
Dielectric Dispersion Modulated Sensing of Yeast Suspension Electroporation


Abstract
:1. Introduction
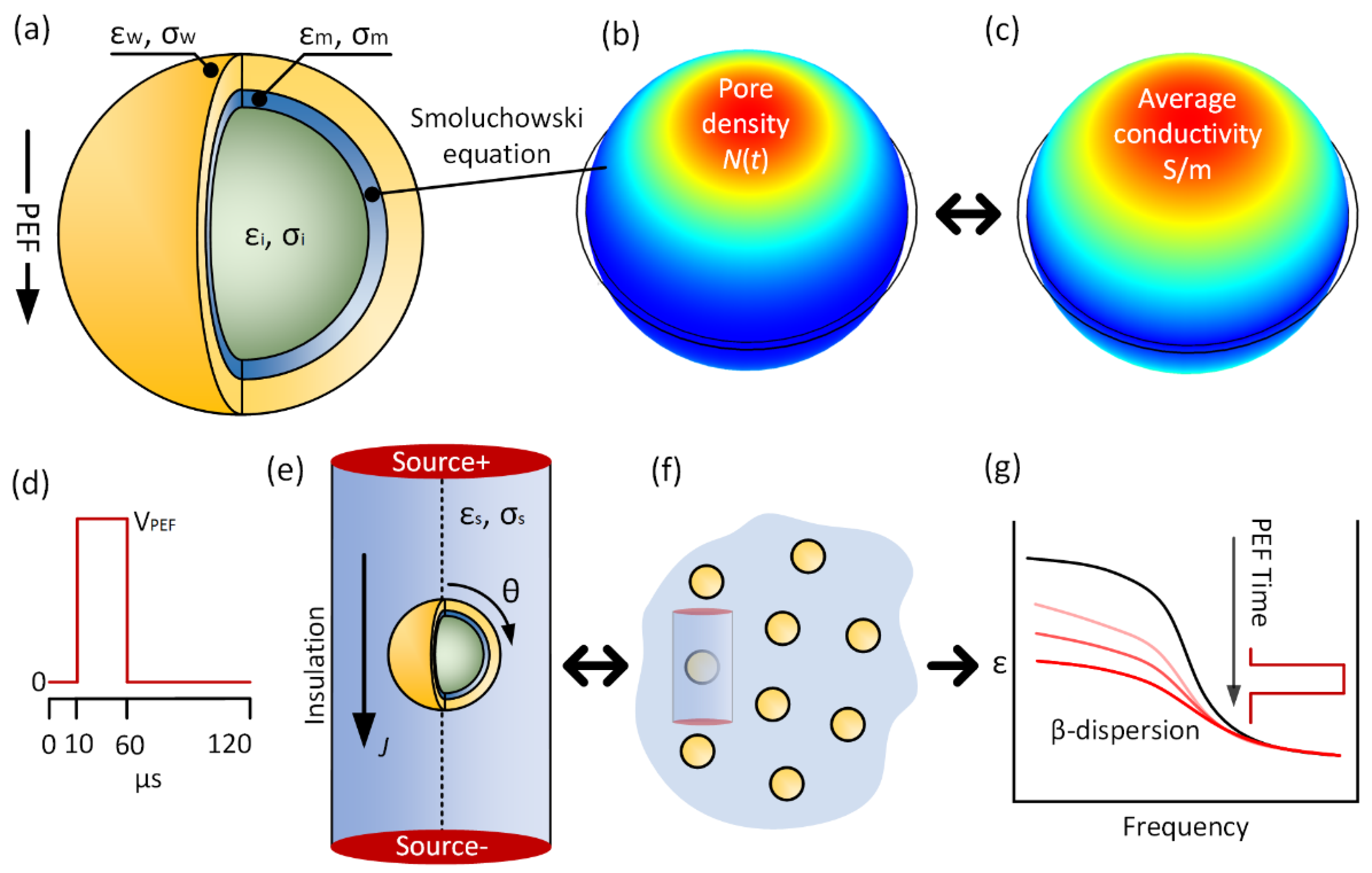
2. Materials and Methods
2.1. Numerical Modeling
2.2. Yeast Cell Passive Properties
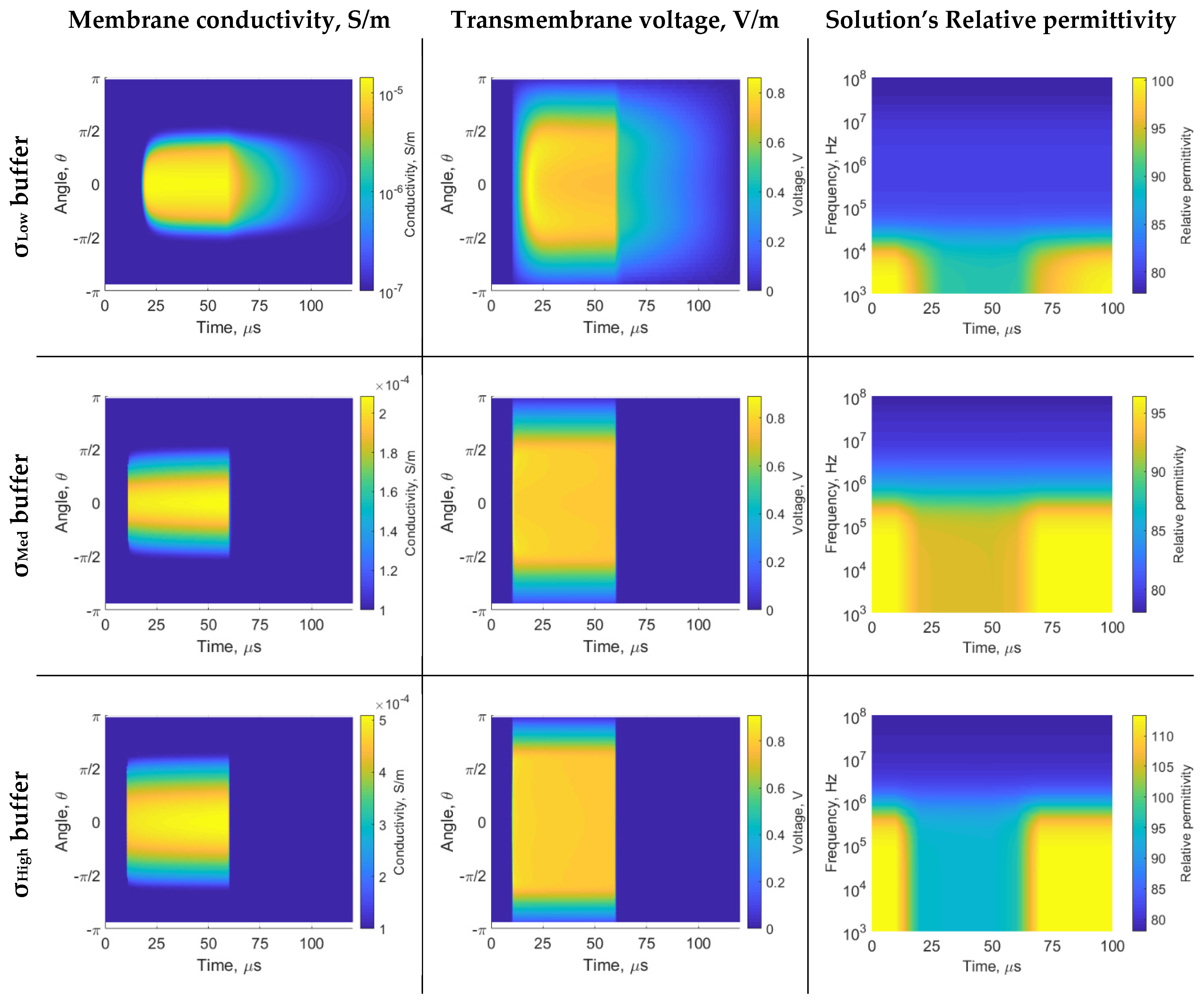
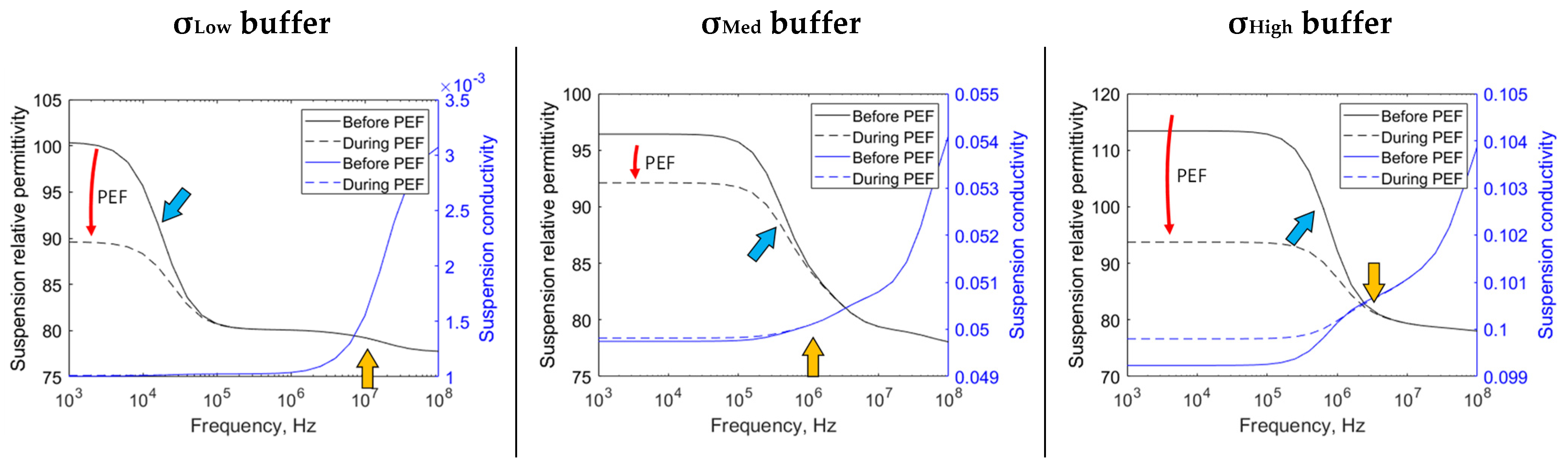
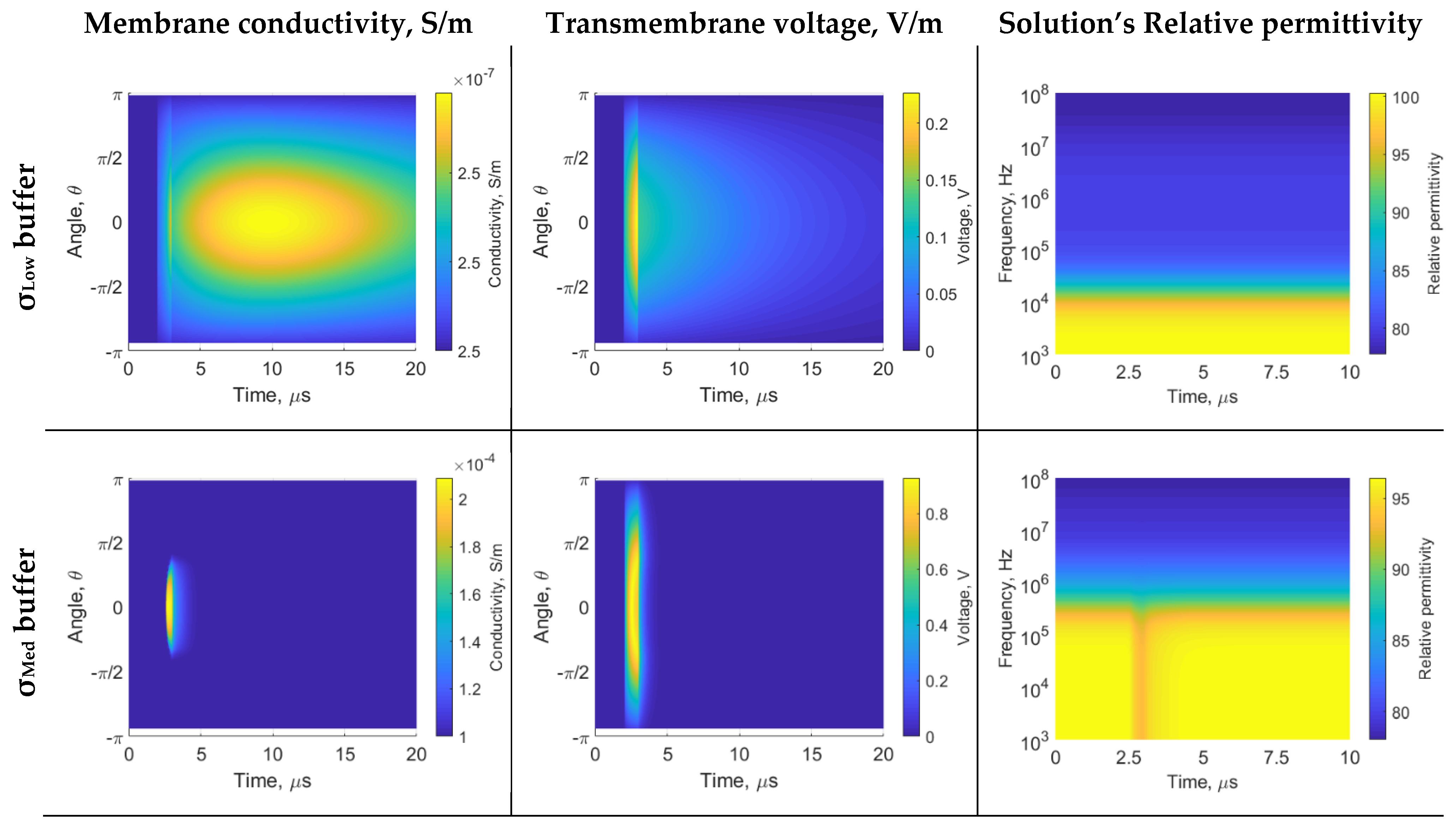
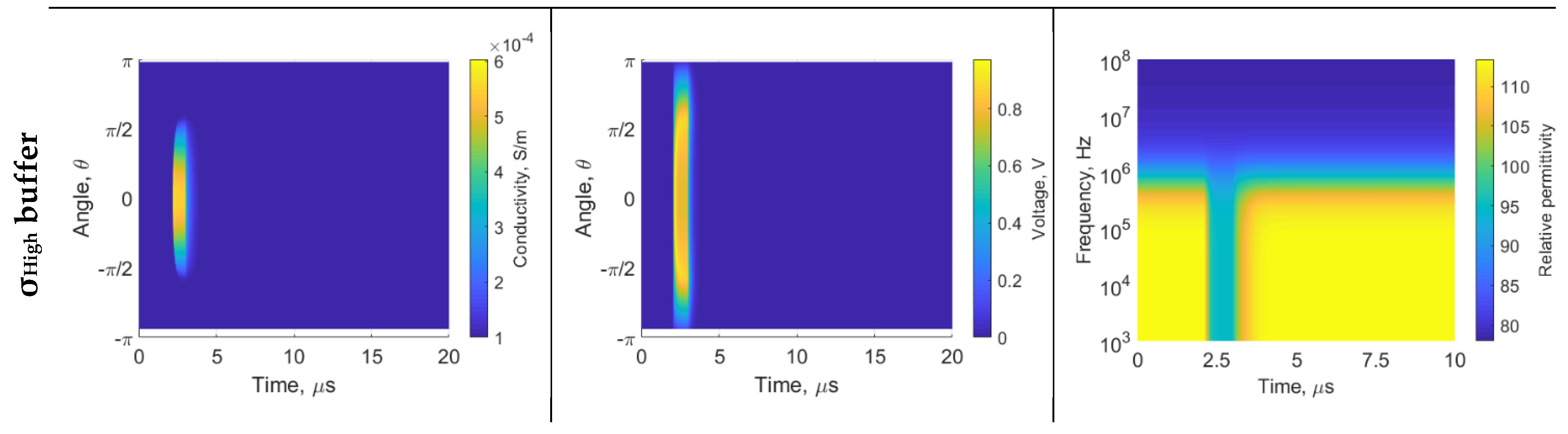
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Symbol | Value | References |
|---|---|---|---|
| External conductivity | σe | See Table 2 | - |
| External permittivity | εe | See Table 2 | - |
| Membrane permittivity | εm | See Table 2 | - |
| Wall membrane conductivity | σw | See Table 2 | - |
| Wall membrane permittivity | εw | See Table 2 | - |
| Cytoplasm conductivity | σi | See Table 2 | - |
| Cytoplasm permittivity | εi | See Table 2 | - |
| Cell radius | R | 5 μm | [30] |
| Thickness membrane | hm | 8 nm | [30,57] |
| Thickness wall | hw | 220 nm | [27,30] |
| Electroporation constant | q | 2.46 | [54,55] |
| Electroporation constant | α | 109 m2s−1 | [42,54,55] |
| Pore density at Vm = 0 V | N0 | 1.5 × 109 m−2 | [42,54,55] |
| Electroporation voltage | Vep | 258 mV | [42,54,55] |
| Pore energy barrier | W0 | 2.65 | [42,54,55] |
| Relative length of the pore | η | 0.15 nm | [42,54,55] |
| Pore radius | rp | 0.8 nm | [42,54,55] |
| Boltzmann constant | k | 1.38 × 10−23 m2kgs−2K−1 | [42,54,55] |
| Temperature | T | 295 K | [42,54,55] |
References
- Ozyigit, I.I. Gene transfer to plants by electroporation: Methods and applications. Mol. Biol. Rep. 2020, 47, 3195–3210. [Google Scholar] [CrossRef]
- Rangel, M.M.M.; Luz, J.C.S.; Oliveira, K.D.; Ojeda, J.; Freytag, J.O.; Suzuki, D.O. Electrochemotherapy in the treatment of neoplasms in dogs and cats. Austral J. Vet. Sci. 2019, 51, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Geboers, B.; Scheffer, H.J.; Graybill, P.M.; Ruarus, A.H.; Nieuwenhuizen, S.; Puijk, R.S.; van den Tol, P.M.; Davalos, R.V.; Rubinsky, B.; de Gruijl, T.D.; et al. High-Voltage Electrical Pulses in Oncology: Irreversible Electroporation, Electrochemotherapy, Gene Electrotransfer, Electrofusion, and Electroimmunotherapy. Radiology 2020, 295, 254–272. [Google Scholar] [CrossRef] [PubMed]
- Pintarelli, G.B.; Ramos, C.T.S.; Da Silva, J.R.; Rossi, M.J.; Suzuki, D.O.H. Sensing of Yeast Inactivation by Electroporation. IEEE Sens. J. 2021, 21, 12027–12035. [Google Scholar] [CrossRef]
- Reineke, K.; Schottroff, F.; Meneses, N.; Knorr, D. Sterilization of liquid foods by pulsed electric fields-an innovative ultra-high temperature process. Front. Microbiol. 2015, 6, 400. [Google Scholar] [CrossRef] [Green Version]
- Ghorbel, A.; Mir, L.M.; García-Sánchez, T. Conductive nanoparticles improve cell electropermeabilization. Nanotechnology 2019, 30, 495101. [Google Scholar] [CrossRef]
- Pintarelli, G.B.; Berkenbrock, J.A.; Rassele, A.; Rangel, M.M.M.; Suzuki, D.O.H. Computer simulation of commercial conductive gels and their application to increase the safety of electrochemotherapy treatment. Med. Eng. Phys. 2019, 74, 99–105. [Google Scholar] [CrossRef]
- Lopes, L.B.; Pintarelli, G.B.; dos Santos, C.S.F.; Suzuki, D.O.H. Computer optimization of conductive gels for electrochemotherapy. Med. Eng. Phys. 2021, 98, 133–139. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, Z.; Xia, X.; Lin, Y. A novel electroporation system for living cell staining and membrane dynamics interrogation. Micromachines 2020, 11, 767. [Google Scholar] [CrossRef]
- Bazzolo, B.; Sieni, E.; Zamuner, A.; Roso, M.; Russo, T.; Gloria, A.; Dettin, M.; Conconi, M.T. Breast cancer cell cultures on electrospun poly(ε-caprolactone) as a potential tool for preclinical studies on anticancer treatments. Bioengineering 2021, 8, 1. [Google Scholar] [CrossRef]
- Kotnik, T.; Kramar, P.; Pucihar, G.; Miklavcic, D.; Tarek, M. Cell membrane electroporation- Part 1: The phenomenon. IEEE Electr. Insul. Mag. 2012, 28, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Haberl, S.; Miklavcic, D.; Sersa, G.; Frey, W.; Rubinsky, B. Cell membrane electroporation-Part 2: The applications. IEEE Electr. Insul. Mag. 2013, 29, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Weaver, J.C.; Smith, K.C.; Esser, A.T.; Son, R.S.; Gowrishankar, T.R. A brief overview of electroporation pulse strength–duration space: A region where additional intracellular effects are expected. Bioelectrochemistry 2012, 87, 236–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubinsky, L.; Guenther, E.; Mikus, P.; Stehling, M.; Rubinsky, B. Electrolytic Effects During Tissue Ablation by Electroporation. Technol. Cancer Res. Treat. 2016, 15, NP95–NP103. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, J.; Kolb, J.F. Time domain dielectric spectroscopy of nanosecond pulsed electric field induced changes in dielectric properties of pig whole blood. Bioelectrochemistry 2015, 103, 28–33. [Google Scholar] [CrossRef]
- Mescia, L.; Chiapperino, M.A.; Bia, P.; Gielis, J.; Caratelli, D. Modeling of Electroporation Induced by Pulsed Electric Fields in Irregularly Shaped Cells. IEEE Trans. Biomed. Eng. 2018, 65, 414–423. [Google Scholar] [CrossRef]
- Kotnik, T.; Rems, L.; Tarek, M.; Miklavčič, D. Membrane Electroporation and Electropermeabilization: Mechanisms and Models. Annu. Rev. Biophys. 2019, 48, 63–91. [Google Scholar] [CrossRef]
- Zhao, Y.; Bhonsle, S.; Dong, S.; Lv, Y.; Liu, H.; Safaai-Jazi, A.; Davalos, R.V.; Yao, C. Characterization of Conductivity Changes During High-Frequency Irreversible Electroporation for Treatment Planning. IEEE Trans. Biomed. Eng. 2018, 65, 1810–1819. [Google Scholar] [CrossRef]
- Pavlin, M.; Kandušer, M.; Reberšek, M.; Pucihar, G.; Hart, F.X.; Magjarevićcacute, R.; Miklavčič, D. Effect of Cell Electroporation on the Conductivity of a Cell Suspension. Biophys. J. 2005, 88, 4378–4390. [Google Scholar] [CrossRef] [Green Version]
- Schwan, H.P. Electrical Properties of Tissue and Cell Suspensions. In Advances in Biological and Medical Physics; Elsevier: Amsterdam, The Netherlands, 1957; pp. 147–209. [Google Scholar]
- Pauly, H.; Schwan, H.P. Über die Impedanz einer Suspension von kugelförmigen Teilchen mit einer Schale. Zeitschrift für Naturforsch. B 1959, 14, 125–131. [Google Scholar] [CrossRef]
- Weaver, J.; Astumian, R. The response of living cells to very weak electric fields: The thermal noise limit. Science 1990, 247, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Gowrishankar, T.R.; Weaver, J.C. An approach to electrical modeling of single and multiple cells. Proc. Natl. Acad. Sci. USA 2003, 100, 3203–3208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Xie, X.; Duan, Y.; Wang, L.; Cheng, Z.; Cheng, J. A review of impedance measurements of whole cells. Biosens. Bioelectron. 2016, 77, 824–836. [Google Scholar] [CrossRef] [PubMed]
- Bastos, J.P.A.; Sadowski, N. Magnetic Materials and 3D Finite Element Modeling; CRC Press: Boca Raton, FL, USA, 2017; Volume 40, ISBN 9781315215624. [Google Scholar]
- Asami, K.; Yonezawa, T. Dielectric behavior of wild-type yeast and vacuole-deficient mutant over a frequency range of 10 kHz to 10 GHz. Biophys. J. 1996, 71, 2192–2200. [Google Scholar] [CrossRef] [Green Version]
- Hölzel, R.; Lamprecht, I. Dielectric properties of yeast cells as determined by electrorotation. Biochim. Biophys. Acta-Biomembr. 1992, 1104, 195–200. [Google Scholar] [CrossRef]
- Raicu, V.; Raicu, G.; Turcu, G. Dielectric properties of yeast cells as simulated by the two-shell model. Biochim. Biophys. Acta-Bioenerg. 1996, 1274, 143–148. [Google Scholar] [CrossRef] [Green Version]
- El Zakhem, H.; Lanoisellé, J.-L.; Lebovka, N.I.; Nonus, M.; Vorobiev, E. The early stages of Saccharomyces cerevisiae yeast suspensions damage in moderate pulsed electric fields. Colloids Surf. B Biointerfaces 2006, 47, 189–197. [Google Scholar] [CrossRef]
- Lyu, C.; Wang, J.; Powell-Palm, M.; Rubinsky, B. Simultaneous electroporation and dielectrophoresis in non-electrolytic micro/nano-electroporation. Sci. Rep. 2018, 8, 2481. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.A.; de Oliveira, P.X.; Pereira, L.S.; Rodrigues, J.; Suzuki, D.O.H. Sensitivity Analysis of a Nuclear Electroporation Model—A Theoretical Study. IEEE Trans. Dielectr. Electr. Insul. 2021, 28, 1850–1858. [Google Scholar] [CrossRef]
- Suzuki, D.O.H.; Ramos, A.; Ribeiro, M.C.M.; Cazarolli, L.H.; Silva, F.R.M.B.; Leite, L.D.; Marques, J.L.B. Theoretical and Experimental Analysis of Electroporated Membrane Conductance in Cell Suspension. IEEE Trans. Biomed. Eng. 2011, 58, 3310–3318. [Google Scholar] [CrossRef]
- Gimsa, J.; Wachner, D. Analytical Description of the Transmembrane Voltage Induced on Arbitrarily Oriented Ellipsoidal and Cylindrical Cells. Biophys. J. 2001, 81, 1888–1896. [Google Scholar] [CrossRef] [Green Version]
- Saulis, G.; Lapė, R.; Pranevičiūtė, R.; Mickevičius, D. Changes of the solution pH due to exposure by high-voltage electric pulses. Bioelectrochemistry 2005, 67, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Sherba, J.J.; Hogquist, S.; Lin, H.; Shan, J.W.; Shreiber, D.I.; Zahn, J.D. The effects of electroporation buffer composition on cell viability and electro-transfection efficiency. Sci. Rep. 2020, 10, 3053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aouida, M.; Tounekti, O.; Belhadj, O.; Mir, L.M. Comparative roles of the cell wall and cell membrane in limiting uptake of xenobiotic molecules by Saccharomyces cerevisiae. Antimicrob. Agents Chemother. 2003, 47, 2012–2014. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Qian, K.; Zhang, L.; Deng, H.; Li, X.; Zhou, J.; Wang, J. Anisotropic conductivity for single-cell electroporation simulation with tangentially dispersive membrane. Electrochim. Acta 2021, 385, 138426. [Google Scholar] [CrossRef]
- Yan, Z.; Hao, C.; Yin, L.; Liu, K.; Qiu, J. Simulation of the influence of temperature on the dynamic process of electroporation based on finite element analysis. IEEE Trans. Plasma Sci. 2021, 49, 2839–2850. [Google Scholar] [CrossRef]
- Chiapperino, M.A.; Bia, P.; Lamacchia, C.M.; Mescia, L. Electroporation modelling of irregular nucleated cells including pore radius dynamics. Electronics 2019, 8, 1477. [Google Scholar] [CrossRef] [Green Version]
- Schmeer, M.; Seipp, T.; Pliquett, U.; Kakorin, S.; Neumann, E. Mechanism for the conductivity changes caused by membrane electroporation of CHO cell-pellets. Phys. Chem. Chem. Phys. 2004, 6, 5564–5574. [Google Scholar] [CrossRef]
- Kakorin, S.; Neumann, E. Ionic conductivity of electroporated lipid bilayer membranes. Bioelectrochemistry 2002, 56, 163–166. [Google Scholar] [CrossRef]
- Pucihar, G.; Miklavčič, D.; Kotnik, T. A time-dependent numerical model of transmembrane voltage inducement and electroporation of irregularly shaped cells. IEEE Trans. Biomed. Eng. 2009, 56, 1491–1501. [Google Scholar] [CrossRef] [Green Version]
- Ramos, A.; Schneider, A.L.S.; Suzuki, D.O.H.; Marques, J.L.B. Sinusoidal signal analysis of electroporation in biological cells. IEEE Trans. Biomed. Eng. 2012, 59, 2965–2973. [Google Scholar] [CrossRef] [PubMed]
- Arnold, W.M.; Schmutzler, R.K.; Schmutzler, A.G.; van der Ven, H.; Al-Hasani, S.; Krebs, D.; Zimmermann, U. Electro-rotation of mouse oocytes: Single-cell measurements of zona-intact and zona-free cells and of the isolated zona pellucida. Biochim. Biophys. Acta (BBA)-Biomembr. 1987, 905, 454–464. [Google Scholar] [CrossRef]
- Ramos, A.; Suzuki, D.O.H.; Marques, J.L.B. Numerical study of the electrical conductivity and polarization in a suspension of spherical cells. Bioelectrochemistry 2006, 68, 213–217. [Google Scholar] [CrossRef]
- Al-Hilphy, A.R.S. Electrical Field (AC) for Non Thermal Milk Pasteurization. J. Nutr. Food Sci. 2012, 2, 177–181. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Chen, X.; Han, F. Simulation of Cell Dielectric Properties Based on COMSOL. Proc. Int. Conf. Artif. Life Robot. 2018, 23, 603–606. [Google Scholar] [CrossRef]
- Teissié, J.; Rols, M.P. An experimental evaluation of the critical potential difference inducing cell membrane electropermeabilization. Biophys. J. 1993, 65, 409–413. [Google Scholar] [CrossRef] [Green Version]
- Silve, A.; Leray, I.; Poignard, C.; Mir, L.M. Impact of external medium conductivity on cell membrane electropermeabilization by microsecond and nanosecond electric pulses. Sci. Rep. 2016, 6, 19957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muraji, M.; Taniguchi, H.; Tatebe, W.; Berg, H. Examination of the relationship between parameters to determine electropermeability of Saccharomyces cerevisiae. Bioelectrochem. Bioenerg. 1999, 48, 485–488. [Google Scholar] [CrossRef]
- Djuzenova, C.S.; Zimmermann, U.; Frank, H.; Sukhorukov, V.L.; Richter, E.; Fuhr, G. Effect of medium conductivity and composition on the uptake of propidium iodide into electropermeabilized myeloma cells. Biochim. Biophys. Acta-Biomembr. 1996, 1284, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Di Biasio, A.; Ambrosone, L.; Cametti, C. The Dielectric Behavior of Nonspherical Biological Cell Suspensions: An Analytic Approach. Biophys. J. 2010, 99, 163–174. [Google Scholar] [CrossRef] [Green Version]
- Stirke, A.; Celiesiute-Germaniene, R.; Zimkus, A.; Zurauskiene, N.; Simonis, P.; Dervinis, A.; Ramanavicius, A.; Balevicius, S. The link between yeast cell wall porosity and plasma membrane permeability after PEF treatment. Sci. Rep. 2019, 9, 14731. [Google Scholar] [CrossRef] [Green Version]
- Neu, J.C.; Krassowska, W. Asymptotic model of electroporation. Phys. Rev. E 1999, 59, 3471–3482. [Google Scholar] [CrossRef]
- Smith, K.C.; Weaver, J.C. Active Mechanisms Are Needed to Describe Cell Responses to Submicrosecond, Megavolt-per-Meter Pulses: Cell Models for Ultrashort Pulses. Biophys. J. 2008, 95, 1547–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rems, L.; Ušaj, M.; Kandušer, M.; Reberšek, M.; Miklavčič, D.; Pucihar, G. Cell electrofusion using nanosecond electric pulses. Sci. Rep. 2013, 3, 3382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Holzel, R.; Pethig, R.; Wang, X.-B. Differences in the AC electrodynamics of viable and non-viable yeast cells determined through combined dielectrophoresis and electrorotation studies. Phys. Med. Biol. 1992, 37, 1499–1517. [Google Scholar] [CrossRef]





| Symbol | Value |
|---|---|
| Infinitesimal volume cylinder heigh | 30 µm |
| Infinitesimal volume cylinder radius | 16.8 µm |
| Cell wall thickness | 220 ηm 1 |
| Cell membrane thickness | 8 ηm 1 |
| Cell radius | 4 µm 1 |
| Parameter | σLow Buffer | σMed Buffer | σHigh Buffer |
|---|---|---|---|
| External solution conductivity (σs) | 1 × 10−3 [S/m] | 50 × 10−3 [S/m] | 0.1 [S/m] |
| External solution permittivity (ɛs) | 78 | 78 1 | 77 |
| Cytoplasm’s conductivity (σi) | 0.2 [S/m] | 0.55 [S/m] | 0.6 [S/m] |
| Cytoplasm’s relative permittivity (ɛi) | 50 | 50 1 | 58 |
| Initial membrane’s conductivity (σm0) | 0.25 × 10−6 [S/m] | 0.1 × 10−3 [S/m] | 0.1 × 10−3 [S/m] 2 |
| Membrane’s relative permittivity (ɛm) | 6 | 7.6 | 5.2 |
| Cell wall’s conductivity (σw) | 14 × 10−3 [S/m] | 5 × 10−3 [S/m] | 20 × 10−3 [S/m] |
| Cell wall’s relative permittivity (ɛw) | 60 | 60 1 | 60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pintarelli, G.B.; da Silva, J.R.; Yang, W.; Suzuki, D.O.H. Dielectric Dispersion Modulated Sensing of Yeast Suspension Electroporation. Sensors 2022, 22, 1811. https://doi.org/10.3390/s22051811
Pintarelli GB, da Silva JR, Yang W, Suzuki DOH. Dielectric Dispersion Modulated Sensing of Yeast Suspension Electroporation. Sensors. 2022; 22(5):1811. https://doi.org/10.3390/s22051811
Chicago/Turabian StylePintarelli, Guilherme B., Jessica R. da Silva, Wuqiang Yang, and Daniela O. H. Suzuki. 2022. "Dielectric Dispersion Modulated Sensing of Yeast Suspension Electroporation" Sensors 22, no. 5: 1811. https://doi.org/10.3390/s22051811
APA StylePintarelli, G. B., da Silva, J. R., Yang, W., & Suzuki, D. O. H. (2022). Dielectric Dispersion Modulated Sensing of Yeast Suspension Electroporation. Sensors, 22(5), 1811. https://doi.org/10.3390/s22051811