Chain Modeling of Molecular Communications for Body Area Network


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- First, we develop a chain model of molecular communication based on biological signaling. The proposed model considers the biological interactions among hormone, Ca and neural signals, which is more general than the model of [8].
- Second, we propose an implementable amplify-and-forward relaying mechanism instead of decode-and-forward relaying as in [8]. In addition, multiple astrocytes are utilized instead of a nano-machine, elevating the reliability of relaying from Ca signaling to neural signaling.
- Third, based on the work in [8], we examined the relations between communication performance and more parameters of the proposed model. We also found that source coding is efficient in improving the communication performance, which may provide a guidance for nano-machine design.
2. Related Work
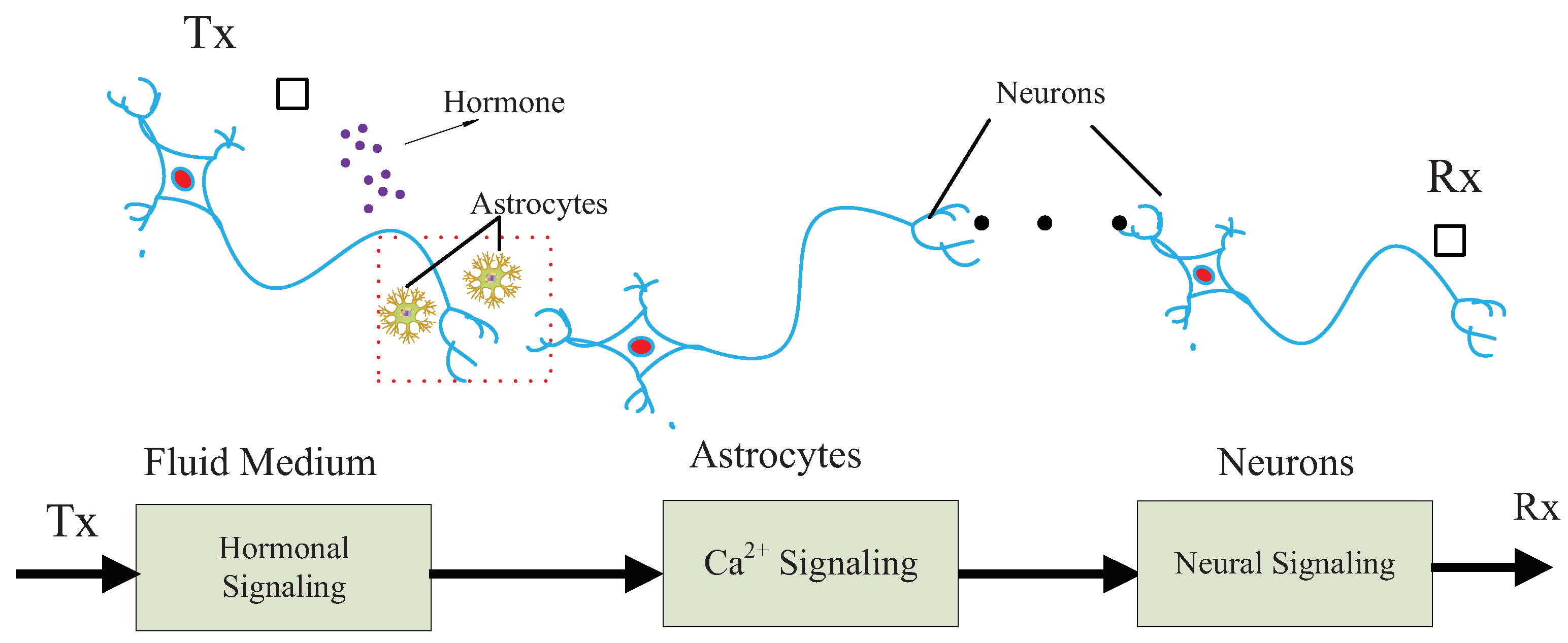
3. Overview of the Communication System
4. Hormonal Signaling Model
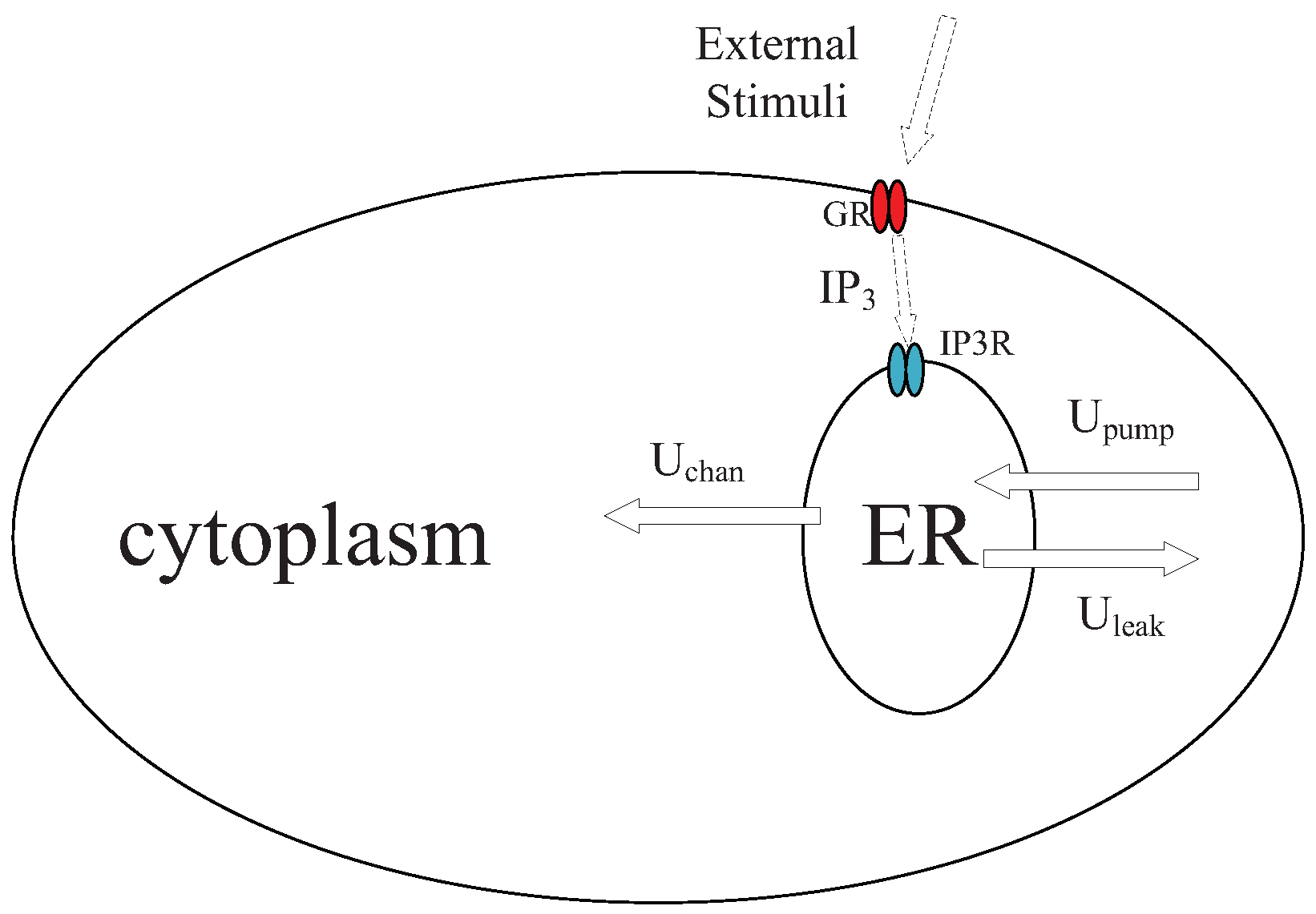
5. Calcium Signaling Model
6. Neural Signaling Model
6.1. Amplify-and-Forward Neural Firing
6.2. Axonal Transmission
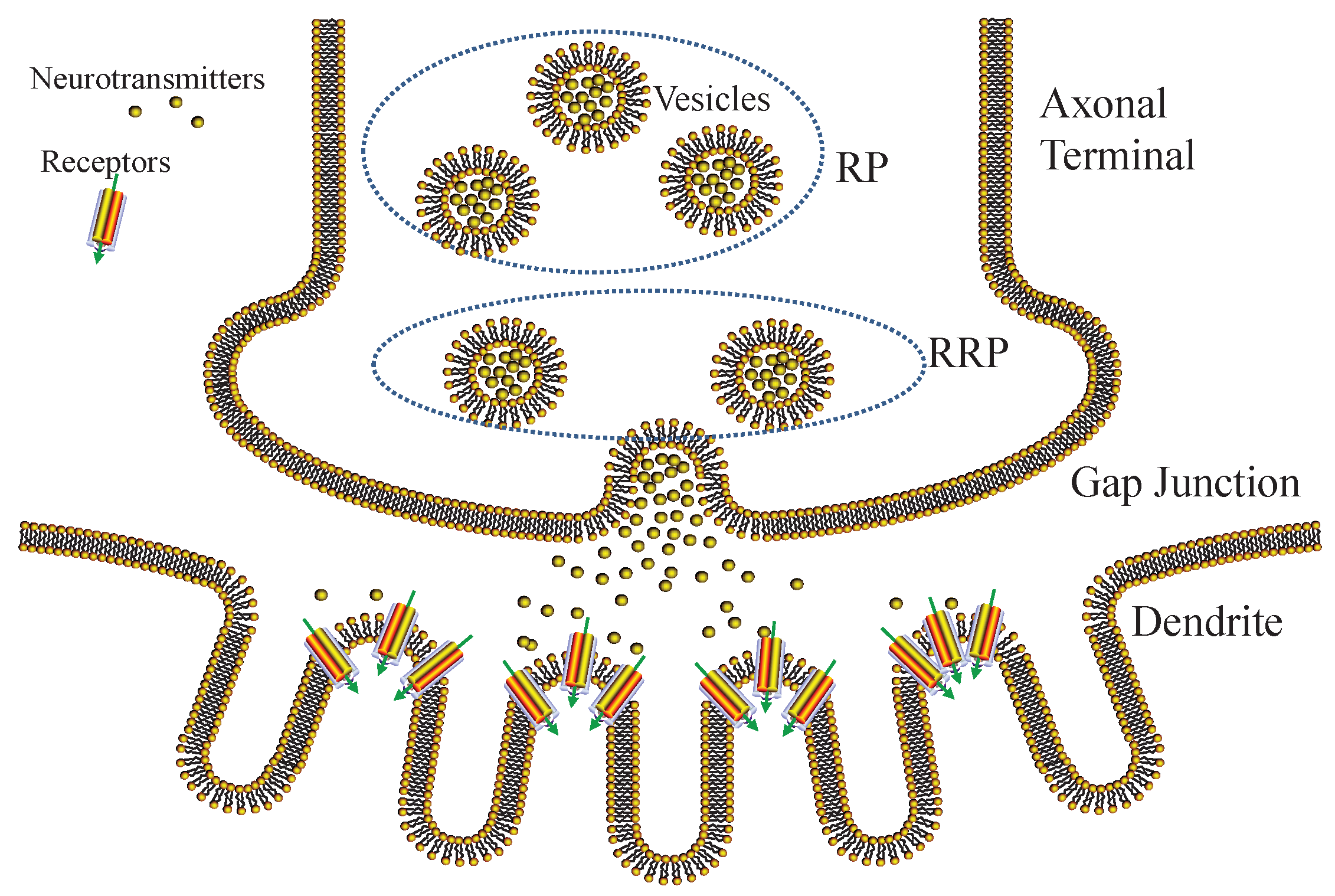
6.3. Gap Junctional Transmission
6.3.1. Vesicle Release
6.3.2. Queueing Model of Neurotransmitters
6.4. Postsynaptic Response
6.4.1. Postsynaptic Potential
6.4.2. Neural Decoding
7. Channel Capacity and Transmission Delay
7.1. Channel Capacity
7.2. Delay
8. Performance Evaluation
8.1. Simulation Design
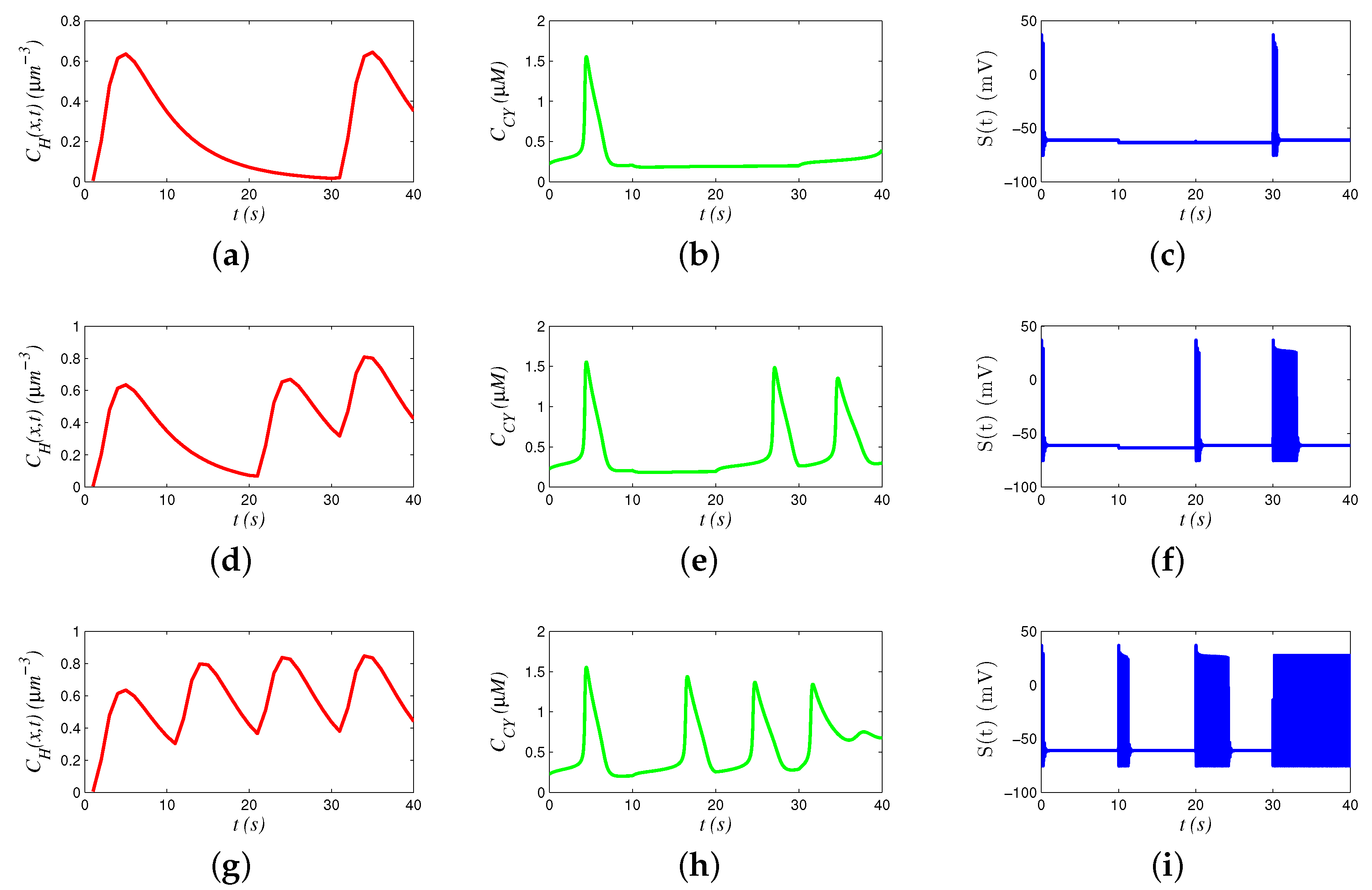
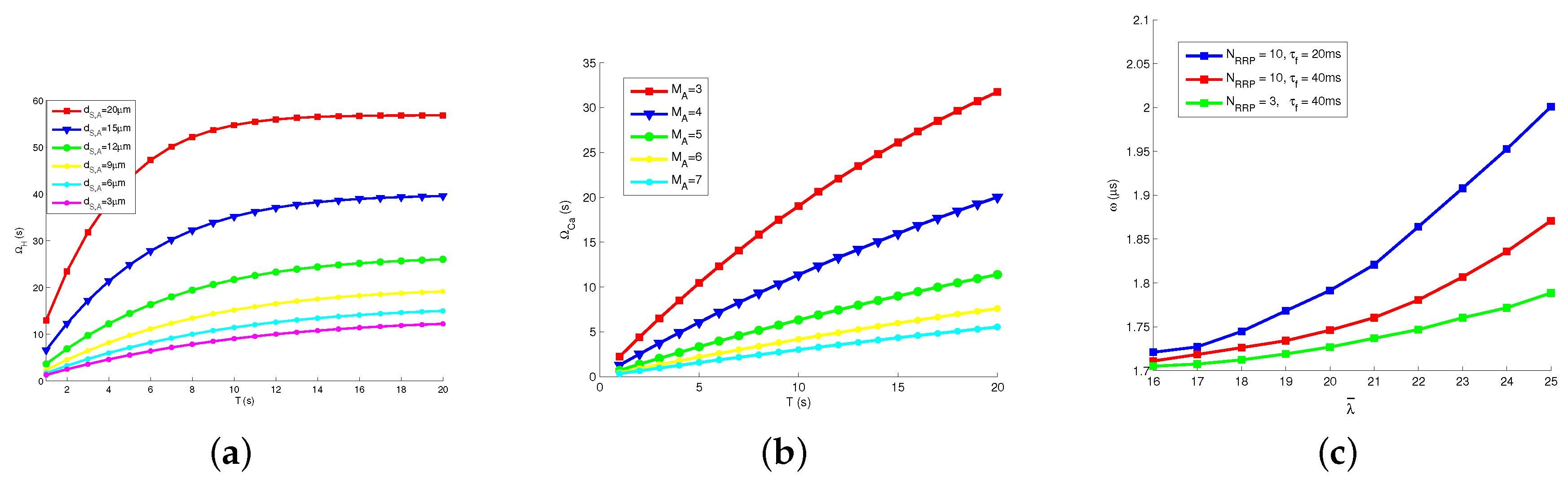
8.2. Behaviors of Signals
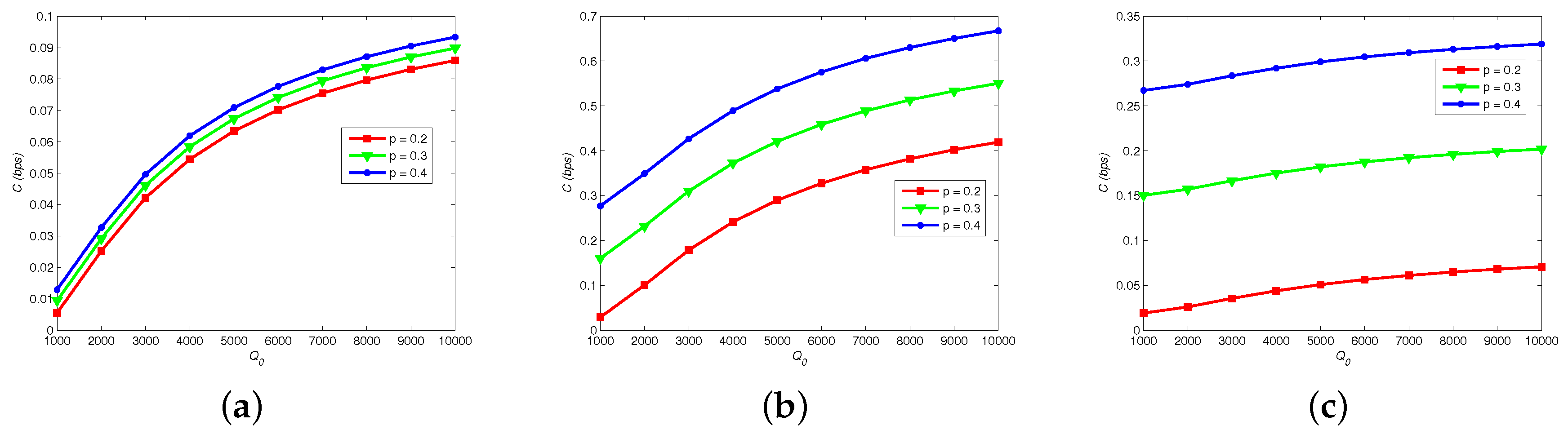
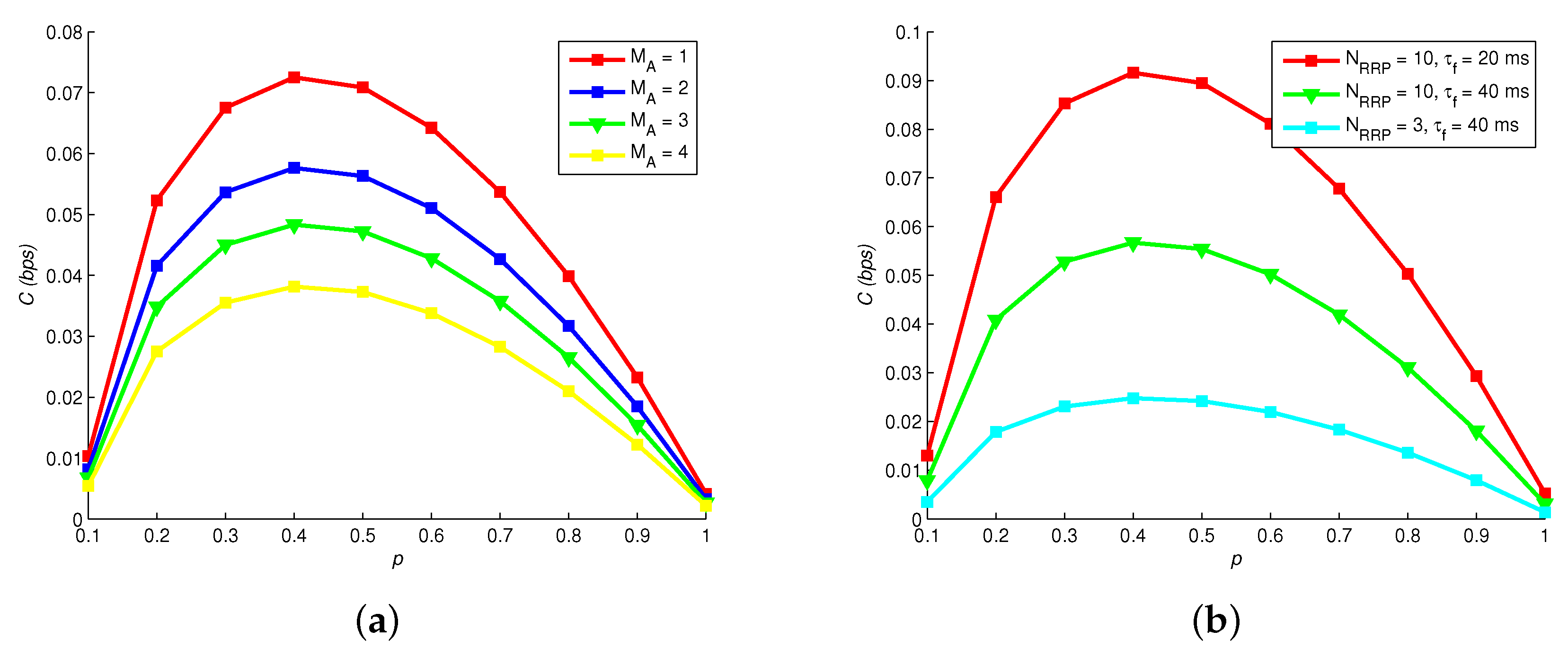
8.3. Channel Capacity
8.4. Transmission Delay
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cavallari, R.; Martelli, F.; Rosini, R.; Buratti, C.; Verdone, R. A survey on wireless body area networks: Technologies and design challenges. IEEE Commun. Surv. Tutor. 2014, 16, 1635–1657. [Google Scholar] [CrossRef]
- Nakano, T.; Eckford, A.W.; Haraguchi, T. Molecular Communication; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Wu, D.; Yan, J.; Wang, H.; Wu, D.; Wang, R. Social attribute aware incentive mechanism for device-to-device video distribution. IEEE Trans. Multimed. 2017, 19, 1908–1920. [Google Scholar] [CrossRef]
- Wu, D.; Yan, J.; Wang, H.; Wu, D.; Wang, R. Dynamic trust relationships aware data privacy protection in mobile crowd-sensing. IEEE Internet Things J. 2018, 5, 2958–2970. [Google Scholar] [CrossRef]
- Qi, H.; Peng, Y.; Gao, Q.; Zhang, C. Applications of nanomaterials in electrogenerated chemiluminescence biosensors. Sensors 2009, 9, 674–695. [Google Scholar] [CrossRef] [PubMed]
- Lecrux, C.; Hamel, E. Nanonetworks: The neurovascular unit in brain function and disease. Acta Physiol. 2011, 203, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Christian, C.A.; Mobley, J.L.; Moenter, S.M. Diurnal and estradiol-dependent changes in gonadotropin-releasing hormone neuron firing activity. Proc. Natl. Acad. Sci. USA 2005, 102, 15682–15687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, P.; Mao, Y.; Liu, Q.; Zhai, H. A diffusion-neuron hybrid channel for molecular communication. In Proceedings of the International Symposium on Intelligent Signal Processing and Communication Systems, Xiamen, China, 6–9 November 2017; pp. 855–860. [Google Scholar]
- Liu, Q.; Yang, K. Channel capacity analysis of a diffusion-based molecular communication system with ligand receptors. Int. J. Commun. Syst. 2015, 28, 1508–1520. [Google Scholar] [CrossRef]
- Satish, K.T.; Tadi, R.T.R.; Prabhat, K.U.; Daniel, B.D.C. Joint Optimization of Molecular Resource Allocation and Relay Positioning in Diffusive Nanonetworks. IEEE Access 2018, 66, 67681–67687. [Google Scholar]
- Jamali, V.; Ahmadzadeh, A.; Farsad, N.; Schober, R. Constant-composition codes for maximum likelihood detection without csi in diffusive molecular communications. IEEE Trans. Commun. 2018, 66, 1981–1995. [Google Scholar] [CrossRef]
- Bicen, A.O.; Akyildiz, I.F.; Balasubramaniam, S.; Koucheryavy, Y. Linear channel modeling and error analysis for intra/inter cellular Ca2+ molecular communication. IEEE Trans. NanoBiosci. 2016, 15, 488–498. [Google Scholar] [CrossRef]
- Taynnan, B.M.; Balasubramaniam, S.; Jennings, B. Comparative end-to-end analysis of Ca2+ signaling-based molecular communication in biological tissues. IEEE Trans. Commun. 2015, 63, 5128–5142. [Google Scholar] [CrossRef]
- He, P.; Nakano, T.; Mao, Y.; Lio, P.; Liu, Q.; Yang, K. Stochastic Channel Switching of Frequency-encoded Signals in Molecular Communication Networks. IEEE Commun. Lett. 2018, 22, 332–335. [Google Scholar] [CrossRef]
- Malak, D.; Akan, O. A Communication Theoretical Analysis of Synaptic Multiple-Access Channel in Hippocampal-Cortical Neurons. IEEE Trans. Commun. 2013, 61, 2457–2467. [Google Scholar] [CrossRef]
- Aghababaiyan, K.; Shah-Mansouri, V.; Maham, B. Axonal channel capacity in neuro-spike communication. IEEE Trans. NanoBiosci. 2018, 17, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Bilgin, B.A.; Akan, O.B. A fast algorithm for analysis of molecular communication in artificial synapse. IEEE Trans. NanoBiosci. 2017, 16, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Liu, Q.; Wang, H.; Yang, Q.; Wang, R. Cache Less for More: Exploiting Cooperative Video Caching and Delivery in D2D Communications. IEEE Trans. Multimed. 2018. [Google Scholar] [CrossRef]
- Li, Y.; Liu, J.; Cao, B.; Wang, C. Joint Optimization of Radio and Virtual Machine Resources with Uncertain User Demands in Mobile Cloud Computing. IEEE Trans. Multimed. 2017, 20, 2427–2438. [Google Scholar] [CrossRef]
- Wu, D.; Zhang, F.; Wang, H.; Wang, R. Security-oriented opportunistic data forwarding in mobile social networks. Future Gener. Comput. Syst. 2017, 87, 803–815. [Google Scholar] [CrossRef]
- Li, Y.; Liang, Y.; Liu, Q.; Wang, H. Resources Allocation in Multi-Cell D2D Communications for Internet of Things. IEEE Internet Things J. 2018, 5, 4100–4108. [Google Scholar] [CrossRef]
- Wang, R.; Yan, J.; Wu, D.; Wang, H.; Yang, Q. Knowledge-centric edge computing based on virtualized d2d communication systems. IEEE Commun. Mag. 2018, 56, 32–38. [Google Scholar] [CrossRef]
- Wu, D.; Shi, H.; Wang, H.; Wang, R.; Fang, H. A Feature based Learning System for Internet of Things Applications. IEEE Internet Things J. 2018. [Google Scholar] [CrossRef]
- Pardridge, W.M.; Mietus, L.J. Transport of steroid hormones through the rat blood-brain barrier. Endocrinology 1979, 105, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Villalobos, C.; Lucia, N.; Javier, G. Anterior pituitary thyrotropes are multifunctional cells. Am. J. Physiol. Endocrinol. Metab. 2004, 287, 1166–1170. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, B.A.; Weiss, S. Generation of neurons and astrocytes from isolated cells of the adult mammalian central nervous system. Science 1992, 255, 1707–1710. [Google Scholar] [CrossRef] [PubMed]
- Roth, B.J.; Yagodin, S.V.; Holtzclaw, L.; Russell, J.T. A mathematical model of agonist-induced propagation of calcium waves in astrocytes. Cell Calcium 1995, 87, 53–64. [Google Scholar] [CrossRef]
- Flavio, F. Network Neuroscience; Elsevier Academic Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Palmer, J.D.; Edmunds, L.N. Cellular and molecular bases of biological clocks. Bioscience 1988, 38, 161–162. [Google Scholar] [CrossRef]
- Kolesnick, R.N.; Gershengorn, M.C. Direct evidence that burst but not sustained secretion of prolactin stimulated by thyrotropin-releasing hormone is dependent on elevation of cytoplasmic calcium. J. Biol. Chem. 1985, 260, 5217–5220. [Google Scholar]
- Nakano, T.; Okaie, Y.; Liu, J.Q. Channel Model and Capacity Analysis of Molecular Communication with Brownian Motion. IEEE Commun. Lett. 2012, 16, 797–800. [Google Scholar] [CrossRef]
- Di, G.A.; Barbi, M.; Chillemi, S.; Alloisio, S.; Nobile, M. Calcium signalling in astrocytes and modulation of neural activity. Biosystems 2007, 89, 74–83. [Google Scholar]
- Navarrete, M.; Araque, A. Endocannabinoids mediate neuron-astrocyte communication. Neuron 2008, 57, 883–893. [Google Scholar] [CrossRef]
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfister, J.P.; Toyoizumi, T.; Barber, D.; Gerstner, W. Optimal spike-timing-dependent plasticity for precise action potential firing in supervised learning. Neural Comput. 2006, 18, 1318–1348. [Google Scholar] [CrossRef] [PubMed]
- Rall, W. Optimal Cable theory for dendritic neurons. In Methods in Neuronal Modeling; MIT Press: Cambridge, MA, USA, 1989; pp. 9–92. [Google Scholar]
- Sreekumaran, E.; Ramakrishna, T.; Madhav, T.R.; Anandh, D.; Prabhu, B.M.; Sulekha, S.; Bindu, P.N.; Raju, T.R. Loss of dendritic connectivity in CA1, CA2, and CA3 neurons in hippocampus in rat under aluminum toxicity: Antidotal effect of pyridoxine. Brain Res. Bull. 2003, 59, 421–427. [Google Scholar] [CrossRef]
- Matveev, V.; Wang, X.J. Implications of all-or-none synaptic transmission and short-term depression beyond vesicle depletion: A computational study. Neuroscience 2000, 24, 1575–1588. [Google Scholar] [CrossRef]
- Pyle, J.L.; Kavalali, E.T.; Piedras-Rentería, E.S.; Tsien, R.W. Rapid reuse of readily releasable pool vesicles at hippocampal synapses. Neuron 2000, 28, 221–231. [Google Scholar] [CrossRef]
- Li, W.; Fretwell, R.J.; Kouvatsos, D.D. Performance Analysis of Queues with Batch Poisson Arrival and GE Service. In Proceedings of the 2011 7th International Conference on Wireless Communications, Networking and Mobile Computing, Wuhan, China, 23–25 September 2011; pp. 1033–1036. [Google Scholar]
- Rall, W. Distinguishing theoretical synaptic potentials computed for different soma-dendritic distributions of synaptic input. Neurophysiology 1967, 30, 1138–1168. [Google Scholar] [CrossRef] [PubMed]
- Bekkers, J.M. Origin of variability in quantal size in cultured hippocampal neurons and hippocampal slices. Neurobiology 1967, 87, 5359–5362. [Google Scholar] [CrossRef]
- Massey, J.F.J. The Kolmogorov-Smirnov test for goodness of fit. J. Am. Stat. Assoc. 1951, 46, 68–78. [Google Scholar] [CrossRef]
- Bogazzi, F.; Bartalena, L.; Brogioni, S.; Burelli, A.; Manetti, L.; Tanda, M.L.; Gasperi, M.; Martino, E. Thyroid vascularity and blood flow are not dependent on serum thyroid hormone levels: Studies in vivo by color flow Doppler sonography. Eur. J. Endocrinol. 1999, 140, 452–456. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, P.; Han, X.; Liu, H. Chain Modeling of Molecular Communications for Body Area Network. Sensors 2019, 19, 395. https://doi.org/10.3390/s19020395
He P, Han X, Liu H. Chain Modeling of Molecular Communications for Body Area Network. Sensors. 2019; 19(2):395. https://doi.org/10.3390/s19020395
Chicago/Turabian StyleHe, Peng, Xiaojuan Han, and Hanyong Liu. 2019. "Chain Modeling of Molecular Communications for Body Area Network" Sensors 19, no. 2: 395. https://doi.org/10.3390/s19020395
APA StyleHe, P., Han, X., & Liu, H. (2019). Chain Modeling of Molecular Communications for Body Area Network. Sensors, 19(2), 395. https://doi.org/10.3390/s19020395