
Detecting Grapevine Virus Infections in Red and White Winegrape Canopies Using Proximal Hyperspectral Sensing



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Plant Virus Testing
2.2. Spectral Data Collection
2.3. Data Processing and Modelling
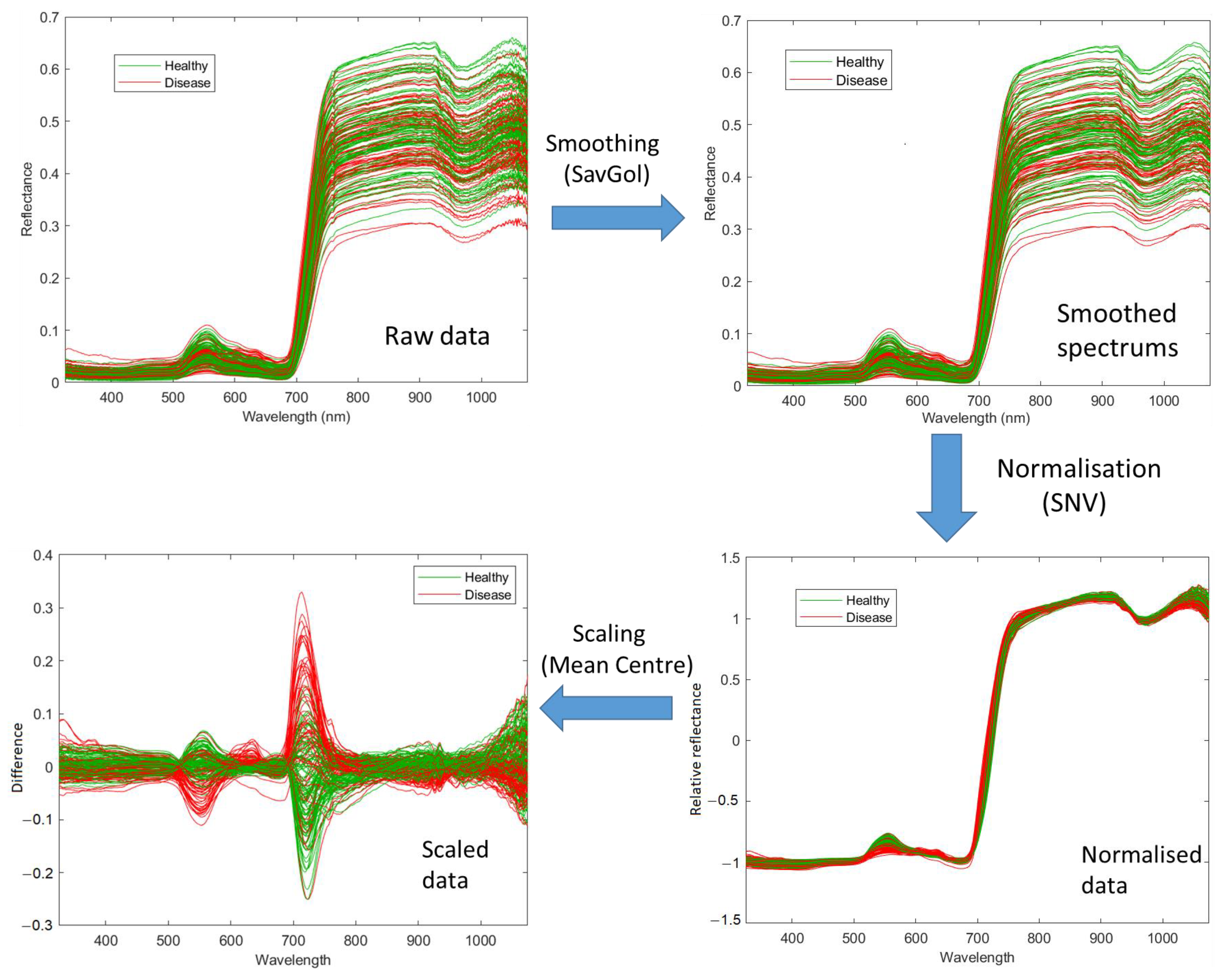
2.3.1. Spectral Data Pre-Processing
2.3.2. Outlier Removal
2.3.3. Cross-Validation
2.3.4. Modelling
3. Results
3.1. Virus Test Results
3.2. Disease Symptomology
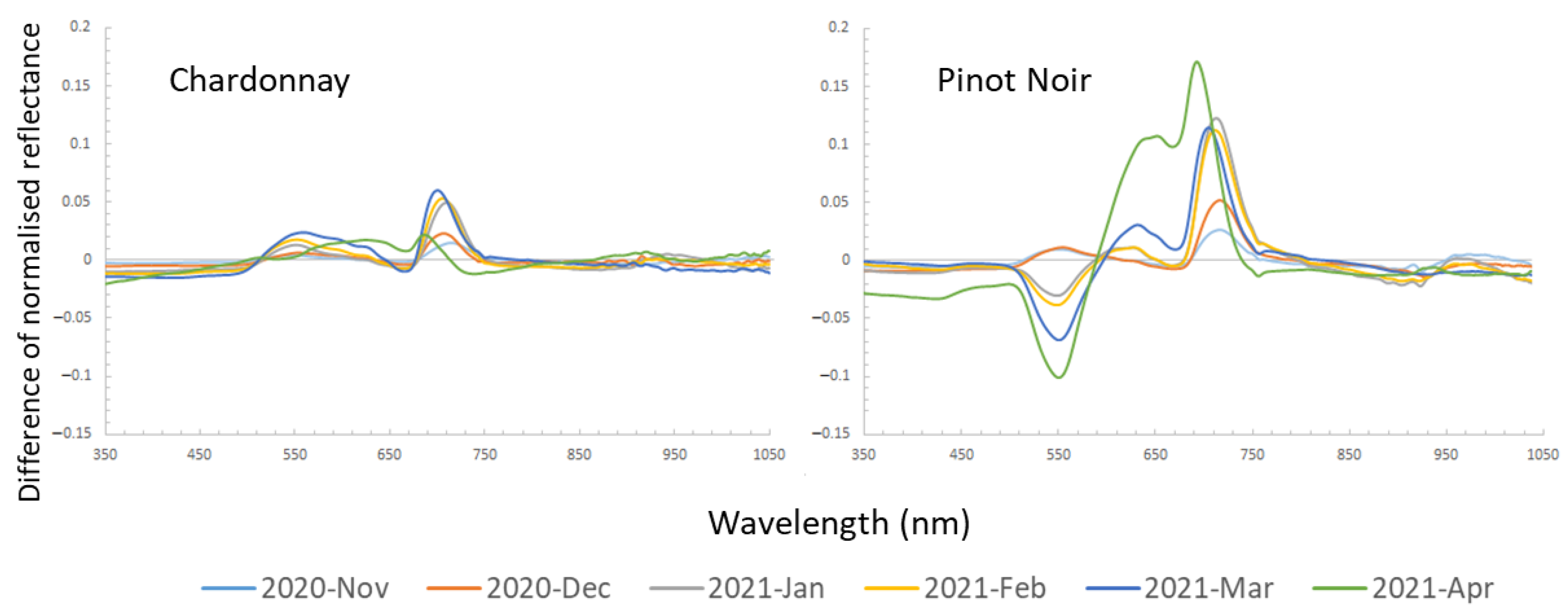
3.3. Critical Spectral Regions for Disease Classification
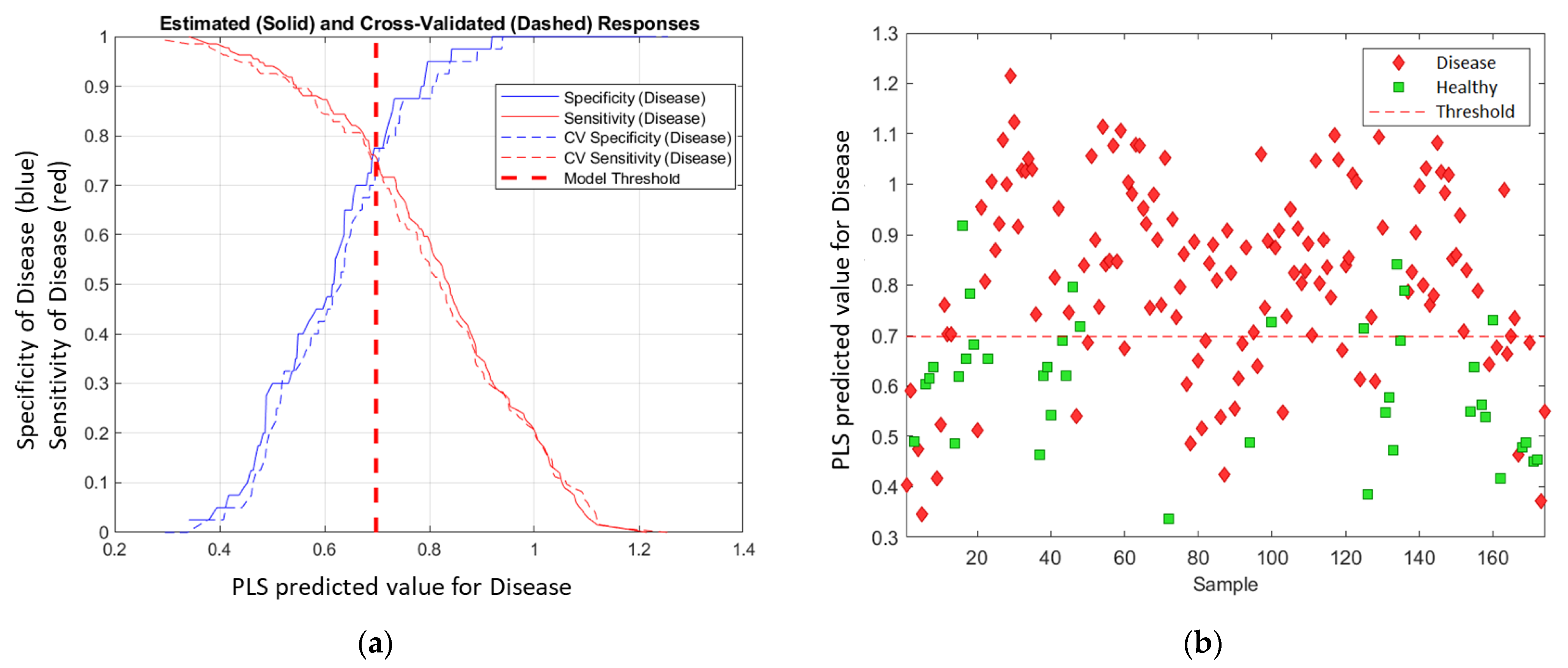
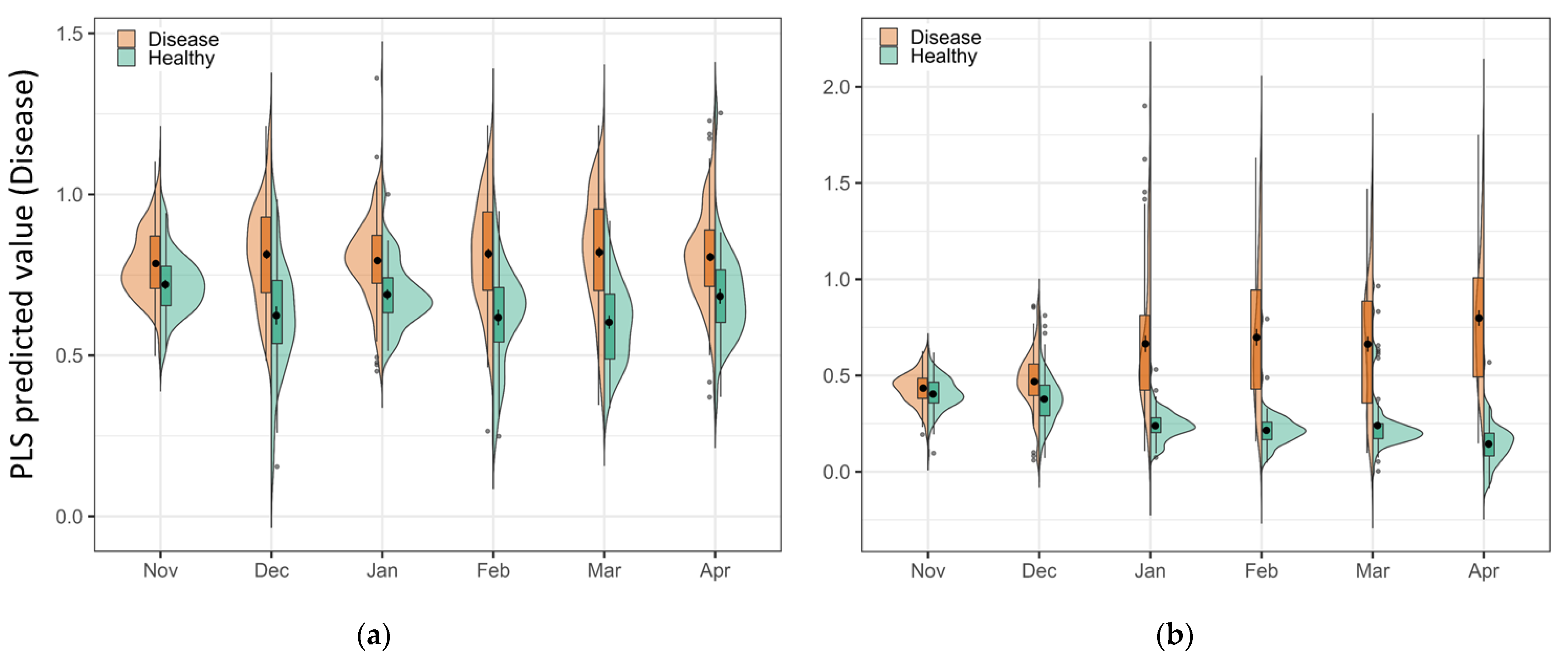
3.4. The Model Results
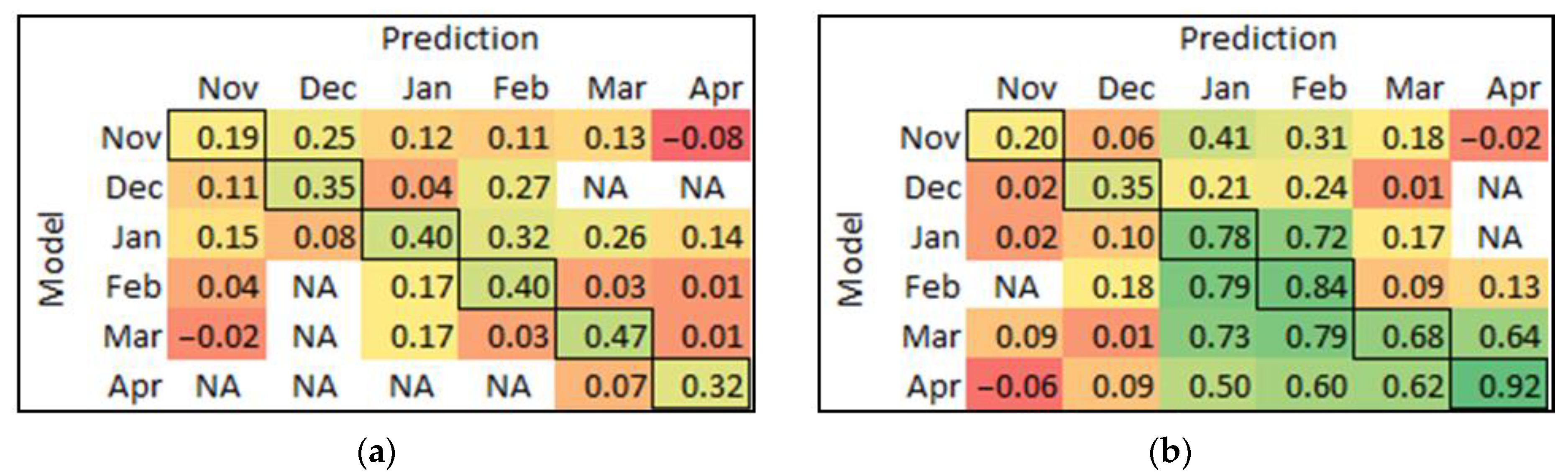
3.5. Model Prediction Matrix
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naidu, R.; Rowhani, A.; Fuchs, M.; Golino, D.; Martelli, G.P. Grapevine leafroll: A complex viral disease affecting a high-value fruit crop. Plant Dis. 2014, 98, 1172–1185. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Keller, K.E.; Rennaker, C.; Martin, R.R. Influence of grapevine leafroll associated viruses (GLRaV-2 and -3) on the fruit composition of Oregon Vitis vinifera L. cv. Pinot noir: Free amino acids, sugars, and organic acids. Food Chem. 2009, 117, 99–105. [Google Scholar] [CrossRef]
- Wolpert, J.A.; Vilas, E.P. Effect of mild leafroll disease on growth, yield, and fruit maturity indices of Riesling and Zinfandel. Am. J. Enol. Vitic. 1992, 43, 367. [Google Scholar] [CrossRef]
- Alabi, O.J.; Casassa, L.F.; Gutha, L.R.; Larsen, R.C.; Henick-Kling, T.; Harbertson, J.F.; Naidu, R.A. Impacts of Grapevine leafroll disease on fruit yield and grape and wine chemistry in a wine grape (Vitis vinifera L.) cultivar. PLoS ONE 2016, 11, e0149666. [Google Scholar] [CrossRef] [Green Version]
- Douglas, N.; Krüger, K. Transmission efficiency of Grapevine leafroll-associated virus 3 (GLRaV-3) by the mealybugs Planococcus ficus and Pseudococcus longispinus (Hemiptera: Pseudococcidae). Eur. J. Plant Pathol. 2008, 122, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Charles, J.; Froud, K.; Van den Brink, R.; Allan, D. Mealybugs and the spread of grapevine leafroll-associated virus 3 (GLRaV-3) in a New Zealand vineyard. Australas. Plant Pathol. 2009, 38, 576–583. [Google Scholar] [CrossRef]
- Atallah, S.; Gomez, M.; Fuchs, M.; Martinson, T. Economic impact of grapevine leafroll disease on Vitis viniferacv. Cabernet franc in Finger Lakes vineyards of New York. Am. J. Enol. Vitic. 2012, 63, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Zorloni, A.; Prati, S.; Bianco, P.A.; Belli, G. Transmission of Grapevine virus A and Grapevine leafroll-associated virus 3 by Heliococcus bohemicus. J. Plant Pathol. 2006, 88, 325–328. [Google Scholar]
- Martelli, G.P.; Ghanem-Sabanadzovic, N.A.; Agranovsky, A.A.; Rwahnih, M.A.; Dolja, V.V.; Dovas, C.I.; Fuchs, M.; Gugerli, P.; Hu, J.S.; Jelkmann, W.; et al. Taxonomic revision of the family Closteroviridae with special reference to the grapevine leafroll-associated members of the genus Ampelovirus and the putative species unassigned to the family. J. Plant Pathol. 2012, 94, 7–19. [Google Scholar]
- Constable, F.E.; Rodoni, B.C. Grapevine Leafroll-Associated Viruses; Wine Australia: Adelaide, Australia, 2014. [Google Scholar]
- Fortusini, A.; Scattini, G.; Prati, S.; Cinquanta, S.; Belli, G. Transmission of grapevine leafroll virus 1 (GLRaV-1) and grapevine virus A (GVA) by scale insects. In Proceedings of the 12th Meeting of ICVG, Lisbon, Portugal, 29 September 1997; pp. 121–122. [Google Scholar]
- Wu, Q.; Habili, N.; Constable, F.; Al Rwahnih, M.A.; Goszczynski, D.E.; Wang, Y.; Pagay, V. Virus pathogens in Australian vineyards with an emphasis on Shiraz Disease. Viruses 2020, 12, 818. [Google Scholar] [CrossRef]
- Bell, V.A.; Lester, P.J.; Pietersen, G.; Hall, A.J. The management and financial implications of variable responses to grapevine leafroll disease. J. Plant Pathol. 2021, 103, 5–15. [Google Scholar] [CrossRef]
- Rubio, L.; Galipienso, L.; Ferriol, I. Detection of plant viruses and disease management: Relevance of genetic diversity and evolution. Front. Plant Sci. 2020, 11, 1092. [Google Scholar] [CrossRef]
- Maree, H.J.; Almeida, R.P.; Bester, R.; Chooi, K.M.; Cohen, D.; Dolja, V.V.; Fuchs, M.F.; Golino, D.A.; Jooste, A.E.; Martelli, G.P.; et al. Grapevine leafroll-associated virus 3. Front. Microbiol. 2013, 4, 82. [Google Scholar] [CrossRef] [Green Version]
- Bell, V.A.; Blouin, A.G.; Cohen, D.; Hedderley, D.I.; Oosthuizen, T.; Spreeth, N.; Lester, P.J.; Pietersen, G. Visual symptom identification of grapevine leafroll-associated virus 3 in red berry cultivars supports virus management by roguing. J. Plant Pathol. 2017, 99, 477–482. [Google Scholar]
- Legrand, P. Biological assays for plant viruses and other graft-transmissible pathogens diagnoses: A review. EPPO Bull. 2015, 45, 240–251. [Google Scholar] [CrossRef]
- Thomas, S.; Kuska, M.T.; Bohnenkamp, D.; Brugger, A.; Alisaac, E.; Wahabzada, M.; Behmann, J.; Mahlein, A.-K. Benefits of hyperspectral imaging for plant disease detection and plant protection: A technical perspective. J. Plant Dis. Prot. 2018, 125, 5–20. [Google Scholar] [CrossRef]
- Naidu, R.A.; Perry, E.M.; Pierce, F.J.; Mekuria, T. The potential of spectral reflectance technique for the detection of Grapevine leafroll-associated virus-3 in two red-berried wine grape cultivars. Comput. Electron. Agric. 2009, 66, 38–45. [Google Scholar] [CrossRef]
- Sinha, R.; Khot, L.R.; Rathnayake, A.P.; Gao, Z.; Naidu, R.A. Visible-near infrared spectroradiometry-based detection of grapevine leafroll-associated virus 3 in a red-fruited wine grape cultivar. Comput. Electron. Agric. 2019, 162, 165–173. [Google Scholar] [CrossRef]
- Pagay, V.; Habili, N.; Wu, Q.; Coleman, D. Rapid and non-destructive detection of Shiraz disease and grapevine leafroll disease on asymptomatic grapevines in Australian vineyards. In Proceedings of the 19th Congress of the International Council for the study of Virus and Virus-like Diseases of Grapevine, Santiago, Chile, 9–12 April 2018. [Google Scholar]
- Junges, A.H.; Almança, M.A.K.; Fajardo, T.V.M.; Ducati, J.R. Leaf hyperspectral reflectance as a potential tool to detect diseases associated with vineyard decline. Trop. Plant Pathol. 2020, 45, 522–533. [Google Scholar] [CrossRef]
- Gao, Z.; Khot, L.R.; Naidu, R.A.; Zhang, Q. Early detection of grapevine leafroll disease in a red-berried wine grape cultivar using hyperspectral imaging. Comput. Electron. Agric. 2020, 179, 105807. [Google Scholar] [CrossRef]
- Fahey, T.; Pham, H.; Gardi, A.; Sabatini, R.; Stefanelli, D.; Goodwin, I.; Lamb, D.W. Active and passive electro-optical sensors for health assessment in food crops. Sensors 2021, 21, 171. [Google Scholar] [CrossRef]
- Bendel, N.; Kicherer, A.; Backhaus, A.; Köckerling, J.; Maixner, M.; Bleser, E.; Klück, H.-C.; Seiffert, U.; Voegele, R.T.; Töpfer, R. Detection of Grapevine leafroll-associated virus 1 and 3 in white and red grapevine cultivars using hyperspectral imaging. Remote Sens. 2020, 12, 1693. [Google Scholar] [CrossRef]
- Pinheiro, F.; Gusmo dos Anjos, W.d.P. Optical Sensors Applied in Agricultural Crops. In Optical Sensors—New Developments and Practical Applications; InTech: Rang-Du-Fliers, France, 2014. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, C.; Sagan, V.; Maimaitiyiming, M.; Maimaitijiang, M.; Bhadra, S.; Kwasniewski, M.T. Early detection of plant viral disease using hyperspectral imaging and deep learning. Sensors 2021, 21, 742. [Google Scholar] [CrossRef] [PubMed]
- Lowe, A.; Harrison, N.; French, A.P. Hyperspectral image analysis techniques for the detection and classification of the early onset of plant disease and stress. Plant Methods 2017, 13, 80. [Google Scholar] [CrossRef]
- Wang, Y.M.; Ostendorf, B.; Gautam, D.; Habili, N.; Pagay, V. Plant viral disease detection: From molecular diagnosis to optical sensing technology—A multidisciplinary review. Remote Sens. 2022, 14, 1542. [Google Scholar] [CrossRef]
- Mendez, K.M.; Reinke, S.N.; Broadhurst, D.I. A comparative evaluation of the generalised predictive ability of eight machine learning algorithms across ten clinical metabolomics data sets for binary classification. Metabolomics 2019, 15, 150. [Google Scholar] [CrossRef] [Green Version]
- Barker, M.; Rayens, W. Partial least squares for discrimination. J. Chemom. 2003, 17, 166–173. [Google Scholar] [CrossRef]
- Höskuldsson, A. PLS regression methods. J. Chemom. 1988, 2, 211–228. [Google Scholar] [CrossRef]
- Wold, S.; Sjöström, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemom. Intell. Lab. Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Ballabio, D.; Consonni, V. Classification tools in chemistry. Part 1: Linear models. PLS-DA. Anal. Methods 2013, 5, 3790–3798. [Google Scholar] [CrossRef]
- Copley, T.; Aliferis, K.; Jabaji, S. Maple bark biochar affects rhizoctonia solani Metabolism and increases damping-off severity. Phytopathology 2015, 105, 1334–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, K.; Anderegg, J.; Mikaberidze, A.; Karisto, P.; Mascher, F.; McDonald, B.A.; Walter, A.; Hund, A. Hyperspectral canopy sensing of Wheat Septoria Tritici Blotch Disease. Front. Plant Sci. 2018, 9, 1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, R.-Q.; Jin, J.-J.; Li, Q.-M.; Su, Z.-Z.; Yu, X.-J.; Tang, Y.; Luo, S.-M.; He, Y.; Li, X.-L. Early detection of magnaporthe oryzae-infected barley leaves and lesion visualization based on hyperspectral imaging. Front. Plant Sci. 2018, 9, 1962. [Google Scholar] [CrossRef]
- Deng, H.; He, R.; Xia, H.; Xu, N.; Deng, Q.; Liang, D.; Lin, L.; Liao, L.; Xiong, B.; Xie, X.; et al. Ultra-HPLC-MS pseudo-targeted metabolomic profiling reveals metabolites and associated metabolic pathway alterations in Asian plum (Prunus salicina) fruits in response to gummosis disease. Funct. Plant Biol. 2022, 49, 936–945. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Chu, B.; Zhang, C.; Liu, F.; Jiang, L.; He, L. Hyperspectral imaging for presymptomatic detection of tobacco disease with successive projections algorithm and machine-learning classifiers. Sci. Rep. 2017, 7, 4125. [Google Scholar] [CrossRef] [Green Version]
- Monis, J.; Bestwick, R.K. Detection and localization of Grapevine Leafroll Associated Closteroviruses in greenhouse and tissue culture grown plants. Am. J. Enol. Vitic. 1996, 47, 199. [Google Scholar] [CrossRef]
- Bioreba, A.G. Double Antibody Sandwich Enzyme-Linked Immunosorbent Assay (DAS-ELISA): Test Specifications. Available online: https://www.bioreba.ch/saas/CustomUpload/374O357O340O370O356O369O350O321O360O366O369O356O353O352O350O320O326O/ELISA_Test_procedure_efd_and_es.pdf (accessed on 7 February 2023).
- Engel, J.; Gerretzen, J.; Szymańska, E.; Jansen, J.J.; Downey, G.; Blanchet, L.; Buydens, L.M.C. Breaking with trends in pre-processing? TrAC Trends Anal. Chem. 2013, 50, 96–106. [Google Scholar] [CrossRef]
- Rinnan, Å.; Nørgaard, L.; Berg, F.v.d.; Thygesen, J.; Bro, R.; Engelsen, S.B. Chapter 2—Data Pre-processing. In Infrared Spectroscopy for Food Quality Analysis and Control; Sun, D.-W., Ed.; Academic Press: San Diego, CA, USA, 2009; pp. 29–50. [Google Scholar] [CrossRef]
- Savitzky, A.; Golay, M.J.E. Smoothing and Differentiation of Data by Simplified Least Squares Procedures. Anal. Chem. 1964, 36, 1627–1639. [Google Scholar] [CrossRef]
- Barnes, R.J.; Dhanoa, M.S.; Lister, S.J. Standard normal variate transformation and de-trending of near-infrared diffuse reflectance spectra. Appl. Spectrosc. 1989, 43, 772–777. [Google Scholar] [CrossRef]
- Hofer, M. Mean centering. In The International Encyclopedia of Communication Research Methods; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 1–3. [Google Scholar] [CrossRef]
- Miller, C.E. Chemometrics in Process Analytical Technology (PAT). In Process Analytical Technology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; pp. 353–438. [Google Scholar] [CrossRef]
- Kuligowski, J.; Pérez-Guaita, D.; Quintás, G. Application of discriminant analysis and cross-validation on proteomics data. In Statistical Analysis in Proteomics; Jung, K., Ed.; Springer New York: New York, NY, USA, 2016; pp. 175–184. [Google Scholar] [CrossRef]
- Rácz, A.; Bajusz, D.; Héberger, K. Modelling methods and cross-validation variants in QSAR: A multi-level analysis$. SAR QSAR Environ. Res. 2018, 29, 661–674. [Google Scholar] [CrossRef]
- Götz, O.; Liehr-Gobbers, K.; Krafft, M. Evaluation of structural equation models using the Partial Least Squares (PLS) approach. In Handbook of Partial Least Squares: Concepts, Methods and Applications; Esposito Vinzi, V., Chin, W.W., Henseler, J., Wang, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 691–711. [Google Scholar] [CrossRef]
- Lê Cao, K.-A.; Welham, Z.M. PLS-Discriminant Aalysis (PLS-DA). In Multivariate Data Integration Using R: Methods and Applications with the MixOmics Package; Routledge: Oxford, UK, 2021; pp. 201–231. [Google Scholar] [CrossRef]
- Wold, S. Cross-validatory estimation of the number of components in factor and principal components models. Technometrics 1978, 20, 397–405. [Google Scholar] [CrossRef]
- Matthews, B.W. Comparison of the predicted and observed secondary structure of T4 phage lysozyme. Biochim. Biophys. Acta (BBA) Protein Struct. 1975, 405, 442–451. [Google Scholar] [CrossRef]
- Chicco, D.; Jurman, G. The advantages of the Matthews correlation coefficient (MCC) over F1 score and accuracy in binary classification evaluation. BMC Genom. 2020, 21, 6. [Google Scholar] [CrossRef] [Green Version]
- Shabanian, M.; Xiao, H.; Meng, B. Seasonal dynamics and tissue distribution of two major viruses associated with grapevine Leafroll under cool climate condition. Eur. J. Plant Pathol. 2020, 158, 1017–1031. [Google Scholar] [CrossRef]
- Bertazzon, N.; Forte, V.; Filippin, L.; Causin, R.; Maixner, M.; Angelini, E. Association between genetic variability and titre of Grapevine Pinot gris virus with disease symptoms. Plant Pathol. 2017, 66, 949–959. [Google Scholar] [CrossRef]
- Martelli, G.P. Grapevine virology highlights 2006–2009. In Proceedings of the 16th Meeting of the International Council for the Study of Virus and Virus-like Diseases of the Grapevine, Dijon, France, 31 August–4 September 2009; pp. 15–23. [Google Scholar]
- Montero, R.; El aou ouad, H.; Pacifico, D.; Marzachì, C.; Castillo, N.; García, E.; Del Saz, N.F.; Florez-Sarasa, I.; Flexas, J.; Bota, J. Effects of Grapevine leafroll-associated virus 3 on the physiology in asymptomatic plants of Vitis vinifera. Ann. Appl. Biol. 2017, 171, 155–171. [Google Scholar] [CrossRef]
- Chooi, K.M.; Bell, V.A.; Blouin, A.G.; Cohen, D.; Mundy, D.; Henshall, W.; MacDiarmid, R.M. Grapevine leafroll-associated virus 3 genotype influences foliar symptom development in New Zealand vineyards. Viruses 2022, 14, 1348. [Google Scholar] [CrossRef]
- Gutha, L.R.; Casassa, L.F.; Harbertson, J.F.; Naidu, R.A. Modulation of flavonoid biosynthetic pathway genes and anthocyanins due to virus infection in grapevine (Vitis vinifera L.) leaves. BMC Plant Biol. 2010, 10, 187. [Google Scholar] [CrossRef] [Green Version]
- Gould, K.S. Nature’s Swiss army knife: The diverse protective roles of anthocyanins in leaves. J. Biomed. Biotechnol. 2004, 2004, 314–320. [Google Scholar] [CrossRef] [Green Version]
- Merzlyak, M.N.; Chivkunova, O.B.; Solovchenko, A.E.; Naqvi, K.R. Light absorption by anthocyanins in juvenile, stressed, and senescing leaves. J. Exp. Bot. 2008, 59, 3903–3911. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, V.; Stefova, M.; Vojnoski, B.; Dörnyei, Á.; Márk, L.; Dimovska, V.; Stafilov, T.; Kilár, F. Identification of polyphenolic compounds in red and white grape varieties grown in R. Macedonia and changes of their content during ripening. Food Res. Int. 2011, 44, 2851–2860. [Google Scholar] [CrossRef]
- Rustioni, L.; Rocchi, L.; Failla, O. Effect of anthocyanin absence on white berry grape (Vitis vinifera L.). Vitis 2015, 54, 239–242. [Google Scholar]
- Eitel, J.U.H.; Vierling, L.A.; Litvak, M.E.; Long, D.S.; Schulthess, U.; Ager, A.A.; Krofcheck, D.J.; Stoscheck, L. Broadband, red-edge information from satellites improves early stress detection in a New Mexico conifer woodland. Remote Sens. Environ. 2011, 115, 3640–3646. [Google Scholar] [CrossRef]
- Asmaryan, S.; Warner, T.A.; Muradyan, V.; Nersisyan, G. Mapping tree stress associated with urban pollution using the WorldView-2 Red Edge band. Remote Sens. Lett. 2013, 4, 200–209. [Google Scholar] [CrossRef]
- Horler, D.N.H.; Dockray, M.; Barber, J. The red edge of plant leaf reflectance. Int. J. Remote Sens. 1983, 4, 273–288. [Google Scholar] [CrossRef]
- Smith, K.L.; Steven, M.D.; Colls, J.J. Use of hyperspectral derivative ratios in the red-edge region to identify plant stress responses to gas leaks. Remote Sens. Environ. 2004, 92, 207–217. [Google Scholar] [CrossRef]
- Borràs, E.; Ferré, J.; Boqué, R.; Mestres, M.; Aceña, L.; Busto, O. Data fusion methodologies for food and beverage authentication and quality assessment—A review. Anal. Chim. Acta 2015, 891, 1–14. [Google Scholar] [CrossRef]
- Feng, Z.; Song, L.; Duan, J.; He, L.; Zhang, Y.; Wei, Y.; Feng, W. Monitoring wheat powdery mildew based on hyperspectral, thermal infrared, and rgb image data fusion. Sensors 2021, 22, 31. [Google Scholar] [CrossRef] [PubMed]
- Owomugisha, G.; Nuwamanya, E.; Quinn, J.A.; Biehl, M.; Mwebaze, E. Early detection of plant diseases using spectral data. In Proceedings of the 3rd International Conference on Applications of Intelligent Systems, Las Palmas de Gran Canaria, Spain, 7–9 January 2020; pp. 1–6. [Google Scholar] [CrossRef] [Green Version]
- Golhani, K.; Balasundram, S.K.; Vadamalai, G.; Pradhan, B. A review of neural networks in plant disease detection using hyperspectral data. Inf. Process. Agric. 2018, 5, 354–371. [Google Scholar] [CrossRef]
- Li, S.; Song, W.; Fang, L.; Chen, Y.; Ghamisi, P.; Benediktsson, J.A. Deep learning for hyperspectral image classification: An overview. IEEE Trans. Geosci. Remote Sens. 2019, 57, 6690–6709. [Google Scholar] [CrossRef] [Green Version]
- Mishra, P.; Polder, G.; Vilfan, N. Close range spectral imaging for disease detection in plants using autonomous platforms: A review on recent studies. Curr. Robot. Rep. 2020, 1, 43–48. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, S.L.; Staid, M.; Staid, M.; Cooper, M.L. Remote hyperspectral imaging of grapevine leafroll-associated virus 3 in Cabernet Sauvignon vineyards. Comput. Electron. Agric. 2016, 130, 109–117. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diseased (GLRaV-1 + GVA) | Healthy | Total | |
|---|---|---|---|
| Chardonnay | 134 | 40 | 174 |
| Pinot Noir | 72 | 101 | 173 |
| Calibration Model Results | Cross-Validation Results | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Confusion Matrix | Confusion Matrix | |||||||||||||
| Time | LVs | Predicted | Actual Disease | Actual Healthy | Sensitivity | F1-Score | Accuracy | MCC | Actual Disease | Actual Healthy | Sensitivity | F1-Score | Accuracy | MCC |
| Nov | 2 | Disease | 80 | 15 | 0.60 | 0.70 | 0.60 | 0.19 | 77 | 14 | 0.57 | 0.68 | 0.59 | 0.19 |
| Healthy | 54 | 25 | 0.63 | 0.42 | 57 | 26 | 0.65 | 0.42 | ||||||
| Dec | 3 | Disease | 95 | 12 | 0.71 | 0.79 | 0.71 | 0.35 | 92 | 13 | 0.69 | 0.77 | 0.68 | 0.31 |
| Healthy | 39 | 28 | 0.70 | 0.52 | 42 | 27 | 0.68 | 0.50 | ||||||
| Jan | 3 | Disease | 96 | 10 | 0.72 | 0.80 | 0.72 | 0.40 | 91 | 15 | 0.68 | 0.76 | 0.67 | 0.26 |
| Healthy | 38 | 30 | 0.75 | 0.56 | 43 | 25 | 0.63 | 0.46 | ||||||
| Feb | 3 | Disease | 98 | 11 | 0.73 | 0.81 | 0.73 | 0.40 | 96 | 16 | 0.72 | 0.78 | 0.69 | 0.28 |
| Healthy | 36 | 29 | 0.73 | 0.55 | 38 | 24 | 0.60 | 0.47 | ||||||
| Mar | 4 | Disease | 102 | 9 | 0.76 | 0.83 | 0.76 | 0.47 | 99 | 11 | 0.74 | 0.81 | 0.74 | 0.41 |
| Healthy | 32 | 31 | 0.78 | 0.60 | 35 | 29 | 0.73 | 0.56 | ||||||
| Apr | 4 | Disease | 88 | 11 | 0.66 | 0.76 | 0.67 | 0.32 | 86 | 15 | 0.64 | 0.73 | 0.64 | 0.22 |
| Healthy | 46 | 28 | 0.72 | 0.50 | 48 | 24 | 0.62 | 0.43 | ||||||
| Calibration Model Results | Cross-Validation Results | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Confusion Matrix | Confusion Matrix | |||||||||||||
| Time | LVs | Predicted | Actual Disease | Actual Healthy | Sensitivity | F1-Score | Accuracy | MCC | Actual Disease | Actual Healthy | Sensitivity | F1-Score | Accuracy | MCC |
| Nov | 1 | Disease | 43 | 40 | 0.60 | 0.55 | 0.60 | 0.20 | 43 | 46 | 0.60 | 0.53 | 0.57 | 0.14 |
| Healthy | 29 | 61 | 0.60 | 0.64 | 29 | 55 | 0.54 | 0.59 | ||||||
| Dec | 3 | Disease | 48 | 33 | 0.68 | 0.63 | 0.67 | 0.35 | 45 | 34 | 0.63 | 0.60 | 0.65 | 0.29 |
| Healthy | 23 | 68 | 0.67 | 0.71 | 26 | 67 | 0.66 | 0.69 | ||||||
| Jan | 2 | Disease | 63 | 10 | 0.88 | 0.87 | 0.89 | 0.78 | 64 | 12 | 0.89 | 0.86 | 0.88 | 0.77 |
| Healthy | 9 | 91 | 0.90 | 0.91 | 8 | 89 | 0.88 | 0.90 | ||||||
| Feb | 2 | Disease | 61 | 3 | 0.85 | 0.90 | 0.92 | 0.84 | 58 | 2 | 0.81 | 0.88 | 0.91 | 0.81 |
| Healthy | 11 | 98 | 0.97 | 0.93 | 14 | 99 | 0.98 | 0.93 | ||||||
| Mar | 3 | Disease | 54 | 9 | 0.75 | 0.80 | 0.84 | 0.68 | 56 | 12 | 0.78 | 0.80 | 0.84 | 0.67 |
| Healthy | 18 | 92 | 0.91 | 0.87 | 16 | 89 | 0.88 | 0.86 | ||||||
| Apr | 5 | Disease | 66 | 1 | 0.92 | 0.95 | 0.96 | 0.92 | 66 | 3 | 0.92 | 0.94 | 0.95 | 0.89 |
| Healthy | 6 | 100 | 0.99 | 0.97 | 6 | 98 | 0.97 | 0.96 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.M.; Ostendorf, B.; Pagay, V. Detecting Grapevine Virus Infections in Red and White Winegrape Canopies Using Proximal Hyperspectral Sensing. Sensors 2023, 23, 2851. https://doi.org/10.3390/s23052851
Wang YM, Ostendorf B, Pagay V. Detecting Grapevine Virus Infections in Red and White Winegrape Canopies Using Proximal Hyperspectral Sensing. Sensors. 2023; 23(5):2851. https://doi.org/10.3390/s23052851
Chicago/Turabian StyleWang, Yeniu Mickey, Bertram Ostendorf, and Vinay Pagay. 2023. "Detecting Grapevine Virus Infections in Red and White Winegrape Canopies Using Proximal Hyperspectral Sensing" Sensors 23, no. 5: 2851. https://doi.org/10.3390/s23052851
APA StyleWang, Y. M., Ostendorf, B., & Pagay, V. (2023). Detecting Grapevine Virus Infections in Red and White Winegrape Canopies Using Proximal Hyperspectral Sensing. Sensors, 23(5), 2851. https://doi.org/10.3390/s23052851