Characterization of Simple and Double Yeast Cells Using Dielectrophoretic Force Measurement

 , ,
, ,
Abstract
:1. Introduction
2. Theoretical Model
2.1. Linear Relationship between Velocity and Position of a Single Cell
2.2. Linear Relationship Between Velocity and Position of a Coupled Cell
2.3. Mathematical Relationship between Velocity and Position of a Single and Coupled Cell
3. Materials and Methods
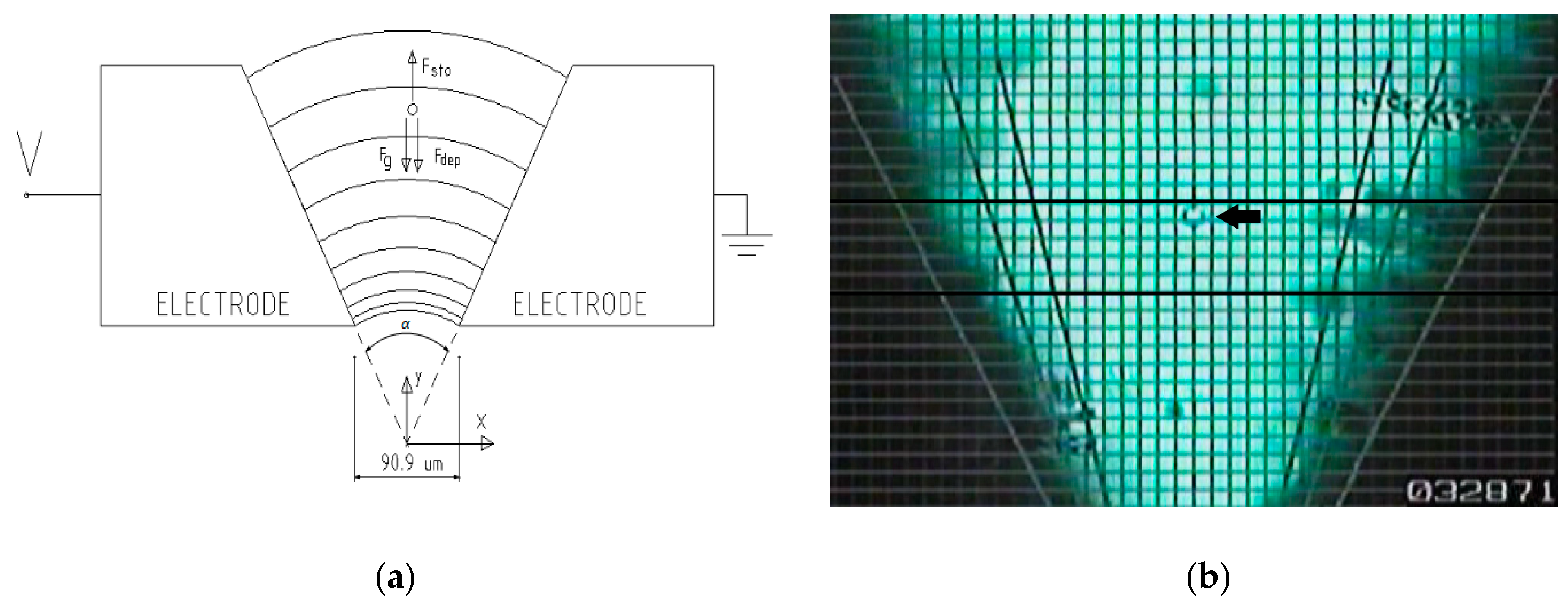
3.1. Dielectrophoretic Device
3.2. Cellular Suspension
3.3. Cell Radius Measurement
3.4. Experimental Procedure
4. Results
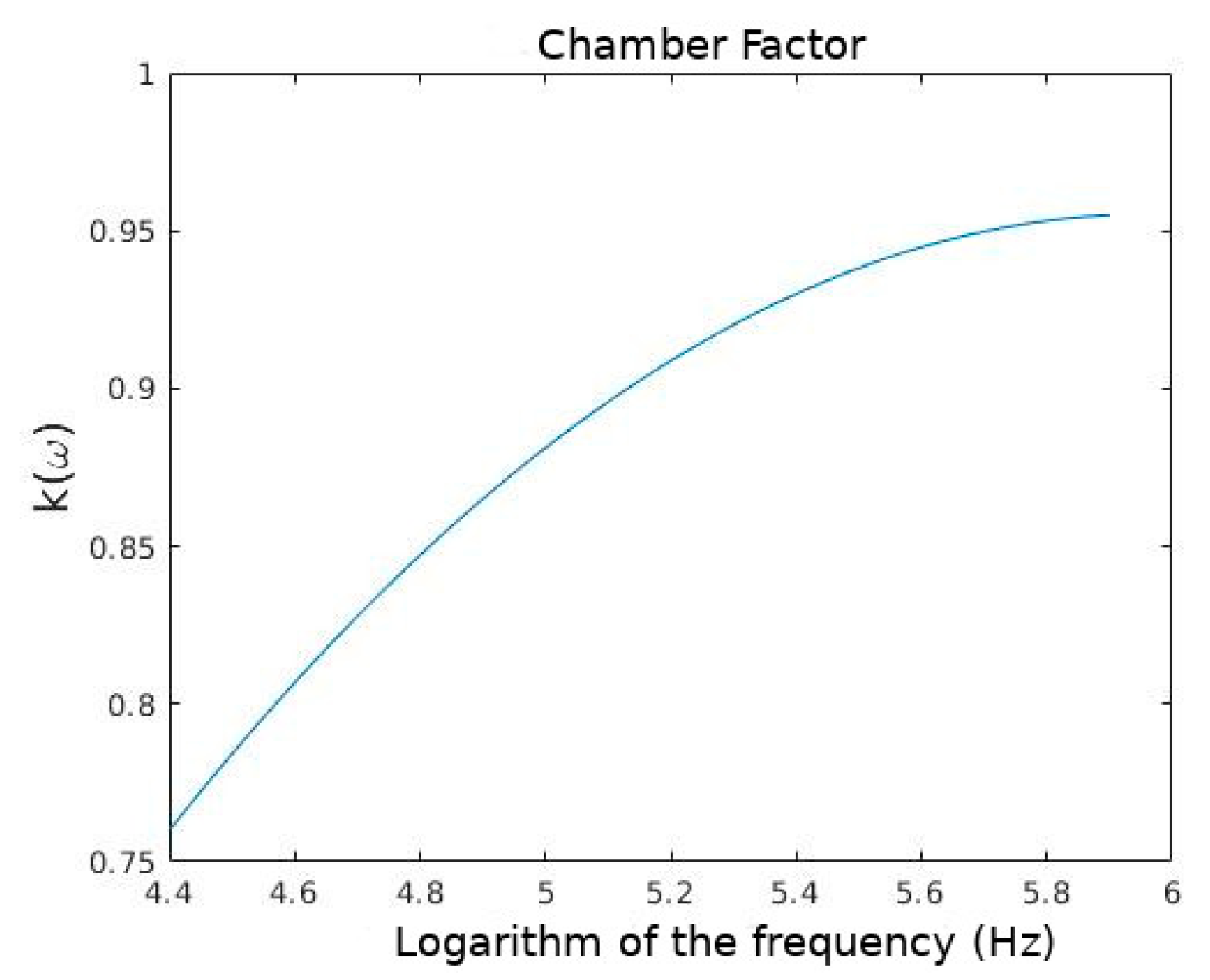
4.1. Measurement of the Electric Properties of the Dielectrophoretic Chamber
4.2. Cell Radius Measurement
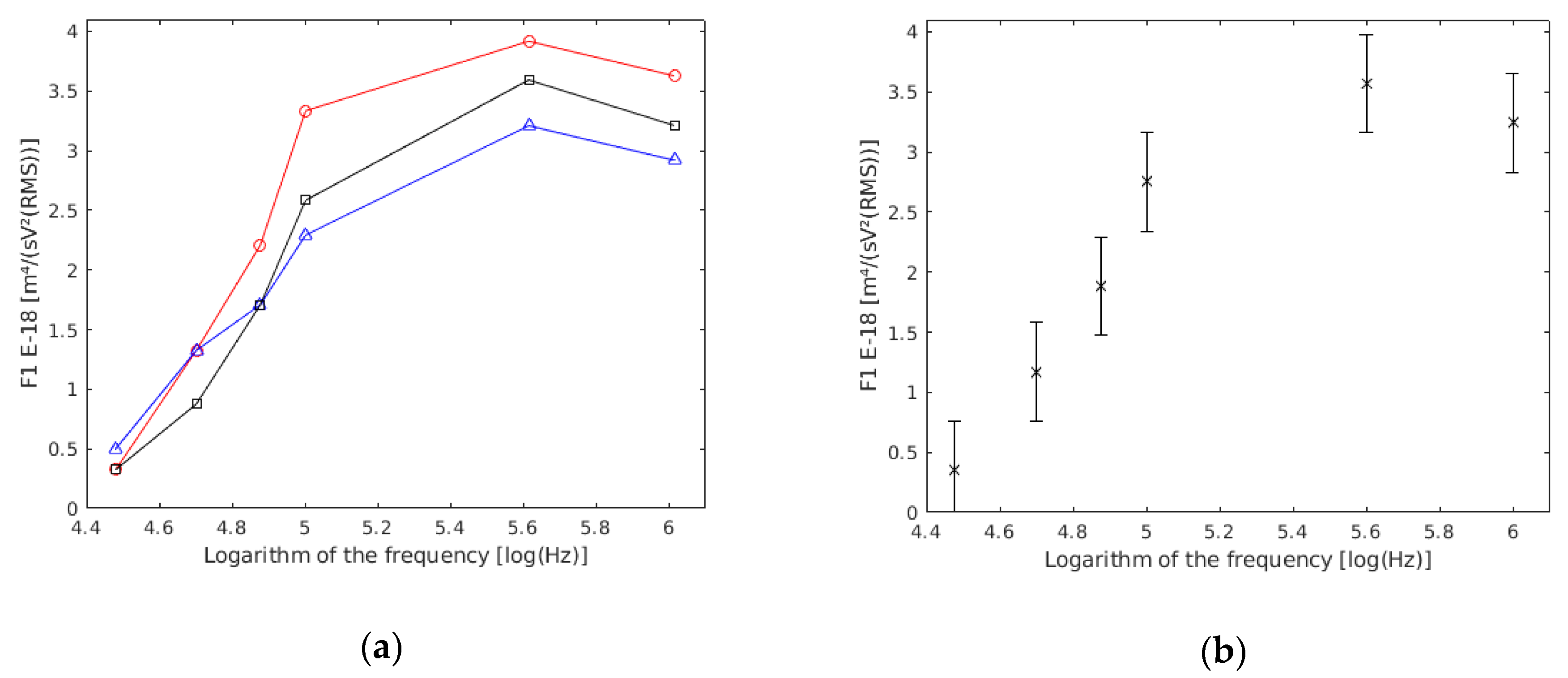
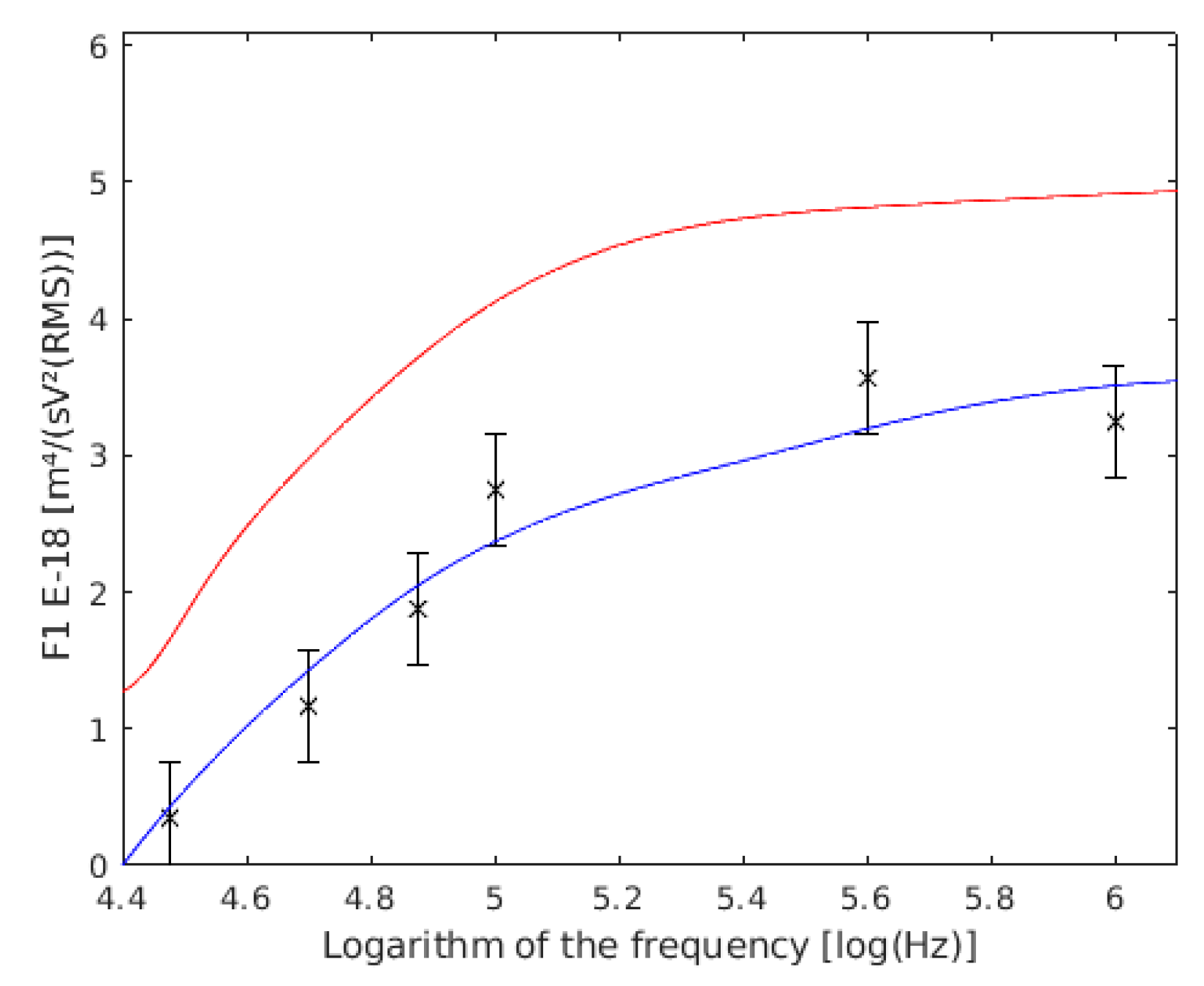
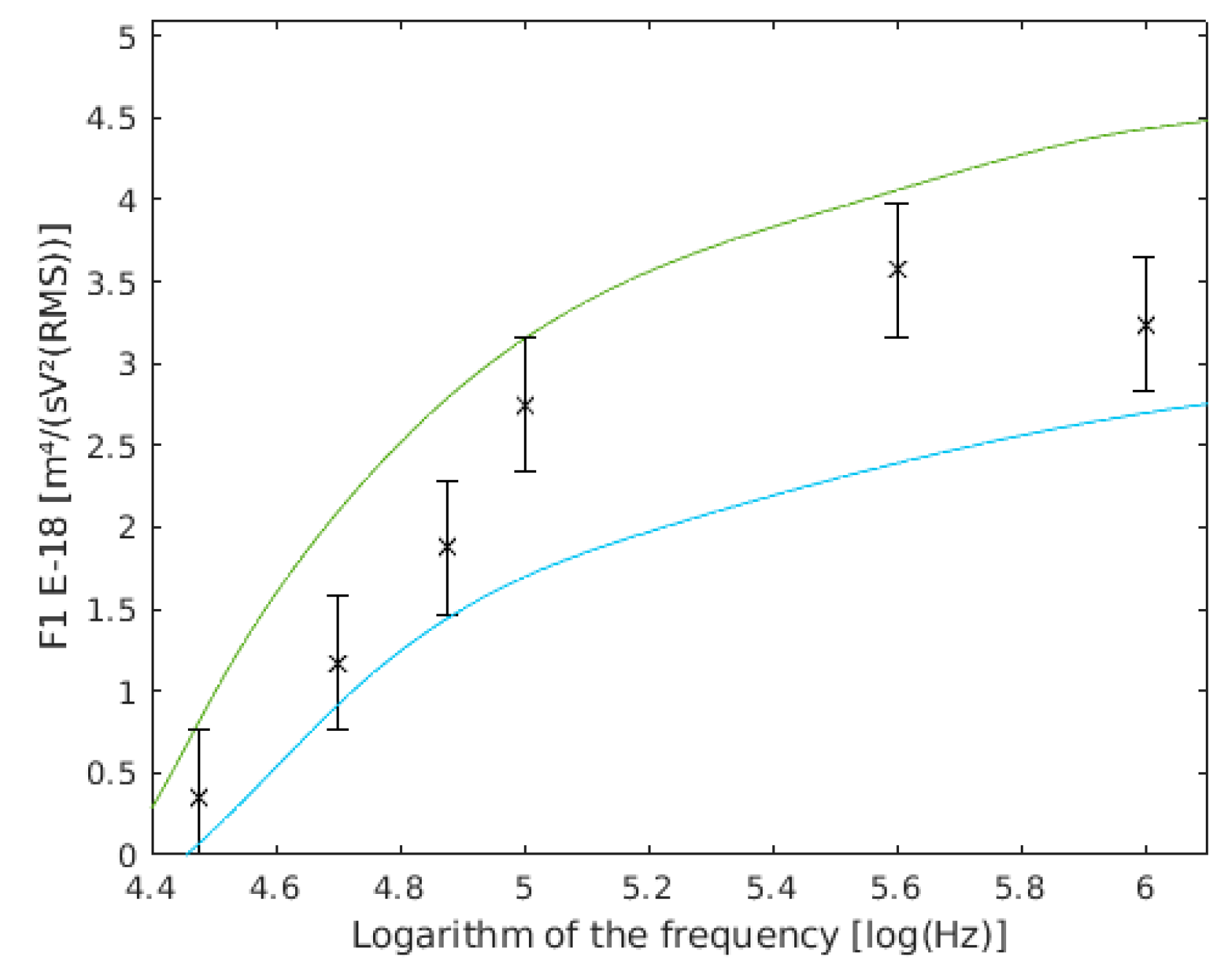
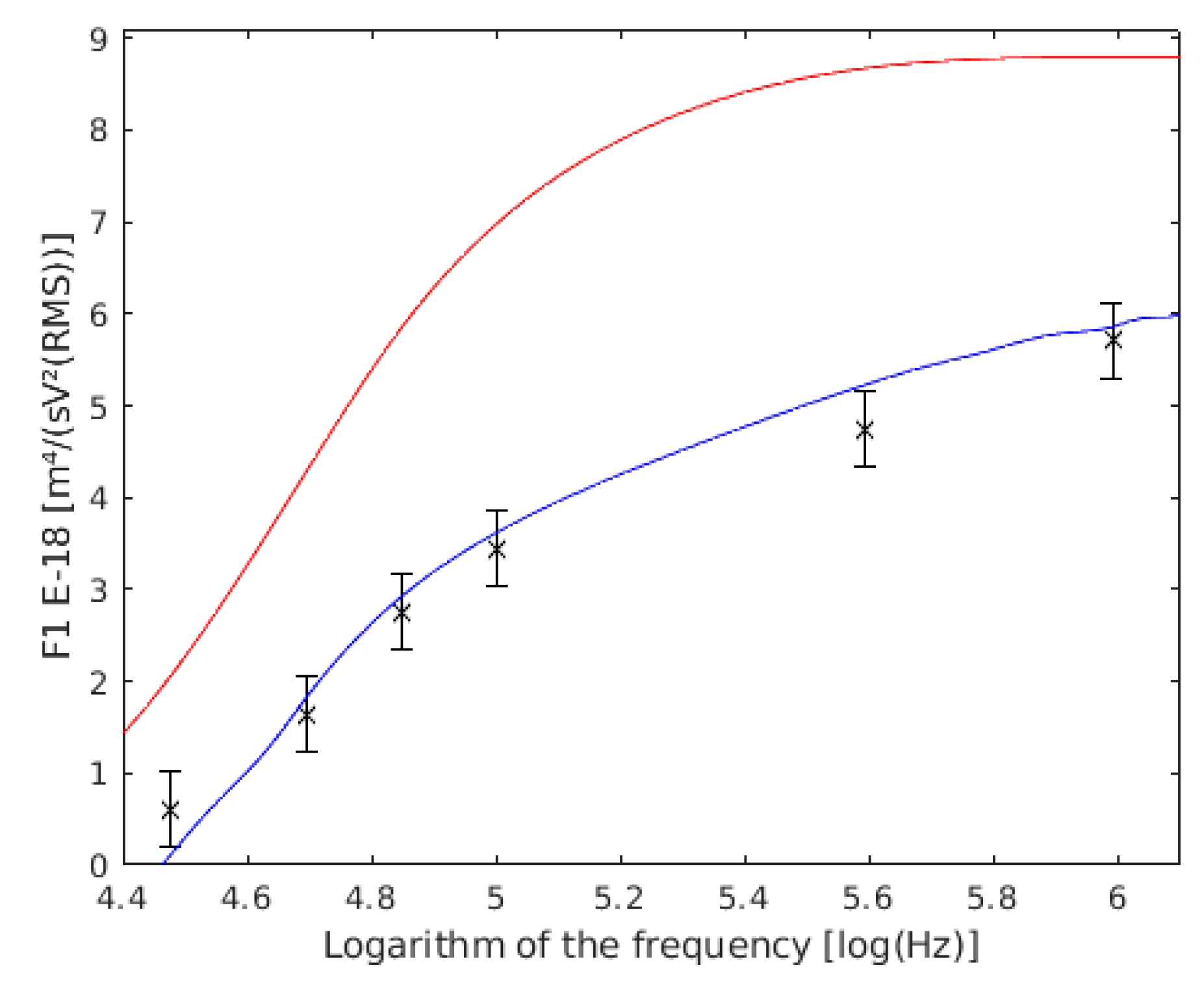
4.3. Single Yeast Cells
- The difference between experiments was due to different solutions, camera mounting, etc.
- Cells in a culture were not all the same (e.g., cells dying, metabolic changes, diameter, etc.).
- The measured cell was different from the ideal theoretical model. For example, too many cells at the time of measurement, too many pearl chains on the electrodes which distorts the electric field in a point, accidental and occasional thermal or mechanical movements, error in the recording of manual data collection, etc.
4.4. Coupled Yeast Cells
5. Discussion
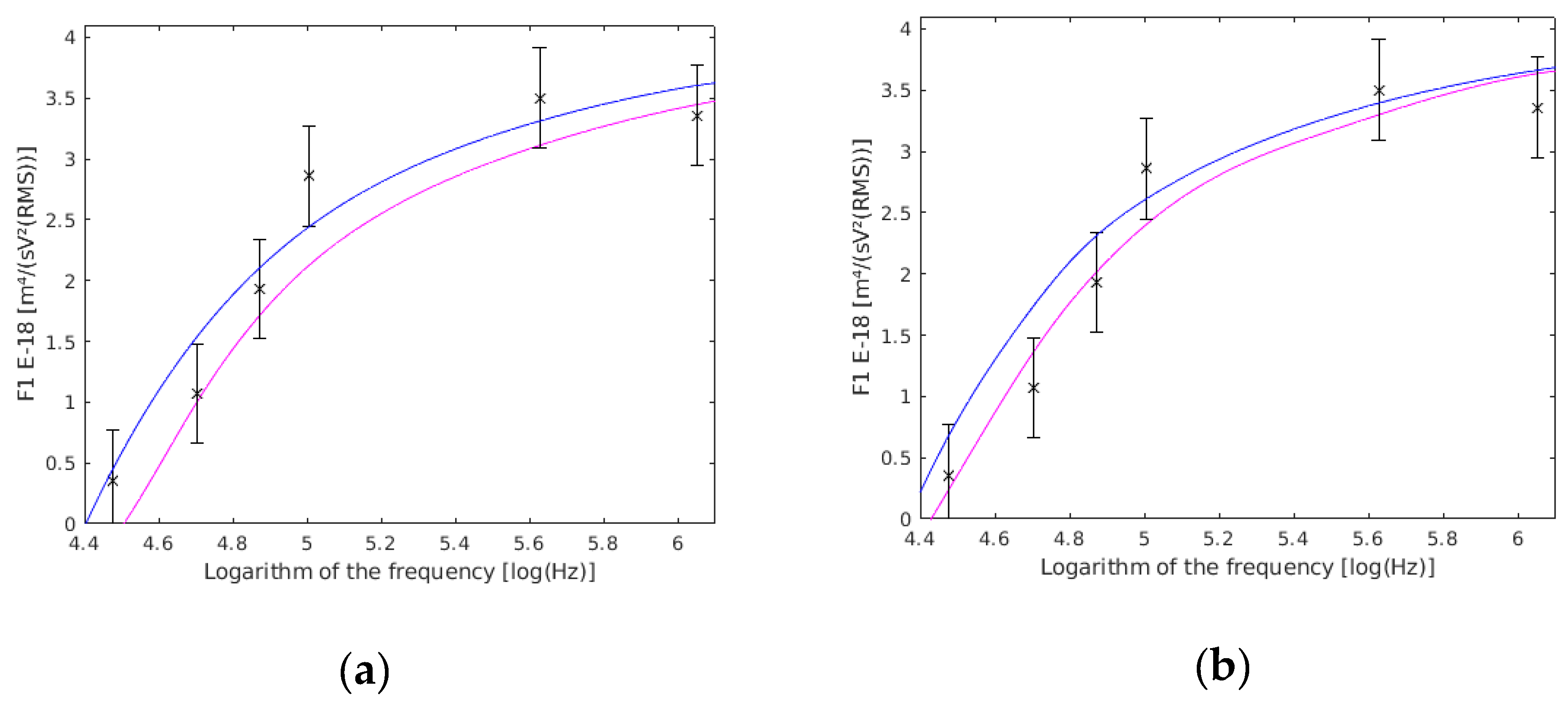
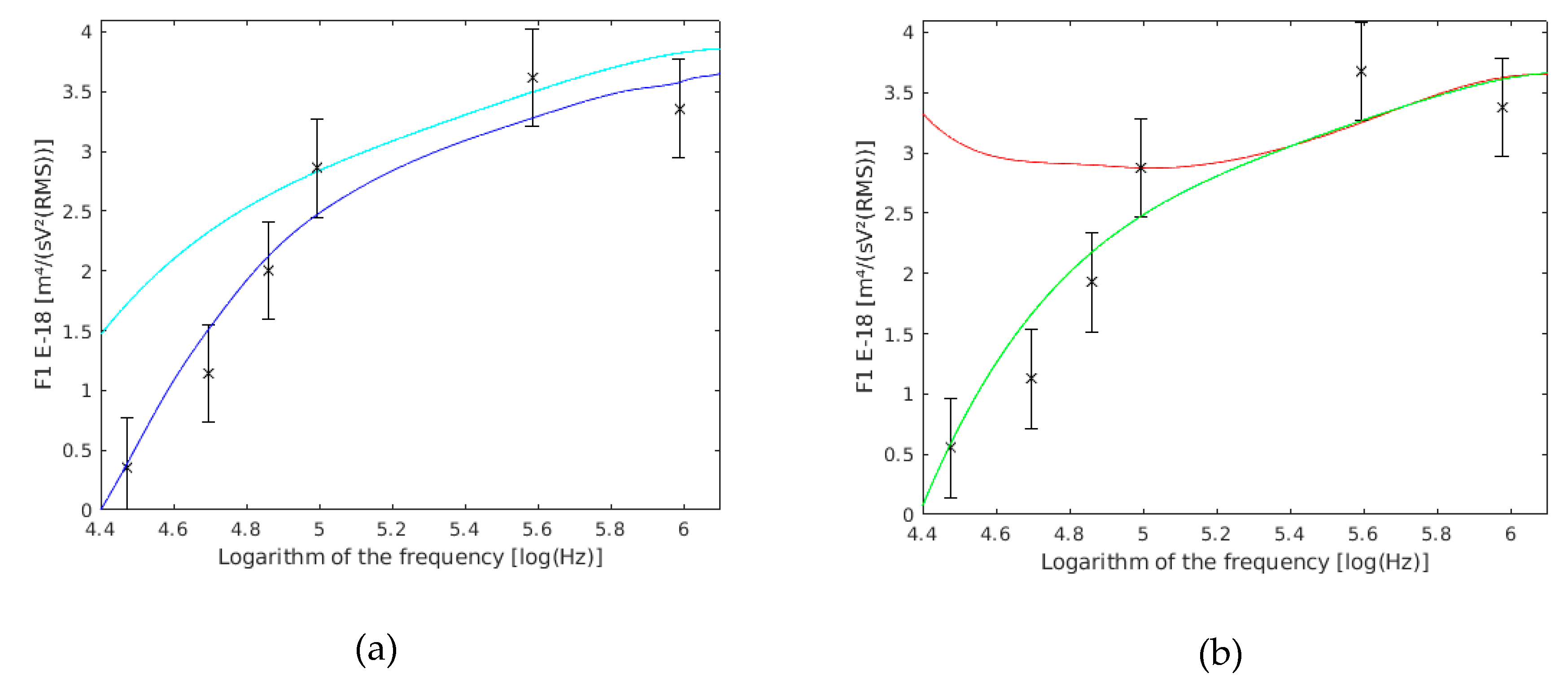
5.1. Single Yeast Cells
5.1.1. Cell Radius
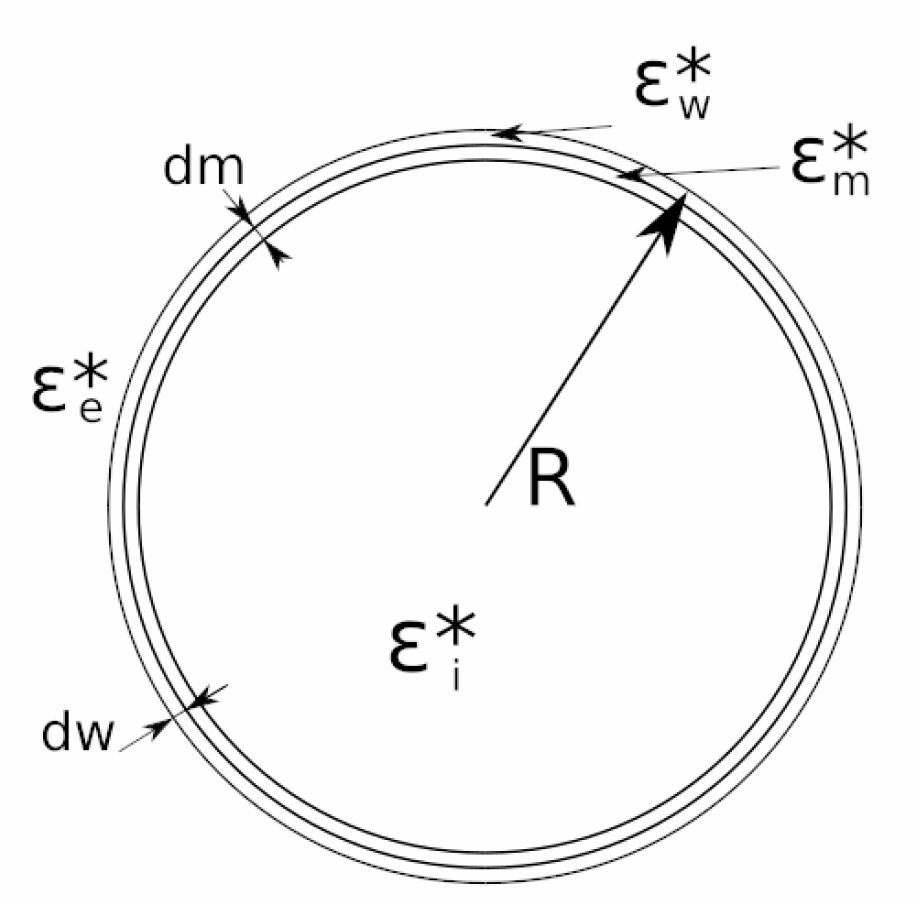
5.1.2. Cell Membrane and Wall Thickness
5.1.3. Cell Wall Conductivity and Permittivity
5.1.4. Cell Membrane Relative Permittivity and Conductivity
5.1.5. Cell Cytoplasm Conductivity and Relative Permittivity
5.2. Coupled Yeast Cells
6. Conclusions
- Conductivity: it would be very useful to vary the solution conductivity so as to look for the membrane capacity. Nonetheless, it is not possible, because of the appearance of thermal currents by the time the conductivity is increased.
- Frequency: to obtain a more complete spectrum, the frequency was varied, and the following problems were observed:
- ∘
- The higher the frequency, the lower the velocity—an effect which is not explained by electrode polarization or by dielectric properties of particles.
- ∘
- The frequency was reduced to reach negative dielectrophoresis, when thermal currents appeared, making any negative dielectrophoresis visualization impossible.
Author Contributions
Conflicts of Interest
References
- Pethig, R. Review—Where Is Dielectrophoresis (DEP) Going? J. Electrochem. Soc. 2017, 164, 3049–3055. [Google Scholar] [CrossRef]
- Su, Y.H.; Warren, C.A.; Guerrant, R.L.; Swami, N.S. Dielectrophoretic Monitoring and Interstrain Separation of Intact Clostridium difficile Based on Their S(Surface)-Layers. Anal. Chem. 2014, 86, 10855–10863. [Google Scholar] [CrossRef]
- Gascoyne, P.R.C.; Shim, S. Isolation of Circulating Tumor Cells by Dielectrophoresis. Cancers 2014, 6, 545–579. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Liu, H.; Bander, N.H.; Kirby, B.J. Enrichment of Prostate Cancer Cells from Blood Cells with a Hybrid Dielectrophoresis and Immunocapture Microfluidic System. Biomed. Microdevices 2013, 15, 941–948. [Google Scholar] [CrossRef]
- Ding, J.; Lawrence, R.M.; Jones, P.V.; Hogue, B.G.; Hayes, M.A. Concentration of Sindbis Virus with Optimized Gradient Insulator-Based Dielectrophoresis. Analyst 2016, 141, 1997–2008. [Google Scholar] [CrossRef]
- Fatoyinbo, H.O.; McDonnell, M.C.; Hughes, M.P. Dielectrophoretic Sample Preparation for Environmental Monitoring of Microorganisms: Soil Particle Removal. Biomicrofluidics 2014, 8, 44115. [Google Scholar] [CrossRef]
- Kumemura, M.; Collard, D.; Sakaki, N.; Yamahata, C.; Hosogi, M.; Hashiguchi, G.; Fujita, H. Single-DNA-Molecule Trapping with Silicon Nanotweezers Using Pulsed Dielectrophoresis. J. Micromech. Microeng. 2011, 21, 54020. [Google Scholar] [CrossRef]
- Flanagan, L.A.; Lu, J.; Wang, L.; Marchenko, S.A.; Jeon, N.L.; Lee, A.P.; Monuki, E.S. Unique Dielectric Properties Distinguish Stem Cells and Their Differentiated Progeny. Stem Cells 2007, 26, 656–665. [Google Scholar] [CrossRef]
- Nakano, M.; Ding, Z.; Suehiro, J. Dielectrophoresis and Dielectrophoretic Impedance Detection of Adenovirus and Rotavirus. Jpn. J. Appl. Phys. 2016, 55, 17001. [Google Scholar] [CrossRef]
- Sonnenberg, A.; Marciniak, J.Y.; McCanna, J.; Krishnan, R.; Rassenti, L.; Kipps, T.J.; Heller, M.J. Dielectrophoretic Isolation and Detection of cfc-DNA Nanoparticulate Biomarkers and Virus from Blood. Electrophoresis 2013, 34, 1076–1084. [Google Scholar] [CrossRef]
- Nakano, M.; Obara, R.; Ding, Z.; Suehiro, J. Detection of Norovirus and Rotavirus by Dielectrophoretic Impedance Measurement. In Proceedings of the Seventh International Conference on Sensing Technology (ICST), Wellington, New Zealand, 3–5 December 2013; pp. 374–378. [Google Scholar]
- Sonnenberg, A.; Marciniak, J.Y.; Skowronski, E.A.; Manouchehri, S.; Rassenti, L.; Ghia, E.M.; Widhopf, G.F.; Kipps, T.J.; Heller, M.J. Dielectrophoretic Isolation and Detection of Cancer-Related Circulating Cell-Free DNA Biomarkers from Blood and Plasma. Electrophoresis 2014, 35, 1828–1836. [Google Scholar] [CrossRef]
- Ramón-Azcón, J.; Yasukawa, T.; Mizutani, F. Sensitive and Spatially Multiplexed Detection System Based on Dielectrophoretic Manipulation of DNA-Encoded Particles Used as Immunoreactions Platform. Anal. Chem. 2011, 83, 1053–1060. [Google Scholar] [CrossRef]
- Abd Rahman, N.; Ibrahim, F.; Yafouz, B. Dielectrophoresis for Biomedical Sciences Applications: A Review. Sensors 2017, 17, 27. [Google Scholar] [CrossRef]
- Jones, P.V.; DeMichele, A.F.; Kemp, L.; Hayes, M.A. Differentiation of Escherichia Coli Serotypes Using DC Gradient Insulator Dielectrophoresis. Anal. Bioanal. Chem. 2014, 406, 183–192. [Google Scholar] [CrossRef]
- Cruz, J.M.; Garcia-Diego, F.J. Dielectrophoretic Force Measurements in Yeast Cells by the Stokes Method. In Proceedings of the 1997 IEEE Industry Applications Conference Thirty-Second IAS Annual Meeting, New Orleans, LA, USA, 5–9 October 1997; pp. 2012–2018. [Google Scholar] [CrossRef]
- Cruz, J.M.; García-Diego, F.J. Dielectrophoretic Motion of Oblate Spheroidal Particles. Measurements of Motion of Red Blood Cells Using the Stokes Method. J. Phys. D Appl. Phys. 1998, 31, 1745. [Google Scholar] [CrossRef]
- Park, S.; Zhang, Y.; Wang, T.H.; Yang, S. Continuous Dielectrophoretic Bacterial Separation and Concentration from Physiological Media of High Conductivity. Lab. Chip 2011, 11, 2893–2900. [Google Scholar] [CrossRef]
- Cherukulappurath, S.; Lee, S.H.; Campos, A.; Haynes, C.L.; Oh, S.H. Rapid and Sensitive in Situ SERS Detection Using Dielectrophoresis. Chem. Mater. 2014, 26, 2445–2452. [Google Scholar] [CrossRef]
- Henslee, E.A.; Sano, M.B.; Rojas, A.D.; Schmelz, E.M.; Davalos, R.V. Selective Concentration of Human Cancer Cells Using Contactless Dielectrophoresis. Electrophoresis 2011, 32, 2523–2529. [Google Scholar] [CrossRef]
- Morimoto, A.; Mogami, T.; Watanabe, M.; Iijima, K.; Akiyama, Y.; Katayama, K.; Futami, T.; Yamamoto, N.; Sawada, T.; Koizumi, F.; et al. High-Density Dielectrophoretic Microwell Array for Detection, Capture, and Single-Cell Analysis of Rare Tumor Cells in Peripheral Blood. PLoS One 2015, 10. [Google Scholar] [CrossRef]
- Huh, D.; Gu, W.; Kamotani, Y.; Grotberg, J.B.; Takayama, S. Microfluidics for Flow Cytometric Analysis of Cells and Particles. Physiol. Meas. 2005, 26. [Google Scholar] [CrossRef]
- Yafouz, B.; Kadri, N.A.; Rothan, H.A.; Yusof, R.; Ibrahim, F. Discriminating Dengue-Infected Hepatic Cells (WRL-68) Using Dielectrophoresis. Electrophoresis 2016, 37, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Alshareef, M.; Metrakos, N.; Juarez Perez, E.; Azer, F.; Yang, F.; Yang, X.; Wang, G. Separation of Tumor Cells with Dielectrophoresis-Based Microfluidic Chip. Biomicrofluidics 2013, 7, 11803. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Graham, K.A.; Johannessen, A.C.; Costea, D.E.; Labeed, F.H. Human Oral Cancer Cells with Increasing Tumorigenic Abilities Exhibit Higher Effective Membrane Capacitance. Integr. Biol. 2014, 6, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.; Hughes, M.P.; Mulhall, H.J.; Oreffo, R.O.C.; Labeed, F.H. Characterization of Human Skeletal Stem and Bone Cell Populations Using Dielectrophoresis. J. Tissue Eng. Regen. Med. 2013, 9, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Manczak, R.; Saada, S.; Provent., T.; Dalmay, C.; Bessette, B.; Bégaud, G.S.; Battu, S.; Blondy, P.; Jauberteau, M.O.; Kaynak, C.B.; et al. UHF-Dielectrophoresis Crossover Frequency as a New Marker for Discrimination of Glioblastoma Undifferentiated Cells. IEEE J. Electromagnet. RF Microwaves Med. Biol. 2019, 3, 191–198. [Google Scholar] [CrossRef]
- Yun, H.; Kim, K.; Lee, W.G. Cell manipulation in Microfluidics. Biofabrication 2013, 5, 22001. [Google Scholar] [CrossRef] [PubMed]
- Shafiee, H.; Jahangir, M.; Inci, F.; Wang, S.; Willenbrecht, R.B.; Giguel, F.F.; Tsibris, A.M.; Kuritzkes, D.R.; Demirci, U. Acute On-Chip HIV Detection Through Label-Free Electrical Sensing of Viral Nano-Lysate. Small 2013, 9, 2553–2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgarella, S.; Di Bari, M. A Portable and Integrated Instrument for Cell Manipulation by Dielectrophoresis. Electrophoresis 2015, 36, 1466–1470. [Google Scholar] [CrossRef]
- Burgarella, S.; Merlo, S.; Dell’Anna, B.; Zarola, G.; Bianchessi, M. A Modular Micro-Fluidic Platform for Cells Handling by Dielectrophoresis. Microelectron. Eng. 2010, 87, 2124–2133. [Google Scholar] [CrossRef]
- Hyun, C.; Kaur, H.; McNabb, D.S.; Li, J. Dielectrophoretic Stretching of DNA Tethered to a Fiber Tip. Nanotechnology 2015, 26, 125501. [Google Scholar] [CrossRef]
- Manouchehri, S.; Ibsen, S.; Wright, J.; Rassenti, L.; Ghia, E.M.; Widhopf, G.F.; Kipps, T.J.; Heller, M.J. Dielectrophoretic Recovery of DNA from Plasma for the Identification of Chronic Lymphocytic Leukemia Point Mutations. Int. J. Hematol. Oncol. 2016, 1, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Heller, M.J.; Krishnan, R.; Sonnenberg, A. Rapid Detection of Cancer Related DNA Nanoparticulate Biomarkers and Nanoparticles in Whole Blood. Int. Soc. Opt. Photonics 2010, 7759. [Google Scholar] [CrossRef]
- Schröder, U.C.; Ramoji, A.; Glaser, U.; Sachse, S.; Leiterer, C.; Csaki, A.; Huübner, U.; Fritzsche, W.; Pfister, W.; Bauer, M.; et al. Combined Dielectrophoresis–Raman Setup for the Classification of Pathogens Recovered from the Urinary Tract. Anal. Chem. 2013, 85, 10717–10724. [Google Scholar] [CrossRef]
- Du, E.; Dao, M.; Suresh, S. Quantitative Biomechanics of Healthy and Diseased Human Red Blood Cells Using Dielectrophoresis in a Microfluidic System. Extrem. Mech. Lett. 2014, 1, 35–41. [Google Scholar] [CrossRef]
- Bakewell, D.J.; Bailey, J.; Holmes, D. Real-Time Dielectrophoretic Signaling and Image Quantification Methods for Evaluating Electrokinetic Properties of Nanoparticles. Electrophoresis 2015, 36, 1443–1450. [Google Scholar] [CrossRef] [PubMed]
- Afshar, S.; Fazelkhah, A.; Salimi, E.; Butler, M.; Thomson, D.J.; Bridges, G.E. Dielectric Properties of Single Cells Subjected to Heat Shock Using DEP Cytometry. IEEE Trans. Microwave Theory Tech. 2018, 12, 5933–5940. [Google Scholar] [CrossRef]
- Pohl, H.A. Dielectrophoresis: the behavior of neutral matter in nonuniform electric fields; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1978. [Google Scholar]
- Akshay, K.; Saurin, P.; Shizhi, Q.; Guoqing, H.; Xiangchun, X. Joule heating effects on reservoir-based dielectrophoresis. Electrophoresis 2013, 35, 721–727. [Google Scholar] [CrossRef] [Green Version]
- Asuka, N.; Alexandra, R. Protein Dielectrophoresis: Advances, Challenges, and Applications. Electrophoresis 2013, 34, 1085–1096. [Google Scholar] [CrossRef]
- Pethig, R.; Huang, Y.; Wang, X.; Burt, J.P.H. Positive and Negative Dielectrophoretic Collection of Colloidal Particles Using Interdigitated Castellated Microelectrodes. J. Phys. D Appl. Phys. 1992, 25, 881. [Google Scholar] [CrossRef]
- Kaler, K.V.; Jones, T.B. Dielectrophoretic Spectra of Single Cells Determined by Feedback-Controlled Levitation. Biophys. J. 1990, 57, 173–182. [Google Scholar] [CrossRef]
- Becker, F.F.; Wang, X.B.; Huang, Y.; Pethig, R.; Vykoukal, J.; Gascoyne, P.R. Separation of Human Breast Cancer Cells From Blood by Differential Dielectric Affinity. Proc. Natl. Acad. Sci. USA 1995, 92, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, X.B.; Tame, J.A.; Pethig, R. Electrokinetic Behaviour of Colloidal Particles in Travelling Electric Fields: Studies Using Yeast Cells. J. Phys. D Appl. Phys. 1993, 26, 1528. [Google Scholar] [CrossRef]
- Gascoyne, P.R.C.; Huang, Y.; Pethig, R.; Vykoukal, J.; Becker, F.F. Dielectrophoretic Separation of Mammalian Cells Studied by Computerized Image Analysis. Meas. Sci. Technol. 1992, 3, 439. [Google Scholar] [CrossRef]
- Wang, X.B.; Huang, Y.; Burt, J.P.H.; Markx, G.H.; Pethig, R. Selective Dielectrophoretic Confinement of Bioparticles in Potential Energy Wells. J. Phys. D Appl. Phys. 1993, 26, 1278. [Google Scholar] [CrossRef]
- Reynolds, O. An Experimental Investigation of the Circumstances Which Determine Whether the Motion of Water Shall Be Direct or Sinuous, and of the Law of Resistance in Parallel Channels. Philos. Trans. R. Soc. Lond. 1883, 174, 935–982. [Google Scholar]
- Reddy, S.; Moore, L.R.; Sun, L.; Zborowski, M.; Chalmers, J.J. Determination of the Magnetic Susceptibility of Labeled Particles by Video Imaging. Chem. Eng. Sci. 1996, 51, 947–956. [Google Scholar] [CrossRef]
- Happel, J.; Brenner, H. Low Reynolds number Hydrodynamcis with special applications to particulate media; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1983; pp. 1–553. [Google Scholar]
- Schwan, H.P. Linear and Nonlinear Electrode Polarization and Biological Materials. Ann. Biomed. Eng. 1992, 20, 269–288. [Google Scholar] [CrossRef]
- Jones, T.B. Electromechanics of particles; Cambridge University Press: Cambridge, UK; New York, NY, USA.
- Pierce, J.S. Institute of Brewing: Analysis Committee Measurement of Yeast Viability. J. Inst. Brew. 1970, 76, 442–443. [Google Scholar] [CrossRef]
- Miller, R.D.; Jones, T.B. On the Effective Dielectric Constant of Columns or Layers of Dielectric Spheres. J. Phys. D Appl. Phys. 1988, 21, 527. [Google Scholar] [CrossRef]
- Sancho, M.; Martínez, G.; Muñoz, S.; Sebastián, J.L.; Pethig, R. Interaction between Cells in Dielectrophoresis and Electrorotation Experiments. Biomicrofluidics 2010, 4, 22802. [Google Scholar] [CrossRef]
- Asami, K.; Yonezawa, T. Dielectric Behavior of Wild-Type Yeast and Vacuole-Deficient Mutant Over a Frequency Range of 10 kHz to 10 GHz. Biophys. J. 1996, 71, 2192–2200. [Google Scholar] [CrossRef]
- Hölzel, R.; Lamprecht, I. Dielectric Properties of Yeast Cells as Determined by Electrorotation. Biochim. Biophys. Acta 1992, 1104, 195–200. [Google Scholar] [CrossRef]
- Chan, K.L.; Gascoyne, P.R.; Becker, F.F.; Pethig, R. Electrorotation of Liposomes: Verification of Dielectric Multi-Shell Model for Cells. Biochim. Biophys. Acta 1997, 1349, 182–196. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Single cells | ||
| Coupled cells |
| Parallel Resistance (Ω) | 18,500 |
| Parallel Capacitance (F) |
| Frequency (kHz) | Standard Error | Selected Cells | Measured Cells | |
|---|---|---|---|---|
| 30 | 0.35 | 0.4 | 8 | 42 |
| 50 | 1.17 | 0.1 | 36 | 61 |
| 75 | 1.88 | 0.1 | 52 | 72 |
| 100 | 2.75 | 0.1 | 48 | 69 |
| 400 | 3.57 | 0.1 | 57 | 95 |
| 1000 | 3.24 | 0.1 | 72 | 103 |
| Frequency (kHz) | Standard Error | Selected Cells | Measured Cells | |
|---|---|---|---|---|
| 30 | 0.63 | 0.2 | 32 | 49 |
| 50 | 1.55 | 0.2 | 47 | 58 |
| 75 | 2.83 | 0.1 | 56 | 67 |
| 100 | 3.39 | 0.1 | 56 | 64 |
| 400 | 4.60 | 0.1 | 58 | 85 |
| 1000 | 5.59 | 0.1 | 67 | 80 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Diego, F.-J.; Rubio-Chavarría, M.; Beltrán, P.; Espinós, F.J. Characterization of Simple and Double Yeast Cells Using Dielectrophoretic Force Measurement. Sensors 2019, 19, 3813. https://doi.org/10.3390/s19173813
García-Diego F-J, Rubio-Chavarría M, Beltrán P, Espinós FJ. Characterization of Simple and Double Yeast Cells Using Dielectrophoretic Force Measurement. Sensors. 2019; 19(17):3813. https://doi.org/10.3390/s19173813
Chicago/Turabian StyleGarcía-Diego, Fernando-Juan, Mario Rubio-Chavarría, Pedro Beltrán, and Francisco J. Espinós. 2019. "Characterization of Simple and Double Yeast Cells Using Dielectrophoretic Force Measurement" Sensors 19, no. 17: 3813. https://doi.org/10.3390/s19173813
APA StyleGarcía-Diego, F. -J., Rubio-Chavarría, M., Beltrán, P., & Espinós, F. J. (2019). Characterization of Simple and Double Yeast Cells Using Dielectrophoretic Force Measurement. Sensors, 19(17), 3813. https://doi.org/10.3390/s19173813