
Early Detection of Aphid Infestation and Insect-Plant Interaction Assessment in Wheat Using a Low-Cost Electronic Nose (E-Nose), Near-Infrared Spectroscopy and Machine Learning Modeling





Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and Insect Material, and Experimental Design Description
2.2. Physiological Measurements
2.3. Near-Infrared Spectroscopy Measurements
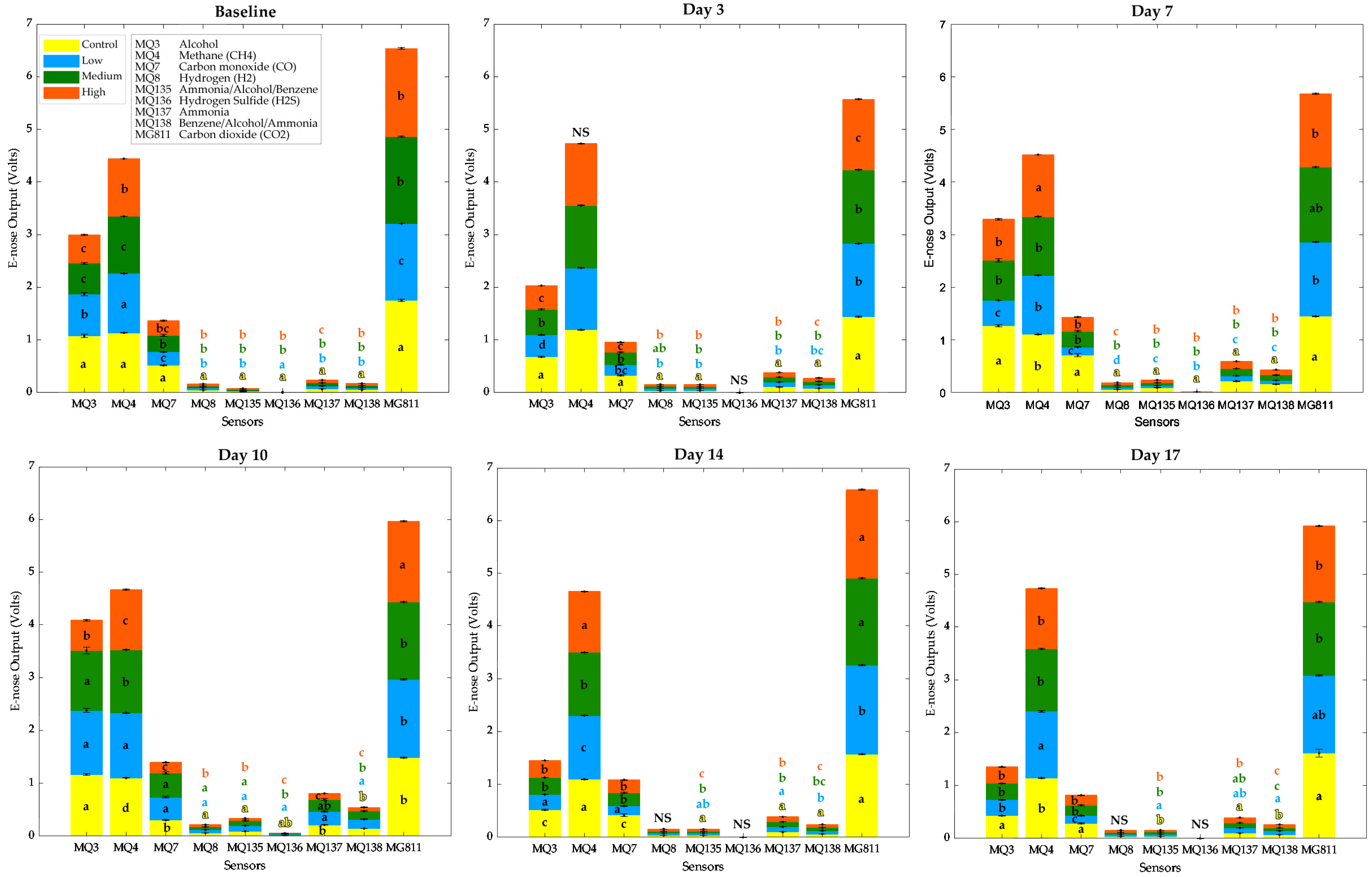
2.4. Electronic Nose Measurements
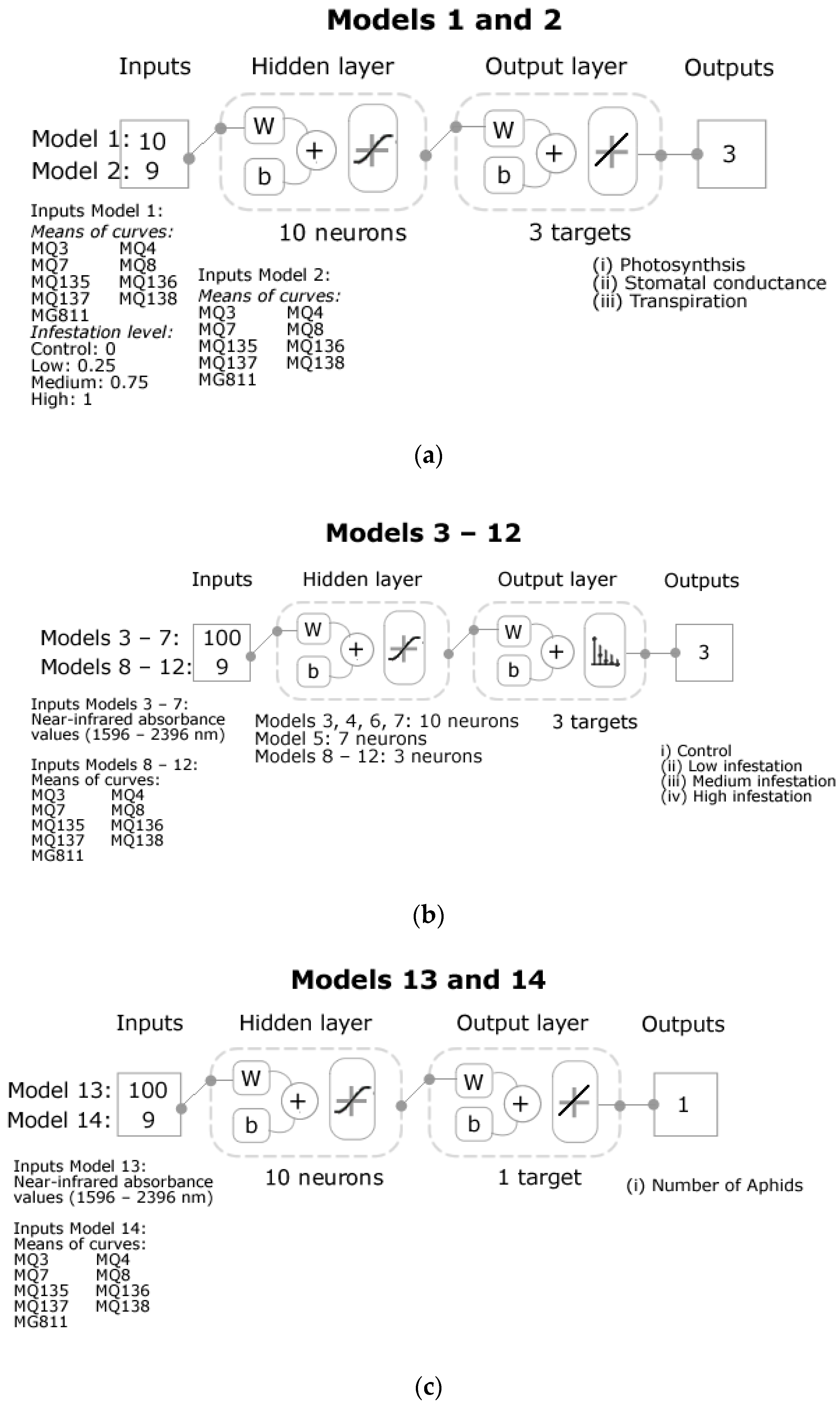
2.5. Statistical Analysis and Machine Learning Modeling
3. Results
4. Discussion
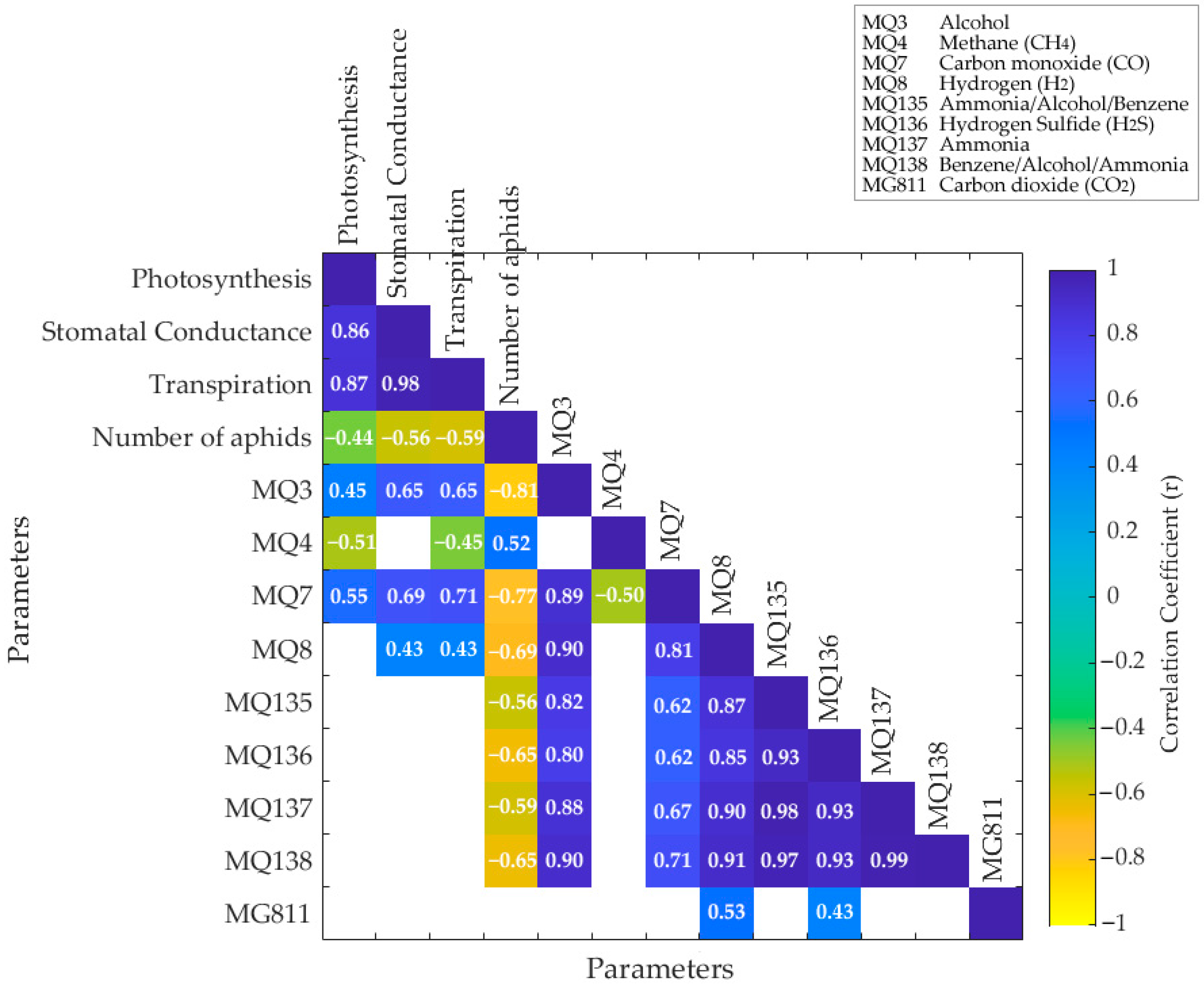
4.1. Physiological Response of Plants to Insect Infestation
4.2. Chemical Fingerprinting and Volatile Compounds’ Response to Insect Infestation
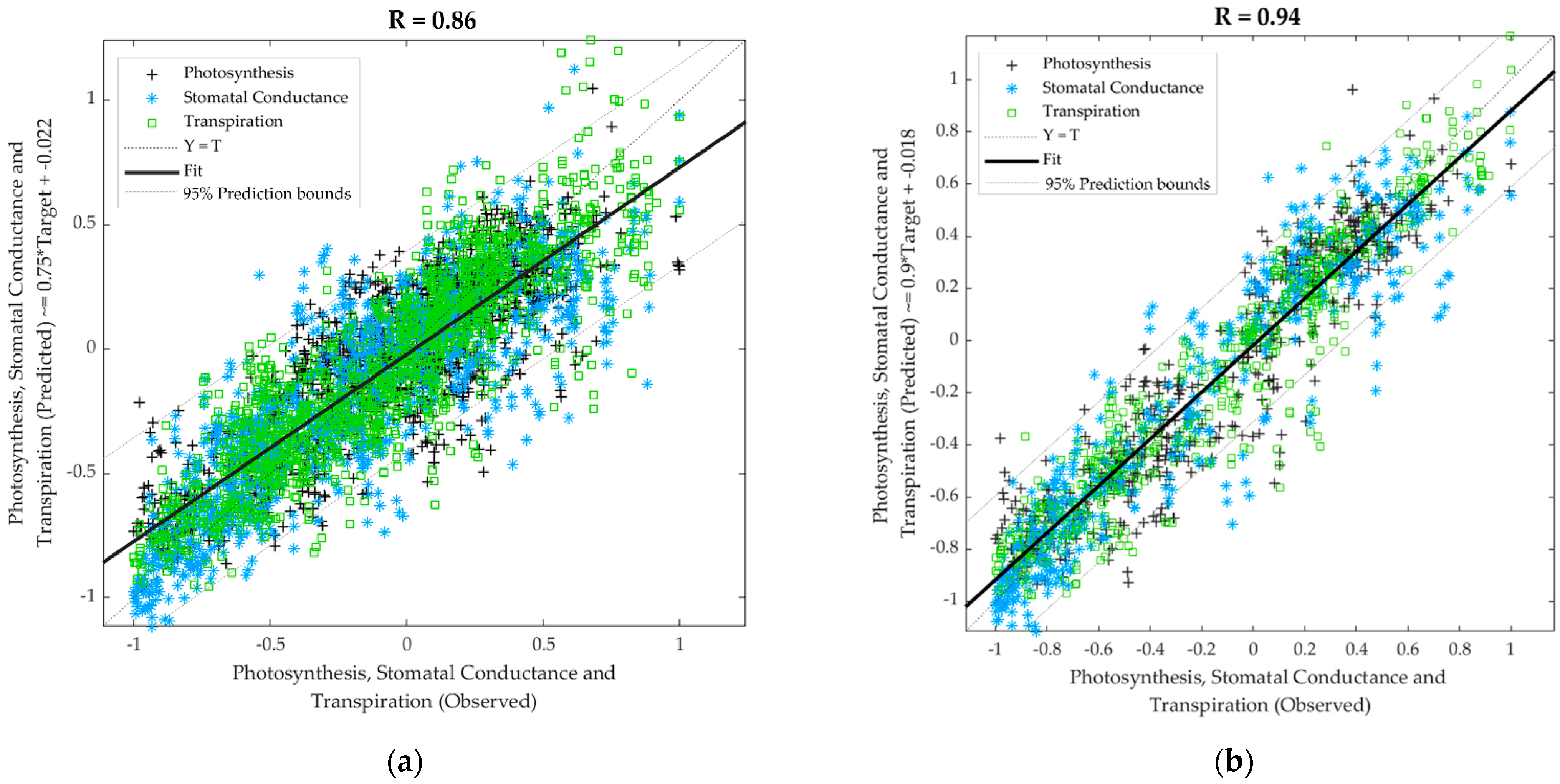
4.3. Machine Learning Models Developed
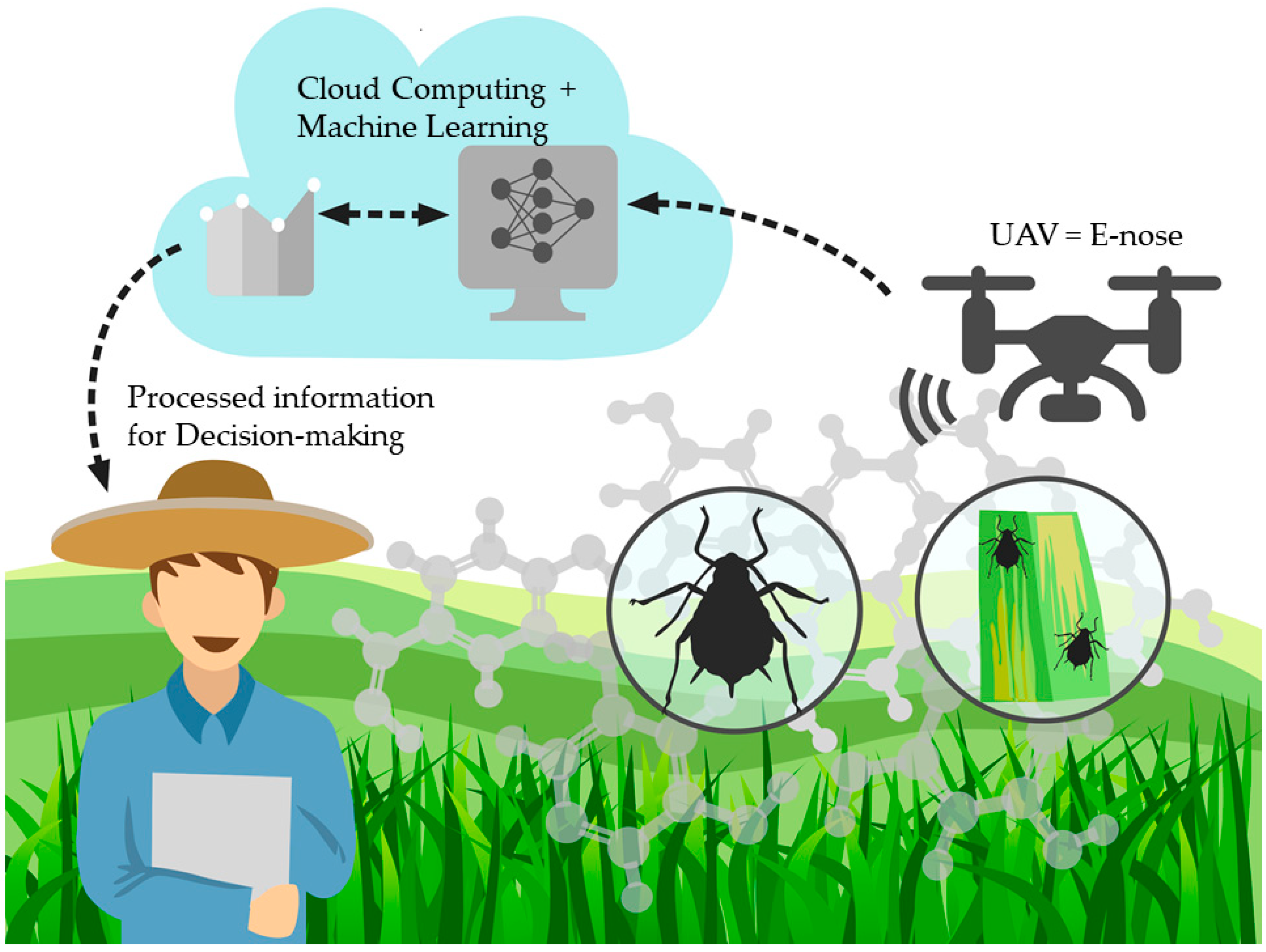
4.4. Deployment Method for ML Models Developed Proposed Using UAV
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wollmann, J.; Schlesener, D.C.H.; Ferreira, M.S.; Krüger, A.; Bernardi, D.; Garcia, J.A.B.; Nunes, A.M.; Garcia, M.S.; Garcia, F. Population Dynamics of Drosophila suzukii (Diptera: Drosophilidae) in Berry Crops in Southern Brazil. Neotrop. Èntomol. 2019, 48, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Satterfield, D.A.; Sillett, T.S.; Chapman, J.W.; Altizer, S.; Marra, P.P. Seasonal insect migrations: Massive, influential, and overlooked. Front. Ecol. Environ. 2020, 18, 335–344. [Google Scholar] [CrossRef]
- Saha, T.; Chandran, N. Chemical ecology and pest management: A review. Int. J. Cardiovasc. Sci. 2017, 5, 618–621. [Google Scholar]
- Kim, K.; Huang, Q.; Lei, C. Advances in insect phototaxis and application to pest management: A review. Pest Manag. Sci. 2019, 75, 3135–3143. [Google Scholar] [CrossRef] [PubMed]
- Preti, M.; Verheggen, F.; Angeli, S. Insect pest monitoring with camera-equipped traps: Strengths and limitations. J. Pest Sci. 2020, 94, 203–217. [Google Scholar] [CrossRef]
- Milosavljević, I.; Hoddle, C.D.; Mafra-Neto, A.; Gómez-Marco, F.; Hoddle, M.S. Use of Digital Video Cameras to Determine the Efficacy of Two Trap Types for Capturing Rhynchophorus palmarum (Coleoptera: Curculionidae). J. Econ. Èntomol. 2020, 113, 3028–3031. [Google Scholar] [CrossRef] [PubMed]
- Remboski, T.B.; de Souza, W.D.; de Aguiar, M.S.; Ferreira, P.R., Jr. Identification of Fruit Fly in Intelligent Traps Using Techniques of Digital Image Processing and Machine Learning. In Proceedings of the 33rd Annual ACM Symposium on Applied Computing, Pau, France, 9–13 April 2018; pp. 260–267. [Google Scholar]
- Chulu, F.; Phiri, J.; Nyirenda, M.; Kabemba, M.M.; Nkunika, P.; Chiwamba, S. Developing an automatic identification and early warning and monitoring web based system of fall army worm based on machine learning in developing countries. Zamb. ICT J. 2019, 3, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Barbedo, J.G.A. Detecting and Classifying Pests in Crops Using Proximal Images and Machine Learning: A Review. AI 2020, 1, 312–328. [Google Scholar] [CrossRef]
- Marković, D.; Vujičić, D.; Tanasković, S.; Đorđević, B.; Ranđić, S.; Stamenković, Z. Prediction of Pest Insect Appearance Using Sensors and Machine Learning. Sensors 2021, 21, 4846. [Google Scholar] [CrossRef]
- Kasinathan, T.; Uyyala, S.R. Machine learning ensemble with image processing for pest identification and classification in field crops. Neural Comput. Appl. 2021, 1–14. [Google Scholar] [CrossRef]
- Lima, M.C.F.; Leandro, M.E.D.D.A.; Valero, C.; Coronel, L.C.P.; Bazzo, C.O.G. Automatic Detection and Monitoring of Insect Pests—A Review. Agriculture 2020, 10, 161. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, Y.; Pu, R.; González-Moreno, P.; Yuan, L.; Wu, K.; Huang, W. Monitoring plant diseases and pests through remote sensing technology: A review. Comput. Electron. Agric. 2019, 165, 104943. [Google Scholar] [CrossRef]
- Velásquez, D.; Sánchez, A.; Sarmiento, S.; Toro, M.; Maiza, M.; Sierra, B. A Method for Detecting Coffee Leaf Rust through Wireless Sensor Networks, Remote Sensing, and Deep Learning: Case Study of the Caturra Variety in Colombia. Appl. Sci. 2020, 10, 697. [Google Scholar] [CrossRef] [Green Version]
- Brunelli, D.; Albanese, A.; D’Acunto, D.; Nardello, M. Energy Neutral Machine Learning Based IoT Device for Pest Detection in Precision Agriculture. IEEE Internet Things Mag. 2019, 2, 10–13. [Google Scholar] [CrossRef]
- Poblete, T.; Camino, C.; Beck, P.; Hornero, A.; Kattenborn, T.; Saponari, M.; Boscia, D.; Navas-Cortes, J.; Zarco-Tejada, P. Detection of Xylella fastidiosa infection symptoms with airborne multispectral and thermal imagery: Assessing bandset reduction performance from hyperspectral analysis. ISPRS J. Photogramm. Remote. Sens. 2020, 162, 27–40. [Google Scholar] [CrossRef]
- Hornero, A.; Hernández-Clemente, R.; North, P.; Beck, P.; Boscia, D.; Navas-Cortes, J.; Zarco-Tejada, P. Monitoring the incidence of Xylella fastidiosa infection in olive orchards using ground-based evaluations, airborne imaging spectroscopy and Sentinel-2 time series through 3-D radiative transfer modelling. Remote. Sens. Environ. 2019, 236, 111480. [Google Scholar] [CrossRef]
- Aasen, H.; Honkavaara, E.; Lucieer, A.; Zarco-Tejada, P.J. Quantitative Remote Sensing at Ultra-High Resolution with UAV Spectroscopy: A Review of Sensor Technology, Measurement Procedures, and Data Correction Workflows. Remote. Sens. 2018, 10, 1091. [Google Scholar] [CrossRef] [Green Version]
- Cellini, A.; Blasioli, S.; Biondi, E.; Bertaccini, A.; Braschi, I.; Spinelli, F. Potential Applications and Limitations of Electronic Nose Devices for Plant Disease Diagnosis. Sensors 2017, 17, 2596. [Google Scholar] [CrossRef] [Green Version]
- Cui, S.; Ling, P.; Zhu, H.; Keener, H.M. Plant Pest Detection Using an Artificial Nose System: A Review. Sensors 2018, 18, 378. [Google Scholar] [CrossRef] [Green Version]
- Lampson, B.D.; Khalilian, A.; Greene, J.K.; Han, Y.J.; Degenhardt, D.C. Development of a Portable Electronic Nose for Detection of Cotton Damaged by Nezara viridula (Hemiptera: Pentatomidae). J. Insects 2014, 2014, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Inocente, E.A.A.; Acosta, N.; Keener, H.M.; Zhu, H.; Ling, P.P. Development of Fast E-nose System for Early-Stage Diagnosis of Aphid-Stressed Tomato Plants. Sensors 2019, 19, 3480. [Google Scholar] [CrossRef] [Green Version]
- Ridgway, C.; Chambers, J.; Portero-Larragueta, E.; Prosser, O. Detection of mite infestation in wheat by electronic nose with transient flow sampling. J. Sci. Food Agric. 1999, 79, 2067–2074. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, J. Detection of age and insect damage incurred by wheat, with an electronic nose. J. Stored Prod. Res. 2007, 43, 489–495. [Google Scholar] [CrossRef]
- Wu, J.; Jayas, D.; Zhang, Q.; White, N.; York, R. Feasibility of the application of electronic nose technology to detect insect infestation in wheat. Can. Biosyst. Eng. 2013, 55, 3.1–3.9. [Google Scholar] [CrossRef]
- Ahouandjinou, S.A.R.M.; Kiki, M.P.A.F.; Badoussi, P.E.N.A.; Assogba, K.M. A Multi-level Smart Monitoring System by Combining an E-Nose and Image Processing for Early Detection of FAW Pest in Agriculture. In Proceedings of the Innovations and Interdisciplinary Solutions for Underserved Areas: 4th EAI International Conference, InterSol 2020, Nairobi, Kenya, 8–9 March 2020; pp. 20–32. [Google Scholar] [CrossRef]
- Cheng, L.; Meng, Q.-H.; Lilienthal, A.J.; Qi, P.-F. Development of compact electronic noses: A review. Meas. Sci. Technol. 2021, 32, 062002. [Google Scholar] [CrossRef]
- Poland, T.M.; Rassati, D. Improved biosecurity surveillance of non-native forest insects: A review of current methods. J. Pest Sci. 2018, 92, 37–49. [Google Scholar] [CrossRef]
- Lan, Y.-B.; Zheng, X.-Z.; Westbrook, J.K.; López, J.; Lacey, R.; Hoffmann, W.C. Identification of Stink Bugs Using an Electronic Nose. J. Bionic Eng. 2008, 5, 172–180. [Google Scholar] [CrossRef]
- Zhou, B.; Wang, J. Use of electronic nose technology for identifying rice infestation by Nilaparvata lugens. Sens. Actuators B Chem. 2011, 160, 15–21. [Google Scholar] [CrossRef]
- Viejo, C.G.; Fuentes, S.; Godbole, A.; Widdicombe, B.; Unnithan, R.R. Development of a low-cost e-nose to assess aroma profiles: An artificial intelligence application to assess beer quality. Sens. Actuators B Chem. 2020, 308, 127688. [Google Scholar] [CrossRef]
- Kratky, B. A suspended pot, non-circulating hydroponic method. Acta Hortic. 2004, 648, 83–89. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Genc, Y.; Hayes, J. The Use of Hydroponics in Abiotic Stress Tolerance Research; InTech Rijeka: Rijeka, Croatia, 2012. [Google Scholar]
- McDonald, G.; Umina, P.; Hangartner, S. Oat Aphid Rhophalosiphum padi. Available online: https://cesaraustralia.com/pestnotes/aphids/oat-aphid/ (accessed on 25 November 2020).
- Jarošík, V.; Honěk, A.; Tichopad, A. Comparison of Field Population Growths of Three Cereal Aphid Species on Winter Wheat. Plant Prot. Sci. 2011, 39, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Costamagna, A.C.; Van Der Werf, W.; Bianchi, F.J.J.A.; Landis, D.A. An exponential growth model with decreasing r captures bottom-up effects on the population growth of Aphis glycines Matsumura (Hemiptera: Aphididae). Agric. For. Èntomol. 2007, 9, 297–305. [Google Scholar] [CrossRef]
- Viejo, C.G.; Tongson, E.; Fuentes, S. Integrating a Low-Cost Electronic Nose and Machine Learning Modelling to Assess Coffee Aroma Profile and Intensity. Sensors 2021, 21, 2016. [Google Scholar] [CrossRef] [PubMed]
- Viejo, C.G.; Torrico, D.D.; Dunshea, F.R.; Fuentes, S. Emerging Technologies Based on Artificial Intelligence to Assess the Quality and Consumer Preference of Beverages. Beverages 2019, 5, 62. [Google Scholar] [CrossRef] [Green Version]
- Viejo, C.G.; Torrico, D.D.; Dunshea, F.R.; Fuentes, S. Development of Artificial Neural Network Models to Assess Beer Acceptability Based on Sensory Properties Using a Robotic Pourer: A Comparative Model Approach to Achieve an Artificial Intelligence System. Beverages 2019, 5, 33. [Google Scholar] [CrossRef] [Green Version]
- Sehar, Z.; Jahan, B.; Masood, A.; Anjum, N.A.; Khan, N.A. Hydrogen peroxide potentiates defense system in presence of sulfur to protect chloroplast damage and photosynthesis of wheat under drought stress. Physiol. Plant. 2020, 172, 922–934. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, L.; Shen, Q.; Yang, J.; Han, X.; Tian, F.; Wu, J. Effects of Water Stress on Photosynthesis, Yield, and Water Use Efficiency in Winter Wheat. Water 2020, 12, 2127. [Google Scholar] [CrossRef]
- Shahzad, M.W.; Ghani, H.; Ayyub, M.; Ali, Q.; Ahmad, H.M.; Faisal, M.; Ali, A.; Qasim, M.U. Performance of some Wheat Cultivars against APHID and Its Damage on Yield and Photosynthesis. J. Glob. Innov. Agric. Soc. Sci. 2019, 105–109. [Google Scholar] [CrossRef]
- Li, Y.; Li, H.; Li, Y.; Zhang, S. Improving water-use efficiency by decreasing stomatal conductance and transpiration rate to maintain higher ear photosynthetic rate in drought-resistant wheat. Crop. J. 2017, 5, 231–239. [Google Scholar] [CrossRef]
- Banerjee, K.; Krishnan, P.; Das, B. Thermal imaging and multivariate techniques for characterizing and screening wheat genotypes under water stress condition. Ecol. Indic. 2020, 119, 106829. [Google Scholar] [CrossRef]
- Francesconi, S.; Balestra, G.M. The modulation of stomatal conductance and photosynthetic parameters is involved in Fusarium head blight resistance in wheat. PLoS ONE 2020, 15, e0235482. [Google Scholar] [CrossRef]
- Ahmed, S.S.; Liu, D.; Simon, J.-C. Impact of water-deficit stress on tritrophic interactions in a wheat-aphid-parasitoid system. PLoS ONE 2017, 12, e0186599. [Google Scholar] [CrossRef] [Green Version]
- Pimenta, A.M.; Scafi, S.H.F.; Pasquini, C.; Raimundo Jr, I.M.; Rohwedder, J.J.; Montenegro, M.d.C.B.; Araújo, A.N. Determination of hydrogen peroxide by near infrared spectroscopy. J. Infrared Spectrosc. 2003, 11, 49–53. [Google Scholar] [CrossRef]
- Fuentes, S.; Tongson, E.; Chen, J.; Viejo, C.G. A Digital Approach to Evaluate the Effect of Berry Cell Death on Pinot Noir Wines’ Quality Traits and Sensory Profiles Using Non-Destructive Near-Infrared Spectroscopy. Beverages 2020, 6, 39. [Google Scholar] [CrossRef]
- De Bei, R.; Fuentes, S.; Wirthensohn, M.; Cozzolino, D.; Tyerman, S. Feasibility study on the use of Near Infrared spectroscopy to measure water status of almond trees. Acta Hortic. 2018, 79–84. [Google Scholar] [CrossRef]
- Terhoeven-Urselmans, T.; Bruns, C.; Schmilewski, G.; Ludwig, B. Quality assessment of growing media with near-infrared spectroscopy: Chemical characteristics and plant assays. Eur. J. Hortic. Sci. 2008, 73, 28. [Google Scholar]
- Burns, D.A.; Ciurczak, E.W. Handbook of Near-Infrared Analysis; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Osborne, B.G.; Fearn, T.; Hindle, P.H. Practical NIR Spectroscopy with Applications in Food and Beverage Analysis; Longman scientific and technical: London, UK, 1993. [Google Scholar]
- Cooper, W.R.; Dillwith, J.W.; Puterka, G.J. Salivary Proteins of Russian Wheat Aphid (Hemiptera: Aphididae). Environ. Èntomol. 2010, 39, 223–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elzinga, D.A.; De Vos, M.; Jander, G. Suppression of Plant Defenses by a Myzus persicae (Green Peach Aphid) Salivary Effector Protein. Mol. Plant-Microbe Interact. 2014, 27, 747–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulak, P.; Proc, K.; Pawłowska, M.; Kasprzycka, A.; Berus, W.; Bieganowski, A. Biogas generation from insects breeding post production wastes. J. Clean. Prod. 2019, 244, 118777. [Google Scholar] [CrossRef]
- Hansen, A.; Moran, N.A. Aphid genome expression reveals host-symbiont cooperation in the production of amino acids. Proc. Natl. Acad. Sci. USA 2011, 108, 2849–2854. [Google Scholar] [CrossRef] [Green Version]
- Chou, S.; Chen, J.M.; Yu, H.; Chen, B.; Zhang, X.; Croft, H.; Khalid, S.; Li, M.; Shi, Q. Canopy-Level Photochemical Reflectance Index from Hyperspectral Remote Sensing and Leaf-Level Non-Photochemical Quenching as Early Indicators of Water Stress in Maize. Remote. Sens. 2017, 9, 794. [Google Scholar] [CrossRef] [Green Version]
- Paz, V.S.; Mikkelsen, T.N.; Johnson, M.; Mo, X.; Morillas, L.; Liu, S.; Shen, L.; Garcia, M. Hyperspectral and thermal sensing of stomatal conductance and photosynthesis under water stress for a C3 (soybean) and a C4 (maize) crop. In Proceedings of the EGU General Assembly Conference Abstracts, Vienna, Austria, 7–12 April 2019. [Google Scholar]
- Rossi, L.; Bagheri, M.; Zhang, W.; Chen, Z.; Burken, J.G.; Ma, X. Using artificial neural network to investigate physiological changes and cerium oxide nanoparticles and cadmium uptake by Brassica napus plants. Environ. Pollut. 2018, 246, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Ryu, D.; Fuentes, S.; Chung, H.; Hernández-Montes, E.; O’Connell, M. Adaptive Estimation of Crop Water Stress in Nectarine and Peach Orchards Using High-Resolution Imagery from an Unmanned Aerial Vehicle (UAV). Remote. Sens. 2017, 9, 828. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, S.; De Bei, R.; Pech, J.; Tyerman, S. Computational water stress indices obtained from thermal image analysis of grapevine canopies. Irrig. Sci. 2012, 30, 523–536. [Google Scholar] [CrossRef]
- Fuentes, S.; Tongson, E.J.; De Bei, R.; Viejo, C.G.; Ristic, R.; Tyerman, S.; Wilkinson, K. Non-Invasive Tools to Detect Smoke Contamination in Grapevine Canopies, Berries and Wine: A Remote Sensing and Machine Learning Modeling Approach. Sensors 2019, 19, 3335. [Google Scholar] [CrossRef] [Green Version]
- Yuan, W.-L.; Xu, B.; Ran, G.-C.; Chen, H.-P.; Zhao, P.-Y.; Huang, Q.-L. Application of imidacloprid controlled-release granules to enhance the utilization rate and control wheat aphid on winter wheat. J. Integr. Agric. 2020, 19, 3045–3053. [Google Scholar] [CrossRef]
- Zhang, H.; Garratt, M.P.; Bailey, A.; Potts, S.G.; Breeze, T. Economic valuation of natural pest control of the summer grain aphid in wheat in South East England. Ecosyst. Serv. 2018, 30, 149–157. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, J.; Zhou, H.; Chen, J. Enhancement of Natural Control Function for Aphids by Intercropping and Infochemical Releasers in Wheat Ecosystem. Integr. Biol. Control 2020, 85–116. [Google Scholar] [CrossRef]
- Peairs, F. Development of Integrated Pest Management Approaches for Russian Wheat Aphid in Colorado USA; GRDC Update Paper: Barton, ACT, Australia, 2017. [Google Scholar]
- Singh, B.; Jasrotia, P. Impact of integrated pest management (IPM) module on major insect-pests of wheat and their natural enemies in North-western plains of India. J. Cereal Res. 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Dixon, A.; Kindlmann, P. Population dynamics of aphids. In Insect Populations in Theory and in Practice; Springer: Berlin/Heidelberg, Germany, 1998; pp. 207–230. [Google Scholar]
- Ahmad, T.; Hassan, M.W.; Jamil, M.; Iqbal, J. Population Dynamics of Aphids (Hemiptera: Aphididae) on Wheat Varieties (Triticum aestivum L.) as Affected by Abiotic Conditions in Bahawalpur, Pakistan. Pak. J. Zool. 2016, 48, 1039–1044. [Google Scholar]
- Brabec, M.; Honěk, A.; Pekár, S.; Martinkova, Z. Population Dynamics of Aphids on Cereals: Digging in the Time-Series Data to Reveal Population Regulation Caused by Temperature. PLoS ONE 2014, 9, e106228. [Google Scholar] [CrossRef] [PubMed]
- Xuesong, S.; Zi, L.; Lei, S.; Jiao, W.; Yang, Z. Aphid Identification and Counting Based on Smartphone and Machine Vision. J. Sens. 2017, 2017, 1–7. [Google Scholar] [CrossRef]
- Lins, E.A.; Rodriguez, J.P.M.; Scoloski, S.I.; Pivato, J.; Lima, M.B.; Fernandes, J.M.C.; Pereira, P.R.V.D.S.; Lau, D.; Rieder, R. A method for counting and classifying aphids using computer vision. Comput. Electron. Agric. 2020, 169, 105200. [Google Scholar] [CrossRef]
- Chen, P.; Li, W.; Yao, S.; Ma, C.; Zhang, J.; Wang, B.; Zheng, C.; Xie, C.; Liang, D. Recognition and counting of wheat mites in wheat fields by a three-step deep learning method. Neurocomputing 2021, 437, 21–30. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample/ Parameter | Photosynthesis (µmol CO2 m−2 s−1) | Stomatal Conductance (mol H2O m−2 s−1) | Transpiration (mmol H2O m−2 s−1) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Measurement | BL | D3 | D7 | D10 | D14 | D17 | BL | D3 | D7 | D10 | D14 | D17 | BL | D3 | D7 | D10 | D14 | D17 |
| Control | 6.78 | 9.18 a | 13.78 a | 12.47 a | 13.22 a | 12.75 a | 0.16 | 0.32 a | 0.51 a | 0.55 a | 0.50 a | 0.62 a | 2.35 | 3.60 a | 4.84 a | 4.16 a | 4.00 a | 6.00 a |
| ±0.32 | ±0.01 | ±0.13 | ±0.30 | ±0.02 | ±0.12 | ±0.16 | ±0.01 | ±0.05 | ±0.25 | ±0.02 | ±0.09 | ±0.24 | ±0.02 | ±0.06 | ±0.28 | ±0.02 | ±0.06 | |
| Low | 4.50 | 9.65 ab | 11.34 b | 10.49 b | 11.42 b | 10.27 b | 0.07 | 0.28 a | 0.35 c | 0.36 b | 0.37 bc | 0.40 c | 1.29 | 3.09 b | 3.81 c | 3.53 b | 2.95 c | 4.96 b |
| ±0.23 | ±0.01 | ±0.09 | ±0.40 | ±0.02 | ±0.13 | ±0.41 | ±0.03 | ±0.16 | ±0.27 | ±0.02 | ±0.12 | ±0.27 | ±0.02 | ±0.10 | ±0.23 | ±0.02 | ±0.11 | |
| Medium | 7.03 | 7.52 c | 12.19 b | 10.37 b | 10.70 b | 10.84 b | 0.15 | 0.16 b | 0.44 ab | 0.36 b | 0.33 c | 0.51 b | 2.19 | 2.11 c | 4.34 b | 3.12 b | 2.79 c | 5.36 b |
| ±0.34 | ±02 | ±0.16 | ±0.65 | ±0.02 | ±0.20 | ±0.19 | ±0.02 | ±0.13 | ±0.33 | ±0.03 | ±0.17 | ±0.34 | ±0.03 | ±0.19 | ±0.24 | ±0.03 | ±0.15 | |
| High | 7.03 | 10.93 a | 13.49 a | 10.67 b | 13.25 a | 11.07 b | 0.18 | 0.27 a | 0.44 b | 0.35 b | 0.44 ab | 0.50 b | 2.47 | 2.73 b | 4.43 b | 3.21 b | 3.53 b | 5.34 b |
| ±0.37 | ±0.02 | ±0.15 | ±0.34 | ±0.01 | ±0.09 | ±0.22 | ±0.02 | ±0.07 | ±0.21 | ±0.02 | ±0.10 | ±0.25 | ±0.02 | ±0.09 | ±0.30 | ±0.02 | ±0.11 | |
| Stage | Samples | Observations | R | b | Performance (MSE) |
|---|---|---|---|---|---|
| Model 1—General (all treatments and measurement days)—10 neurons | |||||
| Training | 1008 | 3024 | 0.87 | 0.75 | 0.05 |
| Testing | 432 | 1296 | 0.83 | 0.75 | 0.06 |
| Overall | 1440 | 4320 | 0.86 | 0.75 | - |
| Model 2—Baseline and control—10 neurons | |||||
| Training | 378 | 1134 | 0.95 | 0.90 | 0.02 |
| Testing | 162 | 486 | 0.93 | 0.90 | 0.04 |
| Overall | 540 | 1620 | 0.94 | 0.90 | - |
| Stage | Samples | Accuracy | Error | Performance (MSE) |
|---|---|---|---|---|
| Model 3—Baseline + Day 3—10 neurons | ||||
| Training | 404 | 100% | 0.0% | <0.01 |
| Validation | 86 | 88.4% | 11.6% | 0.05 |
| Testing | 86 | 88.4% | 11.6% | 0.05 |
| Overall | 576 | 96.5% | 3.5% | - |
| Model 4—Day 7—10 neurons | ||||
| Training | 202 | 100% | 0.0% | <0.01 |
| Validation | 43 | 95.3% | 4.7% | 0.02 |
| Testing | 43 | 93.0% | 7.0% | 0.02 |
| Overall | 288 | 98.3% | 1.7% | - |
| Model 5—Day 10—7 neurons | ||||
| Training | 202 | 100% | 0.0% | <0.01 |
| Validation | 43 | 97.7% | 2.3% | 0.01 |
| Testing | 43 | 95.3% | 4.7% | 0.02 |
| Overall | 288 | 99.0% | 1.0% | - |
| Model 6—Day 14—10 neurons | ||||
| Training | 202 | 100% | 0.0% | <0.01 |
| Validation | 43 | 90.7% | 9.3% | 0.05 |
| Testing | 43 | 86.0% | 14.0% | 0.04 |
| Overall | 288 | 96.5% | 3.5% | - |
| Model 7—Day 17—10 neurons | ||||
| Training | 202 | 100% | 0.0% | <0.01 |
| Validation | 43 | 97.7% | 2.3% | 0.01 |
| Testing | 43 | 97.7% | 2.3% | 0.01 |
| Overall | 288 | 99.3% | 0.7% | - |
| Stage | Samples | Accuracy | Error | Performance (MSE) |
|---|---|---|---|---|
| Model 8—Baseline + Day 3—3 neurons | ||||
| Training | 336 | 99.7% | 0.3% | <0.01 |
| Testing | 144 | 95.1% | 4.9% | 0.02 |
| Overall | 480 | 98.3% | 1.7% | - |
| Model 9—Day 7—3 neurons | ||||
| Training | 168 | 100% | 0.0% | <0.01 |
| Testing | 72 | 94.4% | 5.6% | 0.03 |
| Overall | 240 | 98.3% | 1.7% | - |
| Model 10—Day 10—3 neurons | ||||
| Training | 168 | 100% | 0.0% | <0.01 |
| Testing | 72 | 97.2% | 2.8% | 0.01 |
| Overall | 240 | 99.2% | 0.8% | - |
| Model 11—Day 14—3 neurons | ||||
| Training | 168 | 98.8% | 1.2% | <0.01 |
| Testing | 72 | 97.2% | 2.8% | 0.02 |
| Overall | 240 | 98.3% | 1.7% | - |
| Model 12—Day 17—3 neurons | ||||
| Training | 168 | 97.6% | 2.4% | <0.01 |
| Testing | 72 | 86.1% | 13.9% | 0.06 |
| Overall | 240 | 94.2% | 5.8% | - |
| Stage | Samples | Observations | R | Slope | Performance (MSE) |
|---|---|---|---|---|---|
| Model 13—NIR Day 7–Day 17—10 neurons | |||||
| Training | 605 | 605 | 0.99 | 0.97 | 555 |
| Testing | 259 | 259 | 0.94 | 0.98 | 3078 |
| Overall | 864 | 864 | 0.97 | 0.97 | - |
| Model 14—E-Nose Day 7–Day 17—10 neurons | |||||
| Training | 504 | 504 | 0.99 | 0.98 | 20,014 |
| Testing | 216 | 216 | 0.98 | 0.94 | 40,125 |
| Overall | 720 | 720 | 0.99 | 0.97 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuentes, S.; Tongson, E.; Unnithan, R.R.; Gonzalez Viejo, C. Early Detection of Aphid Infestation and Insect-Plant Interaction Assessment in Wheat Using a Low-Cost Electronic Nose (E-Nose), Near-Infrared Spectroscopy and Machine Learning Modeling. Sensors 2021, 21, 5948. https://doi.org/10.3390/s21175948
Fuentes S, Tongson E, Unnithan RR, Gonzalez Viejo C. Early Detection of Aphid Infestation and Insect-Plant Interaction Assessment in Wheat Using a Low-Cost Electronic Nose (E-Nose), Near-Infrared Spectroscopy and Machine Learning Modeling. Sensors. 2021; 21(17):5948. https://doi.org/10.3390/s21175948
Chicago/Turabian StyleFuentes, Sigfredo, Eden Tongson, Ranjith R. Unnithan, and Claudia Gonzalez Viejo. 2021. "Early Detection of Aphid Infestation and Insect-Plant Interaction Assessment in Wheat Using a Low-Cost Electronic Nose (E-Nose), Near-Infrared Spectroscopy and Machine Learning Modeling" Sensors 21, no. 17: 5948. https://doi.org/10.3390/s21175948
APA StyleFuentes, S., Tongson, E., Unnithan, R. R., & Gonzalez Viejo, C. (2021). Early Detection of Aphid Infestation and Insect-Plant Interaction Assessment in Wheat Using a Low-Cost Electronic Nose (E-Nose), Near-Infrared Spectroscopy and Machine Learning Modeling. Sensors, 21(17), 5948. https://doi.org/10.3390/s21175948