Toward Sub-Diffraction Imaging of Single-DNA Molecule Sensors Based on Stochastic Switching Localization Microscopy




Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
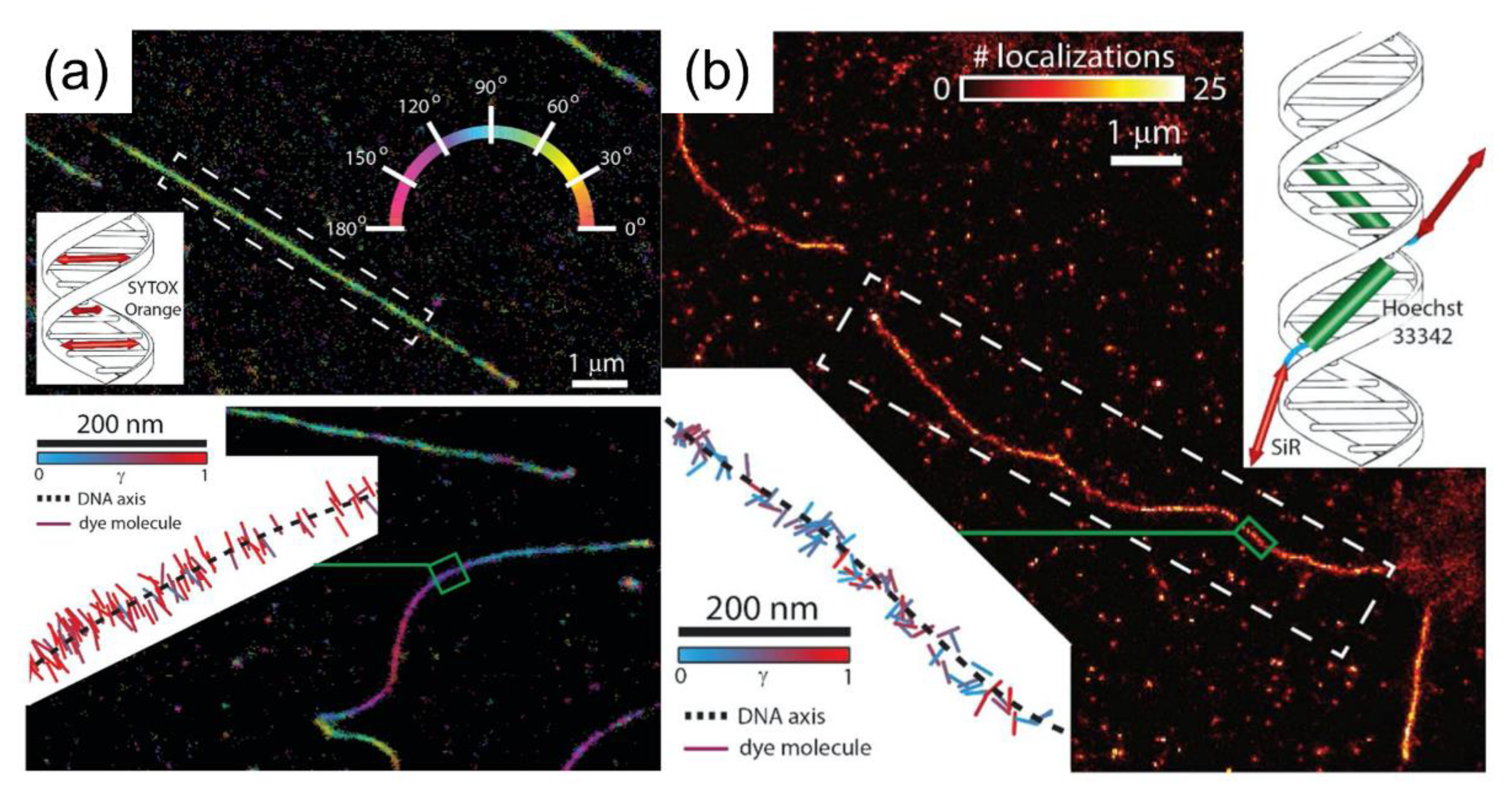{kind=link}
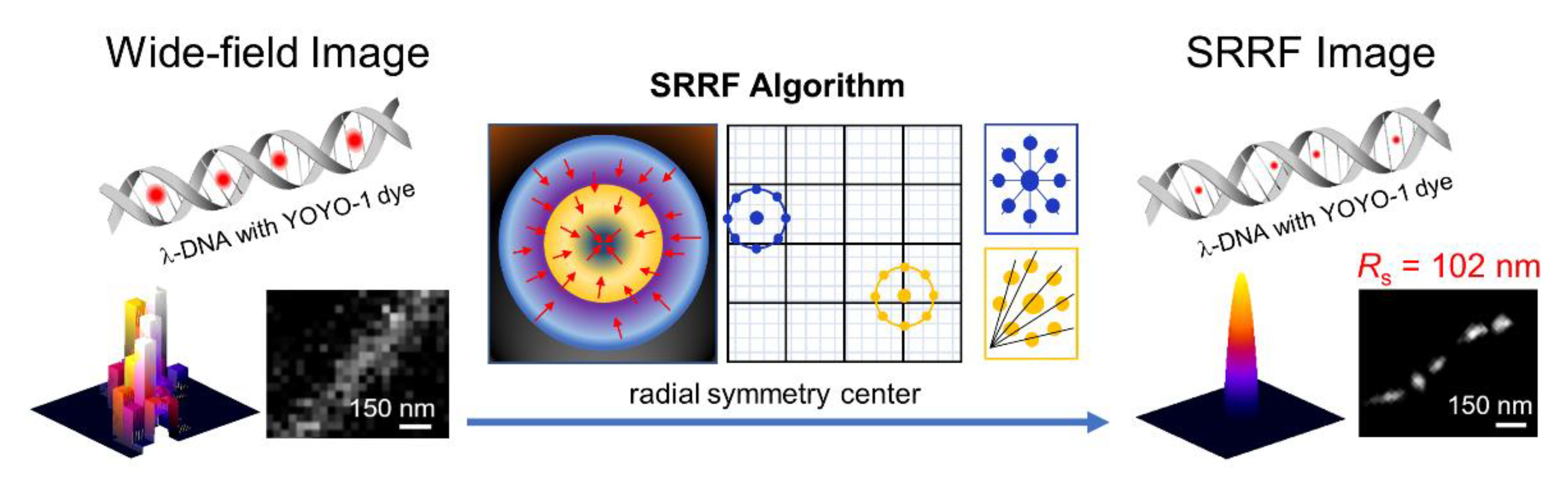{kind=link}
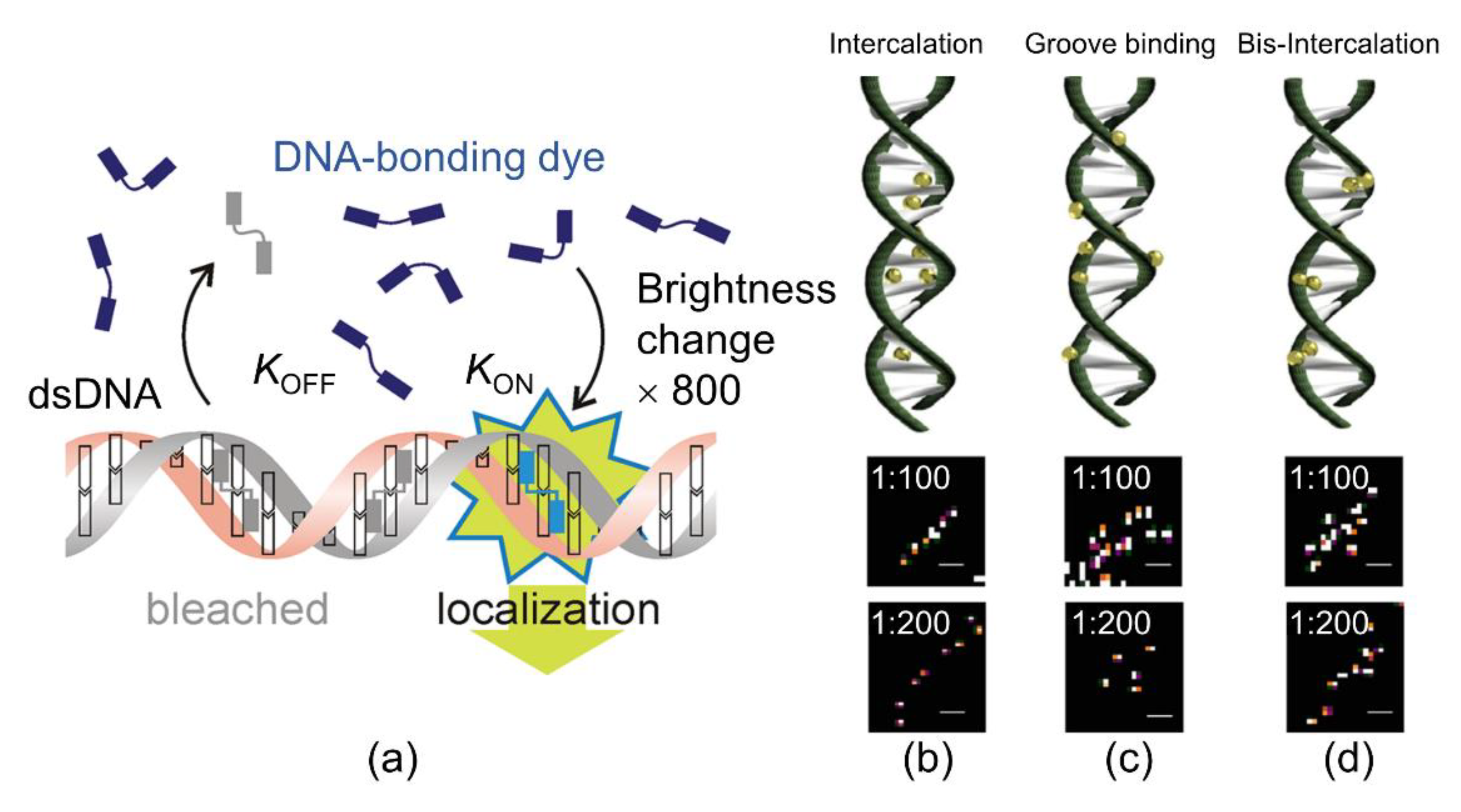{kind=link}
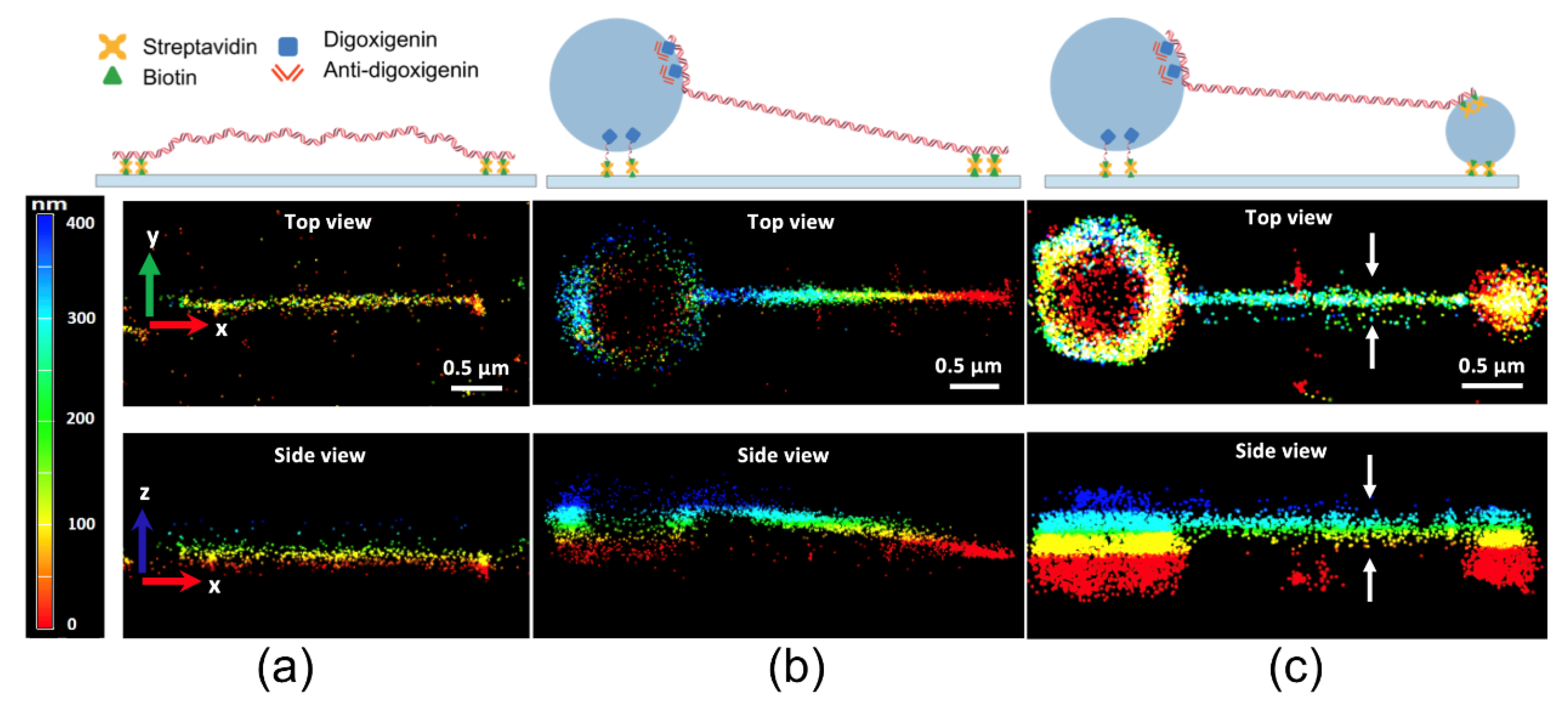{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
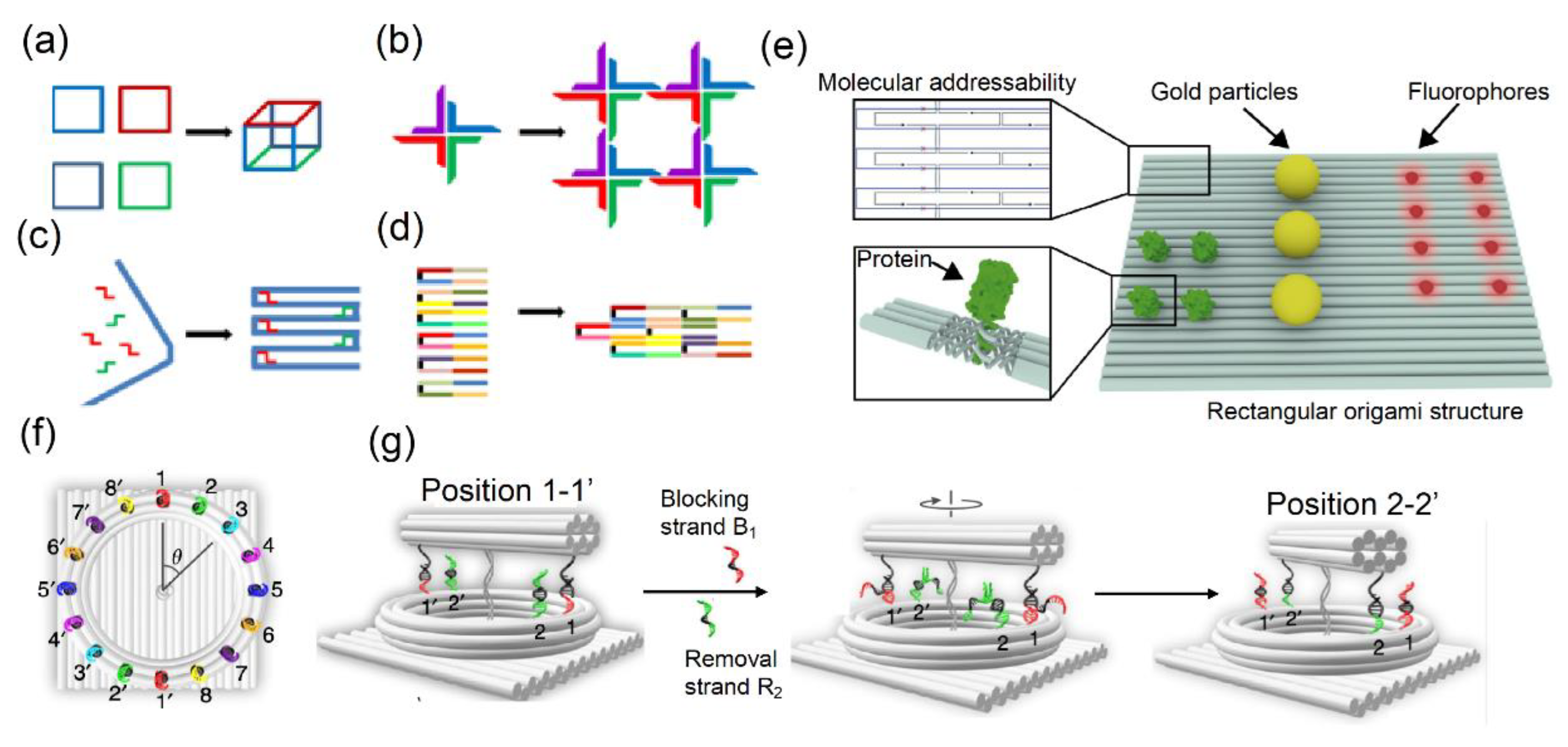
1. Introduction
2. Single-DNA Molecules as Targets in Stochastic Switching-Based Localization Microscopy
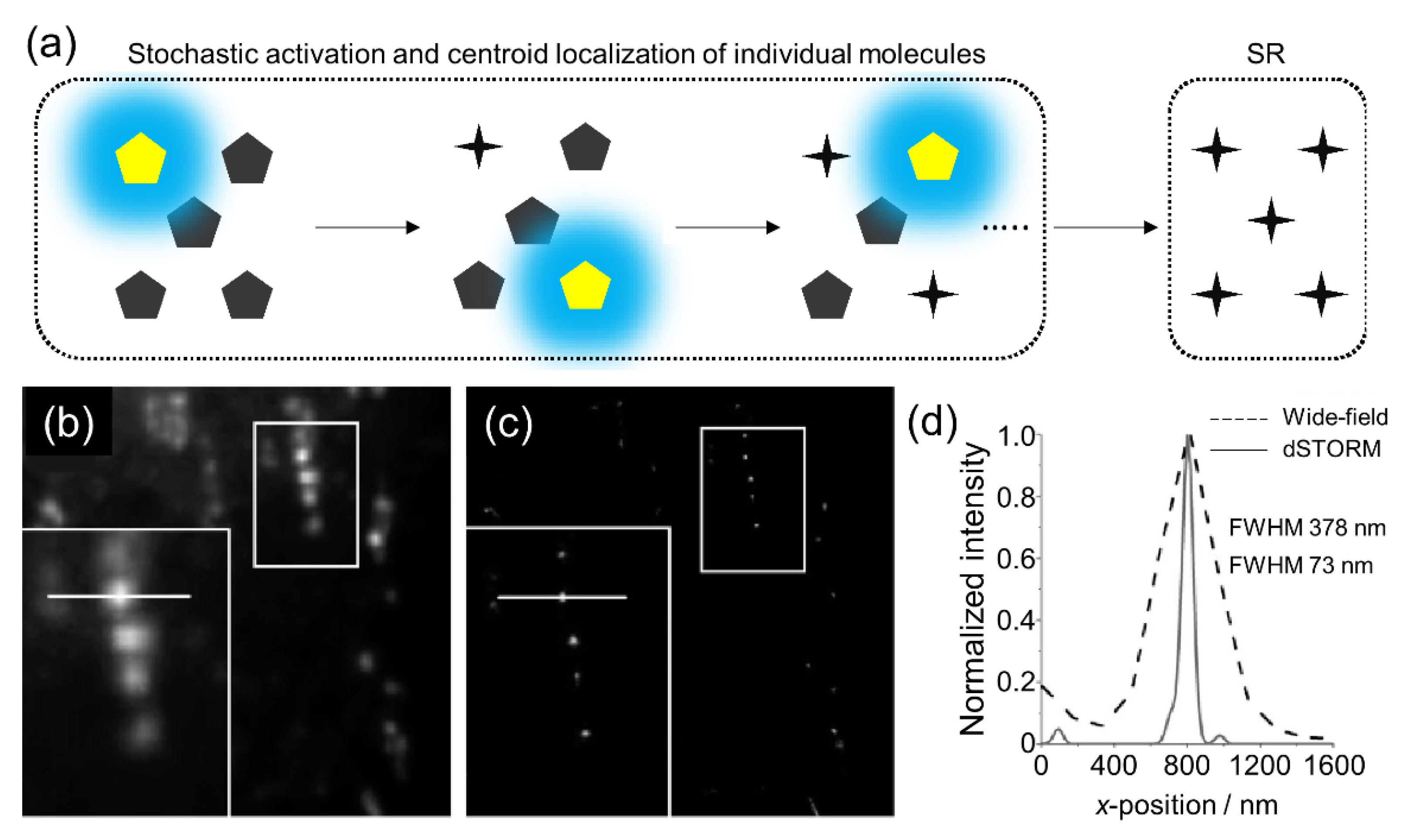
2.1. dSTORM
2.2. BALM
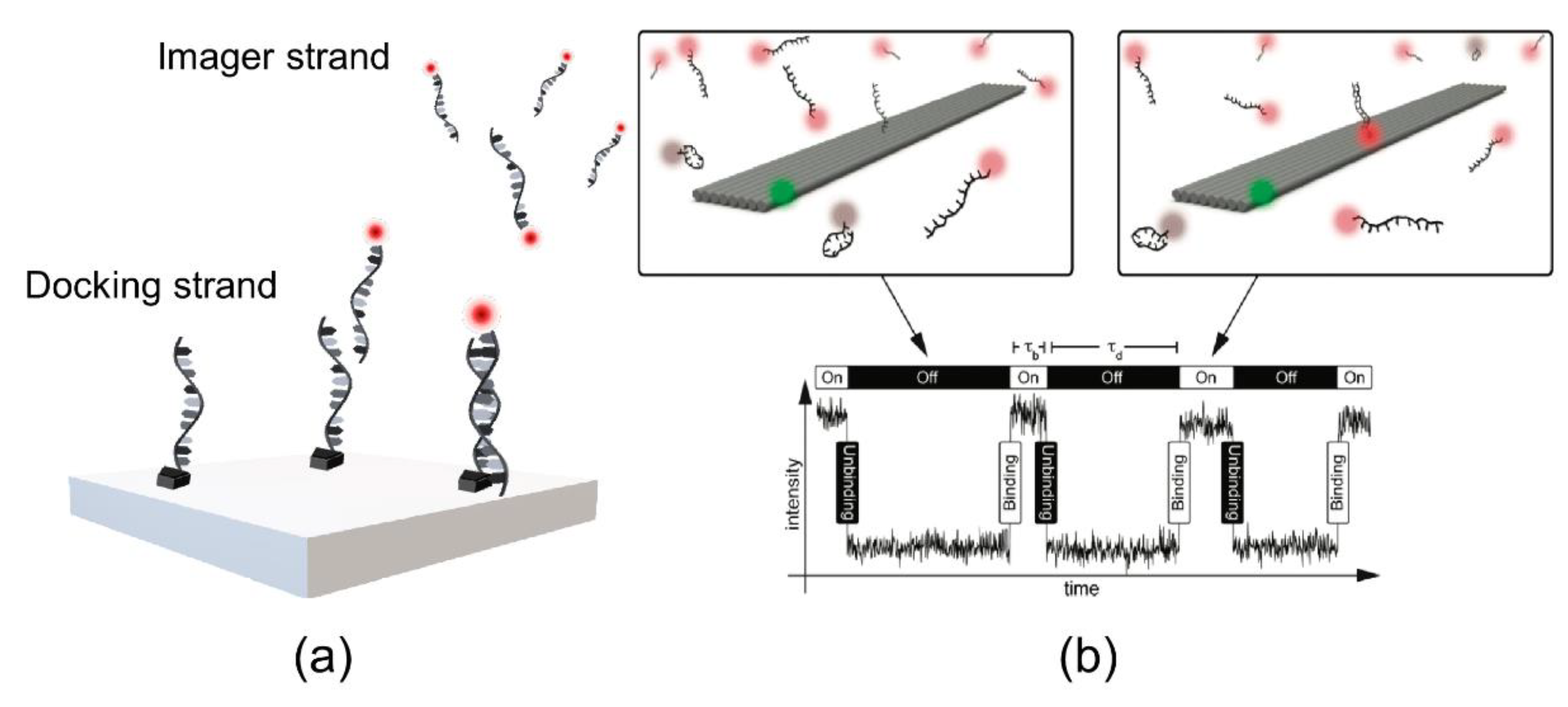
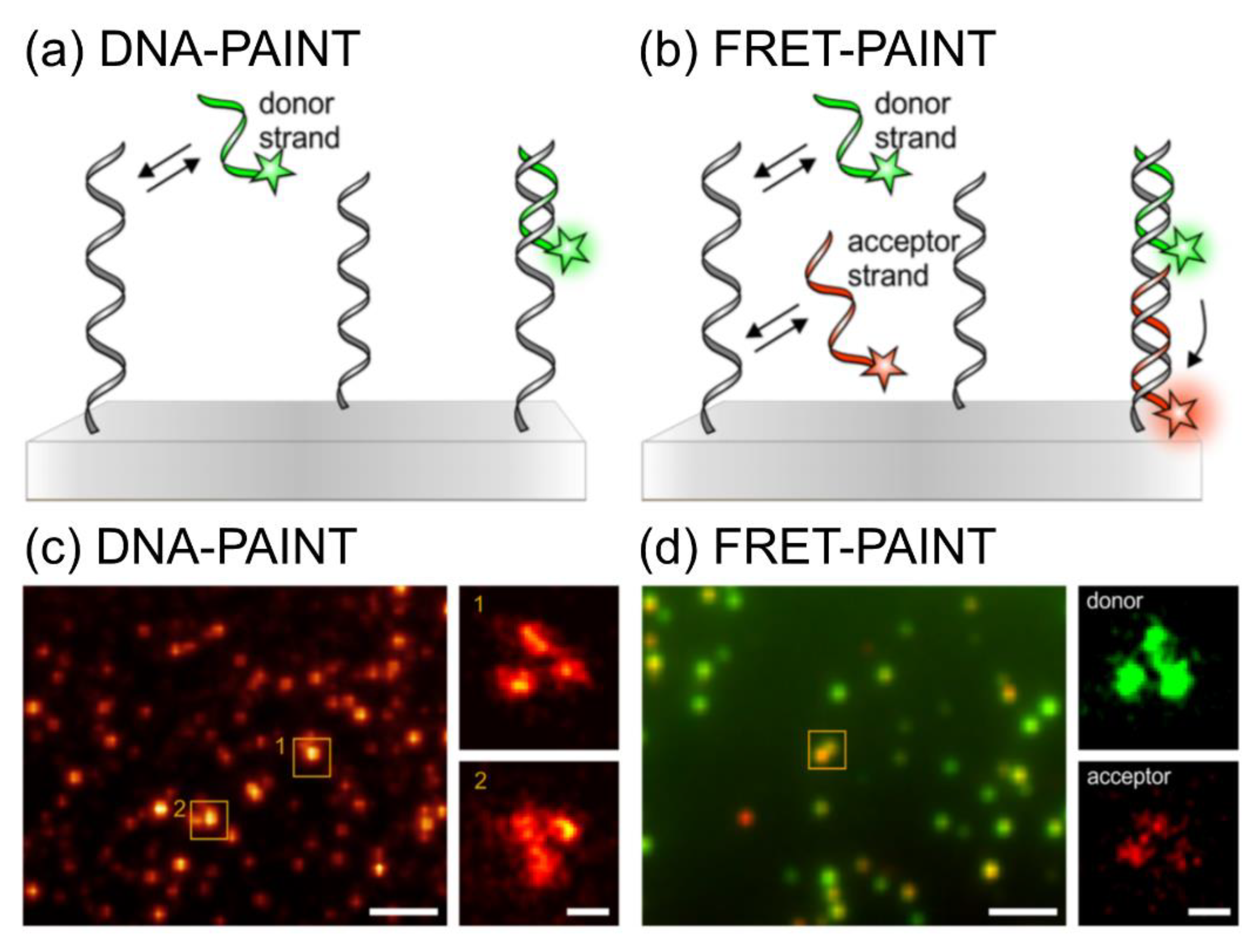
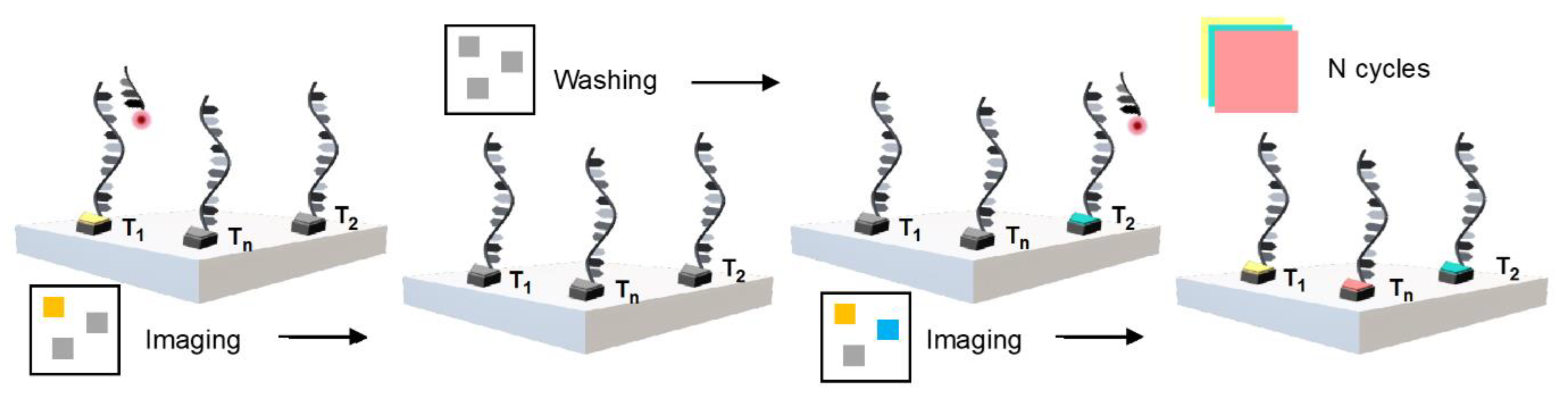
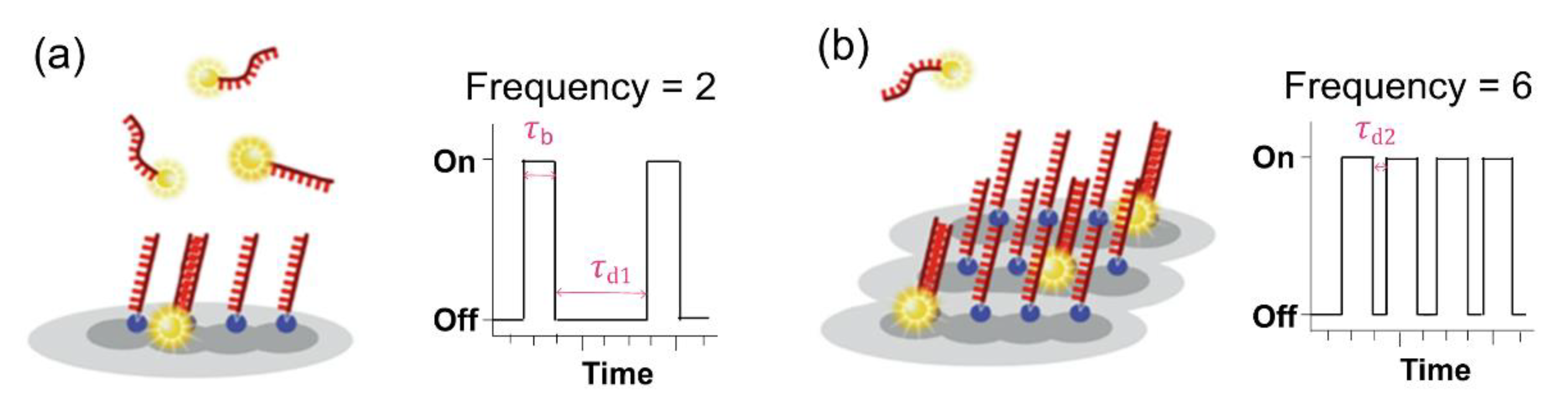
2.3. DNA-PAINT
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ebrahimi, A.; Ravan, H.; Khajouei, S. DNA nanotechnology and bioassay development. TrAC. Trend. Anal. Chem. 2019, 114, 126–142. [Google Scholar] [CrossRef]
- Nummelin, S.; Kommeri, J.; Kostiainen, M.A.; Linko, V. Evolution of structural DNA nanotechnology. Adv. Mater. 2018, 30, 1703721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Horn, J.D. DNA structure and supercoiling: Ribbons and a Yo-Yo model. J. Chem. Educ. 2011, 88, 1264–1267. [Google Scholar] [CrossRef]
- Lin, C.; Liu, Y.; Rinker, S.; Yan, H. DNA tile based self-assembly: Building complex nanoarchitectures. Chem. Phys. Chem. 2006, 7, 1641–1647. [Google Scholar] [CrossRef] [PubMed]
- Paukstelis, P.J.; Nowakowski, J.; Birktoft, J.J.; Seeman, N.C. Crystal structure of a continuous three-dimensional DNA lattice. Chem. Biol. 2004, 11, 1119–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, Y.; Ong, L.L.; Shih, W.M.; Yin, P. Three-dimensional structures self-assembled from DNA bricks. Science 2012, 338, 1177–1183. [Google Scholar] [CrossRef] [Green Version]
- Fu, T.J.; Seeman, N.C. DNA double-crossover molecules. Biochemistry 1993, 32, 3211–3220. [Google Scholar] [CrossRef]
- Ramezani, H.; Dietz, H. Building machines with DNA molecules. Nat. Rev. Genet. 2020, 21, 5–26. [Google Scholar] [CrossRef]
- Ke, Y.; Lindsay, S.; Chang, Y.; Liu, Y.; Yan, H. Self-assembled water-soluble nucleic acid probe tiles for label-free RNA hybridization assays. Science 2008, 319, 180–183. [Google Scholar] [CrossRef]
- Subramanian, H.K.; Chakraborty, B.; Sha, R.; Seeman, N.C. The label-free unambiguous detection and symbolic display of single nucleotide polymorphisms on DNA origami. Nano Lett. 2011, 11, 910–913. [Google Scholar] [CrossRef]
- Pan, J.; Cha, T.G.; Li, F.; Chen, H.; Bragg, N.A.; Choi, J.H. Visible/near-infrared subdiffraction imaging reveals the stochastic nature of DNA walkers. Sci. Adv. 2017, 3, e1601600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Jain, S.; Dilbaghi, N.; Ahluwalia, A.S.; Hassan, A.A.; Kim, K.H. Advanced selection methodologies for DNAzymes in sensing and healthcare applications. Trends Biochem. Sci. 2019, 44, 190–213. [Google Scholar] [CrossRef] [PubMed]
- Steinhauer, C.; Jungmann, R.; Sobey, T.L.; Simmel, F.C.; Tinnefeld, P. DNA origami as a nanoscopic ruler for super-resolution microscopy. Angew. Chem. Int. Ed. 2009, 48, 8870–8873. [Google Scholar] [CrossRef] [PubMed]
- Betzig, E.; Patterson, G.H.; Sougrat, R.; Lindwasser, O.W.; Olenych, S.; Bonifacino, J.S.; Davidson, M.W.; Lippincott-Schwartz, J.; Hess, H.F. Imaging intracellular fluorescent proteins at nanometer resolution. Science 2006, 313, 1642–1645. [Google Scholar] [CrossRef] [Green Version]
- Rust, M.J.; Bates, M.; Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy (STORM). Nat. Methods 2006, 3, 793–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heilemann, M.; Van De Linde, S.; Schüttpelz, M.; Kasper, R.; Seefeldt, B.; Mukherjee, A.; Tinnefeld, P.; Sauer, M. Subdiffraction-resolution fluorescence imaging with conventional fluorescent probes. Angew. Chem. Int. Ed. 2008, 47, 6172–6176. [Google Scholar] [CrossRef]
- Jungmann, R.; Steinhauer, C.; Scheible, M.; Kuzyk, A.; Tinnefeld, P.; Simmel, F.C. Single-molecule kinetics and super-resolution microscopy by fluorescence imaging of transient binding on DNA origami. Nano Lett. 2010, 10, 4756–4761. [Google Scholar] [CrossRef]
- Gradišar, H.; Jerala, R. Self-assembled bionanostructures: Proteins following the lead of DNA nanostructures. J. Nanobiotechnol. 2014, 12, 4. [Google Scholar] [CrossRef] [Green Version]
- Holliday, R. A mechanism for gene conversion in fungi. Genet. Res. 1964, 5, 282–304. [Google Scholar] [CrossRef] [Green Version]
- Ko, S.H.; Su, M.; Zhang, C.; Ribbe, A.E.; Jiang, W.; Mao, C. Synergistic self-assembly of RNA and DNA molecules. Nat. Chem. 2010, 2, 1050–1055. [Google Scholar] [CrossRef]
- He, Y.; Chen, Y.; Liu, H.; Ribbe, A.E.; Mao, C. Self-assembly of hexagonal DNA two-dimensional (2D) arrays. J. Am. Chem. Soc. 2005, 127, 12202–12203. [Google Scholar] [CrossRef] [PubMed]
- Xin, L.; Zhou, C.; Duan, X.; Liu, N. A rotary plasmonic nanoclock. Nat. Commun. 2019, 10, 5394. [Google Scholar] [CrossRef] [PubMed]
- Schlichthaerle, T.; Strauss, M.T.; Schueder, F.; Woehrstein, J.B.; Jungmann, R. DNA nanotechnology and fluorescence applications. Curr. Opin. Biotechnol. 2016, 39, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Jensen, E.; Crossman, D.J. Technical review: Types of imaging-direct STORM. Anat. Rec. 2014, 297, 2227–2231. [Google Scholar] [CrossRef] [PubMed]
- Benke, A.; Manley, S. Live-cell dSTORM of cellular DNA based on direct DNA labeling. Chem. Biol. Chem. 2011, 13, 298–301. [Google Scholar] [CrossRef] [Green Version]
- Flors, C.; Ravarani, C.N.; Dryden, D.T. Super-resolution imaging of DNA labelled with intercalating dyes. Chem. Phys. Chem. 2009, 10, 2201–2204. [Google Scholar] [CrossRef] [PubMed]
- Flors, C. Photoswitching of monomeric and dimeric DNA-intercalating cyanine dyes for super-resolution microscopy applications. Photochem. Photobiol. Sci. 2010, 9, 643–648. [Google Scholar] [CrossRef]
- Backer, A.S.; Lee, M.Y.; Moerner, W.E. Enhanced DNA imaging using super-resolution microscopy and simultaneous single-molecule orientation measurements. Optica 2016, 3, 659–666. [Google Scholar] [CrossRef] [Green Version]
- Gustafsson, N.; Culley, S.; Ashdown, G.; Owen, D.M.; Pereira, P.M.; Henriques, R. Fast live-cell conventional fluorophore nanoscopy with ImageJ through super-resolution radial fluctuations. Nat. Commun. 2016, 7, 12471. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.T.T.; Kang, S.H. Base pair distance in single-DNA molecule via TIRF-based super-resolution radial fluctuations-stream module. Bull. Korean Chem. Soc. 2020, 41, 476–479. [Google Scholar] [CrossRef]
- Schoen, I.; Ries, J.; Klotzsch, E.; Ewers, H.; Vogel, V. Binding-activated localization microscopy of DNA structures. Nano Lett. 2011, 11, 4008–4011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, G.; Chakkarapani, S.K.; Ju, S.; Ahn, S.; Kang, S.H. Super-resolution morphological dissemination of intercalating dye in single DNA molecules via binding activated localization microscopy. Chin. Chem. Lett. 2018, 29, 505–508. [Google Scholar] [CrossRef]
- Yardimci, S.; Burnham, D.R.; Terry, S.Y.A.; Yardimci, H. Three-dimensional super-resolution fluorescence imaging of DNA. Sci. Rep. 2020, 10, 12504. [Google Scholar] [CrossRef] [PubMed]
- Meijering, A.E.C.; Biebricher, A.S.; Sitters, G.; Brouwer, I.; Peterman, E.J.G.; Wuite, G.J.L.; Heller, I. Imaging unlabeled proteins on DNA with super-resolution. Nucleic Acids Res. 2020, 48, e34. [Google Scholar] [CrossRef] [PubMed]
- Sharonov, A.; Hochstrasser, R.M. Wide-field subdiffraction imaging by accumulated binding of diffusing probes. Proc. Nat. Acad. Sci. USA 2006, 103, 18911–18916. [Google Scholar] [CrossRef] [Green Version]
- Nieves, D.J.; Gaus, K.; Baker, M.A.B. DNA-based super-resolution microscopy: DNA-PAINT. Genes 2018, 9, 621. [Google Scholar] [CrossRef] [Green Version]
- Schueder, F.; Stein, J.; Stehr, F.; Auer, A.; Sperl, B.; Strauss, M.T.; Schwille, P.; Jungmann, R. An order of magnitude faster DNA-PAINT imaging by optimized sequence design and buffer conditions. Nat. Methods 2019, 16, 1101–1104. [Google Scholar] [CrossRef]
- Raab, M.; Schmied, J.J.; Jusuk, I.; Forthmann, C.; Tinnefeld, P. Fluorescence microscopy with 6 nm resolution on DNA origami. Chem. Phys. Chem. 2014, 15, 2431–2435. [Google Scholar] [CrossRef]
- Raab, M.; Vietz, C.; Stefani, F.D.; Acuna, G.P.; Tinnefeld, P. Shifting molecular localization by plasmonic coupling in a single-molecule mirage. Nat. Commun. 2017, 8, 13966. [Google Scholar] [CrossRef] [Green Version]
- Stehr, F.; Stein, J.; Schueder, F.; Schwille, P.; Jungmann, R. Flat-top TIRF illumination boosts DNA-PAINT imaging and quantification. Nat. Commun. 2019, 10, 1268. [Google Scholar] [CrossRef]
- Deussner-Helfmann, N.S.; Auer, A.; Strauss, M.T.; Donlin-Asp, P.; Malkusch, S.; Dietz, M.S.; Barth, H.D.; Schuman, E.; Jungmann, R.; Heilemann, M. Correlating DNA-PAINT and single-molecule FRET for multiplexed super-resolution imaging. SPIE 2020, 11246, 112460L. [Google Scholar]
- Deußner-Helfmann, N.S.; Auer, A.; Strauss, M.T.; Malkusch, S.; Dietz, M.S.; Barth, H.D.; Jungmann, R.; Heilemann, M. Correlative single-molecule FRET and DNA-PAINT imaging. Nano Lett. 2018, 18, 4626–4630. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Park, S.; Hohng, S. Accelerated FRET-PAINT microscopy. Mol. Brain 2018, 11, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jungmann, R.; Avendaño, M.S.; Woehrstein, J.B.; Dai, M.; Shih, W.M.; Yin, P. Multiplexed 3D cellular super-resolution imaging with DNA-PAINT and Exchange-PAINT. Nat. Methods 2014, 11, 313–318. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Woehrstein, J.B.; Donoghue, N.; Dai, M.; Avendaño, M.S.; Schackmann, R.C.; Zoeller, J.J.; Wang, S.S.H.; Tillberg, P.W.; Park, D.; et al. Rapid sequential in situ multiplexing with DNA exchange imaging in neuronal cells and tissues. Nano Lett. 2017, 17, 6131–6139. [Google Scholar] [CrossRef] [Green Version]
- Schueder, F.; Strauss, M.T.; Hoerl, D.; Schnitzbauer, J.; Schlichthaerle, T.; Strauss, S.; Yin, P.; Harz, H.; Leonhardt, H.; Jungmann, R. Universal super-resolution multiplexing by DNA exchange. Angew. Chem. Int. Ed. 2017, 56, 4052–4055. [Google Scholar] [CrossRef] [Green Version]
- Agasti, S.S.; Wang, Y.; Schueder, F.; Sukumar, A.; Jungmann, R.; Yin, P. DNA-barcoded labeling probes for highly multiplexed Exchange-PAINT imaging. Chem. Sci. 2017, 8, 3080–3091. [Google Scholar] [CrossRef] [Green Version]
- Dai, M.; Jungmann, R.; Yin, P. Optical imaging of individual biomolecules in densely packed clusters. Nat. Nanotechnol. 2016, 11, 798–807. [Google Scholar] [CrossRef] [Green Version]
- Werbin, J.L.; Avendaño, M.S.; Becker, V.; Jungmann, R.; Yin, P.; Danuser, G.; Sorger, P.K. Multiplexed exchange-PAINT imaging reveals ligand-dependent EGFR and Met interactions in the plasma membrane. Sci. Rep. 2017, 7, 12150. [Google Scholar] [CrossRef]
- Jungmann, R.; Avendaño, M.S.; Dai, M.; Woehrstein, J.; Agasti, S.S.; Feiger, Z.; Rodal, A.; Yin, P. Quantitative super-resolution imaging with qPAINT. Nat. Methods 2016, 13, 439–442. [Google Scholar] [CrossRef]
- Lin, R.; Clowsley, A.H.; Lutz, T.; Baddeley, D.; Soeller, C. 3D super-resolution microscopy performance and quantitative analysis assessment using DNA-PAINT and DNA origami test samples. Methods 2020, 174, 56–71. [Google Scholar] [CrossRef] [PubMed]
- Delcanale, P.; Miret-Ontiveros, B.; Arista-Romero, M.; Pujals, S.; Albertazzi, L. Nanoscale mapping functional sites on nanoparticles by points accumulation for imaging in nanoscale topography (PAINT). ACS Nano 2018, 12, 7629–7637. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.A.B.; Nieves, D.J.; Hilzenrat, G.; Berengut, J.F.; Gaus, K.; Lee, L.K. Stoichiometric quantification of spatially dense assemblies with qPAINT. Nanoscale 2019, 11, 12460–12464. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Liu, Y. Super-resolution localization microscopy: Toward high throughput, high quality, and low cost. APL Photonics 2020, 5, 060902. [Google Scholar] [CrossRef]
- Xu, K.; Babcock, H.P.; Zhuang, X. Dual-objective STORM reveals three-dimensional filament organization in the actin cytoskeleton. Nat. Methods 2012, 9, 185–188. [Google Scholar] [CrossRef]
- Filius, M.; Cui, T.J.; Ananth, A.N.; Docter, M.W.; Hegge, J.W.; Van Der Oost, J.; Joo, C. High-speed super-resolution imaging using protein-assisted DNA-PAINT. Nano Lett. 2020, 20, 2264–2270. [Google Scholar] [CrossRef]
- Auer, A.; Schlichthaerle, T.; Woehrstein, J.B.; Schueder, F.; Strauss, M.T.; Grabmayr, H.; Jungmann, R. Nanometer-scale multiplexed super-resolution imaging with an economic 3D-DNA-PAINT microscope. Chem. Phys. Chem. 2018, 19, 3024–3034. [Google Scholar] [CrossRef]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Batjikh, I.; Kang, S.H. Toward Sub-Diffraction Imaging of Single-DNA Molecule Sensors Based on Stochastic Switching Localization Microscopy. Sensors 2020, 20, 6667. https://doi.org/10.3390/s20226667
Lee S, Batjikh I, Kang SH. Toward Sub-Diffraction Imaging of Single-DNA Molecule Sensors Based on Stochastic Switching Localization Microscopy. Sensors. 2020; 20(22):6667. https://doi.org/10.3390/s20226667
Chicago/Turabian StyleLee, Seungah, Indra Batjikh, and Seong Ho Kang. 2020. "Toward Sub-Diffraction Imaging of Single-DNA Molecule Sensors Based on Stochastic Switching Localization Microscopy" Sensors 20, no. 22: 6667. https://doi.org/10.3390/s20226667
APA StyleLee, S., Batjikh, I., & Kang, S. H. (2020). Toward Sub-Diffraction Imaging of Single-DNA Molecule Sensors Based on Stochastic Switching Localization Microscopy. Sensors, 20(22), 6667. https://doi.org/10.3390/s20226667