Predictive Modeling of a Leaf Conceptual Midpoint Quasi-Color (CMQ) Using an Artificial Neural Network


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Datasets
2.2. Quantification of Pigments and Measuring Leaf Color
2.3. Parametric Models for Predicting CMQ Coordinates
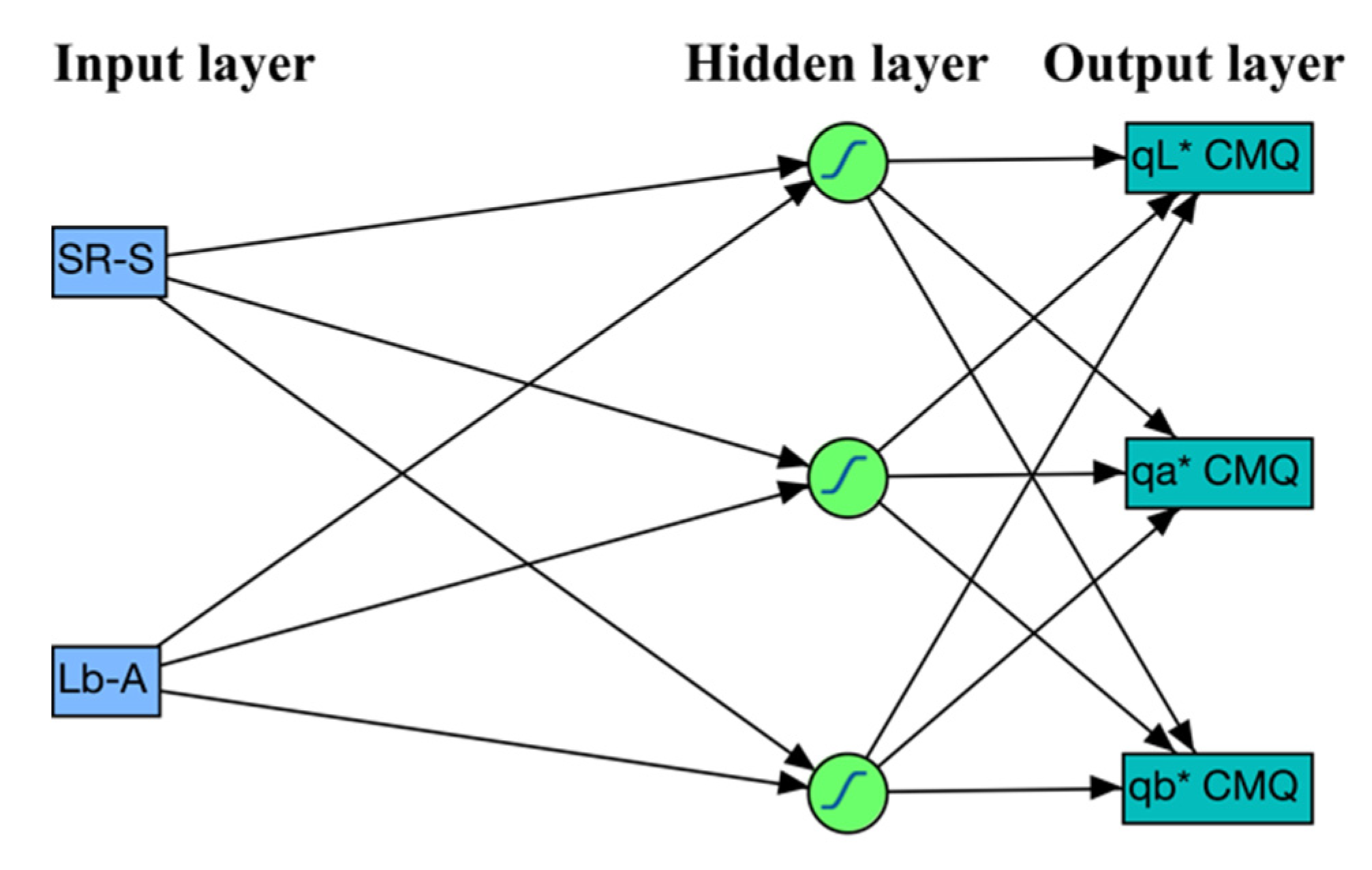
2.4. ANN Models for Predicting CMQ Coordinates
2.5. Calculating CMQ Coordinates from the Content of Pigments
2.6. Testing the ANN Model on Other Plant Species
2.7. Statistical Analyses
3. Results and Discussion
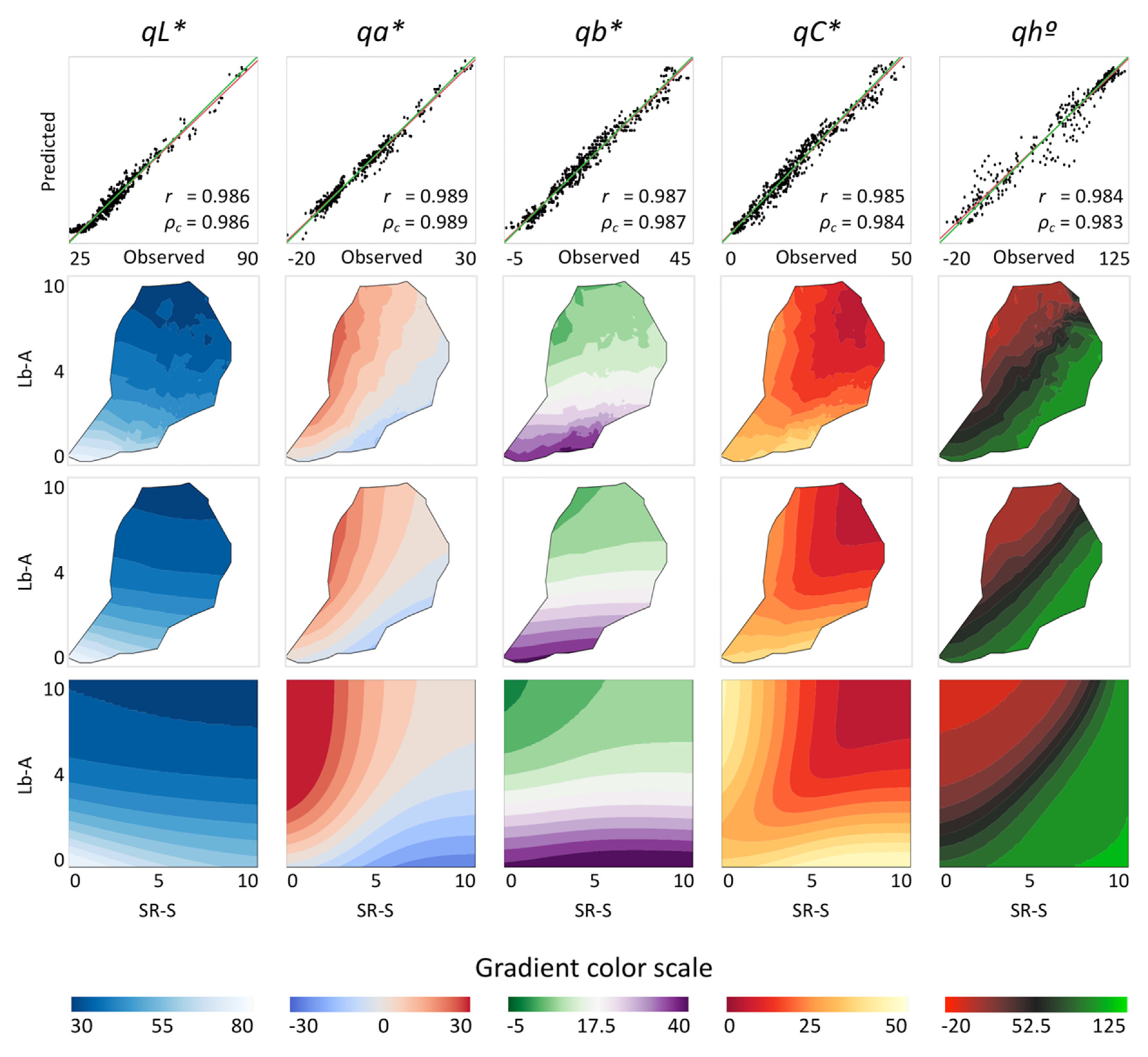
3.1. Content of Pigments in Lettuce Leaves, Leaf Color and Comparison of Models to Predict CMQ Coordinates
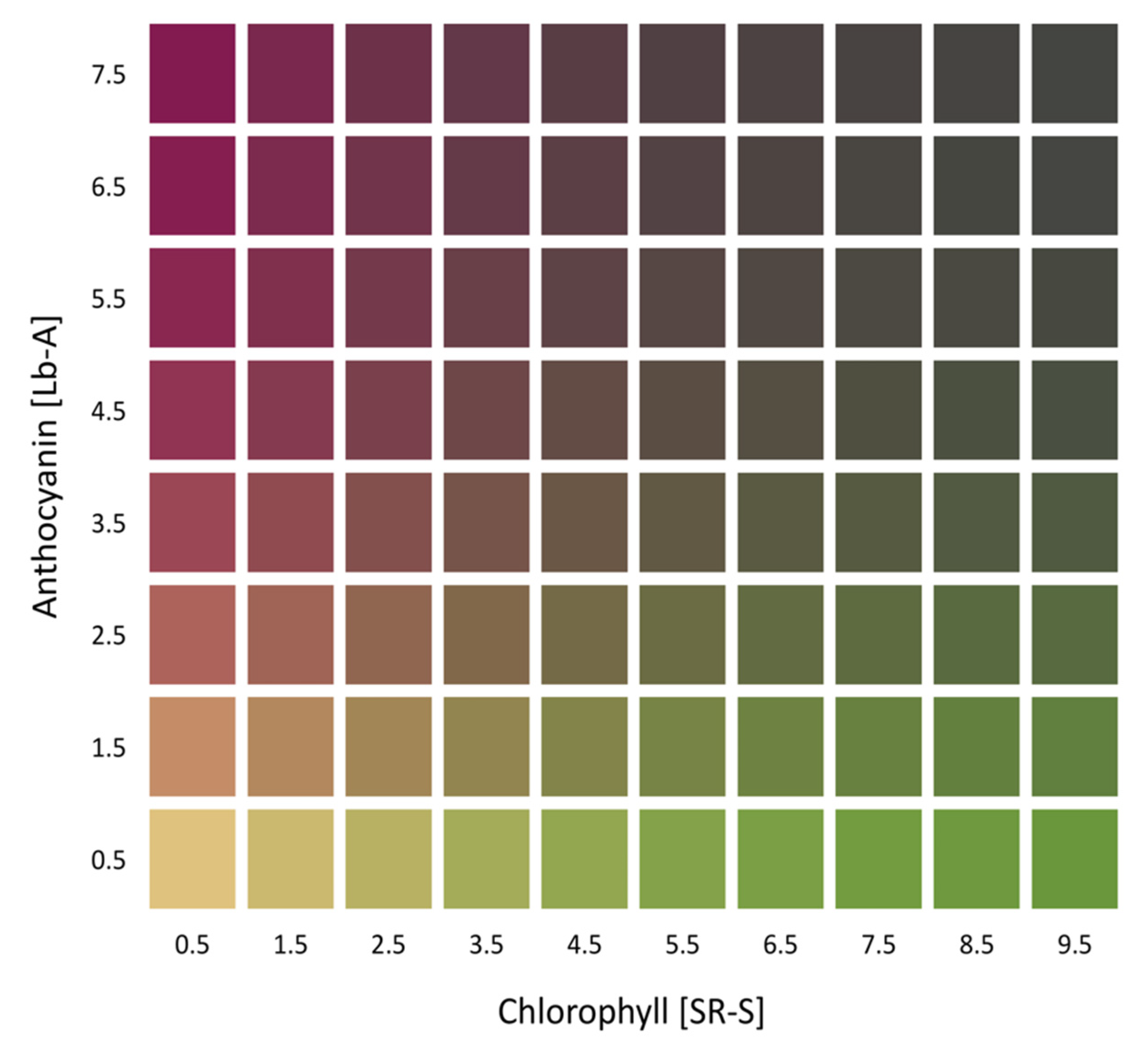
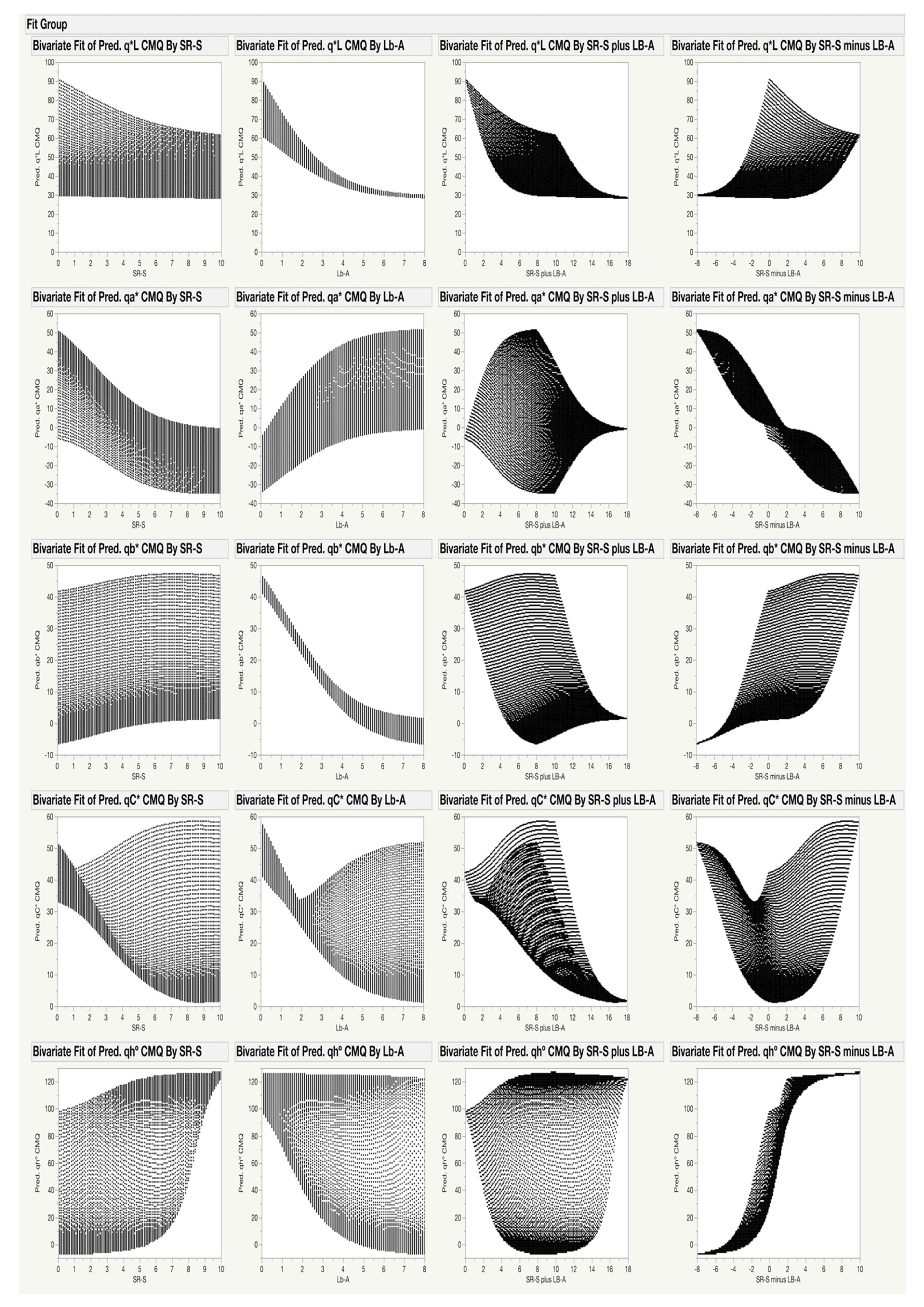
3.2. Modeling the Gamut of CMQ Coordinates
3.3. Performance of the ANN Model on Herbaceous Leaves
3.4. Dorsiventral Distribution of Pigments
3.5. Application of CMQ in Plant Research
4. Conclusions
Supplementary Materials
Funding
Conflicts of Interest
References
- Boldt, J.K.; Meyer, M.H.; Erwin, J.E. Foliar anthocyanins: A horticultural review. Hortic. Rev. 2014, 42, 209–252. [Google Scholar]
- Mahlein, A.-K. Plant disease detection by imaging sensors—Parallels and specific demands for precision agriculture and plant phenotyping. Plant. Dis. 2016, 100, 241–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simko, I.; Jimenez-Berni, J.A.; Sirault, X.R. Phenomic approaches and tools for phytopathologists. Phytopathology 2017, 107, 6–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Junker, L.V.; Ensminger, I. Relationship between leaf optical properties, chlorophyll fluorescence and pigment changes in senescing Acer saccharum leaves. Tree Physiol. 2016, 36, 694–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braitmaier, M.; Diepstraten, J.; Ertl, T. Real-time rendering of seasonal influenced trees. In Proceedings of the Theory and Practice of Computer Graphics, Bournemouth, UK, 8–10 June 2004; pp. 152–159. [Google Scholar]
- Miao, T.; Zhao, C.; Guo, X.; Lu, S. A framework for plant leaf modeling and shading. Math. Comput. Model. 2013, 58, 710–718. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, C.; Lu, S.; Guo, X. Survey on modeling and visualization of plant leaf color. In Proceedings of the Third International Symposium on Plant Growth Modeling, Simulation, Visualization and Applications, Beijing, China, 9–13 November 2009; pp. 417–424. [Google Scholar]
- Yi, W.-L.; He, H.-J.; Wang, L.-P.; Yang, H.-Y. Modeling and simulation of leaf color based on virtual rice. In Proceedings of the International Conference on Materials, Manufacturing and Mechanical Engineering, Beijing, China, 30–31 December 2016; pp. 334–343. [Google Scholar]
- Zhou, N.; Dong, W.; Mei, X. Realistic simulation of seasonal variant maples. In Proceedings of the Second International Symposium on Plant Growth Modeling and Applications, Beijing, China, 13–17 November 2006; pp. 295–301. [Google Scholar]
- Lancaster, J.E.; Lister, C.E.; Reay, P.F.; Triggs, C.M. Influence of pigment composition on skin color in a wide range of fruit and vegetables. J. Am. Soc. Hortic. Sci. 1997, 122, 594–598. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, L.; He, H.; Guo, X. Visual simulation of cucumber leaf color based on the relative content of chlorophyll. Trans. Chin. Soc. Agric. Mach. 2014, 45, 250–254. [Google Scholar]
- Shen, J.; Zou, Z.; Zhang, X.; Zhou, L.; Wang, Y.; Fang, W.; Zhu, X. Metabolic analyses reveal different mechanisms of leaf color change in two purple-leaf tea plant (Camellia sinensis L.) cultivars. Hortic. Res. 2018, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, D.; Shi, P.; Omasa, K. Estimating rice chlorophyll content and leaf nitrogen concentration with a digital still color camera under natural light. Plant. Methods. 2014, 10, 36. [Google Scholar] [CrossRef] [Green Version]
- Gazula, A.; Kleinhenz, M.D.; Scheerens, J.C.; Ling, P.P. Anthocyanin levels in nine lettuce (Lactuca sativa) cultivars: Influence of planting date and relations among analytic, instrumented, and visual assessments of color. HortScience 2007, 42, 232–238. [Google Scholar] [CrossRef]
- Gonçalves, B.; Silva, A.P.; Moutinho-Pereira, J.; Bacelar, E.; Rosa, E.; Meyer, A.S. Effect of ripeness and postharvest storage on the evolution of colour and anthocyanins in cherries (Prunus avium L.). Food Chem. 2007, 103, 976–984. [Google Scholar]
- Fairchild, M.D. Color Appearance Models, 3rd ed.; John Wiley & Sons: Chichester, UK, 2013. [Google Scholar]
- Simko, I.; Hayes, R.J.; Mou, B.; McCreight, J.D. Lettuce and Spinach. In Yield Gains in Major, U.S. Field Crops; Smith, S., Diers, B., Specht, J., Carver, B., Eds.; American Society of Agronomy, Crop Science Society of America, and Soil Science Society of America: Madison, WI, USA, 2014; pp. 53–86. [Google Scholar]
- Simko, I.; Hayes, R.J.; Furbank, R.T. Non-destructive phenotyping of lettuce plants in early stages of development with optical sensors. Front. Plant. Sci. 2016, 7, 1985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parisi, G.I.; Kemker, R.; Part, J.L.; Kanan, C.; Wermter, S. Continual lifelong learning with neural networks: A review. Neural Netw. 2019, 113, 54–71. [Google Scholar] [CrossRef] [PubMed]
- Lazarovits, J.; Sindhwani, S.; Tavares, A.J.; Zhang, Y.; Song, F.; Audet, J.; Krieger, J.R.; Syed, A.M.; Stordy, B.; Chan, W.C. Supervised learning and mass spectrometry predicts the in vivo fate of nanomaterials. ACS Nano 2019, 13, 8023–8034. [Google Scholar] [CrossRef] [PubMed]
- Hinton, G.E.; Salakhutdinov, R.R. Reducing the dimensionality of data with neural networks. Science 2006, 313, 504–507. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Patuwo, B.E.; Hu, M.Y. Forecasting with artificial neural networks: The state of the art. Int. J. Forecast. 1998, 14, 35–62. [Google Scholar] [CrossRef]
- Arab, M.M.; Yadollahi, A.; Shojaeiyan, A.; Ahmadi, H. Artificial neural network genetic algorithm as powerful tool to predict and optimize in vitro proliferation mineral medium for G × N15 rootstock. Front. Plant. Sci. 2016, 7, 1526. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, M.S.; Tan, J.S.; Mohamad, R.; Mokhtar, M.N.; Ariff, A.B. Comparative analyses of response surface methodology and artificial neural network on medium optimization for Tetraselmis sp. FTC209 grown under mixotrophic condition. Sci. World J. 2013, 2013, 948940. [Google Scholar] [CrossRef] [Green Version]
- McCann, M.C.; Defernez, M.; Urbanowicz, B.R.; Tewari, J.C.; Langewisch, T.; Olek, A.; Wells, B.; Wilson, R.H.; Carpita, N.C. Neural network analyses of infrared spectra for classifying cell wall architectures. Plant. Physiol. 2007, 143, 1314–1326. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.-Y. Application of artificial neural network for detecting Phalaenopsis seedling diseases using color and texture features. Comput. Electron. Agric. 2007, 57, 3–11. [Google Scholar] [CrossRef]
- Toda, Y.; Okura, F. How convolutional neural networks diagnose plant disease. Plant. Phenomics. 2019, 2019, 9237136. [Google Scholar] [CrossRef]
- Ghodsvali, A.; Farzaneh, V.; Bakhshabadi, H.; Zare, Z.; Karami, Z.; Mokhtarian, M.; Carvalho, I.S. Screening of the aerodynamic and biophysical properties of barley malt. Int. Agrophysics. 2016, 30, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Ferentinos, K.; Albright, L. Predictive neural network modeling of pH and electrical conductivity in deep–trough hydroponics. Trans. Asae 2002, 45, 2007. [Google Scholar] [CrossRef] [Green Version]
- Moon, T.; Ahn, T.I.; Son, J.E. Forecasting root-zone electrical conductivity of nutrient solutions in closed-loop soilless cultures via a recurrent neural network using environmental and cultivation information. Front. Plant. Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Cavallo, D.P.; Cefola, M.; Pace, B.; Logrieco, A.F.; Attolico, G. Non-destructive automatic quality evaluation of fresh-cut iceberg lettuce through packaging material. J. Food Eng. 2018, 223, 46–52. [Google Scholar] [CrossRef]
- Taghadomi-Saberi, S.; Omid, M.; Emam-Djomeh, Z.; Ahmadi, H. Evaluating the potential of artificial neural network and neuro-fuzzy techniques for estimating antioxidant activity and anthocyanin content of sweet cherry during ripening by using image processing. J. Sci. Food Agric. 2014, 94, 95–101. [Google Scholar] [CrossRef]
- Wang, Z.-W.; Duan, H.-W.; Hu, C.-Y. Modelling the respiration rate of guava (Psidium guajava L.) fruit using enzyme kinetics, chemical kinetics and artificial neural network. Eur. Food Res. Technol. 2009, 229, 495–503. [Google Scholar] [CrossRef]
- Gonzalez-Fernandez, I.; Iglesias-Otero, M.; Esteki, M.; Moldes, O.; Mejuto, J.; Simal-Gandara, J. A critical review on the use of artificial neural networks in olive oil production, characterization and authentication. Crit. Rev. Food Sci. Nutr. 2019, 59, 1913–1926. [Google Scholar] [CrossRef]
- The PLANTS Database. United States Department of Agriculture, Natural Resources Conservation Service, Washington, D.C. Available online: https://plants.sc.egov.usda.gov (accessed on 14 November 2018).
- Parry, C.; Blonquist, J.M., Jr.; Bugbee, B. In situ measurement of leaf chlorophyll concentration: Analysis of the optical/absolute relationship. Plant Cell Environ. 2014, 37, 2508–2520. [Google Scholar] [CrossRef]
- van den Berg, A.K.; Perkins, T.D. Nondestructive estimation of anthocyanin content in autumn sugar maple leaves. HortScience 2005, 40, 685–686. [Google Scholar] [CrossRef] [Green Version]
- Gotwalt, C.M. JMP Neural Network Methodology; SAS Institute: Cary, NC, USA, 2012. [Google Scholar]
- Lin, L.I.-K. A concordance correlation coefficient to evaluate reproducibility. Biometrics 1989, 255–268. [Google Scholar] [CrossRef]
- Basri, M.; Abd Rahman, R.N.Z.R.; Ebrahimpour, A.; Salleh, A.B.; Gunawan, E.R.; Abd Rahman, M.B. Comparison of estimation capabilities of response surface methodology (RSM) with artificial neural network (ANN) in lipase-catalyzed synthesis of palm-based wax ester. BMC Biotechnol. 2007, 7, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, K.M.; Survase, S.A.; Saudagar, P.S.; Lele, S.; Singhal, R.S. Comparison of artificial neural network (ANN) and response surface methodology (RSM) in fermentation media optimization: Case study of fermentative production of scleroglucan. Biochem. Eng. J. 2008, 41, 266–273. [Google Scholar] [CrossRef]
- Pakalapati, H.; Arumugasamy, S.K.; Khalid, M. Comparison of response surface methodology and feedforward neural network modeling for polycaprolactone synthesis using enzymatic polymerization. Biocatal. Agric. Biotechnol. 2019, 18, 101046. [Google Scholar] [CrossRef]
- Hatier, J.-H.B.; Gould, K.S. Black coloration in leaves of Ophiopogon planiscapus ‘Nigrescens’. Leaf optics, chromaticity, and internal light gradients. Funct. Plant. Biol. 2007, 34, 130–138. [Google Scholar] [CrossRef]
- Mampholo, B.M.; Maboko, M.M.; Soundy, P.; Sivakumar, D. Phytochemicals and overall quality of leafy lettuce (Lactuca sativa L.) varieties grown in closed hydroponic system. J. Food Qual. 2016, 39, 805–815. [Google Scholar] [CrossRef]
- Murakami, P.F.; Turner, M.R.; van den Berg, A.K.; Schaberg, P.G. An Instructional Guide for Leaf Color Analysis Using Digital Imaging Software; General Technical Report NE-327; US Department of Agriculture, Forest Service, Northeastern Research Station: Newton Square, PA, USA, 2005.
- Zhang, X.; Goldberg, M.D. Monitoring fall foliage coloration dynamics using time-series satellite data. Remote Sens. Environ. 2011, 115, 382–391. [Google Scholar] [CrossRef]
- Mochizuki, S.; Cai, D.; Komori, T.; Kimura, H.; Hori, R. Virtual autumn coloring system based on biological and fractal model. In Proceedings of the Ninth Pacific Conference on Computer Graphics and Applications. Pacific Graphics, Tokyo, Japan, 16–18 October 2001; pp. 348–354. [Google Scholar]
- Owen, W.G.; Lopez, R.G. Customizing crop foliage color with LEDs: Red leaf lettuce. Greenh. Grow. 2015, 46–52. [Google Scholar]
- Simko, I. Genetic variation in response to N, P, or K deprivation in baby leaf lettuce. Horticulturae 2020, 6, 15. [Google Scholar] [CrossRef]
- Camejo, D.; Frutos, A.; Mestre, T.C.; del Carmen Piñero, M.; Rivero, R.M.; Martínez, V. Artificial light impacts the physical and nutritional quality of lettuce plants. Hortic. Environ. Biotechnol. 2020, 61, 69–82. [Google Scholar] [CrossRef]
- Owen, W.G.; Lopez, R.G. End-of-production supplemental lighting with red and blue light-emitting diodes (LEDs) influences red pigmentation of four lettuce varieties. HortScience 2015, 50, 676–684. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Training Set (480 Samples) | Testing Set (124 Samples) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R2 | RMSE | R2 | RMSE | |||||||||
| qL* | qa* | qb* | qL* | qa* | qb* | qL* | qa* | qb* | qL* | qa* | qb* | |
| Full factorial | 0.945 | 0.844 | 0.917 | 2.37 | 2.72 | 3.04 | 0.963 | 0.857 | 0.913 | 2.31 | 2.65 | 3.31 |
| Quadratic polynomial | 0.962 | 0.957 | 0.971 | 2.00 | 1.73 | 1.84 | 0.978 | 0.945 | 0.965 | 1.83 | 1.77 | 2.13 |
| Response surface | 0.970 | 0.964 | 0.972 | 1.76 | 1.58 | 1.82 | 0.982 | 0.950 | 0.965 | 1.69 | 1.66 | 2.15 |
| Artificial neural network | 0.969 | 0.979 | 0.974 | 1.81 | 1.23 | 1.71 | 0.981 | 0.979 | 0.971 | 1.71 | 1.08 | 1.91 |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simko, I. Predictive Modeling of a Leaf Conceptual Midpoint Quasi-Color (CMQ) Using an Artificial Neural Network. Sensors 2020, 20, 3938. https://doi.org/10.3390/s20143938
Simko I. Predictive Modeling of a Leaf Conceptual Midpoint Quasi-Color (CMQ) Using an Artificial Neural Network. Sensors. 2020; 20(14):3938. https://doi.org/10.3390/s20143938
Chicago/Turabian StyleSimko, Ivan. 2020. "Predictive Modeling of a Leaf Conceptual Midpoint Quasi-Color (CMQ) Using an Artificial Neural Network" Sensors 20, no. 14: 3938. https://doi.org/10.3390/s20143938
APA StyleSimko, I. (2020). Predictive Modeling of a Leaf Conceptual Midpoint Quasi-Color (CMQ) Using an Artificial Neural Network. Sensors, 20(14), 3938. https://doi.org/10.3390/s20143938