
Developing a Method to Estimate the Downstream Metabolite Signals from Hyperpolarized [1-13C]Pyruvate

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Signal Estimation and Correction
2.2. MR Dynamic Signal Simulations
2.3. In Vitro Study: Cell Preparation and Irradiation
2.4. [1-13C]Pyruvate Hyperpolarization and In Vitro Experiments
2.5. Data Analysis
2.5.1. In Vitro Analysis

2.5.2. Simulations
3. Results
3.1. Simulation
3.2. In Vitro Study
3.2.1. Before and after In Vitro Signal Correction
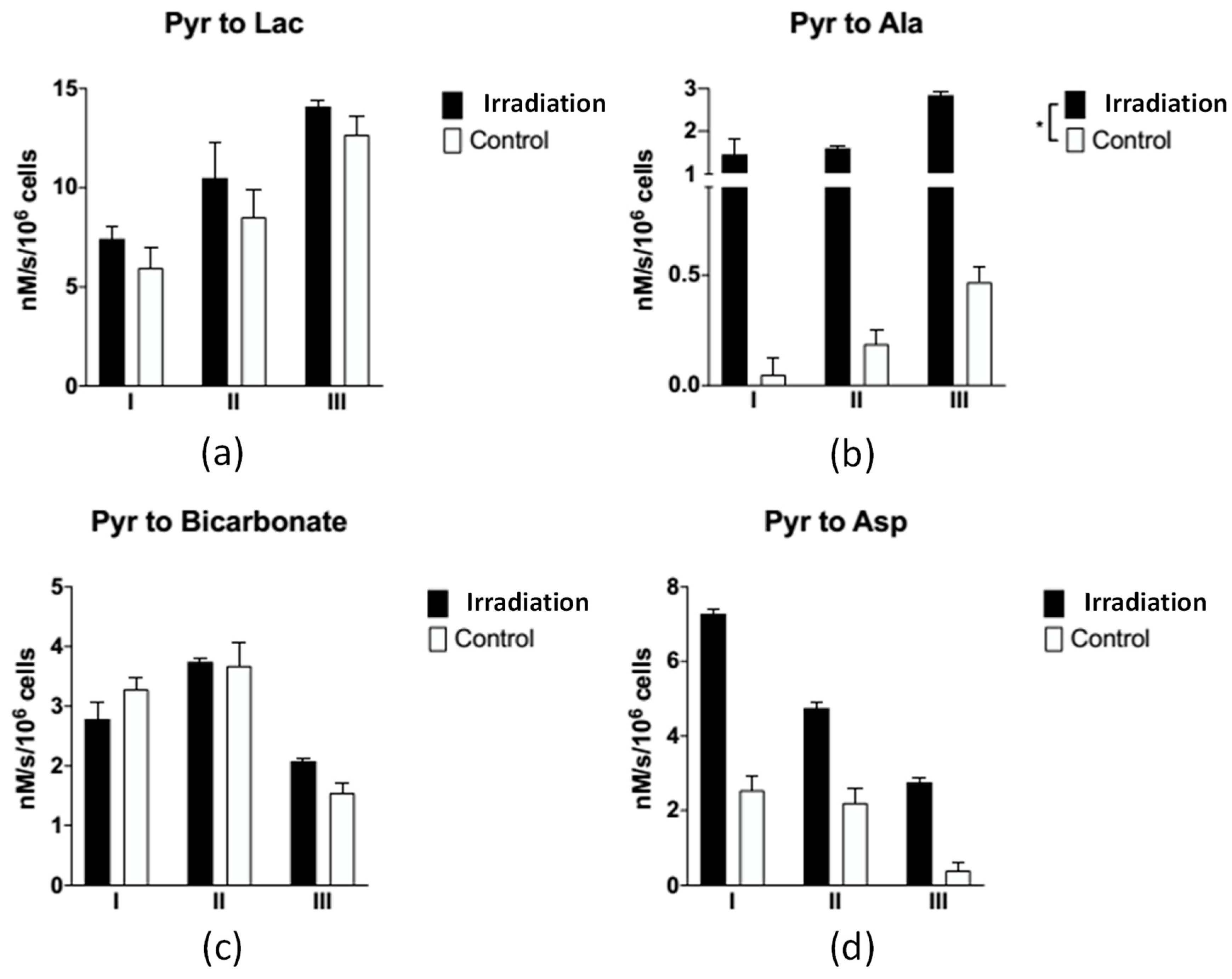
3.2.2. A Metabolite Signal Comparison between Irradiated and Control Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dominguez, H.; Rollin, C.; Guyonvarch, A.; Guerquin-Kern, J.-L.; Cocaign-Bousquet, M.; Lindley, N.D. Carbon-flux distribution in the central metabolic pathways of Corynebacterium glutamicum during growth on fructose. Eur. J. Biochem. 1998, 254, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O. On the Origin of Cancer Cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef] [PubMed]
- de la Cruz-López, K.G.; Castro-Muñoz, L.J.; Reyes-Hernández, D.O.; García-Carrancá, A.; Manzo-Merino, J. Lactate in the Regulation of Tumor Microenvironment and Therapeutic Approaches. Front. Oncol. 2019, 9, 1143. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, J.K.; Holmes, E.; Kinross, J.M.; Darzi, A.W.; Takats, Z.; Lindon, J.C. Metabolic phenotyping in clinical and surgical environments. Nature 2012, 491, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Larson, S.M.; Schwartz, L.H. 18F-FDG PET as a candidate for “qualified biomarker”: Functional assessment of treatment response in oncology. J. Nucl. Med. 2006, 47, 901–903. [Google Scholar] [PubMed]
- Menzel, M.I.; Farrell, E.V.; Janich, M.A.; Khegai, O.; Wiesinger, F.; Nekolla, S.; Otto, A.M.; Haase, A.; Schulte, R.F.; Schwaiger, M. Multimodal Assessment of In Vivo Metabolism with Hyperpolarized [1-13C] MR Spectroscopy and 18F-FDG PET Imaging in Hepatocellular Carcinoma Tumor–Bearing Rats. J. Nucl. Med. 2013, 54, 1113–1119. [Google Scholar] [CrossRef] [Green Version]
- Saito, K.; Matsumoto, S.; Takakusagi, Y.; Matsuo, M.; Morris, H.D.; Lizak, M.J.; Munasinghe, J.P.; Devasahayam, N.; Subramanian, S.; Mitchell, J.B.; et al. 13C-MR Spectroscopic Imaging with Hyperpolarized [1-13C]Pyruvate Detects Early Response to Radiotherapy in SCC Tumors and HT-29 Tumors. Clin. Cancer Res. 2015, 21, 5073–5081. [Google Scholar] [CrossRef] [Green Version]
- Ardenkjær-Larsen, J.H.; Fridlund, B.; Gram, A.; Hansson, G.; Hansson, L.; Lerche, M.H.; Servin, R.; Thaning, M.; Golman, K. Increase in signal-to-noise ratio of >10,000 times in liquid-state NMR. Proc. Natl. Acad. Sci. USA 2003, 100, 10158–10163. [Google Scholar] [CrossRef] [Green Version]
- Ardenkjaer-Larsen, J.H.; Leach, A.M.; Clarke, N.; Urbahn, J.; Anderson, D.; Skloss, T.W. Dynamic nuclear polarization polarizer for sterile use intent. NMR Biomed. 2011, 24, 927–932. [Google Scholar] [CrossRef]
- Kurhanewicz, J.; Vigneron, D.B.; Brindle, K.; Chekmenev, E.Y.; Comment, A.; Cunningham, C.H.; DeBerardinis, R.J.; Green, G.G.; Leach, M.O.; Rajan, S.S. Analysis of cancer metabolism by imaging hyperpolarized nuclei: Prospects for translation to clinical research. Neoplasia 2011, 13, 81–97. [Google Scholar] [CrossRef] [Green Version]
- Sibisi, S.; Skilling, J.; Brereton, R.G.; Laue, E.D.; Staunton, J. Maximum entropy signal processing in practical NMR spectroscopy. Nature 1984, 311, 446–447. [Google Scholar] [CrossRef]
- Chen, G.Y.; Bui, T.D. Multiwavelets denoising using neighboring coefficients. IEEE Signal Process. Lett. 2003, 10, 211–214. [Google Scholar] [CrossRef]
- Ding, Y.; Selesnick, I.W. Artifact-Free Wavelet Denoising: Non-convex Sparse Regularization, Convex Optimization. IEEE Signal Process. Lett. 2015, 22, 1364–1368. [Google Scholar] [CrossRef] [Green Version]
- Madadi, Z.; Anand, G.V.; Premkumar, A.B. Signal Detection in Generalized Gaussian Noise by Nonlinear Wavelet Denoising. IEEE Trans. Circuits Syst. I Regul. Pap. 2013, 60, 2973–2986. [Google Scholar] [CrossRef]
- Chang, S.G.; Bin, Y.; Vetterli, M. Spatially adaptive wavelet thresholding with context modeling for image denoising. IEEE Trans. Image Process. 2000, 9, 1522–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sendur, L.; Selesnick, I.W. Bivariate shrinkage functions for wavelet-based denoising exploiting interscale dependency. IEEE Trans. Signal Process. 2002, 50, 2744–2756. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; He, Z.; Feng, G.; Xiao, B. An Improved Wavelet Denoising Algorithm for Wideband Radar Targets Detection. Circuits Syst. Signal Process. 2013, 32, 2003–2026. [Google Scholar] [CrossRef]
- Mallat, S.G. A theory for multiresolution signal decomposition: The wavelet representation. IEEE Trans. Pattern Anal. Mach. Intell. 1989, 11, 674–693. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, M.; Anderson, C.L.; Freed, J.H. A New Wavelet Denoising Method for Selecting Decomposition Levels and Noise Thresholds. IEEE Access 2016, 4, 3862–3877. [Google Scholar] [CrossRef]
- Rowland, B.; Merugumala, S.K.; Liao, H.; Creager, M.A.; Balschi, J.; Lin, A.P. Spectral improvement by fourier thresholding of in vivo dynamic spectroscopy data. Magn. Reson. Med. 2016, 76, 978–985. [Google Scholar] [CrossRef] [Green Version]
- Bier, E.A.; Robertson, S.H.; Schrank, G.M.; Rackley, C.; Mammarappallil, J.G.; Rajagopal, S.; McAdams, H.P.; Driehuys, B. A protocol for quantifying cardiogenic oscillations in dynamic 129Xe gas exchange spectroscopy: The effects of idiopathic pulmonary fibrosis. NMR Biomed. 2019, 32, e4029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brender, J.R.; Kishimoto, S.; Merkle, H.; Reed, G.; Hurd, R.E.; Chen, A.P.; Ardenkjaer-Larsen, J.H.; Munasinghe, J.; Saito, K.; Seki, T.; et al. Dynamic Imaging of Glucose and Lactate Metabolism by 13C-MRS without Hyperpolarization. Sci. Rep. 2019, 9, 3410. [Google Scholar] [CrossRef] [PubMed]
- Ratakack, M.T.; VanOstenburg, D.O. Computer Simulation Teaches Fourier Transform NMR. Comput. Phys. Educ. 1994, 8, 279–285. [Google Scholar] [CrossRef]
- Daniels, C.J.; McLean, M.A.; Schulte, R.F.; Robb, F.J.; Gill, A.B.; McGlashan, N.; Graves, M.J.; Schwaiger, M.; Lomas, D.J.; Brindle, K.M.; et al. A comparison of quantitative methods for clinical imaging with hyperpolarized 13C-pyruvate. NMR Biomed. 2016, 29, 387–399. [Google Scholar] [CrossRef] [Green Version]
- Rangan, S.R. A new human cell line (FaDu) from a hypopharyngeal carcinoma. Cancer 1972, 29, 117–121. [Google Scholar] [CrossRef]
- Kasten-Pisula, U.; Menegakis, A.; Brammer, I.; Borgmann, K.; Mansour, W.Y.; Degenhardt, S.; Krause, M.; Schreiber, A.; Dahm-Daphi, J.; Petersen, C.; et al. The extreme radiosensitivity of the squamous cell carcinoma SKX is due to a defect in double-strand break repair. Radiother. Oncol. J. Eur. Soc. Ther. Radiol. Oncol. 2009, 90, 257–264. [Google Scholar] [CrossRef]
- Suit, H.D.; Zietman, A.; Tomkinson, K.; Ramsay, J.; Gerweck, L.; Sedlacek, R. Radiation response of xenografts of a human squamous cell carcinoma and a glioblastoma multiforme: A progress report. Int. J. Radiat. Oncol. Biol. Phys. 1990, 18, 365–373. [Google Scholar] [CrossRef]
- Toulany, M.; Dittmann, K.; Krüger, M.; Baumann, M.; Rodemann, H.P. Radioresistance of K-Ras mutated human tumor cells is mediated through EGFR-dependent activation of PI3K-AKT pathway. Radiother. Oncol. J. Eur. Soc. Ther. Radiol. Oncol. 2005, 76, 143–150. [Google Scholar] [CrossRef]
- Dittmann, K.; Mayer, C.; Paasch, A.; Huber, S.; Fehrenbacher, B.; Schaller, M.; Rodemann, H.P. Nuclear EGFR renders cells radio-resistant by binding mRNA species and triggering a metabolic switch to increase lactate production. Radiother. Oncol. 2015, 116, 431–437. [Google Scholar] [CrossRef]
- Lai, Y.C.; Hsieh, C.Y.; Lu, K.Y.; Sung, C.H.; Ho, H.Y.; Cheng, M.L.; Chen, A.P.; Ng, S.H.; Chen, F.H.; Lin, G. Monitoring Early Glycolytic Flux Alterations Following Radiotherapy in Cancer and Immune Cells: Hyperpolarized Carbon-13 Magnetic Resonance Imaging Study. Metabolites 2021, 11, 518. [Google Scholar] [CrossRef]
- Hurd, R.E.; Yen, Y.-F.; Chen, A.; Ardenkjaer-Larsen, J.H. Hyperpolarized 13C metabolic imaging using dissolution dynamic nuclear polarization. J. Magn. Reson. Imaging 2012, 36, 1314–1328. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, H.A.I.; Comment, A.; Schwitter, J. Assessment of Aspartate and Bicarbonate Produced From Hyperpolarized [1-13C]Pyruvate as Markers of Renal Gluconeogenesis. Front. Physiol. 2021, 12, 792769. [Google Scholar] [CrossRef] [PubMed]
- Krysztofiak, A.; Szymonowicz, K.; Hlouschek, J.; Xiang, K.; Waterkamp, C.; Larafa, S.; Goetting, I.; Vega-Rubin-de-Celis, S.; Theiss, C.; Matschke, V.; et al. Metabolism of cancer cells commonly responds to irradiation by a transient early mitochondrial shutdown. iScience 2021, 24, 103366. [Google Scholar] [CrossRef]
- Merritt, M.E.; Harrison, C.; Sherry, A.D.; Malloy, C.R.; Burgess, S.C. Flux through hepatic pyruvate carboxylase and phosphoenolpyruvate carboxykinase detected by hyperpolarized 13C magnetic resonance. Proc. Natl. Acad. Sci. USA 2011, 108, 19084–19089. [Google Scholar] [CrossRef] [Green Version]
- Hassel, B. Pyruvate carboxylation in neurons. J. Neurosci. Res. 2001, 66, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.-Y.; Hsieh, Y.-T.; Huang, J.-M.; Liu, C.-J.; Chuang, L.-T.; Huang, P.-C.; Kuo, T.-Y.; Chia, H.-Y.; Chou, C.-Y.; Chang, C.-W.; et al. Determination of Pyruvate Metabolic Fates Modulates Head and Neck Tumorigenesis. Neoplasia 2019, 21, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Schniewind, I.; Hadiwikarta, W.W.; Grajek, J.; Poleszczuk, J.; Richter, S.; Peitzsch, M.; Muller, J.; Klusa, D.; Beyreuther, E.; Lock, S.; et al. Cellular plasticity upon proton irradiation determines tumor cell radiosensitivity. Cell Rep. 2022, 38, 110422. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Funchain, P.; Retuerto, M.; Jurevic, R.J.; Fowler, N.; Burkey, B.; Eng, C.; Ghannoum, M.A. Metabolomic analysis identifies differentially produced oral metabolites, including the oncometabolite 2-hydroxyglutarate, in patients with head and neck squamous cell carcinoma. BBA Clin. 2017, 7, 8–15. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KAB (×10−2) (s−1) | KAC (×10−3) (s−1) | |
|---|---|---|
| Raw data | 1.00 ± 0.04 | 5.11 ± 0.23 |
| Processed data | 1.05 ± 0.03 | 5.42 ± 0.18 |
| Processed a | Raw a | NMR b | ||||
|---|---|---|---|---|---|---|
| IR. | Con. | IR. | Con. | IR. | Con. | |
| Pyr → Lac | 0.172 | 0.166 | ||||
| Pyr → Bic | 0.017 | 0.016 | ||||
| Experiment I | Experiment II | Experiment III | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Irradiated | Control | Ratio b | Irradiated | Control | Ratio | Irradiated | Control | Ratio | |
| lac/pyr a | 8.00 × 10−2 | 1.60 × 10−1 | 5.00 × 10−1 | 1.72 × 10−1 | 1.66 × 10−1 | 1.04 × 100 | 3.30 × 10−1 | 2.50 × 10−1 | 1.32 × 100 |
| bic/pyr | 3.00 × 10−2 | 4.00 × 10−2 | 7.50 × 10−1 | 1.68 × 10−2 | 1.62 × 10−2 | 1.04 × 100 | 2.48 × 10−2 | 2.69 × 10−2 | 9.20 × 10−1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsieh, C.-Y.; Sung, C.-H.; Shen, Y.-L.; Lai, Y.-C.; Lu, K.-Y.; Lin, G. Developing a Method to Estimate the Downstream Metabolite Signals from Hyperpolarized [1-13C]Pyruvate. Sensors 2022, 22, 5480. https://doi.org/10.3390/s22155480
Hsieh C-Y, Sung C-H, Shen Y-L, Lai Y-C, Lu K-Y, Lin G. Developing a Method to Estimate the Downstream Metabolite Signals from Hyperpolarized [1-13C]Pyruvate. Sensors. 2022; 22(15):5480. https://doi.org/10.3390/s22155480
Chicago/Turabian StyleHsieh, Ching-Yi, Cheng-Hsuan Sung, Yi-Liang (Eric) Shen, Ying-Chieh Lai, Kuan-Ying Lu, and Gigin Lin. 2022. "Developing a Method to Estimate the Downstream Metabolite Signals from Hyperpolarized [1-13C]Pyruvate" Sensors 22, no. 15: 5480. https://doi.org/10.3390/s22155480
APA StyleHsieh, C. -Y., Sung, C. -H., Shen, Y. -L., Lai, Y. -C., Lu, K. -Y., & Lin, G. (2022). Developing a Method to Estimate the Downstream Metabolite Signals from Hyperpolarized [1-13C]Pyruvate. Sensors, 22(15), 5480. https://doi.org/10.3390/s22155480