Timing and Modulation of Activity in the Lower Limb Muscles During Indoor Rowing: What Are the Key Muscles to Target in FES-Rowing Protocols?

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Participants
2.2. Experimental Protocol
2.3. EMG and Rowing Machine Recordings
2.4. Estimating the Timing and Side-Differences in the Modulation of Muscle Activity
2.5. Statistics
3. Results
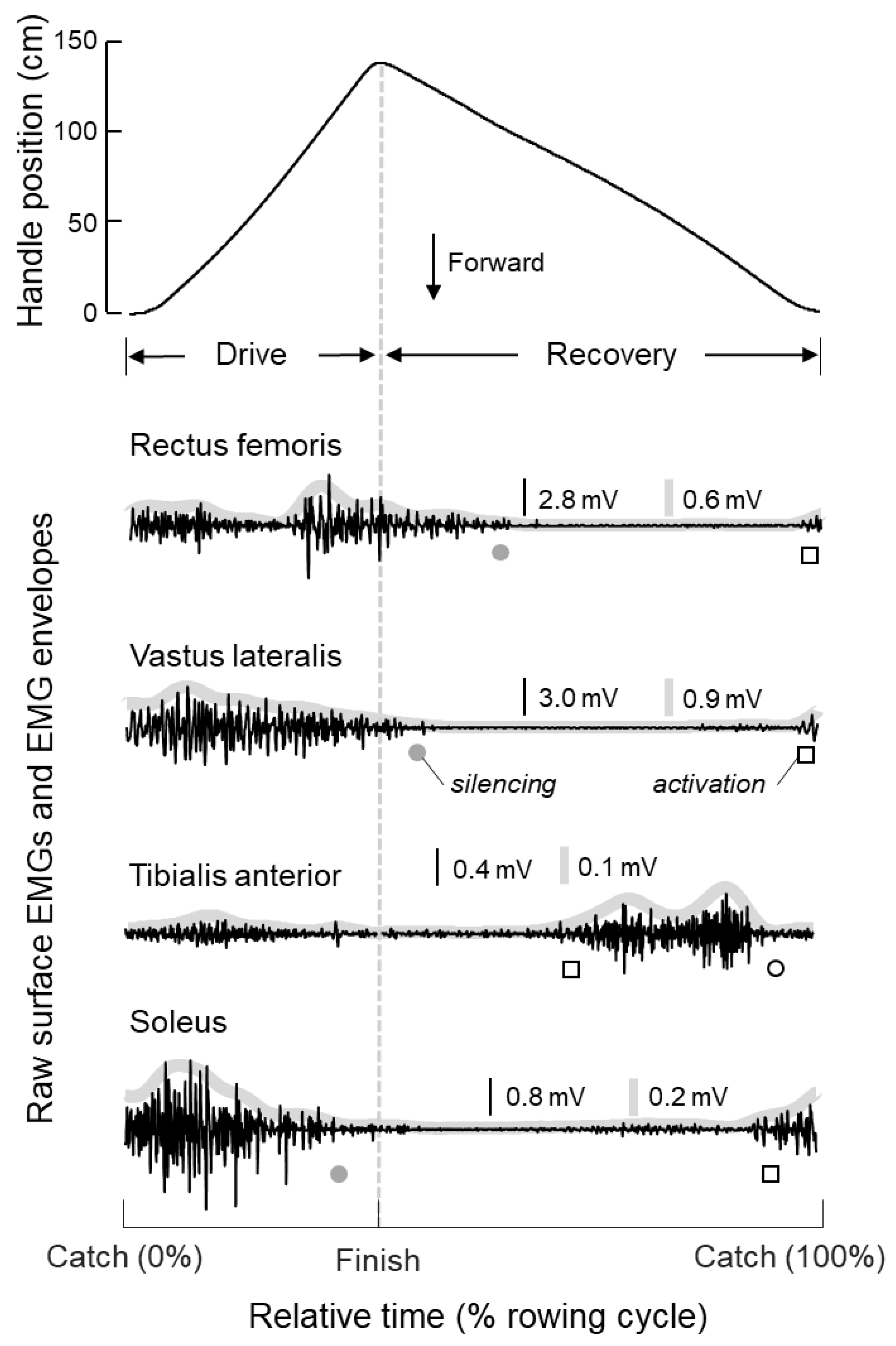
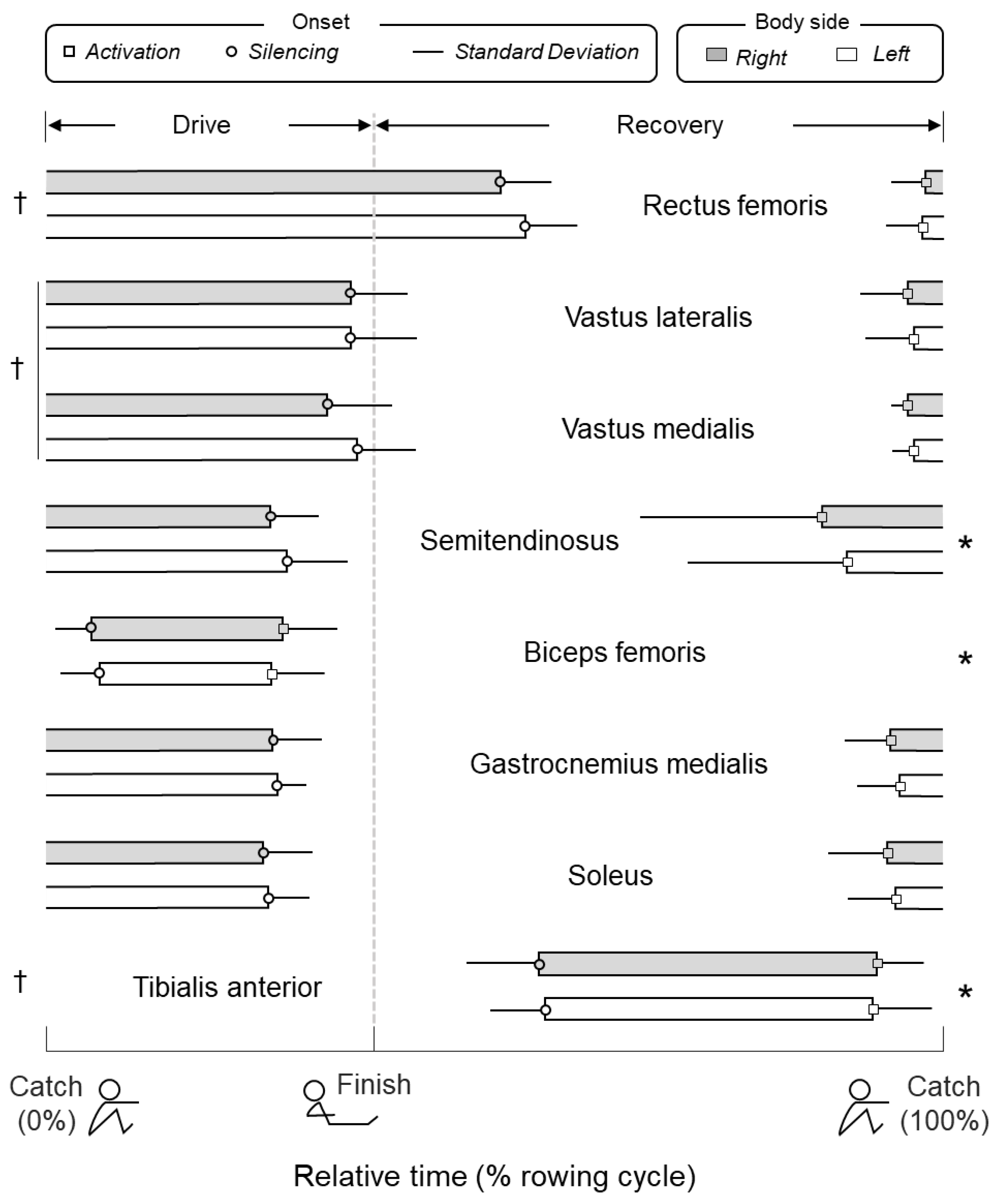
3.1. Timing and Duration of Muscle Activity During Rowing
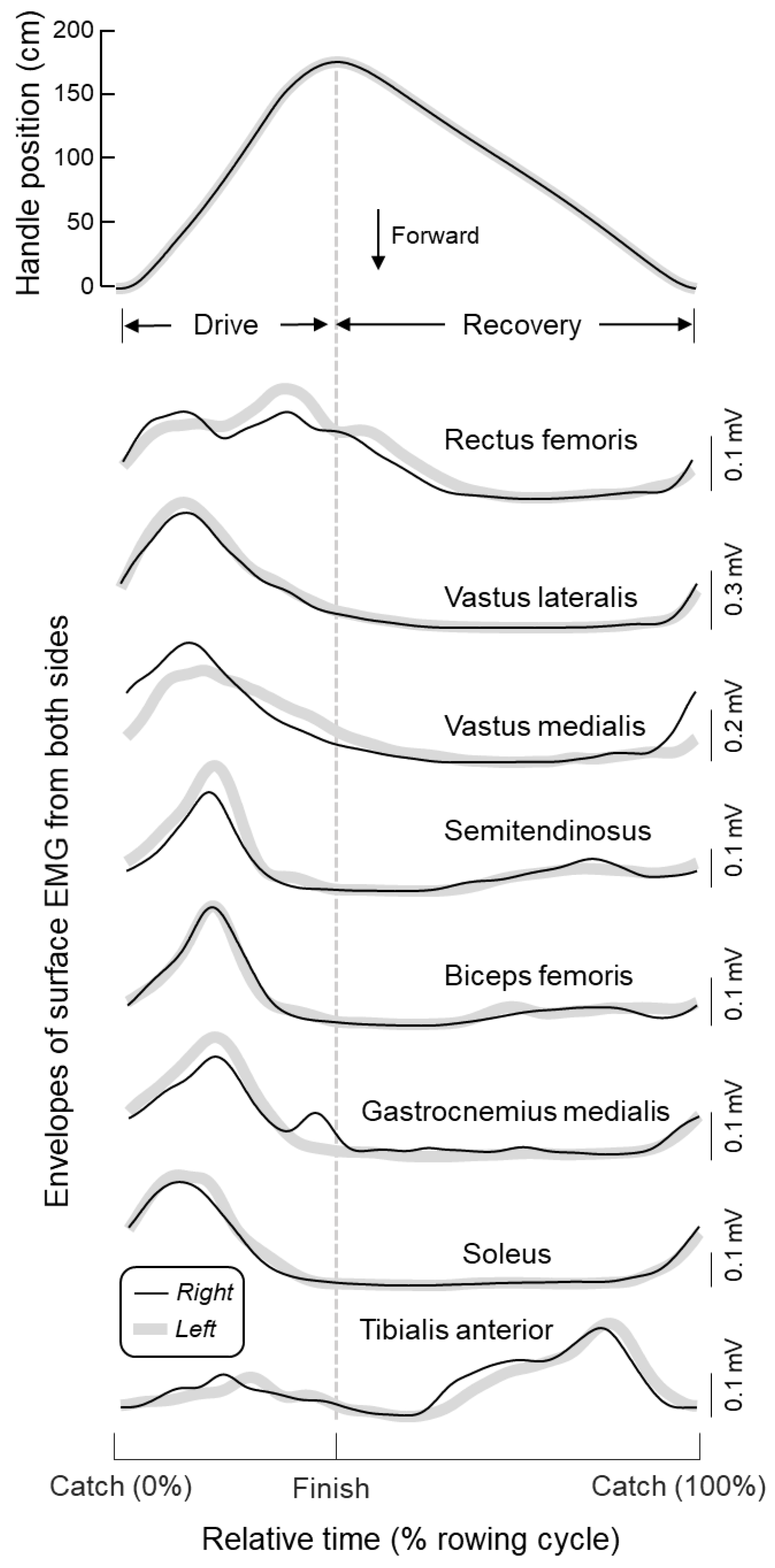
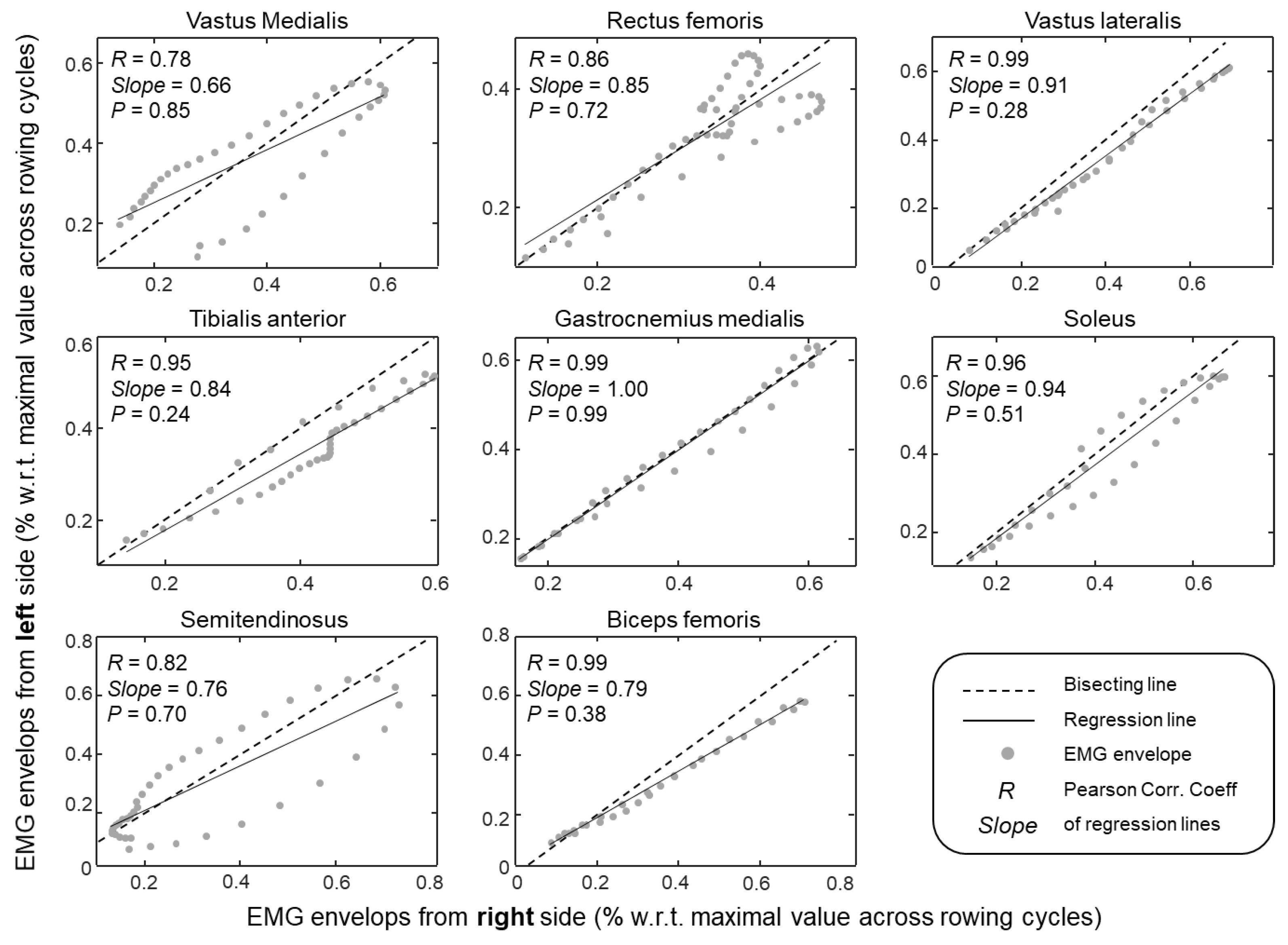
3.2. Modulation of Muscle Activity in Both Legs
4. Discussion
4.1. What Are the Key, Leg Muscles Recruited During Indoor Rowing?
4.2. Identification of Key Leg Muscles to Target in FES-Rowing Applications
4.3. A Few Considerations on the Validity of Current Results
Author Contributions
Acknowledgments
Conflicts of Interest
Ethical Statements
References
- Laskin, J.J.; Ashley, E.A.; Olenik, L.M.; Burnham, R.; Cumming, D.C.; Steadward, R.D.; Wheeler, G.D. Electrical stimulation-assisted rowing exercise in spinal cord injured people. A pilot study. Paraplegia 1993, 31, 534–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheeler, G.D.; Andrews, B.; Lederer, R.; Davoodi, R.; Natho, K.; Weiss, C.; Jeon, J.; Bhambhani, Y.; Steadward, R.D. Functional electric stimulation-assisted rowing: Increasing cardiovascular fitness through functional electric stimulation rowing training in persons with spinal cord injury. Arch. Phys. Med. Rehabil. 2002, 83, 1093–1099. [Google Scholar] [CrossRef]
- Taylor, J.A.; Picard, G.; Widrick, J.J. Aerobic Capacity With Hybrid FES Rowing in Spinal Cord Injury: Comparison With Arms-Only Exercise and Preliminary Findings With Regular Training. PM R 2011, 3, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, R.S.; McCarthy, I.D.; Gall, A.; Stock, C.G.; Shippen, J.; Andrews, B.J. Can FES-rowing mediate bone mineral density in SCI: A pilot study. Spinal Cord. 2014, 52, S4–S5. [Google Scholar] [CrossRef] [PubMed]
- Hettinga, D.M.; Andrews, B.J. The feasibility of functional electrical stimulation indoor rowing for high-energy training and sport. Neuromodulation 2007, 10, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Lambach, R.L.; Stafford, N.E.; Kolesar, J.A.; Kiratli, B.J.; Creasey, G.H.; Gibbons, R.S.; Andrews, B.J.; Beaupre, G.S. Bone changes in the lower limbs from participation in an FES rowing exercise program implemented within two years after traumatic spinal cord injury. J. Spinal Cord Med. 2018, 1–9. [Google Scholar] [CrossRef]
- Draghici, A.E.; Picard, G.; Taylor, J.A.; Shefelbine, S.J. Assessing kinematics and kinetics of functional electrical stimulation rowing. J. Biomech. 2017, 53, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Halliday, S.E.; Zavatsky, A.B.; Hase, K. Can functional electric stimulation-assisted rowing reproduce a race-winning rowing stroke? Arch. Phys. Med. Rehabil. 2004, 85, 1265–1272. [Google Scholar] [CrossRef]
- Shaharudin, S.; Zanotto, D.; Agrawal, S. Muscle Synergy during Wingate Anaerobic Rowing Test of Collegiate Rowers and Untrained Subjects. Int. J. Sport. Sci. 2014, 4, 165–172. [Google Scholar]
- Turpin, N.A.; Gu??vel, A.; Durand, S.; Hug, F. No evidence of expertise-related changes in muscle synergies during rowing. J. Electromyogr. Kinesiol. 2011, 21, 1030–1040. [Google Scholar] [CrossRef]
- Turpin, N.A.; Guével, A.; Durand, S.; Hug, F. Effect of power output on muscle coordination during rowing. Eur. J. Appl. Physiol. 2011, 111, 3017–3029. [Google Scholar] [CrossRef] [PubMed]
- Parkin, S.; Nowicky, A.V.; Rutherford, O.M.; McGregor, A.H. Do oarsmen have asymmetries in the strength of their back and leg muscles? J. Sports Sci. 2001, 19, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.-M.J.; Robertson, D.G.E.; Stothart, J.P. Analysis of Lower Limb Muscle Function in Ergometer Rowing. Int. J. Sport Biomech. 1988, 4, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Pollock, C.L.; Jenkyn, T.R.; Jones, I.C.; Ivanova, T.D.; Garland, S.J. Electromyography and kinematics of the trunk during rowing in elite female rowers. Med. Sci. Sports Exerc. 2009, 41, 628–636. [Google Scholar] [CrossRef]
- Janshen, L.; Mattes, K.; Tidow, G. Muscular coordination of the lower extremities of oarsmen during ergometer rowing. J. Appl. Biomech. 2009, 25, 156–164. [Google Scholar] [CrossRef]
- So, R.C.H.; Tse, M.A.; Wong, S.C.W. Application of Surface Electromyography in Assesing Muscle Recruitment Patterns in Six-Minute Continuous Rowing Effort. J. Strenght Cond. Res. 2007, 21, 724–730. [Google Scholar]
- Readi, N.G.G.; Rosso, V.; Rainoldi, A.; Vieira, T.M.M.M.M. Do sweep rowers symmetrically activate their low back muscles during indoor rowing? Scand. J. Med. Sci. Sports 2015, 25, e339–e352. [Google Scholar] [CrossRef]
- Fohanno, V.; Nordez, A.; Smith, R.; Colloud, F. Asymmetry in elite rowers: Effect of ergometer design and stroke rate. Sport. Biomech. 2015, 14, 310–322. [Google Scholar] [CrossRef]
- Buckeridge, E.M.; Bull, A.M.J.J.; McGregor, A.H. Foot force production and asymmetries in elite rowers. Sports Biomech. 2014, 13, 47–61. [Google Scholar] [CrossRef]
- Guével, A.; Boyas, S.; Guihard, V.; Cornu, C.; Hug, F.; Nordez, A. Thigh muscle activities in elite rowers during on-water rowing. Int. J. Sports Med. 2011, 32, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Andrews, B.J.; Gibbons, R.; Goodey, S.; Poulton, A.; Shippen, J. Towards the Development of Full Motion FES Rowing with Accurate Ergometry: RowStim IV. In Converging Clinical and Engineering Research on Neurorehabilitation III; Masia, L., Micera, S., Akay, M., Pons, J., Eds.; Springer: Cham, Switzerland, 2019; pp. 917–921. [Google Scholar]
- Davoodi, R.; Andrews, B.J.; Wheeler, G.D. Automatic finite state control of FES-assisted indoor rowing exercise after spinal cord injury. Neuromodulation 2002, 5, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Hermens, H.J.; Freriks, B.; Disselhorst-Klug, C.; Rau, G. Development of recommendations for SEMG sensors and sensor placement procedures. J. Electromyogr. Kinesiol. 2000, 10, 361–374. [Google Scholar] [CrossRef]
- Marateb, H.R.; McGill, K.C.; Holobar, A.; Lateva, Z.C.; Mansourian, M.; Merletti, R. Accuracy assessment of CKC high-density surface EMG decomposition in biceps femoris muscle. J. Neural Eng. 2011, 8, 066002. [Google Scholar] [CrossRef] [PubMed]
- Vinti, M.; Gracies, J.M.; Gazzoni, M.; Vieira, T. Localised sampling of myoelectric activity may provide biased estimates of cocontraction for gastrocnemius though not for soleus and tibialis anterior muscles. J. Electromyogr. Kinesiol. 2018, 38, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Cabral, H.V.H.V.; de Souza, L.M.L.L.M.L.; Mello, R.G.T.R.G.T.; Gallina, A.; de Oliveira, L.F.L.F.; Vieira, T.M.T.M. Is the firing rate of motor units in different vastus medialis regions modulated similarly during isometric contractions? Muscle and Nerve 2018, 57, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Vieira, T.M.; Botter, A.; Muceli, S.; Farina, D. Specificity of surface EMG recordings for gastrocnemius during upright standing. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, T.; Cerone, G.L.; Gastaldi, L.; Pastorelli, S.; Oliveira, L.F.; Gazzoni, M.; Botter, A. Design and Test of a Biomechanical Model for the Estimation of Knee Joint Angle During Indoor Rowing: Implications for FES-Rowing Protocols in Paraplegia. IEEE Trans. Neural Syst. Rehabil. Eng. 2018, 26, 2145–2152. [Google Scholar] [CrossRef]
- Allison, G.T. Trunk muscle onset detection technique for EMG signals with ECG artefact. J. Electromyogr. Kinesiol. 2003, 13, 209–216. [Google Scholar] [CrossRef]
- Dawson, B.; Trapp, R.G. Basic & Clinical Biostatistics, 4th ed.; Lange Medical Books-McGraw-Hill, Medical Pub. Division, c2004: New York, NY, USA, 2004. [Google Scholar]
- Vigotsky, A.D.; Halperin, I.; Lehman, G.J.; Trajano, G.S.; Vieira, T.M. Interpreting signal amplitudes in surface electromyography studies in sport and rehabilitation sciences. Front. Physiol. 2018, 8, 985. [Google Scholar] [CrossRef] [Green Version]
- Kuo, A.D. The action of two-joint muscles: The legacy of W. P. Lombard. In Classics in Movement Science; Latash, M.L., Zatsiorsky, V.M., Eds.; Human Kinetics: Champaign, IL, USA, 2001; p. 451. [Google Scholar]
- Lombard, W.P. The Action of Two-Joint Muscles. Am. Phys. Educ. Rev. 1903, 8, 141–145. [Google Scholar] [CrossRef]
- Kirkwood, C.A.; Andrews, B.J.; Mowforth, P. Automatic detection of gait events: A case study using inductive learning techniques. J. Biomed. Eng. 1989, 11, 511–516. [Google Scholar] [CrossRef]
- Kostov, A.; Andrews, B.J.; Popović, D.B.; Stein, R.B.; Armstrong, W.W. Machine learning in control of functional electrical stimulation systems for locomotion. IEEE Trans. Biomed. Eng. 1995, 42, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Davoodi, R.; Andrews, B.J.; Wheeler, G.D.; Lederer, R. Development of an indoor rowing machine with manual FES controller for total body exercise in paraplegia. IEEE Trans. Neural Syst. Rehabil. Eng. 2002, 10, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Davoodi, R.; Andrews, B.J. Fuzzy Logic Control of FES Rowing Exercise in Paraplegia. IEEE Trans. Biomed. Eng. 2004, 51, 541–543. [Google Scholar] [CrossRef]
- Andrews, B.; Gibbons, R.; Wheeler, G. Development of Functional Electrical Stimulation Rowing: The Rowstim Series. Artif. Organs 2017, 41, E203–E212. [Google Scholar] [CrossRef]
- Morbidoni, C.; Cucchiarelli, A.; Fioretti, S.; Di Nardo, F. A deep learning approach to EMG-based classification of gait phases during level ground walking. Electronics 2019, 8, 894. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.-L.; Byun, H.-J.; Song, W.-G.; Son, J.-W.; Lim, J.-T. On pattern classification of EMG signals for walking motions. Artif. Life Robot. 2000, 4, 193–197. [Google Scholar] [CrossRef]
- Arnin, J.; Yamsa-ard, T.; Triponyuwasin, P.; Wongsawat, Y. Development of practical functional electrical stimulation cycling systems based on an electromyography study of the Cybathlon 2016. Eur. J. Transl. Myol. 2017, 27, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Heller, B.W.; Veltink, P.H.; Rijkhoff, N.J.M.; Rutten, W.L.C.; Andrews, B.J. Reconstructing muscle activation during normal walking: A comparison of symbolic and connectionist machine learning techniques. Biol. Cybern. 1993, 69, 327–335. [Google Scholar] [CrossRef]
- Thrasher, A.; Andrews, B.; Wang, F. Control of FES using reinforcement learning: Accelerating the learning rate. In Proceedings of the 19th Annual International Conference of the IEEE Engineering in Medicine and Biology Society. “Magnificent Milestones and Emerging Opportunities in Medical Engineering” (Cat. No.97CH36136), Chicago, IL, USA, 30 October–2 November 1997; pp. 1774–1776. [Google Scholar]
- Černe, T.; Kamnik, R.; Vesnicer, B.; Žganec Gros, J.; Munih, M. Differences between elite, junior and non-rowers in kinematic and kinetic parameters during ergometer rowing. Hum. Mov. Sci. 2013, 32, 691–707. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieira, T.M.; Cerone, G.L.; Stocchi, C.; Lalli, M.; Andrews, B.; Gazzoni, M. Timing and Modulation of Activity in the Lower Limb Muscles During Indoor Rowing: What Are the Key Muscles to Target in FES-Rowing Protocols? Sensors 2020, 20, 1666. https://doi.org/10.3390/s20061666
Vieira TM, Cerone GL, Stocchi C, Lalli M, Andrews B, Gazzoni M. Timing and Modulation of Activity in the Lower Limb Muscles During Indoor Rowing: What Are the Key Muscles to Target in FES-Rowing Protocols? Sensors. 2020; 20(6):1666. https://doi.org/10.3390/s20061666
Chicago/Turabian StyleVieira, Taian M., Giacinto Luigi Cerone, Costanza Stocchi, Morgana Lalli, Brian Andrews, and Marco Gazzoni. 2020. "Timing and Modulation of Activity in the Lower Limb Muscles During Indoor Rowing: What Are the Key Muscles to Target in FES-Rowing Protocols?" Sensors 20, no. 6: 1666. https://doi.org/10.3390/s20061666
APA StyleVieira, T. M., Cerone, G. L., Stocchi, C., Lalli, M., Andrews, B., & Gazzoni, M. (2020). Timing and Modulation of Activity in the Lower Limb Muscles During Indoor Rowing: What Are the Key Muscles to Target in FES-Rowing Protocols? Sensors, 20(6), 1666. https://doi.org/10.3390/s20061666