Climate-Triggered Insect Defoliators and Forest Fires Using Multitemporal Landsat and TerraClimate Data in NE Iran: An Application of GEOBIA TreeNet and Panel Data Analysis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data and Field Mensuration
2.3. Methodology
2.3.1. TreeNet-Based Insect Infestation Mapping
2.3.2. Intensity of Insect Infestation, Severity of Forest Fire, and Climate Hazards
2.3.3. Relationships among Insect Infestation, Forest Fires, and Climate Hazards
Panel data models
Testing for fixed effects and random effects
3. Results
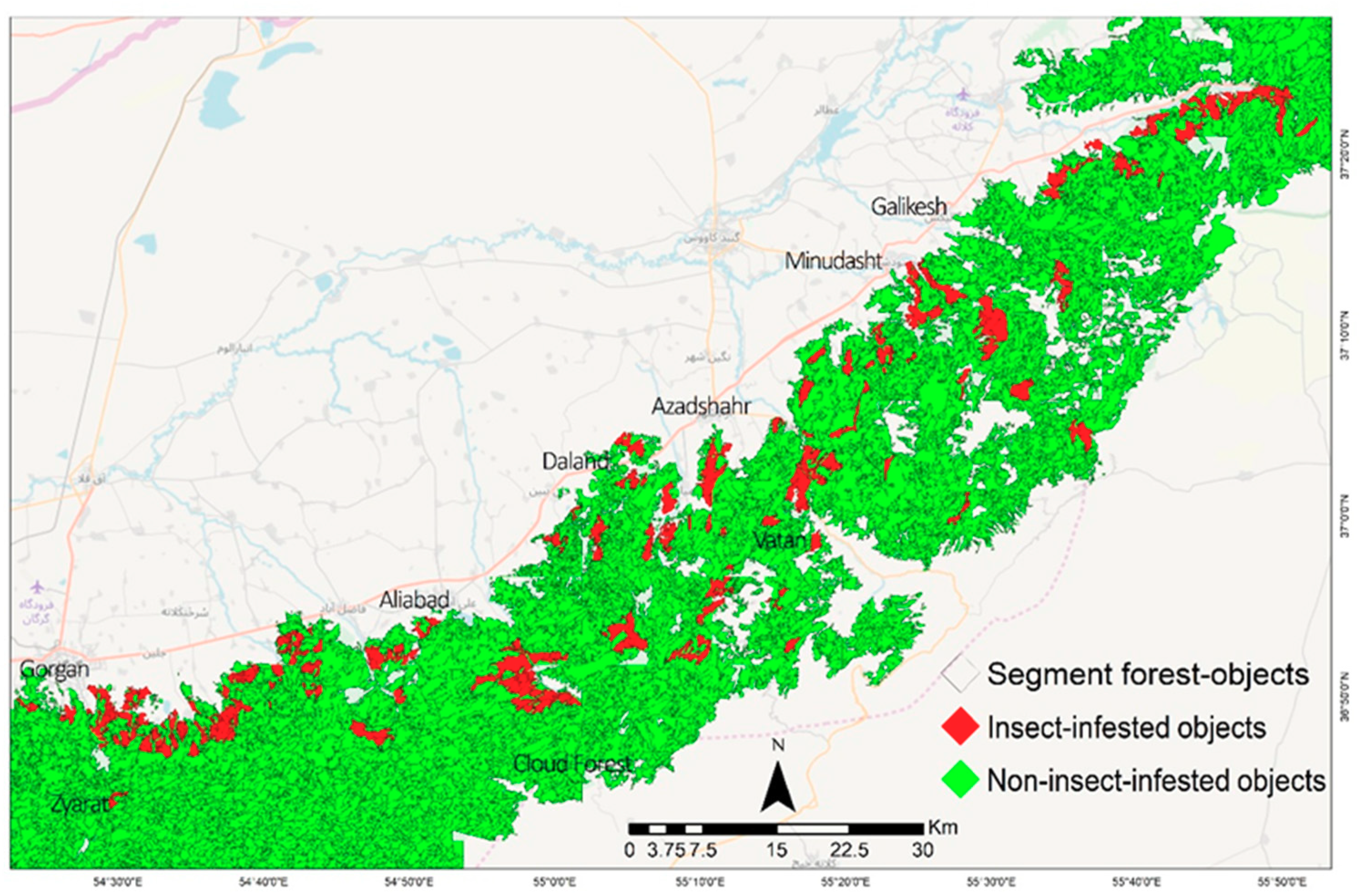
3.1. Insect Defoliation Mapping
3.2. Insect Infestation, Forest Fires, and Climate Hazards Modelling
4. Discussion
5. Conclusions
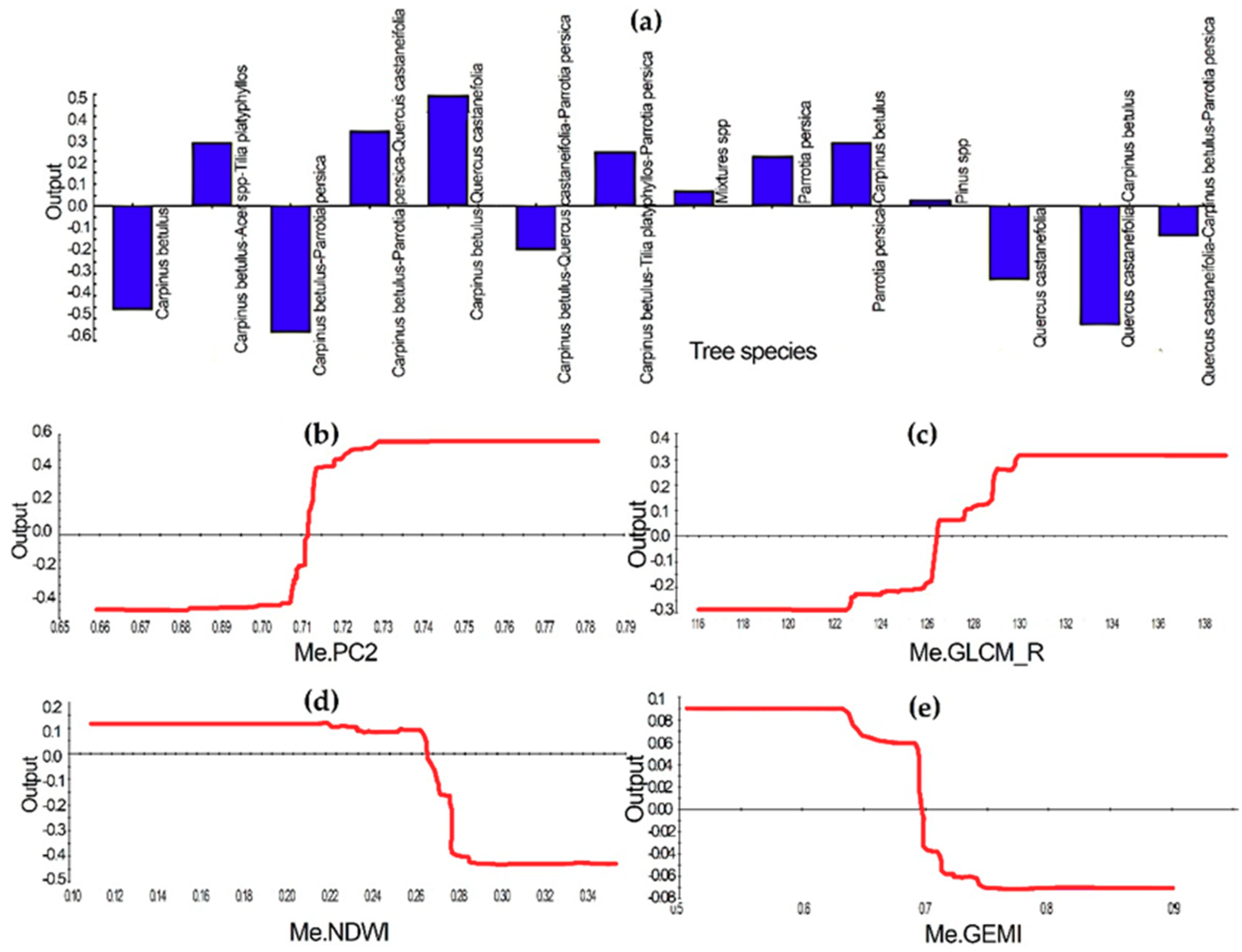
- GEOBIA TreeNet indicated excellent performance with the contribution of Landsat 8 OLI-derived and ancillary object features for discriminating insect-defoliated forests from healthy forests.
- Although the object features of Landsat 8 OLI recorded a higher importance for discriminating insect-defoliated objects, tree species has obtained the second rank of importance following the mean of PC2. In addition, other top image object features were the mean of the red channels derived from GLCM, the mean of NDWI, and the mean of GEMI, respectively.
- The random effects model demonstrated higher performance in comparison with the fixed effects and common effects models to model mutual interaction of the intensity of insect defoliation and the severity of forest fire and their associations with the TerraClimate-derived climate hazards.
- Maximum temperatures significantly triggered both insect outbreaks and forest fires. Although the drought conditions of the current year and the availability of soil moisture of the previous year were significant regarding the intensity of insect infestation, they have indicated neutral effects on the severity of forest fires.
- The severity of forest fires of the previous year has triggered the intensity of insect infestation; however, the insect infestation was not effective for the forest fires.
- Future studies will be required to explore the application of novel satellite images such as Sentinel-2 or the combination of Landsat 8 and Sentinel-2 for monitoring near-real-time insect-induced defoliation, identifying infestations resulting from bark beetles and pathogens, and discriminating between them.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Swetnam, T.W.; Lynch, A.M. Multicentury, Regional-Scale Patterns of Western Spruce Budworm Outbreaks. Ecol. Monogr. 1993, 63, 399–424. [Google Scholar] [CrossRef]
- Rullan-Silva, C.D.; Olthoff, A.E.; La Delgado de Mata, J.A.; Pajares-Alonso, J.A. Remote Monitoring of Forest Insect Defoliation. A Review. For. Syst. 2013, 22, 377–391. [Google Scholar] [CrossRef]
- Hall, R.J.; Castilla, G.; White, J.C.; Cooke, B.J.; Skakun, R.S. Remote sensing of forest pest damage: A review and lessons learned from a Canadian perspective. Can. Entomol. 2016, 148, S296–S356. [Google Scholar] [CrossRef]
- Dash, J.P.; Watt, M.S.; Pearse, G.D.; Heaphy, M.; Dungey, H.S. Assessing very high resolution UAV imagery for monitoring forest health during a simulated disease outbreak. ISPRS J. Photogramm. Remote Sens. 2017, 131, 1–14. [Google Scholar] [CrossRef]
- Senf, C.; Seidl, R.; Hostert, P. Remote sensing of forest insect disturbances: Current state and future directions. Int. J. Appl. Earth Obs. Geoinf. 2017, 60, 49–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, J.H. Stochastic gradient boosting. Comput. Stat. Data Anal. 2002, 38, 367–378. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Dobrowski, S.Z.; Parks, S.A.; Hegewisch, K.C. TerraClimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958–2015. Sci. Data 2018, 5, 170191. [Google Scholar] [CrossRef] [PubMed]
- Latifi, H.; Fassnacht, F.E.; Schumann, B.; Dech, S. Object-based extraction of bark beetle (Ips typographus L.) infestations using multi-date LANDSAT and SPOT satellite imagery. Prog. Phys. Geogr. Earth Environ. 2014, 38, 755–785. [Google Scholar] [CrossRef]
- Meddens, A.J.H.; Hicke, J.A. Spatial and temporal patterns of Landsat-based detection of tree mortality caused by a mountain pine beetle outbreak in Colorado, USA. For. Ecol. Manag. 2014, 322, 78–88. [Google Scholar] [CrossRef]
- De Beurs, K.M.; Townsend, P.A. Estimating the effect of gypsy moth defoliation using MODIS. Remote Sens. Environ. 2008, 112, 3983–3990. [Google Scholar] [CrossRef]
- Senf, C.; Wulder, M.A.; Campbell, E.M.; Hostert, P. Using Landsat to Assess the Relationship between Spatiotemporal Patterns of Western Spruce Budworm Outbreaks and Regional-Scale Weather Variability. Can. J. Remote Sens. 2016, 42, 706–718. [Google Scholar] [CrossRef]
- Babst, F.; Esper, J.; Parlow, E. Landsat TM/ETM+ and tree-ring based assessment of spatiotemporal patterns of the autumnal moth (Epirrita autumnata) in northernmost Fennoscandia. Remote Sens. Environ. 2010, 114, 637–646. [Google Scholar] [CrossRef]
- Townsend, P.A.; Singh, A.; Foster, J.R.; Rehberg, N.J.; Kingdon, C.C.; Eshleman, K.N.; Seagle, S.W. A general Landsat model to predict canopy defoliation in broadleaf deciduous forests. Remote Sens. Environ. 2012, 119, 255–265. [Google Scholar] [CrossRef]
- Rullán-Silva, C.; Olthoff, A.E.; Pando, V.; Pajares, J.A.; Delgado, J.A. Remote monitoring of defoliation by the beech leaf-mining weevil Rhynchaenus fagi in northern Spain. For. Ecol. Manag. 2015, 347, 200–208. [Google Scholar] [CrossRef]
- Meddens, A.J.H.; Hicke, J.A.; Vierling, L.A.; Hudak, A.T. Evaluating methods to detect bark beetle-caused tree mortality using single-date and multi-date Landsat imagery. Remote Sens. Environ. 2013, 132, 49–58. [Google Scholar] [CrossRef]
- Vastaranta, M.; Kantola, T.; Lyytikäinen-Saarenmaa, P.; Holopainen, M.; Kankare, V.; Wulder, M.; Hyyppä, J.; Hyyppä, H. Area-Based Mapping of Defoliation of Scots Pine Stands Using Airborne Scanning LiDAR. Remote Sens. 2013, 5, 1220–1234. [Google Scholar] [CrossRef] [Green Version]
- Adelabu, S.; Mutanga, O.; Adam, E. Evaluating the impact of red-edge band from Rapideye image for classifying insect defoliation levels. ISPRS J. Photogramm. Remote Sens. 2014, 95, 34–41. [Google Scholar] [CrossRef]
- Goodwin, N.R.; Coops, N.C.; Wulder, M.A.; Gillanders, S.; Schroeder, T.A.; Nelson, T. Estimation of insect infestation dynamics using a temporal sequence of Landsat data. Remote Sens. Environ. 2008, 112, 3680–3689. [Google Scholar] [CrossRef]
- Paritsis, J.; Veblen, T.T.; Smith, J.M.; Holz, A. Spatial prediction of caterpillar (Ormiscodes) defoliation in Patagonian Nothofagus forests. Landsc. Ecol. 2011, 26, 791–803. [Google Scholar] [CrossRef]
- Thayn, J.B. Using a remotely sensed optimized Disturbance Index to detect insect defoliation in the Apostle Islands, Wisconsin, USA. Remote Sens. Environ. 2013, 136, 210–217. [Google Scholar] [CrossRef]
- Heurich, M.; Ochs, T.; Andresen, T.; Schneider, T. Object-orientated image analysis for the semi-automatic detection of dead trees following a spruce bark beetle (Ips typographus) outbreak. Eur. J. For. Res. 2010, 129, 313–324. [Google Scholar] [CrossRef]
- Hay, G.J.; Castilla, G. Geographic Object-Based Image Analysis (GEOBIA): A new name for a new discipline. In Object-Based Image Analysis: Spatial Concepts for Knowledge-Driven Remote Sensing Applications; Blaschke, T., Lang, S., Hay, G.J., Eds.; Springer: Berlin, Germany; London, UK, 2008; pp. 75–89. ISBN 978-3-540-77057-2. [Google Scholar]
- Latifi, H.; Schumann, B.; Kautz, M.; Dech, S. Spatial characterization of bark beetle infestations by a multidate synergy of SPOT and Landsat imagery. Environ. Monit. Assess. 2014, 186, 441–456. [Google Scholar] [CrossRef] [PubMed]
- Flower, A.; Gavin, D.G.; Heyerdahl, E.K.; Parsons, R.A.; Cohn, G.M. Drought-triggered western spruce budworm outbreaks in the interior Pacific Northwest: A multi-century dendrochronological record. For. Ecol. Manag. 2014, 324, 16–27. [Google Scholar] [CrossRef]
- Millar, C.I.; Stephenson, N.L. Temperate forest health in an era of emerging megadisturbance. Science 2015, 349, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Kautz, M.; Meddens, A.J.H.; Hall, R.J.; Arneth, A. Biotic disturbances in Northern Hemisphere forests—A synthesis of recent data, uncertainties and implications for forest monitoring and modelling. Glob. Ecol. Biogeogr. 2017, 26, 533–552. [Google Scholar] [CrossRef]
- Ward, S.F.; Aukema, B.H. Anomalous outbreaks of an invasive defoliator and native bark beetle facilitated by warm temperatures, changes in precipitation and interspecific interactions. Ecography 2019, 42, 1068–1078. [Google Scholar] [CrossRef] [Green Version]
- Ryan, M.G.; Vose, J.M. Effects of Climatic Variability and Change on Forest Ecosystems: A Comprehensive Science Synthesis for the U.S. Forest Sector. PNW-GTR-870; 2012. Available online: https://www.fs.usda.gov/treesearch/pubs/42651 (accessed on 2 August 2019).
- Paradis, A.; Elkinton, J.; Hayhoe, K.; Buonaccorsi, J. Role of winter temperature and climate change on the survival and future range expansion of the hemlock woolly adelgid (Adelges tsugae) in eastern North America. Mitig. Adapt. Strateg. Glob. Chang. 2008, 13, 541–554. [Google Scholar] [CrossRef]
- Trần, J.K.; Ylioja, T.; Billings, R.F.; Régnière, J.; Ayres, M.P. Impact of minimum winter temperatures on the population dynamics of Dendroctonus frontalis. Ecol. Appl. 2007, 17, 882–899. [Google Scholar] [CrossRef]
- Pureswaran, D.S.; Roques, A.; Battisti, A. Forest Insects and Climate Change. Curr. For. Rep. 2018, 4, 35–50. [Google Scholar] [CrossRef] [Green Version]
- Jepsen, J.U.; Kapari, L.; Hagen, S.B.; Schott, T.; Vindstad, O.P.L.; Nilssen, A.C.; Ims, R.A. Rapid northwards expansion of a forest insect pest attributed to spring phenology matching with sub-Arctic birch. Glob. Chang. Biol. 2011, 17, 2071–2083. [Google Scholar] [CrossRef]
- Kolb, T.E.; Fettig, C.J.; Ayres, M.P.; Bentz, B.J.; Hicke, J.A.; Mathiasen, R.; Stewart, J.E.; Weed, A.S. Observed and anticipated impacts of drought on forest insects and diseases in the United States. For. Ecol. Manag. 2016, 380, 321–334. [Google Scholar] [CrossRef]
- Fettig, C.J.; Mortenson, L.A.; Bulaon, B.M.; Foulk, P.B. Tree mortality following drought in the central and southern Sierra Nevada, California, U.S. For. Ecol. Manag. 2019, 432, 164–178. [Google Scholar] [CrossRef]
- Littell, J.S.; Peterson, D.L.; Riley, K.L.; Liu, Y.; Luce, C.H. Fire and Drought. In Effects of Drought on Forests and Rangelands in the United States: A Comprehensive Science Synthesis; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 2016; pp. 135–154. [Google Scholar]
- Rouault, G.; Candau, J.-N.; Lieutier, F.; Nageleisen, L.-M.; Martin, J.-C.; Warzée, N. Effects of drought and heat on forest insect populations in relation to the 2003 drought in Western Europe. Ann. For. Sci. 2006, 63, 613–624. [Google Scholar] [CrossRef]
- Ryerson, D.E.; Swetnam, T.W.; Lynch, A.M. A tree-ring reconstruction of western spruce budworm outbreaks in the San Juan Mountains, Colorado, U.S.A. Can. J. For. Res. 2003, 33, 1010–1028. [Google Scholar] [CrossRef]
- Thomson, A.J.; Shepherd, R.F.; Harris, J.W.E.; Silversides, R.H. Relating weather to outbreaks of western spruce budworm, Choristoneura occidentalis (Lepidoptera: Tortricidae), in British Columbia. Can. Entomol. 1984, 116, 375–381. [Google Scholar] [CrossRef]
- Chen, G.; He, Y.; de Santis, A.; Li, G.; Cobb, R.; Meentemeyer, R.K. Assessing the impact of emerging forest disease on wildfire using Landsat and KOMPSAT-2 data. Remote Sens. Environ. 2017, 195, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Hicke, J.A.; Johnson, M.C.; Hayes, J.L.; Preisler, H.K. Effects of bark beetle-caused tree mortality on wildfire. For. Ecol. Manag. 2012, 271, 81–90. [Google Scholar] [CrossRef]
- Agne, M.C.; Woolley, T.; Fitzgerald, S. Fire severity and cumulative disturbance effects in the post-mountain pine beetle lodgepole pine forests of the Pole Creek Fire. For. Ecol. Manag. 2016, 366, 73–86. [Google Scholar] [CrossRef] [Green Version]
- Meigs, G.W.; Campbell, J.L.; Zald, H.S.J.; Bailey, J.D.; Shaw, D.C.; Kennedy, R.E. Does wildfire likelihood increase following insect outbreaks in conifer forests? Ecosphere 2015, 6, 1–24. [Google Scholar] [CrossRef]
- Kane, J.M.; Varner, J.M.; Metz, M.R.; van Mantgem, P.J. Characterizing interactions between fire and other disturbances and their impacts on tree mortality in western U.S. Forests. For. Ecol. Manag. 2017, 405, 188–199. [Google Scholar] [CrossRef]
- Kulakowski, D.; Jarvis, D. Low-severity fires increase susceptibility of lodgepole pine to mountain pine beetle outbreaks in Colorado. For. Ecol. Manag. 2013, 289, 544–550. [Google Scholar] [CrossRef]
- McCarley, T.R.; Kolden, C.A.; Vaillant, N.M.; Hudak, A.T.; Smith, A.M.S.; Kreitler, J. Landscape-scale quantification of fire-induced change in canopy cover following mountain pine beetle outbreak and timber harvest. For. Ecol. Manag. 2017, 391, 164–175. [Google Scholar] [CrossRef] [Green Version]
- Shirvani, Z.; Abdi, O.; Buchroithner, M.F.; Pradhan, B. Analysing Spatial and Statistical Dependencies of Deforestation Affected by Residential Growth: Gorganrood Basin, Northeast Iran. Land Degrad. Dev. 2017, 28, 2176–2190. [Google Scholar] [CrossRef]
- Abdi, O.; Kamkar, B.; Shirvani, Z.; Teixeira da Silva, J.A.; Buchroithner, M.F. Spatial-statistical analysis of factors determining forest fires: A case study from Golestan, Northeast Iran. Geomat. Nat. Hazards Risk 2018, 9, 267–280. [Google Scholar] [CrossRef]
- Abdi, O.; Shirvani, Z.; Buchroithner, M.F. Spatiotemporal drought evaluation of Hyrcanian deciduous forests and semi-steppe rangelands using moderate resolution imaging spectroradiometer time series in Northeast Iran. Land Degrad. Dev. 2018, 29, 2525–2541. [Google Scholar] [CrossRef]
- Abdi, O.; Shirvani, Z.; Buchroithner, M.F. Forest drought-induced diversity of Hyrcanian individual-tree mortality affected by meteorological and hydrological droughts by analyzing moderate resolution imaging spectroradiometer products and spatial autoregressive models over northeast Iran. Agric. For. Meteorol. 2019, 275, 265–276. [Google Scholar] [CrossRef]
- Setayeshgar, F. The Attack of Lymantria Dispar to 420 Hecatres of Hyrcanian Forests: Outbreak of Defoliators in the Forests. Available online: http://khorasannews.com/?nid=18724&type=0 (accessed on 1 July 2014).
- Kia-Daliri, H.; Kazemi-Najafi, S.; Ahangaran, Y. The effect of leaf feeder moth (Erannis defoliaria & Operophtera brumata) on radial growth of 3 tree species in north of Iran (case study: Mashalak, Nowshahr). Iran. J. For. Poplar Res. 2007, 15, 301–309. [Google Scholar]
- Ahmadi, A.; Kavosi, M.R.; Soltanloo, H. Zelkova carpinifolia reservoir from Hyrcanian Forests, Northern Iran, a new sacrifice of Ophiostoma novo-ulmi. Biodiversitas 2014, 15, 48–52. [Google Scholar] [CrossRef]
- F.A.O. Food Chain Crisis Early Warning Bulletin. Alerts on Threats to the Food Chain Affecting Food Security in Countries and Regions No. 18. 2016. Available online: www.fao.org/food-chain-crisis (accessed on 20 July 2019).
- Mirabolfathy, M.; Groenewald, J.Z.; Crous, P.W. The Occurrence of Charcoal Disease Caused by Biscogniauxia mediterranea on Chestnut-Leaved Oak (Quercus castaneifolia) in the Golestan Forests of Iran. Plant Dis. 2011, 95, 876. [Google Scholar] [CrossRef]
- Sagheb Talebi, K.; Sajedi, T.; Pourhashemi, M. Forests of Iran. A Treasure from the Past, a Hope for the Future; Springer: Dordrecht, The Netherlands, 2014; ISBN 978-94-007-7371-4. [Google Scholar]
- U.S. Geological Survey, Earth Resources Observation Science Center. Collection-1 Landsat OLI Level-2 Surface Reflectance (SR) Science Product; U.S. Geological Survey, Earth Resources Observation Science Center: Reston, VA, USA, 2019.
- ASF DAAC. PALSAR Radiometric Terrain Corrected High Res; Includes Material © JAXA/METI 2007; ASF DAAC: Fairbanks, AK, USA, 2019. [Google Scholar]
- Department of Natural Resources and Watershed Management of Golestan. The Booklets of Forest Management Plans: Jafarabad, Golriz, Tavir, Kohmian, Zarringol, Sorkhdari, Rezaeian, Qalemorian, Daland, Vatan, Nodeh-Bon-Qale, Minudasht, Takht, Loveh, Sijan-Kiaram, Korankaftar, Terjenli; Department of Natural Resources and Watershed Management of Golestan: Gorgan, Iran, 1981–2011. [Google Scholar]
- Google Earth Pro V 7.3.2.5491. Sousara, Golestan Province. 37° 00’ 47.58”N, 55° 16’ 30.88”E, Eye alt 10.17 km; DigitalGlobe 2019; Google LLC: Mountain View, CA, USA, 2019. [Google Scholar]
- eCognition Developer. T. 9.0 User Guide; Trimble Germany GmbH: Munich, Germany, 2014. [Google Scholar]
- Crippen, R. Calculating the vegetation index faster. Remote Sens. Environ. 1990, 34, 71–73. [Google Scholar] [CrossRef]
- Pinty, B.; Verstraete, M.M. GEMI: A non-linear index to monitor global vegetation from satellites. Vegetatio 1992, 101, 15–20. [Google Scholar] [CrossRef]
- Kaufman, Y.J.; Tanre, D. Atmospherically resistant vegetation index (ARVI) for EOS-MODIS. IEEE Trans. Geosci. Remote Sens. 1992, 30, 261–270. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Kaufman, Y.J.; Merzlyak, M.N. Use of a green channel in remote sensing of global vegetation from EOS-MODIS. Remote Sens. Environ. 1996, 58, 289–298. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Huete, A.R.; Didan, K.; Miura, T. Development of a two-band enhanced vegetation index without a blue band. Remote Sens. Environ. 2008, 112, 3833–3845. [Google Scholar] [CrossRef]
- Gao, B.-C. NDWI—A normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Singh, A.; Harrison, A. Standardized principal components. Int. J. Remote Sens. 1985, 6, 883–896. [Google Scholar] [CrossRef]
- Kauth, R.J.; Thomas, G.S. The Tasselled Cap—A Graphic Description of the Spectral-Temporal Development of Agricultural Crops as Seen by Landsat; Purdue University: West Lafayette, USA, 1976. [Google Scholar]
- Haralick, R.M.; Shanmugam, K.; Dinstein, I.H. Textural Features for Image Classification. IEEE Trans. Syst. Man Cybern. 1973, 3, 610–621. [Google Scholar] [CrossRef]
- Navulur, K. Multispectral Image Analysis Using the Object-Oriented Paradigm; CRC Press: Boca Raton, FL, USA, 2006; ISBN 9780429146305. [Google Scholar]
- Moore, I.D.; Grayson, R.B.; Ladson, A.R. Digital terrain modelling: A review of hydrological, geomorphological, and biological applications. Hydrol. Process. 1991, 5, 3–30. [Google Scholar] [CrossRef]
- Wilson, J.P.; Gallant, J.C. Terrain Analysis. Principles and Applications; John Wiley & Sons: New York, NY, USA, 2000; ISBN 978-0-471-32188-0. [Google Scholar]
- Riley, S.J.; DeGloria, S.D.; Elliot, R. Index that quantifies topographic heterogeneity. Intermt. J. Sci. 1999, 5, 23–27. [Google Scholar]
- Salford Systems Ltd. Salford Predictive Modeler 8.3; Salford Systems Ltd.: San Diego, USA, 2018. [Google Scholar]
- Hastie, T.; Tibshirani, R.; Friedman, J. The elements of statistical learning: Data mining, inference and prediction. Math. Intell. 2005, 27, 83–85. [Google Scholar] [CrossRef]
- Kandel, K.; Huettmann, F.; Suwal, M.K.; Ram Regmi, G.; Nijman, V.; Nekaris, K.A.I.; Lama, S.T.; Thapa, A.; Sharma, H.P.; Subedi, T.R. Rapid multi-nation distribution assessment of a charismatic conservation species using open access ensemble model GIS predictions: Red panda (Ailurus fulgens) in the Hindu-Kush Himalaya region. Biol. Conserv. 2015, 181, 150–161. [Google Scholar] [CrossRef]
- ESRI. How Fuzzy Overlay Works. Available online: http://desktop.arcgis.com/en/arcmap/10.5/tools/spatial-analyst-toolbox/how-fuzzy-overlay-works.htm (accessed on 1 May 2019).
- Key, C.H.; Benson, N.C. Landscape Assessment: Remote Sensing of Severity, the Normalized Burn Ratio. FIREMON: Fire Effects Monitoring and Inventory System RMRS-GTR-164-CD: LA 1-51; 2006. Available online: http://pubs.er.usgs.gov/publication/2002085 (accessed on 2 August 2019).
- Miller, J.D.; Thode, A.E. Quantifying burn severity in a heterogeneous landscape with a relative version of the delta Normalized Burn Ratio (dNBR). Remote Sens. Environ. 2007, 109, 66–80. [Google Scholar] [CrossRef]
- ESRI. How Fuzzy Membership Works. Available online: http://desktop.arcgis.com/en/arcmap/10.5/tools/spatial-analyst-toolbox/how-fuzzy-membership-works.htm (accessed on 4 April 2019).
- Elhorst, J.P. Spatial Panel Data Models. In Spatial Econometrics: From Cross-Sectional Data to Spatial Panels; Elhorst, J.P., Ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 37–93. ISBN 978-3-642-40340-8. [Google Scholar]
- Greene, W.H. Econometric Analysis, 5th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2002; ISBN 0-13-066189-9. [Google Scholar]
- Elhorst, J.P. Spatial Panel Data Models. In Handbook of Applied Spatial Analysis; Fischer, M.M., Getis, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 377–407. ISBN 978-3-642-03646-0. [Google Scholar]
- Bell, A.; Jones, K. Explaining Fixed Effects: Random Effects Modeling of Time-Series Cross-Sectional and Panel Data. PSRM 2015, 3, 133–153. [Google Scholar] [CrossRef]
- Hausman, J.A. Specification Tests in Econometrics. Econometrica 1978, 46, 1251. [Google Scholar] [CrossRef] [Green Version]
- Baltagi, B.H. Econometric Analysis of Panel Data, 4th ed.; John Wiley & Sons: Chichester, UK, 2008; ISBN 0-470-01456-3. [Google Scholar]
- Carter, G.A. Responses of Leaf Spectral Reflectance to Plant Stress. Am. J. Bot. 1993, 80, 239. [Google Scholar] [CrossRef]
- Carter, G.A.; Cibula, W.G.; Miller, R.L. Narrow-band Reflectance Imagery Compared with ThermalImagery for Early Detection of Plant Stress. J. Plant Physiol. 1996, 148, 515–522. [Google Scholar] [CrossRef]
- Muchoney, D.M.; Haack, B.N. Change Detection for Monitoring Forest Defoliation. Photogramm. Eng. Remote Sens. 1994, 60, 1243–1252. [Google Scholar]
- Moskal, L.M.; Franklin, S.E. Relationship between airborne multispectral image texture and aspen defoliation. Int. J. Remote Sens. 2004, 25, 2701–2711. [Google Scholar] [CrossRef]
- Lottering, R.; Mutanga, O.; Peerbhay, K.; Ismail, R. Detecting and mapping Gonipterus scutellatus induced vegetation defoliation using WorldView-2 pan-sharpened image texture combinations and an artificial neural network. J. Appl. Remote Sens. 2019, 13, 014513. [Google Scholar] [CrossRef]
- Olsson, P.-O.; Jönsson, A.M.; Eklundh, L. A new invasive insect in Sweden—Physokermes inopinatus: Tracing forest damage with satellite based remote sensing. For. Ecol. Manag. 2012, 285, 29–37. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Object features | Input data | Features 1 (No.) |
|---|---|---|
| Spectral features (32) | Blue, Green, Red, NIR, SWIR1, SWIR2 | Mean (6), StdDev (6) of the spectral bands Mean and StdDev of spectral indices (14) (IPVI [61] GEMI [62], ARVI [63], GVI [64], NDVI [65], EVI2 [66], NDWI [67] Principal components (6) [68] Greenness (2), Wetness (2) [69], Brightness (1), Max. diff. (1) |
| Surface texture-features (56) | Single bands and all bands in all directions | GLCMall dir. (Homogeneity (7), Contrast (7), Dissimilarity (7), Entropy (7), Mean (7), Angle second moment (7), StDev (7), Correlation (7)) [70,71] |
| Geometric features (3) | Objects | Area (1), Compactness (1), Asymmetry(1) [71] |
| Ancillary data (4) | ALOS PALSAR Forest data | Topographic Wetness Index [72], Topographic Position Index [73], Terrain Ruggedness Index [74], Forest types |
| Measure | Average | Overall accuracy | Specificity | Sensitivity | Precision | F1 statistic |
|---|---|---|---|---|---|---|
| Percent | 87.15 | 86.76 | 80.56 | 93.75 | 81.08 | 86.96 |
| Model | Constant | SPIt | SPIt − 1 | Tmaxt | Tmaxt − 1 | SoilMt | SoilMt − 1 | Firet − 1 | R2 |
|---|---|---|---|---|---|---|---|---|---|
| Common effects | 0.204 | 0.184 * | 0.070 ns | 0.463 * | 0.165 * | 0.144 * | −0.134 * | 0.171 * | 0.680 |
| Fixed effects | 0.290 ** | −0.039 ns | −0.080 ns | 0.762 ** | 0.367 * | 0.152 * | −0.072 ns | 0.154 * | 0.798 |
| Random effects | 0.210 ** | 0.153 * | 0.048 ns | 0.718 ** | 0.321 * | 0.146 * | −0.126 * | 0.194 * | 0.706 |
| Model | Constant | SPIt | SPIt − 1 | Tmaxt | Tmaxt − 1 | SoilMt | SoilMt − 1 | IIt | IIt − 1 | R2 |
|---|---|---|---|---|---|---|---|---|---|---|
| Common effects | −0.0169 ns | −0.085 ns | 0.030 | 0.385 * | 0.177 * | 0.032 ns | −0.163 ns | −0.059 ns | 0.105 ns | 0.210 |
| Fixed effects | −0.032 ns | −0.113 ns | 0.094 ns | 0.254 ns | 0.213 ** | 0.049 ns | −0.126 ns | −0.031 ns | 0.106 ns | 0.550 |
| Random effects | −0.017 ns | −0.101 ns | 0.117 ns | 0.330 * | 0.196 ** | 0.041 ns | −0.148 ns | −0.020 ns | 0.106 ns | 0.236 |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdi, O. Climate-Triggered Insect Defoliators and Forest Fires Using Multitemporal Landsat and TerraClimate Data in NE Iran: An Application of GEOBIA TreeNet and Panel Data Analysis. Sensors 2019, 19, 3965. https://doi.org/10.3390/s19183965
Abdi O. Climate-Triggered Insect Defoliators and Forest Fires Using Multitemporal Landsat and TerraClimate Data in NE Iran: An Application of GEOBIA TreeNet and Panel Data Analysis. Sensors. 2019; 19(18):3965. https://doi.org/10.3390/s19183965
Chicago/Turabian StyleAbdi, Omid. 2019. "Climate-Triggered Insect Defoliators and Forest Fires Using Multitemporal Landsat and TerraClimate Data in NE Iran: An Application of GEOBIA TreeNet and Panel Data Analysis" Sensors 19, no. 18: 3965. https://doi.org/10.3390/s19183965
APA StyleAbdi, O. (2019). Climate-Triggered Insect Defoliators and Forest Fires Using Multitemporal Landsat and TerraClimate Data in NE Iran: An Application of GEOBIA TreeNet and Panel Data Analysis. Sensors, 19(18), 3965. https://doi.org/10.3390/s19183965