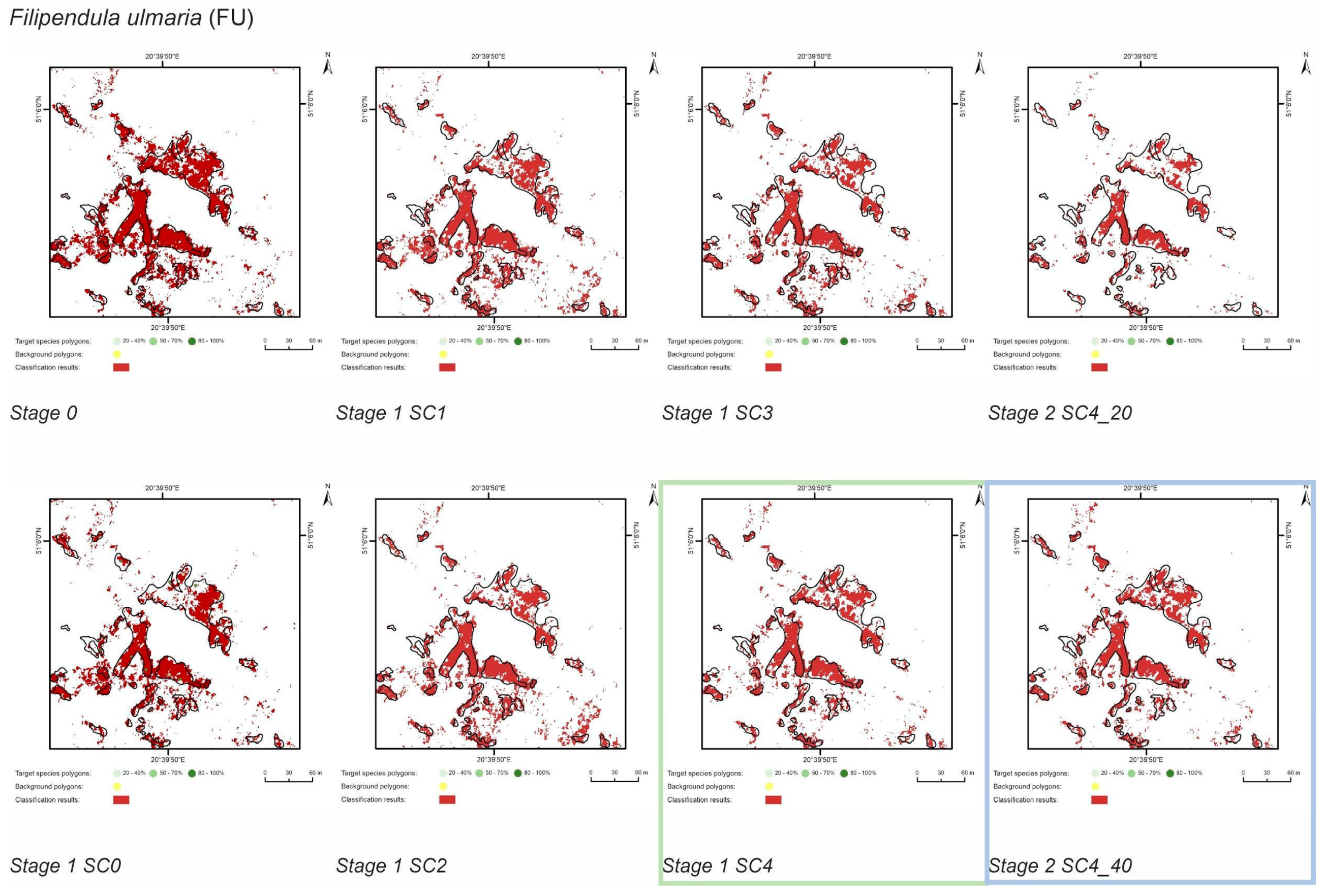
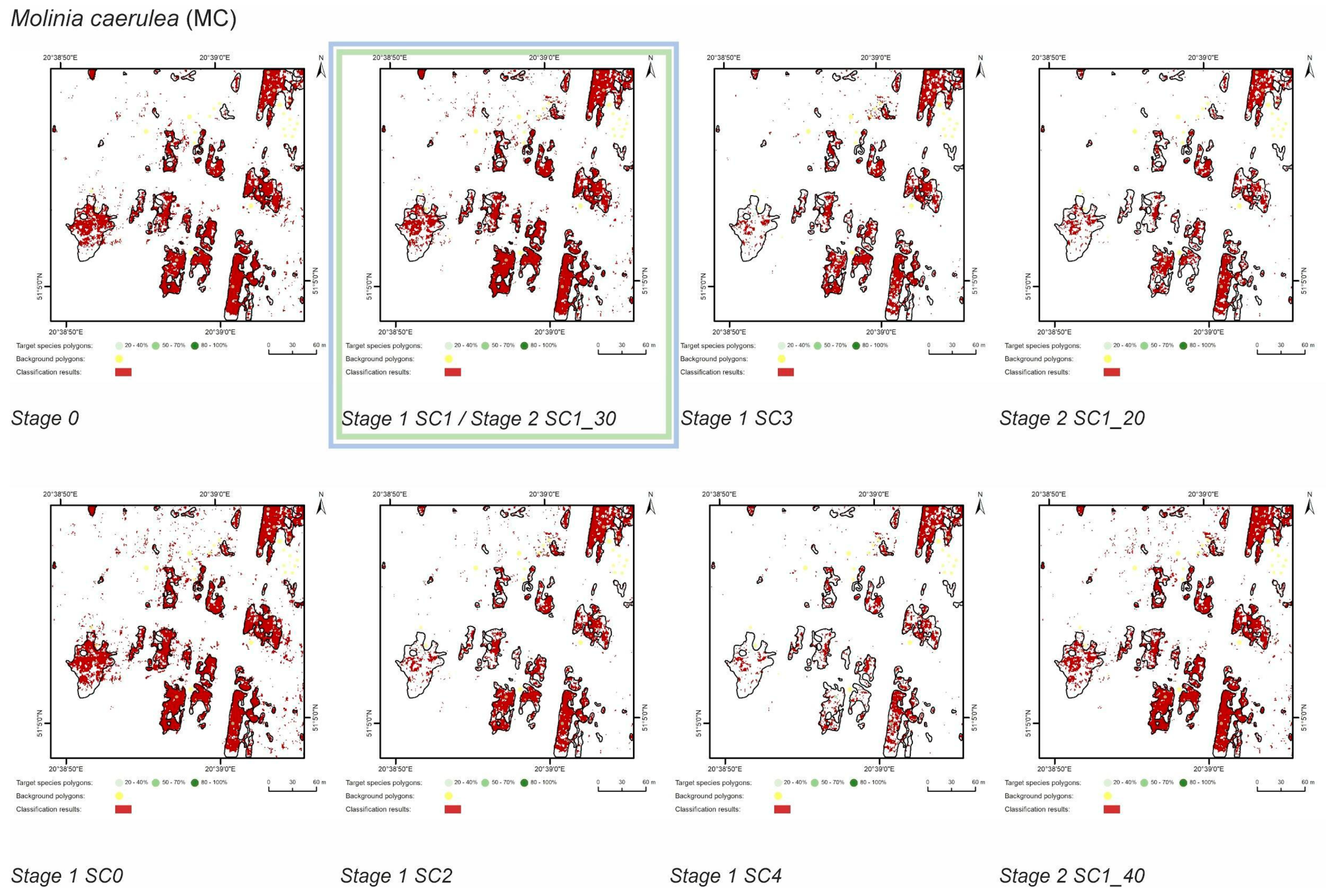
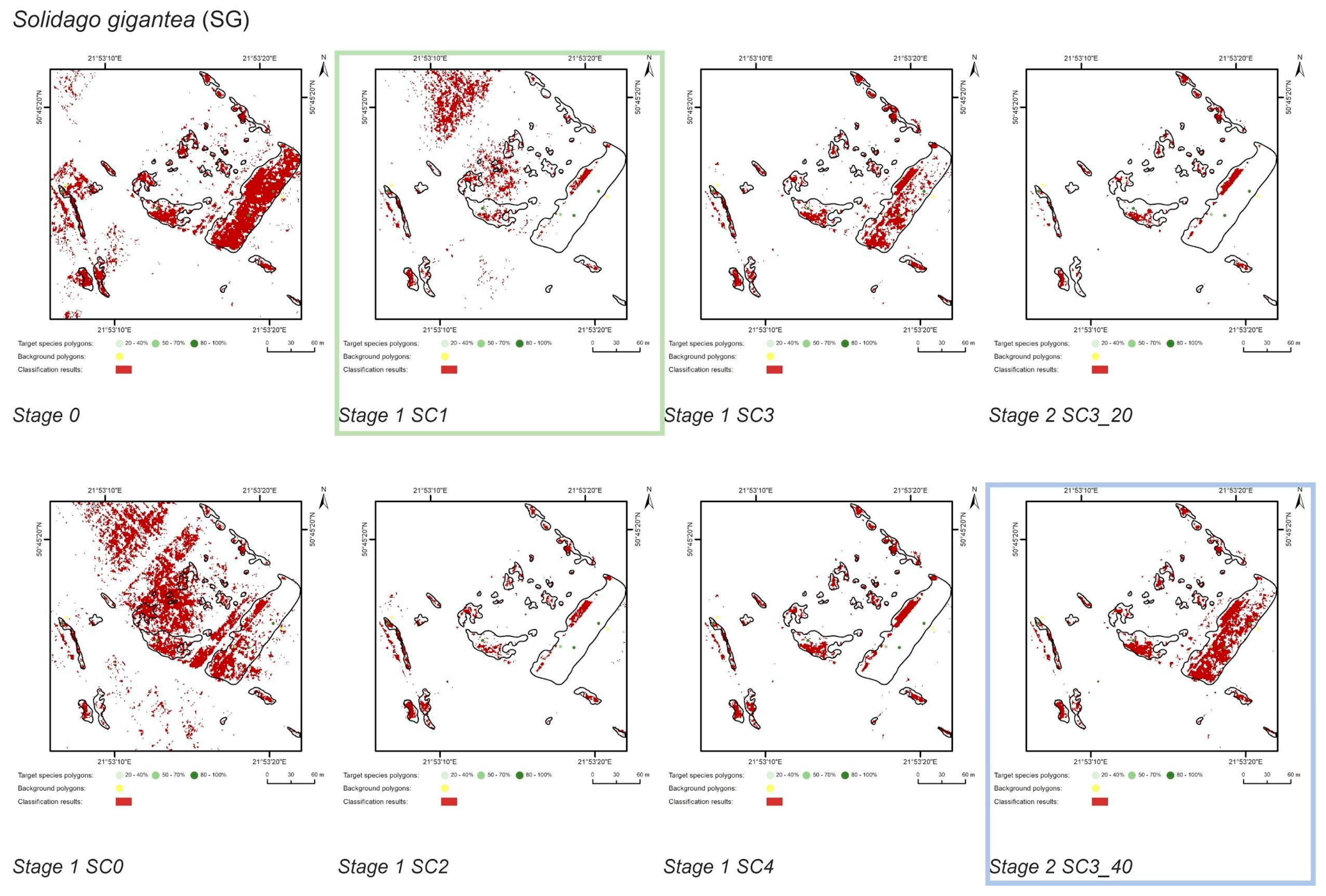
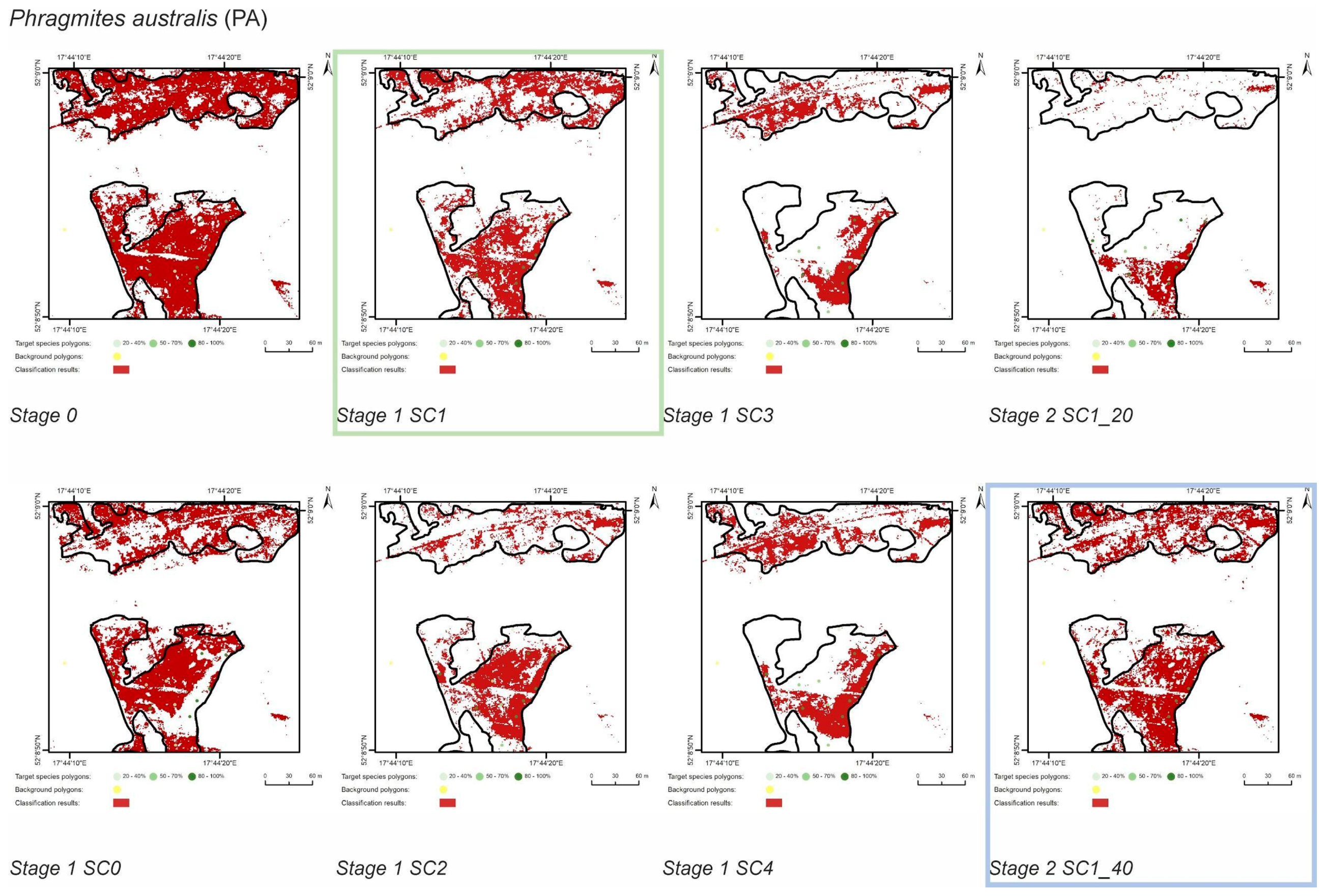
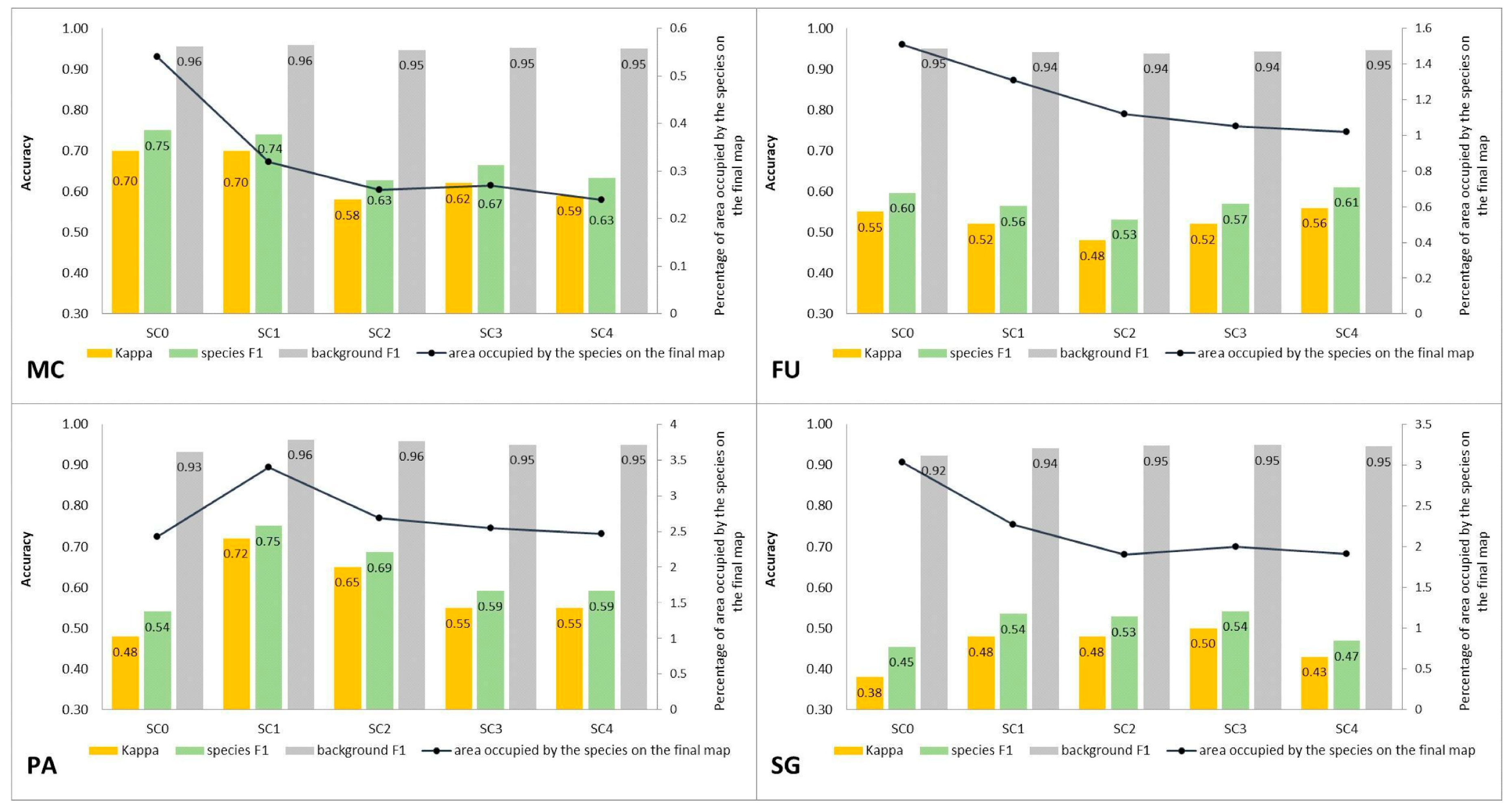
3.1. Classification—Simply Approach (STAGE 0)
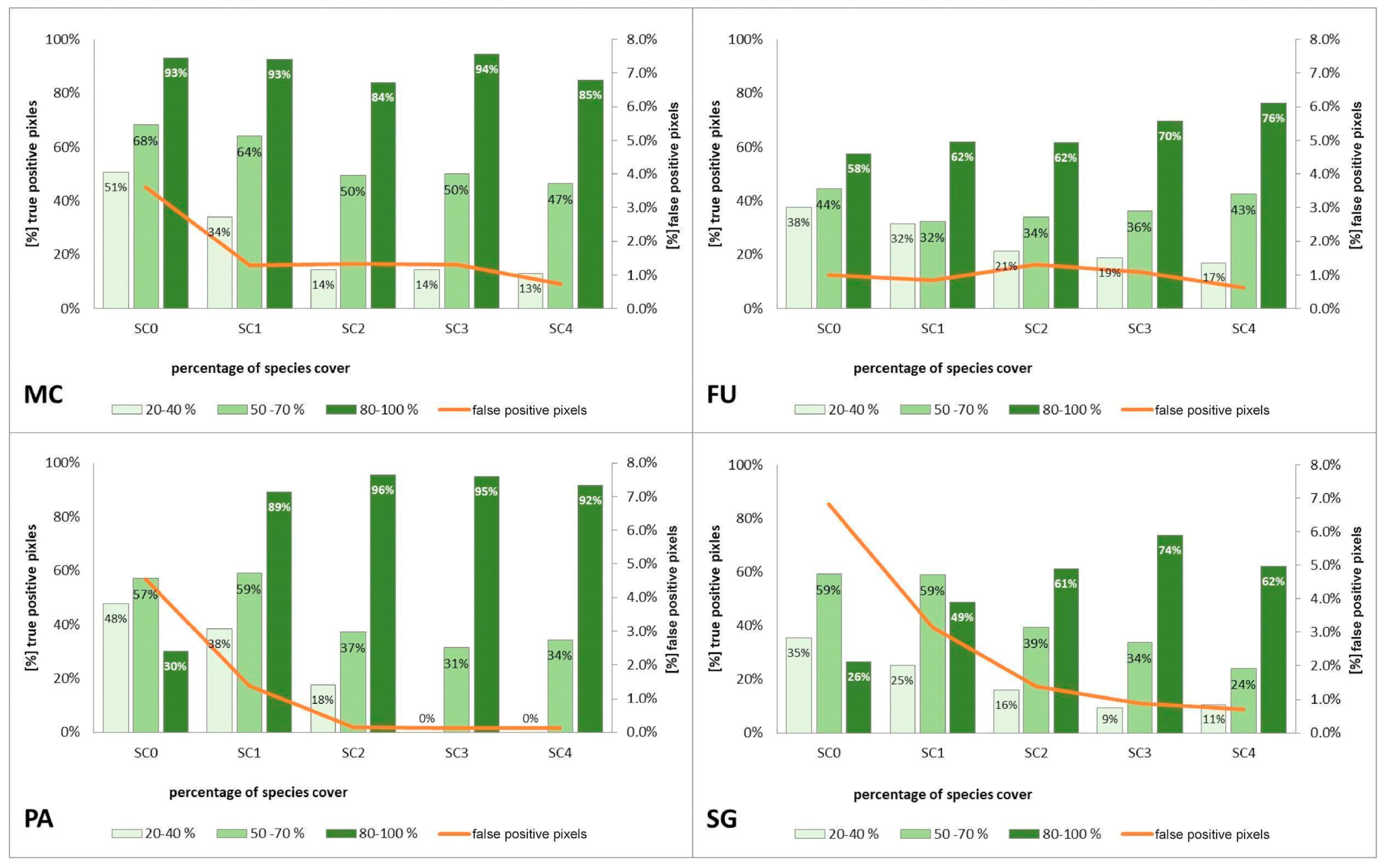
3.2. Classification—Various Cover of Target Species in Training Polygons (STAGE 1)
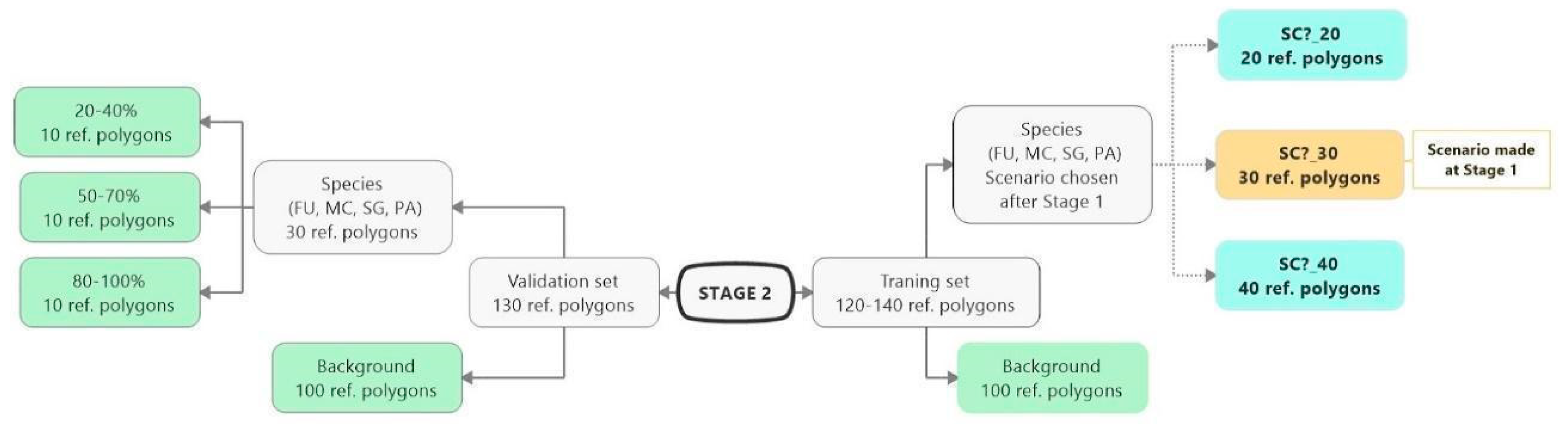
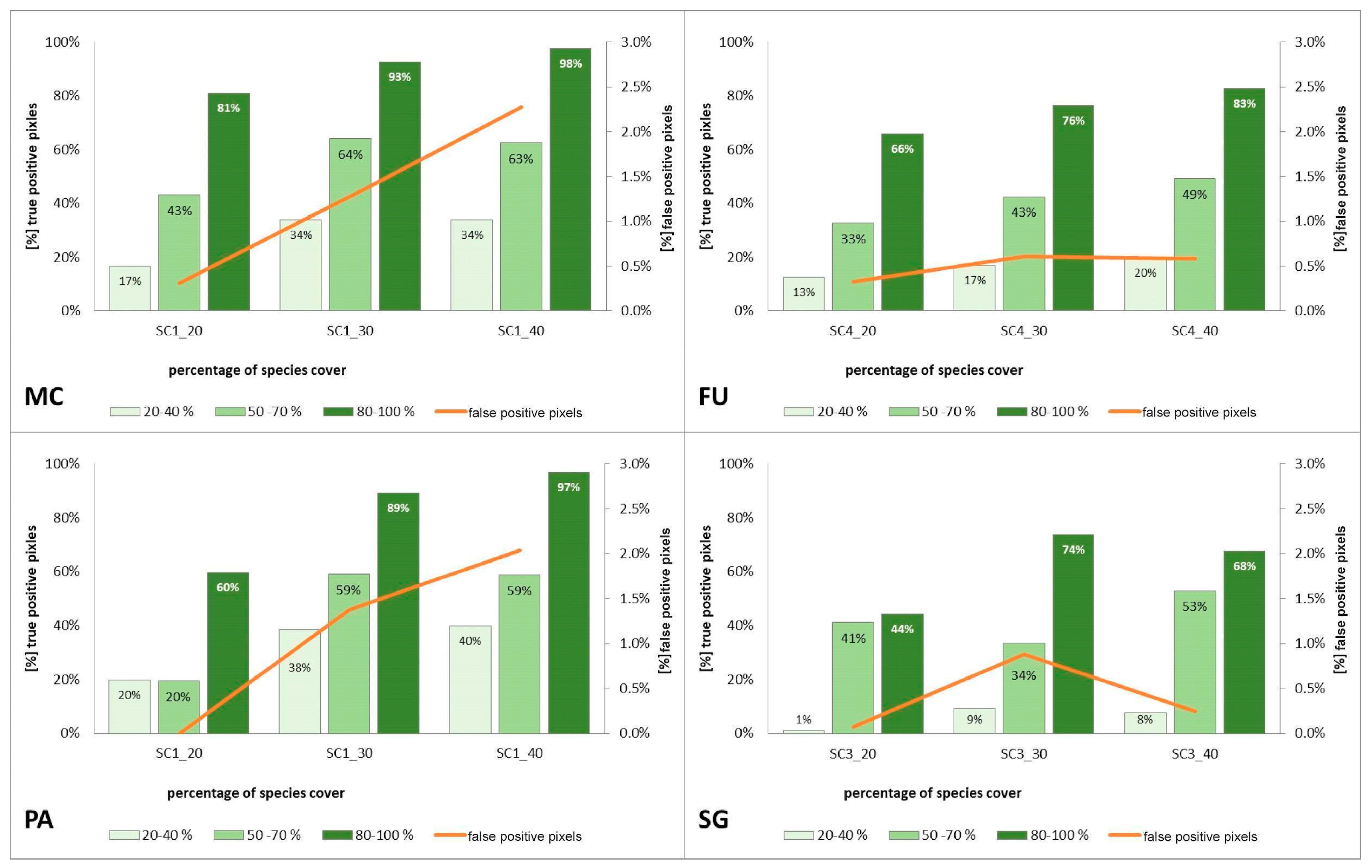
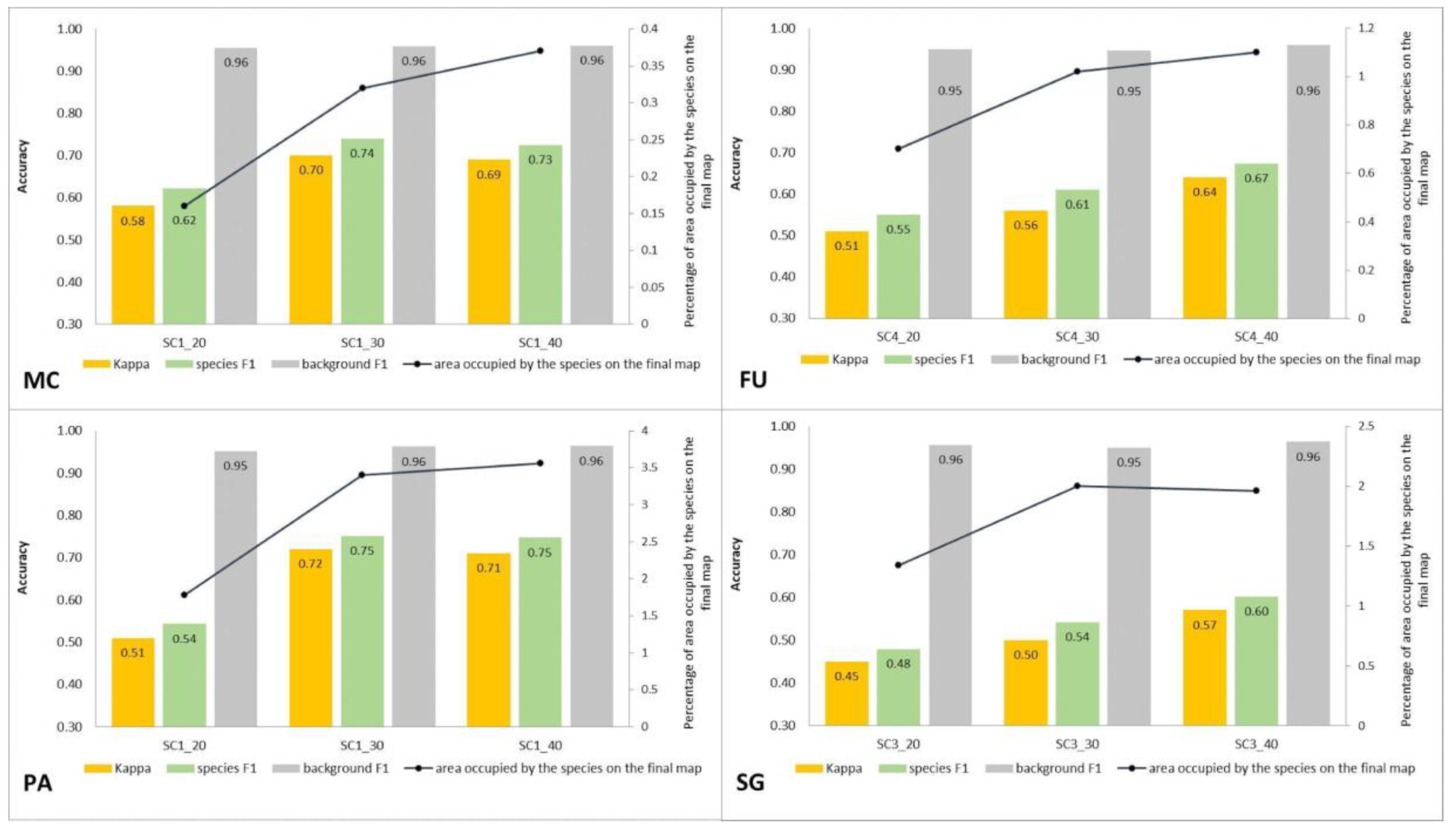
3.3. Classification—Various Number of Target Species Training Polygons (STAGE 2)
4. Discussion
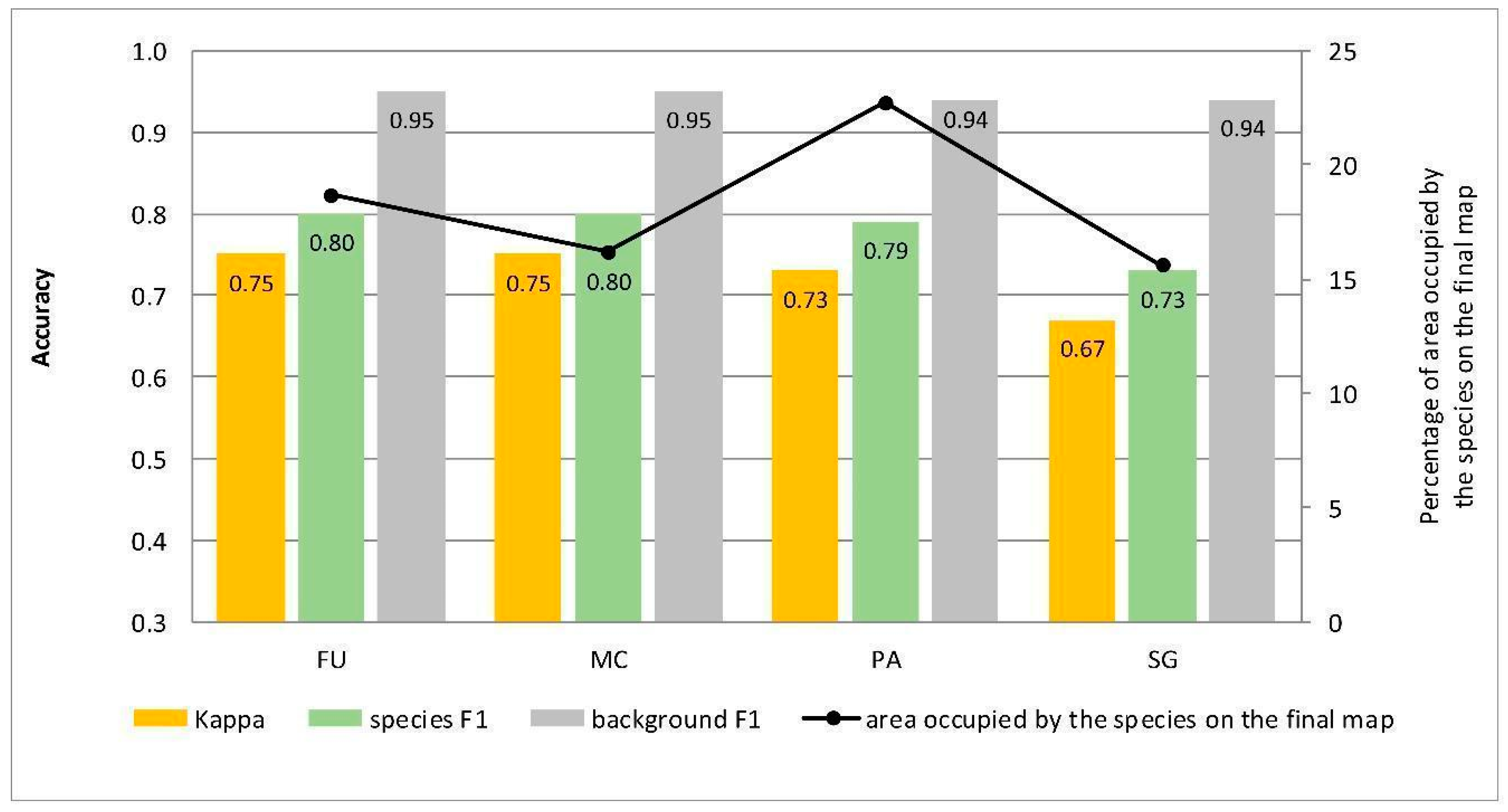
4.1. Effect of the Species Percentage Cover in the Training Dataset on the Classification Results
4.2. Percentage Cover of Target Species that Enables Its Identification
4.3. Impact of the Number of Target Species Training Polygons on the Result of Classification
4.4. Applicability of the Obtained Results
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Carey, M.P.; Sanderson, B.L.; Barnas, K.A.; Olden, J.D. Native invaders—Challenges for science, management, policy, and society. Front. Ecol. Environ. 2012, 10, 373–381. [Google Scholar] [CrossRef]
- Bradley, B.A. Remote detection of invasive plants: A review of spectral, textural and phenological approaches. Biol. Invasions 2014, 16, 1411–1425. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Asner, G.P. Applications of remote sensing to alien invasive plant studies. Sensors 2009, 9, 4869–4889. [Google Scholar] [CrossRef] [PubMed]
- Joshi, C.M.; de Leeuw, J.; van Duren, I.C. Remote sensing and GIS applications for mapping and spatial modelling of invasive species. In Proceedings of the ISPRS Congress: Geo-Imagery Bridging Continents 2004, Istanbul, Turkey, 12–23 July 2004. ISPRS 35, B7. [Google Scholar]
- Skowronek, S.; Asner, G.P.; Feilhauer, H. Performance of one-class classifiers for invasive species mapping using airborne imaging spectroscopy. Ecol. Inform. 2017, 37, 66–76. [Google Scholar] [CrossRef]
- MGGP Aero. Auxiliary Work in WP6 under the Programme “Natural Environment, Agriculture and Forestry” BIOSTRATEG II.: The Innovative Approach Supporting Monitoring of Non-Forest Natura 2000 Habitats, Using Remote Sensing Methods (HabitARS); MGGP Aero: Warszawa, Poland, 2016. [Google Scholar]
- Müllerová, J.; Brůna, J.; Bartaloš, T.; Dvořák, P.; Vítková, M.; Pyšek, P. Timing is important: Unmanned aircraft vs. Satellite imagery in plant invasion monitoring. Front. Plant Sci. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Niphadkar, M.; Nagendra, H. Remote sensing of invasive plants: Incorporating functional traits into the picture. Int. J. Remote Sens. 2016, 37, 3074–3085. [Google Scholar] [CrossRef]
- Baldeck, C.A.; Asner, G.P. Improving remote species identification through efficient training data collection. Remote Sens. 2014, 6, 2682–2698. [Google Scholar] [CrossRef]
- Aneece, I.; Epstein, H. Identifying invasive plant species using field spectroscopy in the VNIR region in successional systems of north-central Virginia. Int. J. Remote Sens. 2017, 38, 100–122. [Google Scholar] [CrossRef]
- Dubula, B.; Tesfamichael, S.G.; Rampedi, I.T. Assessing the potential of remote sensing to discriminate invasive Asparagus laricinus from adjacent land cover types. South Afr. J. Geomat. 2016, 5, 201–213. [Google Scholar] [CrossRef]
- Pelletier, C.; Valero, S.; Inglada, J.; Champion, N.; Sicre, C.M.; Dedieu, G. Effect of training class label noise on classification performances for land cover mapping with satellite image time series. Remote Sens. 2017, 9, 173. [Google Scholar] [CrossRef]
- Royimani, L.; Mutanga, O.; Odindi, J.; Dube, T.; Matongera, T.N. Advancements in satellite remote sensing for mapping and monitoring of alien invasive plant species (AIPs). Phys. Chem. Earth Parts A/B/C 2018. [Google Scholar] [CrossRef]
- Peerbhay, K.; Mutanga, O.; Ismail, R. The identification and remote detection of alien invasive plants in commercial forests: An Overview. S. Afr. J. Geomat. 2016, 5, 49. [Google Scholar] [Green Version]
- Kaneko, K.; Nohara, S. Review of effective vegetation mapping using the UAV (Unmanned Aerial Vehicle) method. J. Geogr. Inf. Syst. 2014, 06, 733–742. [Google Scholar] [CrossRef]
- Glenn, N.F.; Mundt, J.T.; Weber, K.T.; Prather, T.S.; Lass, L.W.; Pettingill, J. Hyperspectral data processing for repeat detection of small infestations of leafy spurge. Remote Sens. Environ. 2005, 95, 399–412. [Google Scholar] [CrossRef]
- Fassnacht, F.E.; Latifi, H.; Stereńczak, K.; Modzelewska, A.; Lefsky, M.; Waser, L.T.; Straub, C.; Ghosh, A. Review of studies on tree species classification from remotely sensed data. Remote Sens. Environ. 2016, 186, 64–87. [Google Scholar] [CrossRef]
- Pontius, R.G.; Millones, M. Death to Kappa: Birth of quantity disagreement and allocation disagreement for accuracy assessment. Int. J. Remote Sens. 2011, 32, 4407–4429. [Google Scholar] [CrossRef]
- Castaldi, F.; Chabrillat, S.; van Wesemael, B. Sampling strategies for soil property mapping using multispectral sentinel-2 and hyperspectral EnMAP satellite data. Remote Sens. 2019, 11, 309. [Google Scholar] [CrossRef]
- Stehman, S.V. Sampling designs for accuracy assessment of land cover. Int. J. Remote Sens. 2009, 30, 5243–5272. [Google Scholar] [CrossRef]
- Manolakis, D.; Shaw, G. Detection algorithms for hyperspectral Imaging applications. IEEE Signal Process. Mag. 2002, 19, 29–43. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudi’k, M.; Dudi’k, D.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Foody, G.M.; Mathur, A. Toward intelligent training of supervised image classifications: Directing training data acquisition for SVM classification. Remote Sens. Environ. 2004, 93, 107–117. [Google Scholar] [CrossRef]
- Millard, K.; Richardson, M. On the importance of training data sample selection in Random Forest image classification: A case study in peatland ecosystem mapping. Remote Sens. 2015, 7, 8489–8515. [Google Scholar] [CrossRef]
- Mather, P.M. Computer Processing of Remotely-Sensed Images: An Introduction; John Wiley: Chichester, UK, 1999. [Google Scholar]
- Pyšek, P.; Richardson, D.M.; Rejmánek, M.; Webster, G.L.; Williamson, M.; Kirschner, J. Alien plants in checklists and floras: Towards better communication between taxonomists and ecologists. Taxon 2004, 53, 131–143. [Google Scholar] [CrossRef]
- Ball, P.W.; Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A.; Chater, A.O.; DeFilipps, R.A.; et al. Flora Europaea; Cambridge University Press: Cambridge, UK, 1972; Volume 3, pp. 6–7. [Google Scholar]
- Taylor, K.; Rowland, A.P.; Jones, H.E. Molinia caerulea (L.) Moench. J. Ecol. 2001, 89, 126–144. [Google Scholar] [CrossRef]
- Shaltout, K.H.; Al-sodany, Y.; Eid, E.M. Biology of Common Reed Phragmites Review and Inquiry; Overview Series, Assiut University Center for Environmental Studies (AUCES): Assiut, Egypt, 2006. [Google Scholar]
- Täckholm, V.; Täckholm, G.; Drar, M. Flora of Egypt; 19. Phragmites. Bulletin of the Faculty of Science 17; Fouad I University: Giza, Egypt, 1941; Volume 1, pp. 209–216. [Google Scholar]
- Holm, L.G.; Plucknett, D.L.; Pancho, J.V.; Herberger, J.P. Phragmites australis (Cav.) Trin. (= P. communis Trin.) and Phragmites karka (Retz.) Trin. In The World’s Worst Weeds “Distribution and Biology”; The University Press of Hawaii: Honolulu, HI, USA, 1977; 609p. [Google Scholar]
- Weber, E.; Jacobs, G. Biological flora of central Europe: Solidago gigantea Aiton. Flora 2005, 200, 109–118. [Google Scholar] [CrossRef]
- Capek, M. The possibility of biological control of imported weeds of the genus Solidago L. in Europe. Acta Inst. For. Zvolensis 1971, 9, 429–441. [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Dull, R.; Wirth, V.; Werner, W.; Paulissen, D. Zeigerwerte von Pflanzen in Mitteleuropa, Scripta Geobotanica; Erich Goltze: Göttingen, Germany, 2001; Volume 18. [Google Scholar]
- Voser-Huber, M.L. Studien an eingeburgerten arten der gattung solidago L. Dissertat. Botan. 1983, 68, 1–97. [Google Scholar]
- Botta-Dukát, Z. Morphological plasticity in the rhizome system of Solidago gigantea (Asteraceae): Comparison of populations in a wet and a dry habitat. Acta Bot. Hung. 2016, 58, 227–240. [Google Scholar] [CrossRef] [Green Version]
- Hestir, E.L.; Khanna, S.; Andrew, M.E.; Santos, M.J.; Viers, J.H.; Greenberg, J.A.; Rajapakse, S.S.; Ustin, S.L. Identification of invasive vegetation using hyperspectral remote sensing in the California Delta ecosystem. Remote Sens. Environ. 2008, 112, 4034–4047. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Cutler, D.R.; Edwards, T.C.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, R.L.; Wood, S.D.; Sheley, R.L. Mapping invasive plants using hyperspectral imagery and Breiman Cutler classifications (randomForest). Remote Sens. Environ. 2006, 100, 356–362. [Google Scholar] [CrossRef]
- Congalton, R.G.; Green, K. Assessing the Accuracy of Remotely Sensed Data: Principles and Practices; CRC Press: Boca Raton, FL, USA, 1999; pp. 105–110. [Google Scholar]
- Lillesand, T.; Kiedfer, R.; Chipman, J. Remote Sensing and Image Interpretation, 6th ed.; John Wiley and Sons, Inc.: New York, NY, USA, 2008; pp. 1–804. [Google Scholar]
- Vegetation Classification Studio Software, Version 2.13/hb. Available online: http://www.definity.pl/vcs (accessed on 12 March 2019).
- Ustin, S.L.; DiPietro, D.; Olmstead, K.; Underwood, E.; Scheer, G.J. Hyperspectral remote sensing for invasive species detection and mapping. IEEE Int. Geosci. Remote Sens. Symp. 2002, 3, 1658–1660. [Google Scholar]
- Dorigo, W.; Lucieer, A.; Podobnikar, T.; Carni, A. Mapping invasive Fallopia japonica by combined spectral, spatial, and temporal analysis of digital orthophotos. Int. J. Appl. Earth Obs. Geoinf. 2012, 19, 185–195. [Google Scholar] [CrossRef]
- Mirik, M.; Ansley, R.J.; Steddom, K.; Jones, D.C.; Rush, C.M.; Michels, G.J.; Elliott, N.C. Remote distinction of a noxious weed (Musk Thistle: Carduus Nutans) using airborne hyperspectral imagery and the support vector machine classifier. Remote Sens. 2013, 5, 612–630. [Google Scholar] [CrossRef]
- Underwood, E.; Ustin, S.; Dipietro, D. Mapping Non-Native Plants Using Hyperspectral Imagery. Remote Sens. Environ. 2003, 86, 150–161. [Google Scholar] [CrossRef]
- Ishii, J.; Washitani, I. Early detection of the invasive alien plant Solidago altissima in moist tall grassland using hyperspectral imagery. Int. J. Remote Sens. 2013, 34, 5926–5936. [Google Scholar] [CrossRef]
- Evangelista, P.H.; Stohlgren, T.J.; Morisette, J.T.; Kumar, S. Mapping invasive tamarisk (Tamarix): A comparison of single-scene and time-series analyses of remotely sensed data. Remote Sens. 2009, 1, 519–533. [Google Scholar] [CrossRef]
- Vilà, M.; Schaffner, U.; Pyšek, P.; Pergl, J.; Jarošík, V.; Hulme, P.E.; Hejda, M. A global assessment of invasive plant impacts on resident species, communities and ecosystems: The interaction of impact measures, invading species’ traits and environment. Glob. Chang. Biol. 2011, 18, 1725–1737. [Google Scholar]
- Millard, K.; Richardson, M. Wetland mapping with LiDAR derivatives, SAR polarimetric decompositions, and LiDAR-SAR fusion using a random forest classifier. Can. J. Remote Sens. 2013, 39, 290–307. [Google Scholar] [CrossRef]
- Peerbhay, K.; Mutanga, O.; Lottering, R.; Bangamwabo, V.; Ismail, R. Detecting bugweed (Solanum mauritianum) abundance in plantation forestry using multisource remote sensing. ISPRS J. Photogramm. Remote Sens. 2016, 121, 167–176. [Google Scholar] [CrossRef]
- Barbosa, J.M.; Asner, G.P.; Martin, R.E.; Baldeck, C.A.; Hughes, F.; Johnson, T. Determining subcanopy Psidium cattleianum invasion in Hawaiian forests using imaging spectroscopy. Remote Sens. 2016, 8, 33. [Google Scholar] [CrossRef]
- De Castro, A.I.; Jurado-Expósito, M.; Peña-Barragán, J.M.; López-Granados, F. Airborne multi-spectral imagery for mapping cruciferous weeds in cereal and legume crops. Precis. Agric. 2012, 13, 302–321. [Google Scholar] [CrossRef]
- Raczko, E.; Zagajewski, B. Comparison of support vector machine, random forest and neural network classifiers for tree species classification on airborne hyperspectral APEX images. Eur. J. Remote Sens. 2017, 50, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Marcinkowska-Ochtyra, A.; Jarocińska, A.; Bzdęga, K.; Tokarska-Guzik, B. Classification of expansive grassland species in different growth stages based on hyperspectral and LiDAR data. Remote Sens. 2018, 10, 2019. [Google Scholar] [CrossRef]
- Mundt, J.T.; Glenn, N.F.; Weber, K.T.; Prather, T.S.; Lass, L.W.; Pettingill, J. Discrimination of hoary cress and determination of its detection limits via hyperspectral image processing and accuracy assessment techniques. Remote Sens. Environ. 2005, 96, 509–517. [Google Scholar] [CrossRef]
- Schmidt, J.; Fassnacht, F.E.; Förster, M.; Schmidtlein, S. Synergetic use of Sentinel-1 and Sentinel-2 for assessments of heathland conservation status. Remote Sens. Ecol. Conserv. 2017, 4, 225–239. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Liaw, A.; Breiman, L. Using Random Forest to Learn Imbalanced Data; University of California: Berkeley, CA, USA, 2004; pp. 1–12. [Google Scholar]
- He, K.S.; Rocchini, D.; Neteler, M.; Nagendra, H. Benefits of hyperspectral remote sensing for tracking plant invasions. Divers. Distrib. 2011, 17, 381–392. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area | No. 1 | No. 1 | No. 2 | No. 3 |
|---|---|---|---|---|
| Target species | Molinia caerulea (MC) | Filipendula ulmaria (FU) | Phragmites australis (PA) | Solidago gigantea (SG) |
| Flight dates and start of field sampling | 27 September 2017 | 07 July 2017 | 16 July 2017 | 09 September 2017 |
| End of field sampling | 29 September 2017 | 13 July 2017 | 18 July 2017 | 19 September 2017 |
| Dominant development phase of target species | fruiting | flowering | flowering | flowering |
| Phenological and structural traits of plants during acquisition | Plants form a dense tussock, floral stems become pinkish–orange as the leaves turn yellow | Plants form a bushy clump, bearing sprays of creamy–white flowers | Plants form dense stands that include flowering bushy panicles and standing dead stems from previous year’s growth | Plants form a bushy mound of deep-green leaves, bearing large clusters of golden-yellow flowers |
| Number of established target species reference polygons | 110 | 110 | 110 | 110 |
| Number of established background polygons | 200 | 200 | 200 | 200 |
| Data collection area [km2] | 40.59 | 40.59 | 10.37 | 35.45 |
| Number of flight lines | 26 | 25 | 11 | 20 |
| Orientation of flight | NS | WE | WE | NS |
| Sensor Type | Data Parameters | Flight Lines Overlap | Swath Width |
|---|---|---|---|
| HySpex VNIR-1800 0.4–0.9 µm | GSD 0.49 [m] | 35 [%] | 440 m |
| HySpex SWIR-384 0.9–2.5 µm | GSD 1.07 [m] | 30 [%] | 410 m |
| Number of the Target Species Reference Polygons and Background Reference Polygons | ||
|---|---|---|
| Percentage Cover of the Target Species | ||
| 20–40% | 50–70% | 80–100% |
| 30 | 30 | 50 |
| Target Species | Stage and Scenario | Control Area of Target Species [m2] | Control Area of Back-Ground [m2] | True Positive [m2] | False Positive [m2] | True Negative [m2] | False Negative [m2] | True Positive [%] | False Positive [%] | True Negative [%] | False Negative [%] |
|---|---|---|---|---|---|---|---|---|---|---|---|
| MC | Stage 0 | 14189 | 85667 | 9784 | 1946 | 83739 | 4387 | 69 | 14 | 98 | 31 |
| Stage 1 SC0 | 10840 | 3449 | 82236 | 3331 | 76 | 24 | 96 | 23 | |||
| Stage 1 SC1 | 9812 | 1774 | 83911 | 4359 | 69 | 13 | 98 | 31 | |||
| Stage 1 SC2 | 7578 | 878 | 84807 | 6593 | 53 | 6 | 99 | 46 | |||
| Stage 1 SC3 | 6953 | 694 | 84991 | 7218 | 49 | 5 | 99 | 51 | |||
| Stage 1 SC4 | 5102 | 460 | 85225 | 9069 | 36 | 3 | 99 | 64 | |||
| Stage 2 SC1_20 | 6123 | 402 | 85283 | 8048 | 43 | 3 | 100 | 57 | |||
| Stage 2 SC1_30 | 9812 | 1774 | 83911 | 4359 | 69 | 13 | 98 | 31 | |||
| Stage 2 SC1_40 | 9343 | 1737 | 83948 | 4828 | 66 | 12 | 98 | 34 | |||
| FU | Stage 0 | 10797 | 89203 | 8134 | 5910 | 83149 | 2663 | 75 | 55 | 93 | 25 |
| Stage 1 SC0 | 6486 | 4406 | 84653 | 4311 | 60 | 41 | 95 | 40 | |||
| Stage 1 SC1 | 7102 | 5006 | 84053 | 3695 | 66 | 46 | 94 | 34 | |||
| Stage 1 SC2 | 6679 | 4590 | 84469 | 4118 | 62 | 43 | 95 | 38 | |||
| Stage 1 SC3 | 6287 | 3454 | 85605 | 4510 | 58 | 32 | 96 | 42 | |||
| Stage 1 SC4 | 6457 | 3126 | 85933 | 4340 | 60 | 29 | 96 | 40 | |||
| Stage 2 SC4_20 | 4684 | 1482 | 87577 | 6113 | 43 | 14 | 98 | 57 | |||
| Stage 2 SC4_30 | 6457 | 3126 | 85933 | 4340 | 60 | 29 | 96 | 40 | |||
| Stage 2 SC4_40 | 6453 | 3057 | 86002 | 4344 | 60 | 28 | 96 | 40 | |||
| SG | Stage 0 | 11477 | 88523 | 7076 | 3398 | 84981 | 4401 | 62 | 30 | 96 | 38 |
| Stage 1 SC0 | 4208 | 10431 | 77948 | 7269 | 37 | 91 | 88 | 63 | |||
| Stage 1 SC1 | 1488 | 4128 | 84251 | 9989 | 13 | 36 | 95 | 87 | |||
| Stage 1 SC2 | 1916 | 581 | 87798 | 9561 | 17 | 5 | 99 | 83 | |||
| Stage 1 SC3 | 4061 | 665 | 87714 | 7416 | 35 | 6 | 99 | 65 | |||
| Stage 1 SC4 | 2575 | 568 | 87811 | 8902 | 22 | 5 | 99 | 78 | |||
| Stage 2 SC3_20 | 1636 | 211 | 88168 | 9841 | 14 | 2 | 100 | 86 | |||
| Stage 2 SC3_30 | 4061 | 665 | 87714 | 7416 | 35 | 6 | 99 | 65 | |||
| Stage 2 SC3_40 | 4770 | 638 | 87741 | 6707 | 42 | 6 | 99 | 58 | |||
| PA | Stage 0 | 33106 | 66894 | 24417 | 3534 | 63216 | 8689 | 74 | 11 | 95 | 26 |
| Stage 1 SC0 | 18279 | 3409 | 63341 | 14827 | 55 | 10 | 95 | 45 | |||
| Stage 1 SC1 | 15426 | 1700 | 65050 | 17680 | 47 | 5 | 97 | 53 | |||
| Stage 1 SC2 | 10979 | 657 | 66093 | 22127 | 33 | 2 | 99 | 67 | |||
| Stage 1 SC3 | 8604 | 356 | 66394 | 24502 | 26 | 1 | 99 | 74 | |||
| Stage 1 SC4 | 10450 | 473 | 66277 | 22656 | 32 | 1 | 99 | 68 | |||
| Stage 2 SC1_20 | 3904 | 315 | 66435 | 29202 | 12 | 1 | 99 | 88 | |||
| Stage 2 SC1_30 | 15426 | 1700 | 65050 | 17680 | 47 | 5 | 97 | 53 | |||
| Stage 2 SC1_40 | 16393 | 1532 | 65218 | 16713 | 50 | 5 | 97 | 50 |
| Target Species | RS Accuracy Measures (Kappa, F1) | Correctly Classified Species Pixels (%) | Compatibility with Field Mapping (Control Area) | Chosen Scenario |
|---|---|---|---|---|
| Molinia caerulea (MC) | SC1 | SC1 | SC1 | SC1 |
| Filipendula ulmaria (FU) | SC4 | SC4 | SC4 | SC4 |
| Solidago gigantea (SG) | SC3 | SC3 | SC3 | SC3 |
| Phragmites australis (PA) | SC1 | SC1 | SC1 | SC1 |
| Target Species | RS Accuracy Measures (Kappa, F1) | Correctly Classified Species Pixels (%) | Compatibility with Field Mapping | Chosen Scenario |
|---|---|---|---|---|
| Molinia caerulea (MC) | SC1_30 | SC1_40 | SC1_30 | SC1_30 |
| Filipendula ulmaria (FU) | SC4_40 | SC4_40 | SC4_40 | SC4_40 |
| Solidago gigantea (SG) | SC3_40 | SC3_40 | SC3_40 | SC3_40 |
| Phragmites australis (PA) | SC1_30 | SC1_40 | SC1_40 | SC1_40 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kopeć, D.; Zakrzewska, A.; Halladin-Dąbrowska, A.; Wylazłowska, J.; Kania, A.; Niedzielko, J. Using Airborne Hyperspectral Imaging Spectroscopy to Accurately Monitor Invasive and Expansive Herb Plants: Limitations and Requirements of the Method. Sensors 2019, 19, 2871. https://doi.org/10.3390/s19132871
Kopeć D, Zakrzewska A, Halladin-Dąbrowska A, Wylazłowska J, Kania A, Niedzielko J. Using Airborne Hyperspectral Imaging Spectroscopy to Accurately Monitor Invasive and Expansive Herb Plants: Limitations and Requirements of the Method. Sensors. 2019; 19(13):2871. https://doi.org/10.3390/s19132871
Chicago/Turabian StyleKopeć, Dominik, Agata Zakrzewska, Anna Halladin-Dąbrowska, Justyna Wylazłowska, Adam Kania, and Jan Niedzielko. 2019. "Using Airborne Hyperspectral Imaging Spectroscopy to Accurately Monitor Invasive and Expansive Herb Plants: Limitations and Requirements of the Method" Sensors 19, no. 13: 2871. https://doi.org/10.3390/s19132871
APA StyleKopeć, D., Zakrzewska, A., Halladin-Dąbrowska, A., Wylazłowska, J., Kania, A., & Niedzielko, J. (2019). Using Airborne Hyperspectral Imaging Spectroscopy to Accurately Monitor Invasive and Expansive Herb Plants: Limitations and Requirements of the Method. Sensors, 19(13), 2871. https://doi.org/10.3390/s19132871