
Estimation of ANT-DBS Electrodes on Target Positioning Based on a New PerceptTM PC LFP Signal Analysis

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subject
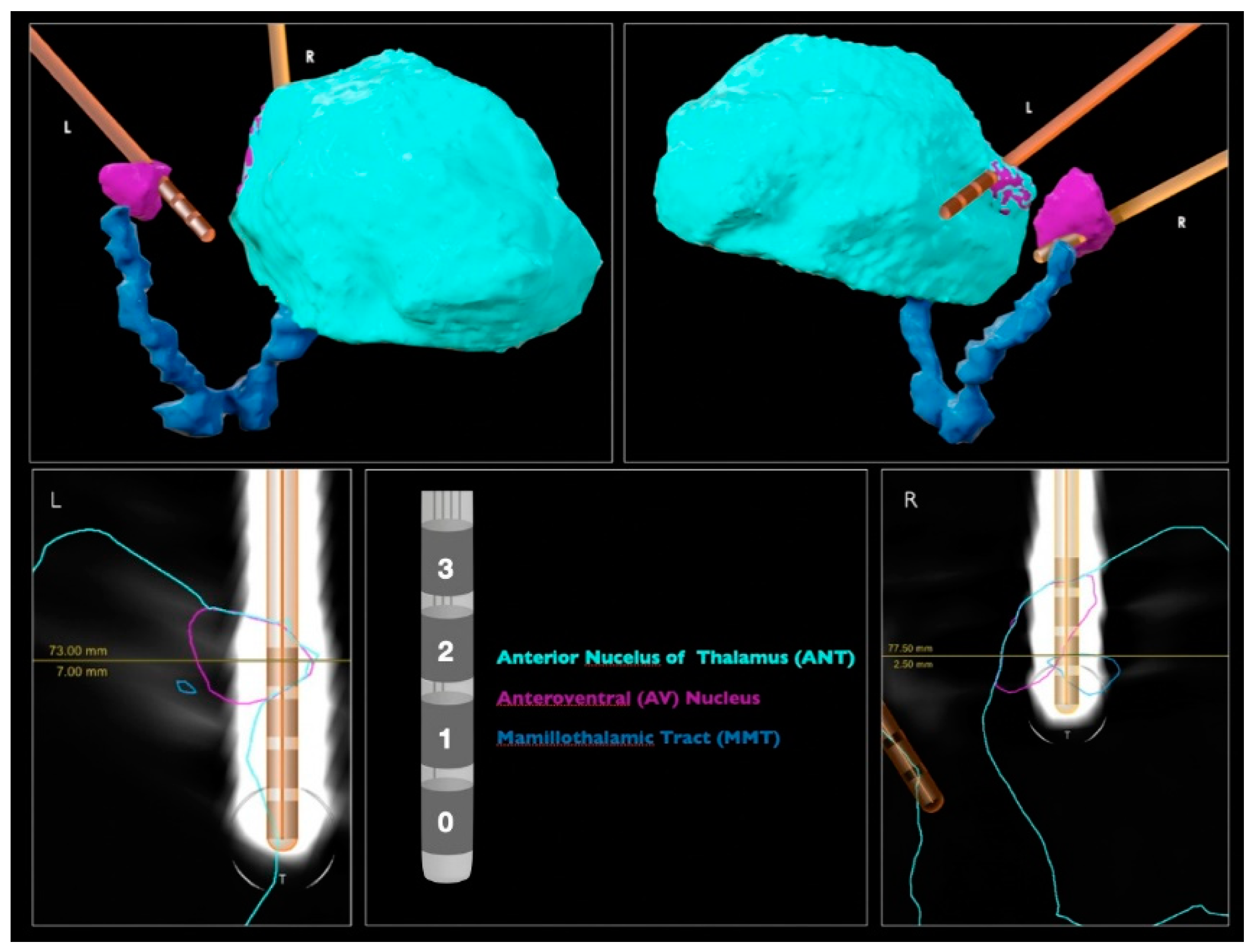
2.2. DBS Lead Contact Localization
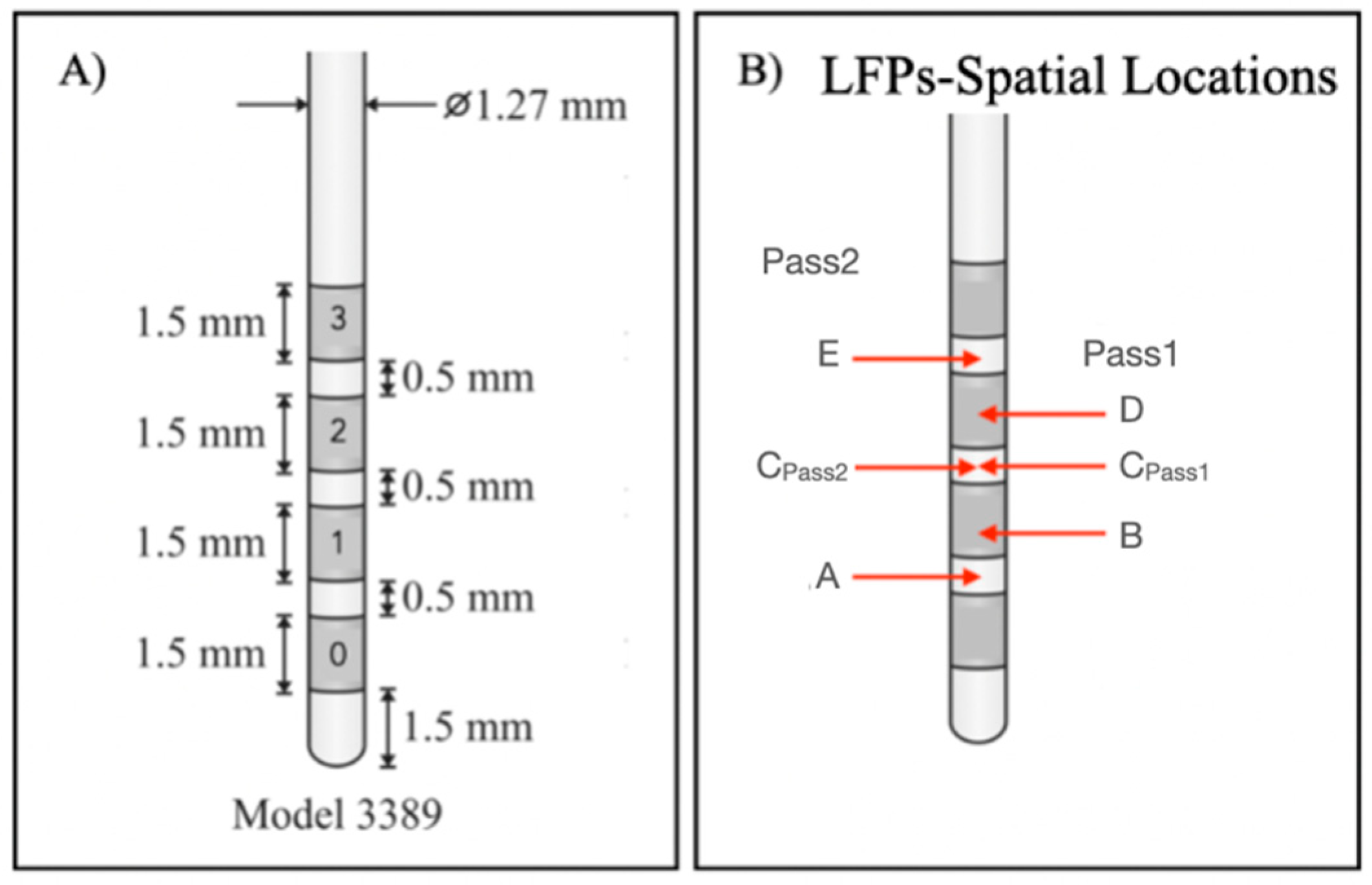
2.3. LFP Recording
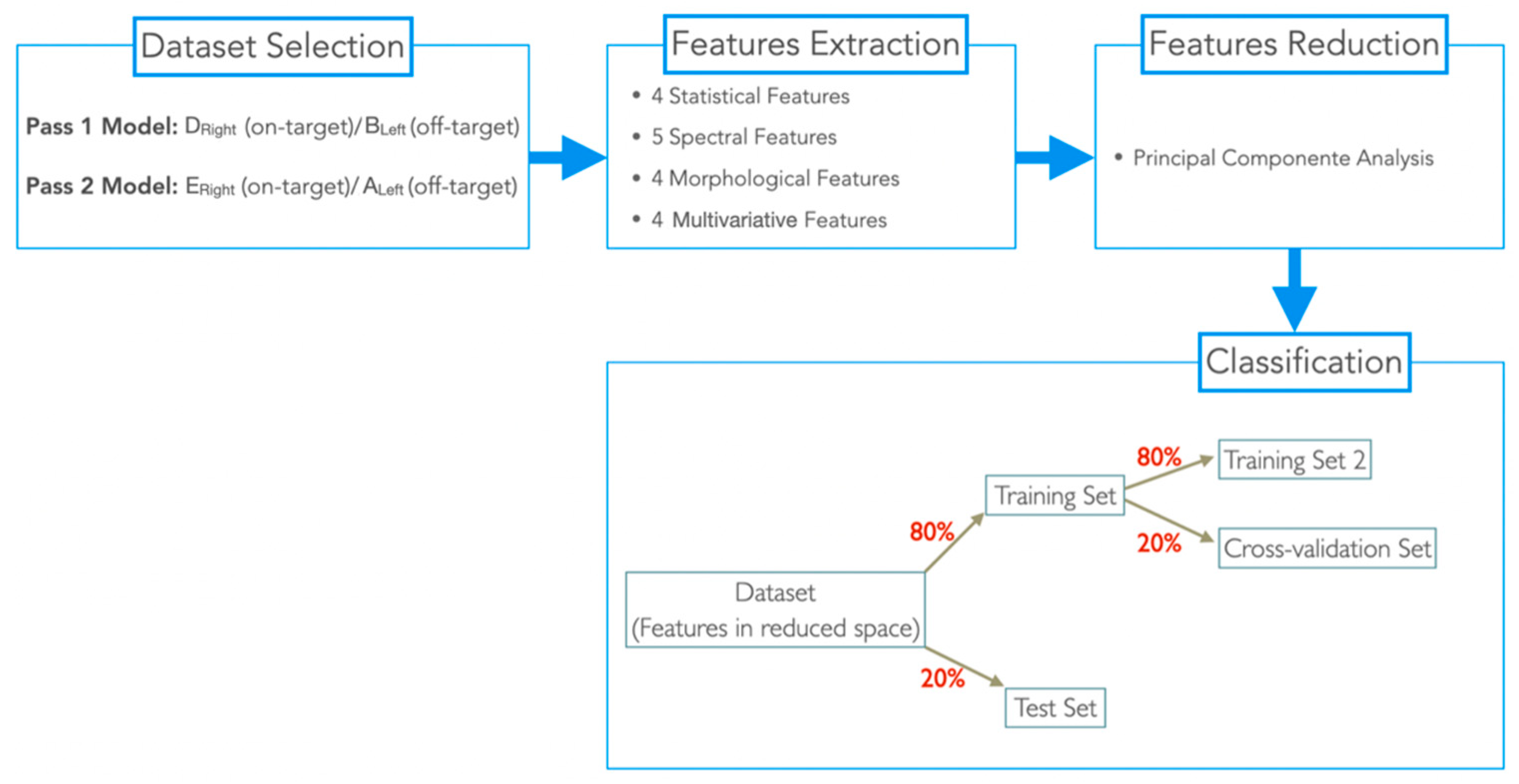
2.4. Dataset Selection
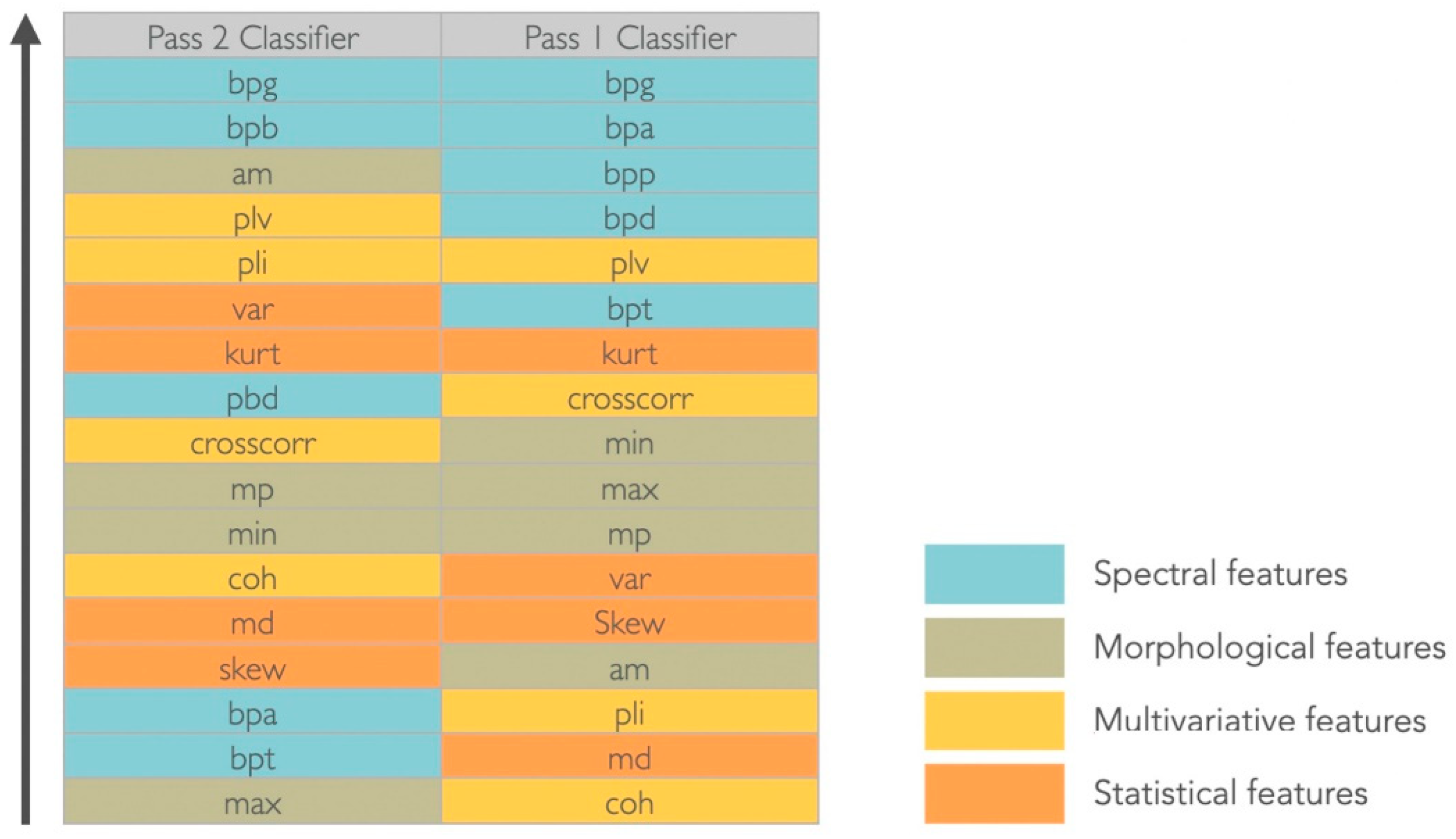
2.5. Feature Extraction
2.6. Feature Reduction
2.7. Classification
2.8. Prediction
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fisher, R.S.; Cross, J.H.; French, J.A.; Higurashi, N.; Hirsch, E.; Jansen, F.E.; Lagae, L.; Moshé, S.L.; Peltola, J.; Roulet Perez, E.; et al. Operational classification of seizure types by the International League Against Epilepsy: Position Paper of the ILAE Commission for Classification and Terminology. Epilepsia 2017, 58, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Kwan, P.; Brodie, M.J. Early identification of refractory epilepsy. N. Engl. J. Med. 2000, 342, 314–319. [Google Scholar] [CrossRef] [PubMed]
- West, S.; Nolan, S.J.; Newton, R. Surgery for epilepsy: A systematic review of current evidence. Epileptic Disord. 2016, 18, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Benbadis, S.R.; Geller, E.; Ryvlin, P.; Schachter, S.; Wheless, J.; Doyle, W.; Vale, F.L. Putting it all together: Options for intractable epilepsy: An updated algorithm on the use of epilepsy surgery and neurostimulation. Epilepsy Behav. 2018, 88, 33–38. [Google Scholar] [CrossRef]
- Mirski, M.A.; Ferrendelli, J.A. Interruption of the mammillothalamic tract prevents seizures in guinea pigs. Science 1984, 226, 72–74. [Google Scholar] [CrossRef]
- Mirski, M.A.; Ferrendelli, J.A. Anterior thalamic mediation of generalized pentylenetetrazol seizures. Brain Res. 1986, 399, 212–223. [Google Scholar] [CrossRef]
- Mirski, M.A.; Rossell, L.A.; Terry, J.B.; Fisher, R.S. Anticonvulsant effect of anterior thalamic high frequency electrical stimulationin the rat. Epilepsy Res. 1997, 28, 89–100. [Google Scholar] [CrossRef]
- Mirski, M.A.; McKeon, A.C.; Ferrendelli, J. Anterior thalamus and substantia nigra: Two distinct structures mediating experimental generalized seizures. Brain Res. 1986, 397, 377–380. [Google Scholar] [CrossRef]
- Gartner, L.P. A Textbook of Neuroanatomy; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Cooper, I.; Upton, A.; Amin, I. Reversibility of chronic neurologic deficits. Some effects of electrical stimulation of the thalamus and internal capsule in man. Stereotact. Funct. Neurosurg. 1980, 43, 244–258. [Google Scholar] [CrossRef]
- Cooper, I.; Upton, A.; Amin, I.; Garnett, S.; Brown, G.; Springman, M. Evoked metabolic responses in the limbic-striate system produced by stimulation of anterior thalamic nucleus in man. Int. J. Neurol. 1984, 18, 179–187. [Google Scholar]
- Upton, A.; Cooper, I.; Springman, M.; Amin, I. Suppression of seizures and psychosis of limbic system origin by chronic stimulation of anterior nucleus of the thalamus. Int. J. Neurol. 1985, 19, 223–230. [Google Scholar] [PubMed]
- Hodaie, M.; Wennberg, R.A.; Dostrovsky, J.O.; Lozano, A.M. Chronic anterior thalamus stimulation for intractable epilepsy. Epilepsia 2002, 43, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Kerrigan, J.F.; Litt, B.; Fisher, R.S.; Cranstoun, S.; French, J.A.; Blum, D.E.; Dichter, M.; Shetter, A.; Baltuch, G.; Jaggi, J.; et al. Electrical stimulation of the anterior nucleus of the thalamus for the treatment of intractable epilepsy. Epilepsia 2004, 45, 346–354. [Google Scholar] [CrossRef]
- Andrade, D.; Zumsteg, D.; Hamani, C.; Hodaie, M.; Sarkissian, S.; Lozano, A.; Wennberg, R. Long-term follow-up of patients with thalamic deep brain stimulation for epilepsy. Neurology 2006, 66, 1571–1573. [Google Scholar] [CrossRef]
- Lee, K.; Jang, K.; Shon, Y. Chronic deep brain stimulation of subthalamic and anterior thalamic nuclei for controlling refractory partial epilepsy. In Advances in Functional and Reparative Neurosurgery; Springer: Berlin/Heidelberg, Germany, 2006; pp. 87–91. [Google Scholar]
- Osorio, I.; Overman, J.; Giftakis, J.; Wilkinson, S.B. High frequency thalamic stimulation for inoperable mesial temporal epilepsy. Epilepsia 2007, 48, 1561–1571. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.J.; Shon, Y.M.; Cho, C.B. Long-term outcome of anterior thalamic nucleus stimulation for intractable epilepsy. Stereotact. Funct. Neurosurg. 2012, 90, 379–385. [Google Scholar] [CrossRef]
- Fisher, R.; Salanova, V.; Witt, T.; Worth, R.; Henry, T.; Gross, R.; Oommen, K.; Osorio, I.; Nazzaro, J.; Labar, D.; et al. Electrical stimulation of the anterior nucleus of thalamus for treatment of refractory epilepsy. Epilepsia 2010, 51, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Salanova, V.; Witt, T.; Worth, R.; Henry, T.R.; Gross, R.E.; Nazzaro, J.M.; Labar, D.; Sperling, M.R.; Sharan, A.; Sandok, E.; et al. Long-term efficacy and safety of thalamic stimulation for drug-resistant partial epilepsy. Neurology 2015, 84, 1017–1025. [Google Scholar] [CrossRef]
- Lehtimäki, K.; Möttönen, T.; Järventausta, K.; Katisko, J.; Tähtinen, T.; Haapasalo, J.; Niskakangas, T.; Kiekara, T.; Öhman, J.; Peltola, J. Outcome based definition of the anterior thalamic deep brain stimulation target in refractory epilepsy. Brain Stimul. 2016, 9, 268–275. [Google Scholar] [CrossRef]
- Järvenpää, S.; Lehtimäki, K.; Rainesalo, S.; Möttönen, T.; Peltola, J. Improving the effectiveness of ANT DBS therapy for epilepsy with optimal current targeting. Epilepsia Open 2020, 5, 406–417. [Google Scholar] [CrossRef]
- Buentjen, L.; Kopitzki, K.; Schmitt, F.C.; Voges, J.; Tempelmann, C.; Kaufmann, J.; Kanowski, M. Direct targeting of the thalamicanteroventral nucleus for deep brain stimulation by T1-weighted magnetic resonance imaging at 3 T. Stereotact. Funct. Neurosurg. 2014, 92, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Möttönen, T.; Katisko, J.; Haapasalo, J.; Tähtinen, T.; Kiekara, T.; Kähärä, V.; Peltola, J.; Öhman, J.; Lehtimäki, K. Defining the anterior nucleus of the thalamus (ANT) as a deep brain stimulation target in refractory epilepsy: Delineation using 3 T MRI and intraoperative microelectrode recording. NeuroImage Clin. 2015, 7, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Hodaie, M.; Cordella, R.; Lozano, A.M.; Wennberg, R.; Dostrovsky, J.O. Bursting activity of neurons in the human anterior thalamic nucleus. Brain Res. 2006, 1115, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ohara, S.; Taghva, A.; Kim, J.; Lenz, F. Spontaneous low threshold spike bursting in awake humans is different in different lateral thalamic nuclei. Exp. Brain Res. 2007, 180, 281–288. [Google Scholar] [CrossRef]
- Schaper, F.L.; Zhao, Y.; Janssen, M.L.; Wagner, G.L.; Colon, A.J.; Hilkman, D.M.; Gommer, E.; Vlooswijk, M.C.; Hoogland, G.; Ackermans, L.; et al. Single-cell recordings to target the anterior nucleus of the thalamus in deep brain stimulation for patients with refractory epilepsy. Int. J. Neural Syst. 2019, 29, 1850012. [Google Scholar] [CrossRef] [PubMed]
- Möttönen, T.; Katisko, J.; Haapasalo, J.; Tähtinen, T.; Saastamoinen, A.; Peltola, J.; Öhman, J.; Lehtimäki, K. The correlation between intraoperative microelectrode recording and 3-Tesla MRI in patients undergoing ANT-DBS for refractory epilepsy. Stereotact. Funct. Neurosurg. 2016, 94, 86–92. [Google Scholar] [CrossRef]
- Levy, R.; Ashby, P.; Hutchison, W.D.; Lang, A.E.; Lozano, A.M.; Dostrovsky, J.O. Dependence of subthalamic nucleus oscillations on movement and dopamine in Parkinson’s disease. Brain 2002, 125, 1196–1209. [Google Scholar] [CrossRef]
- Priori, A.; Foffani, G.; Pesenti, A.; Tamma, F.; Bianchi, A.M.; Pellegrini, M.; Locatelli, M.; Moxon, K.; Villani, R. Rhythm-specific pharmacological modulation of subthalamic activity in Parkinson’s disease. Exp. Neurol. 2004, 189, 369–379. [Google Scholar] [CrossRef]
- Brown, P.; Williams, D. Basal ganglia local field potential activity: Character and functional significance in the human. Clin. Neurophysiol. 2005, 116, 2510–2519. [Google Scholar] [CrossRef]
- Kühn, A.A.; Trottenberg, T.; Kivi, A.; Kupsch, A.; Schneider, G.H.; Brown, P. The relationship between local field potential and neuronal discharge in the subthalamic nucleus of patients with Parkinson’s disease. Exp. Neurol. 2005, 194, 212–220. [Google Scholar] [CrossRef]
- Zaidel, A.; Spivak, A.; Grieb, B.; Bergman, H.; Israel, Z. Subthalamic span of β oscillations predicts deep brain stimulation efficacyfor patients with Parkinson’s disease. Brain 2010, 133, 2007–2021. [Google Scholar] [CrossRef] [PubMed]
- Marceglia, S.; Fumagalli, M.; Priori, A. What neurophysiological recordings tell us about cognitive and behavioral functions of the human subthalamic nucleus. Expert Rev. Neurother. 2011, 11, 139–149. [Google Scholar] [CrossRef]
- Rosa, M.; Marceglia, S.; Barbieri, S.; Priori, A. Local Field Potential and Deep Brain Stimulation (DBS). In Encyclopedia of Computational Neuroscience; Springer: New York, NY, USA, 2014. [Google Scholar]
- Kühn, A.A.; Kupsch, A.; Schneider, G.H.; Brown, P. Reduction in subthalamic 8–35 Hz oscillatory activity correlates with clinicalimprovement in Parkinson’s disease. Eur. J. Neurosci. 2006, 23, 1956–1960. [Google Scholar] [CrossRef]
- Mann, J.M.; Foote, K.D.; Garvan, C.W.; Fernandez, H.H.; Jacobson, C.E.; Rodriguez, R.L.; Haq, I.; Siddiqui, M.S.; Malaty, I.A.; Morishita, T.; et al. Brain penetration effects of microelectrodes and DBS leads in STN or GPi. J. Neurol. Neurosurg. Psychiatry 2009, 80, 794–798. [Google Scholar] [CrossRef] [PubMed]
- Constantoyannis, C.; Berk, C.; Honey, C.R.; Mendez, I.; Brownstone, R.M. Reducing hardware-related complications of deep brain stimulation. Can. J. Neurol. Sci. 2005, 32, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Quinn, E.J.; Blumenfeld, Z.; Velisar, A.; Koop, M.M.; Shreve, L.A.; Trager, M.H.; Hill, B.C.; Kilbane, C.; Henderson, J.M.; Brontë- Stewart, H. Beta oscillations in freely moving Parkinson’s subjects are attenuated during deep brain stimulation. Mov. Disord. 2015, 30, 1750–1758. [Google Scholar] [CrossRef]
- Blumenfeld, Z.; Koop, M.M.; Prieto, T.E.; Shreve, L.A.; Velisar, A.; Quinn, E.J.; Trager, M.H.; Brontë-Stewart, H. Sixty-hertz stimulation improves bradykinesia and amplifies subthalamic low-frequency oscillations. Mov. Disord. 2017, 32, 80–88. [Google Scholar] [CrossRef]
- Neumann, W.J.; Staub-Bartelt, F.; Horn, A.; Schanda, J.; Schneider, G.H.; Brown, P.; Kühn, A.A. Long term correlation of subthalamic beta band activity with motor impairment in patients with Parkinson’s disease. Clin. Neurophysiol. 2017, 128, 2286–2291. [Google Scholar] [CrossRef]
- Syrkin-Nikolau, J.; Koop, M.M.; Prieto, T.; Anidi, C.; Afzal, M.F.; Velisar, A.; Blumenfeld, Z.; Martin, T.; Trager, M.; Bronte-Stewart, H. Subthalamic neural entropy is a feature of freezing of gait in freely moving people with Parkinson’s disease. Neurobiol. Dis. 2017, 108, 288–297. [Google Scholar] [CrossRef]
- Anidi, C.; O’Day, J.J.; Anderson, R.W.; Afzal, M.F.; Syrkin-Nikolau, J.; Velisar, A.; Bronte-Stewart, H.M. Neuromodulation targets pathological not physiological beta bursts during gait in Parkinson’s disease. Neurobiol. Dis. 2018, 120, 107–117. [Google Scholar] [CrossRef]
- Hell, F.; Plate, A.; Mehrkens, J.H.; Bötzel, K. Subthalamic oscillatory activity and connectivity during gait in Parkinson’s disease. NeuroImage: Clin. 2018, 19, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Maling, N.; Lempka, S.F.; Blumenfeld, Z.; Bronte-Stewart, H.; McIntyre, C.C. Biophysical basis of subthalamic local field potentials recorded from deep brain stimulation electrodes. J. Neurophysiol. 2018, 120, 1932–1944. [Google Scholar] [CrossRef] [PubMed]
- Swann, N.C.; de Hemptinne, C.; Thompson, M.C.; Miocinovic, S.; Miller, A.M.; Gilron, R.; Ostrem, J.L.; Chizeck, H.J.; Starr, P.A. Adaptive deep brain stimulation for Parkinson’s disease using motor cortex sensing. J. Neural Eng. 2018, 15, 046006. [Google Scholar] [CrossRef]
- Ramirez Pasos, U.E.; Steigerwald, F.; Reich, M.M.; Matthies, C.; Volkmann, J.; Reese, R. Levodopa modulates functional connectivity in the upper beta band between subthalamic nucleus and muscle activity in tonic and phasic motor activity patterns in Parkinson’s disease. Front. Hum. Neurosci. 2019, 13, 223. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Wang, Q.; Xu, C.; Fang, F.; Fan, J.; Li, L.; Du, Q.; Zhang, R.; Wang, Y.; Lin, Y.; et al. Transcranial direct current stimulation reduces seizure frequency in patients with refractory focal epilepsy: A randomized, double-blind, sham-controlled, and three-arm parallel multicenter study. Brain Stimul. 2020, 13, 109–116. [Google Scholar]
- Goyal, A.; Goetz, S.; Stanslaski, S.; Oh, Y.; Rusheen, A.E.; Klassen, B.; Miller, K.; Blaha, C.D.; Bennet, K.E.; Lee, K. The development of an implantable deep brain stimulation device with simultaneous chronic electrophysiological recording and stimulation in humans. Biosens. Bioelectron. 2021, 176, 112888. [Google Scholar] [CrossRef]
- Jimenez-Shahed, J. Device profile of the percept PC deep brain stimulation system for the treatment of Parkinson’s disease and related disorders. Expert Rev. Med. Devices 2021, 18, 319–332. [Google Scholar] [CrossRef]
- Osorio, I.; Zaveri, H.P.; Frei, M.G.; Arthurs, S. Epilepsy: The Intersection of Neurosciences, Biology, Mathematics, Engineering, and Physics; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Zaveri, H.P.; Duckrow, R.B.; Spencer, S.S. On the use of bipolar montages for time-series analysis of intracranial electroencephalograms. Clin. Neurophysiol. 2006, 117, 2102–2108. [Google Scholar] [CrossRef]
- Bastos, A.M.; Schoffelen, J.M. A tutorial review of functional connectivity analysis methods and their interpretational pitfalls. Front. Syst. Neurosci. 2016, 9, 175. [Google Scholar]
- Georgiy, R.; John, B.; Martha, J.; Richard, N. Patient-specific early seizure detection from scalp electroencephalogram. J. Clin. Neurophysiol. 2010, 27, 163–178. [Google Scholar]
- Wold, S.; Esbensen, K.; Geladi, P. Principal component analysis. Chemom. Intell. Lab. Syst. 1987, 2, 37–52. [Google Scholar]
- Machine Learning with Matlab. Available online: https://www.mathworks.com/matlabcentral/fileexchange/42744-machine-learning-with-matlab (accessed on 2 June 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Description | Neurostimulator | Anatomical Target | Number of Patients |
|---|---|---|---|---|
| Jimenz-Shahed et al. (2021) [50] | Description of the sensing capabilities of the neurostimulator | Percept PC | x | x |
| Goyal et al. (2021) [49] | Description of the sensing capabilities of the neurostimulator | Percept PC | x | x |
| Yang et al. (2020) [48] | Study of which stimulation parameters provide pain relief without triggering after discharges | Activa PC+S | ACC | 3 |
| Passos et al. (2019) [47] | Study the impact of dopaminergic state and movement of beta functional connectivity between basal ganglia and lower motor neurons | Activa PC+S | STN | 8 |
| Anidi et al. (2019) [43] | Study whether STN-DBS affects beta burst dynamics and gait impairments differentially in freezers and non-freezers | Activa PC+S | STN | 12 |
| Hell et al. (2018) [44] | Study of the STN function during gait | Activa PC+S | STN | 10 |
| Maling et al. (2018) [45] | Development of a patient-specific computational framework to analyze LFP recordings | Activa PC+S | STN | 1 |
| Swann et al. (2018) [46] | Study of the disfunction of disturbed neural networks in FLPs | Activa PC+S | STN | 5 |
| Neumann et al. (2017) [41] | Study of long-term association of STN beta activity with parkinsonian motor signs | Activa PC+S | STN | 15 |
| Syrkin-Nikolau et al. (2017) [42] | Study of STN neural features of freezers and non-freezers | Activa PC+S | STN | 14 |
| Blumenfeld et al. (2016) [40] | Study whether the STN alpha/beta oscillation attenuation is causal to the bradykinesia improvement | Activa PC+S | STN | 9 |
| Quinn et al. (2015) [39] | Study whether beta power is similar in different resting postures during forward walking | Activa PC+S | STN | 15 |
| Contacts | Physical Location | Montage | Target % (LH) | Target % (RH) |
|---|---|---|---|---|
| 0–1 | A | Pass2 | − | + |
| 0–2 | B | Pass1 | − | + |
| 1–2 | CPass2 | Pass2 | − | ++ |
| 0–3 | CPass1 | Pass1 | − | ++ |
| 1–3 | D | Pass1 | + | +++ |
| 2–3 | E | Pass2 | ++ | +++ |
| Feature Type | Feature | Description |
|---|---|---|
| Statistical | Variance (var) | Difference (spread) between the normalized squared sum of instantaneous values with the mean value |
| Skewness (skew) | Distortion or asymmetry of the probability density function of the amplitude of time-series | |
| Kurtosis (kurt) | Sharpness of the probability density function of the amplitude of time-series | |
| Median (md) | Value separating the higher half from the lower half of a data sample | |
| Spectral | Band power delta (bpd) | Power spectral density in the delta frequency band (1–3 Hz) |
| Band power theta (pbt) | Power spectral density in the theta frequency band (4–7 Hz) | |
| Band power alpha (pba) | Power spectral density in the alpha frequency band (8–12 Hz) | |
| Band power beta (bpb) | Power spectral density in the beta frequency band (13–30 Hz) | |
| Band power gamma (bpg) | Power spectral density in the gamma frequency band (3–100 Hz) | |
| Morphological | Absolute mean (am) | Absolute average value of a data sample |
| Mean peaks (mp) | Absolute average of the maximum values of a data sample | |
| Maximum (max) | Maximum value of a data sample | |
| Minimum (min) | Minimum value of a data sample | |
| Multivariate | Cross-correlation (crosscorr) | Measure of similarity of two time-series |
| Coherence (coh) | Measures the causal relationship between two signals | |
| Phase locking value (plv) | Measure of the phase synchrony between two time-series | |
| Phase lag index (pli) | Evaluates the phase difference distribution across observations |
| Classifier | Model Parameters | Standard |
|---|---|---|
| SVM | Kernel function: linear, radial, basis function (rbf), polynomial | rbf |
| kNN | Distance metrics: squared Euclidean (seuclidean), Euclidean, correlation, Spearman | seuclidean |
| NN | Hidden layer size (1:10) | 5 |
| Model | Spatial Locations to Predict |
|---|---|
| Pass1 | CPass1-Left, CPass1-Right, BRight, DLeft, ALeft and ERight |
| Pass2 | CPass2-Left, CPass2-Right, ARight, ELeft, BLeft and DRight |
| Spatial Location | Target (LH) | Predictions (LH) | Target (RH) | Predictions (RH) |
|---|---|---|---|---|
| A | − | 54.35% | + | 50.00% |
| B | − | 63.04% | + | 56.52% |
| CPass1 | − | 41.30% | ++ | 52.17% |
| CPass2 | - | 52.17% | ++ | 54.35% |
| D | + | 40.87% | +++ | 60.87% |
| E | ++ | 69.57% | +++ | 60.87% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopes, E.M.; Rego, R.; Rito, M.; Chamadoira, C.; Dias, D.; Cunha, J.P.S. Estimation of ANT-DBS Electrodes on Target Positioning Based on a New PerceptTM PC LFP Signal Analysis. Sensors 2022, 22, 6601. https://doi.org/10.3390/s22176601
Lopes EM, Rego R, Rito M, Chamadoira C, Dias D, Cunha JPS. Estimation of ANT-DBS Electrodes on Target Positioning Based on a New PerceptTM PC LFP Signal Analysis. Sensors. 2022; 22(17):6601. https://doi.org/10.3390/s22176601
Chicago/Turabian StyleLopes, Elodie Múrias, Ricardo Rego, Manuel Rito, Clara Chamadoira, Duarte Dias, and João Paulo Silva Cunha. 2022. "Estimation of ANT-DBS Electrodes on Target Positioning Based on a New PerceptTM PC LFP Signal Analysis" Sensors 22, no. 17: 6601. https://doi.org/10.3390/s22176601
APA StyleLopes, E. M., Rego, R., Rito, M., Chamadoira, C., Dias, D., & Cunha, J. P. S. (2022). Estimation of ANT-DBS Electrodes on Target Positioning Based on a New PerceptTM PC LFP Signal Analysis. Sensors, 22(17), 6601. https://doi.org/10.3390/s22176601