

A Supervised Machine Learning Approach to Classify Brain Morphology of Professional Visual Artists versus Non-Artists

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Imagery Questionnaire
2.3. Brain Data Collection
2.4. Preprocessing
2.5. Supervised Machine Learning Procedure
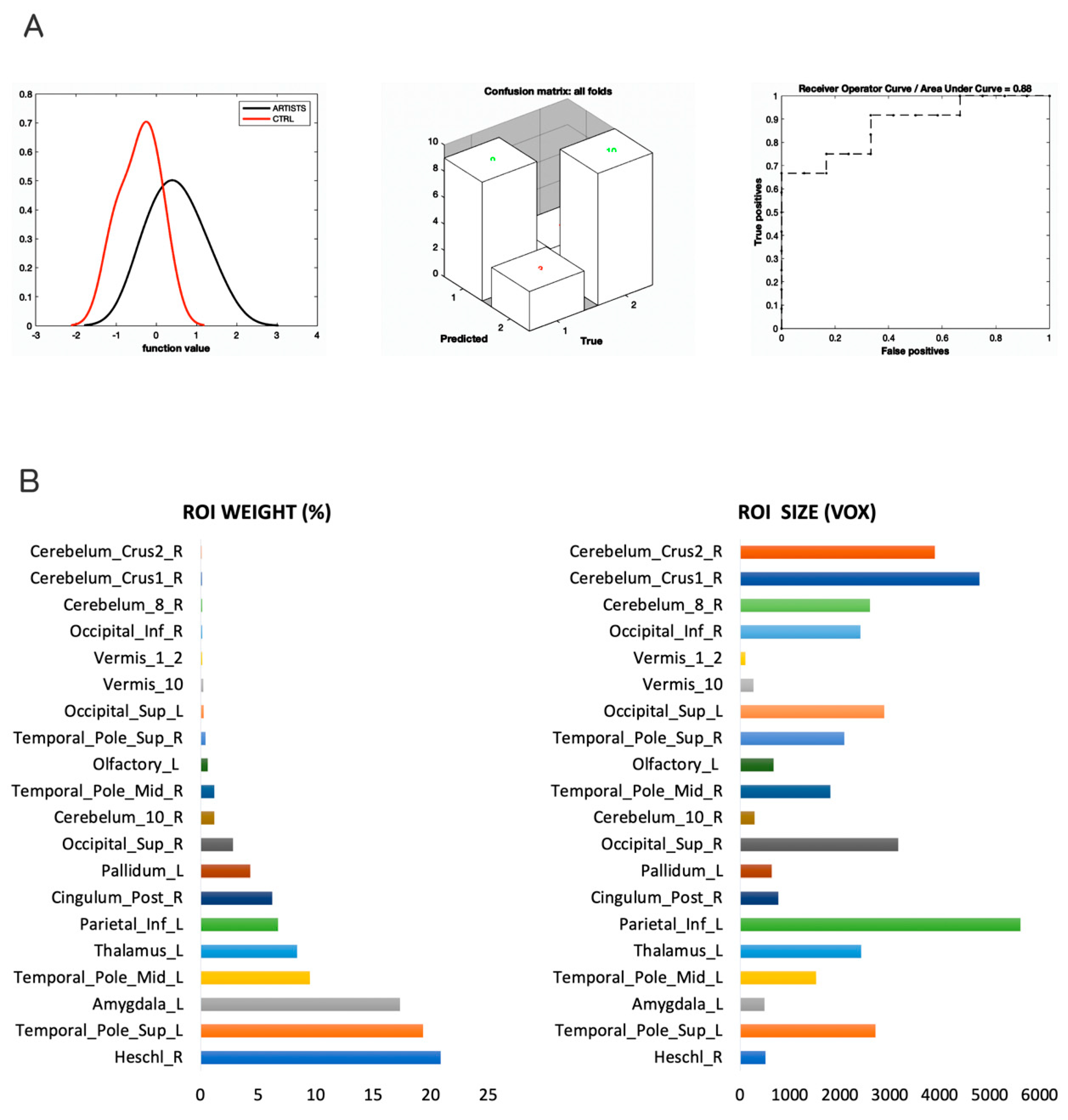
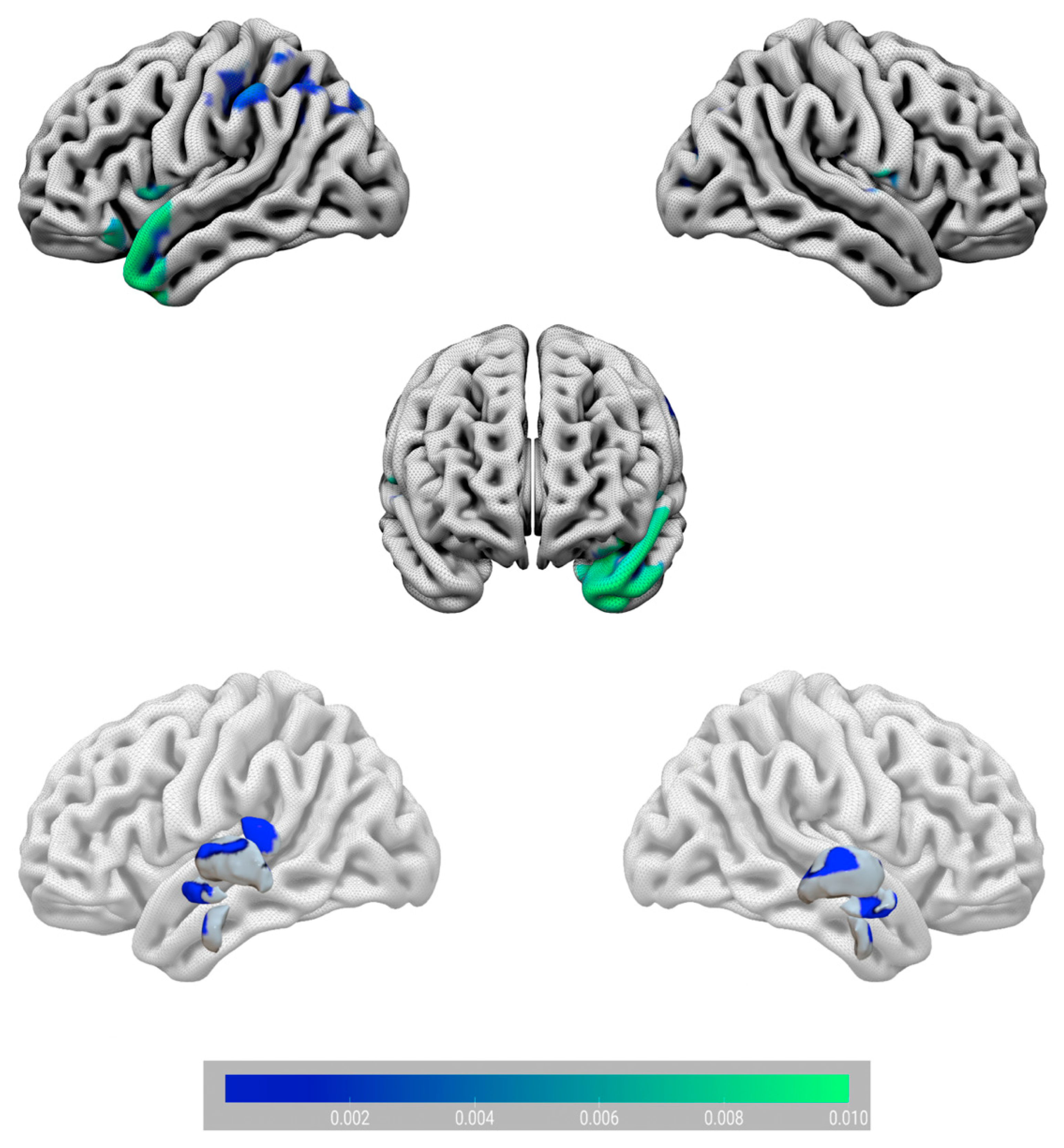
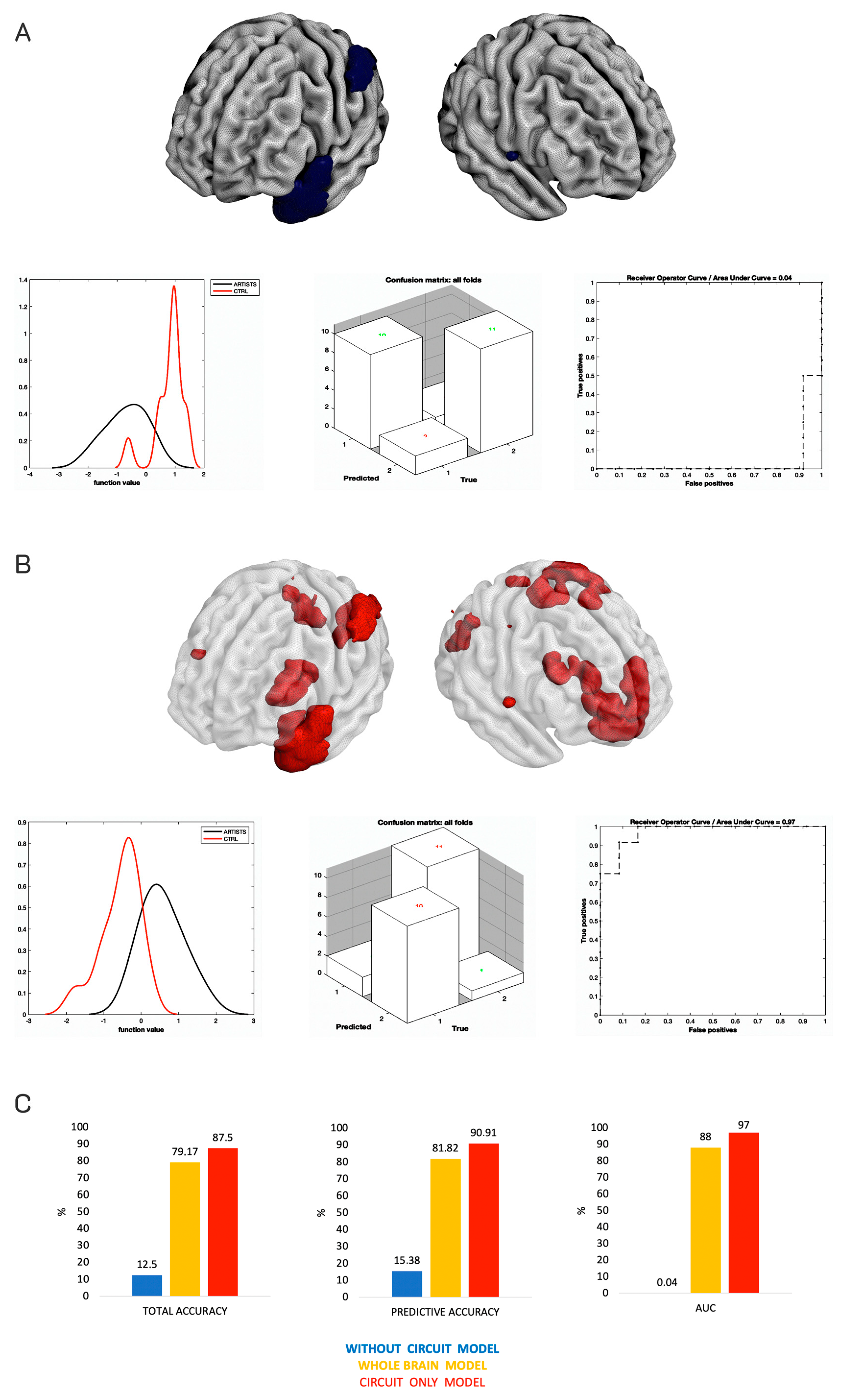
3. Results
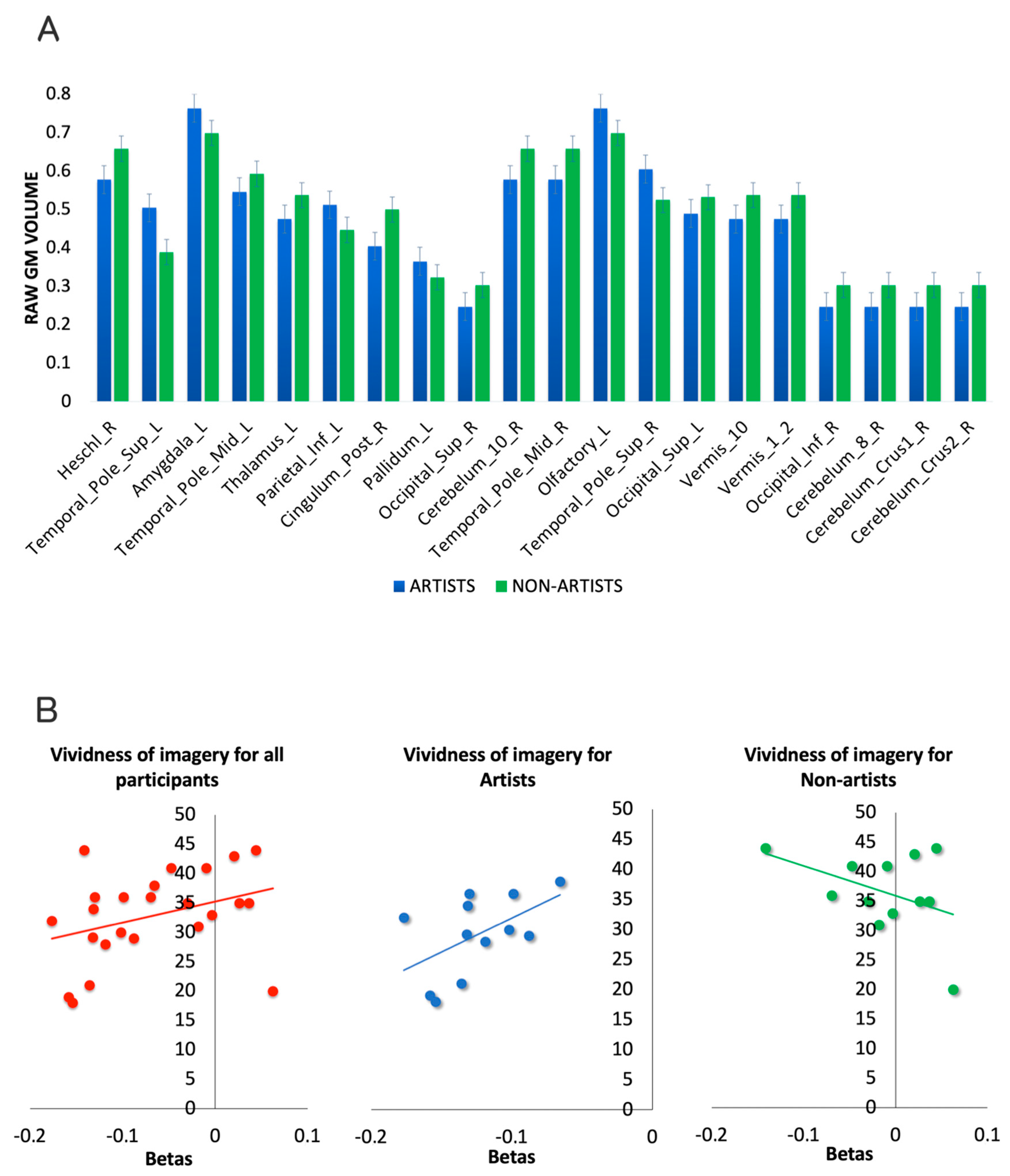
Additional Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Pisapia, N.; Rastelli, C. Creativity as an Information-Based Process. Riv. Internazionale Filos. Psicol. 2022, 13, 1–18. [Google Scholar]
- Simon, H.A. The Logic of Heuristic Decision Making. In Models of Discovery; Springer: Berlin/Heidelberg, Germany, 1977; pp. 154–175. [Google Scholar]
- Feist, G.J. The Psychology of Science and the Origins of the Scientific Mind; Yale University Press: New Haven, CT, USA, 2006; ISBN 9780300110746. [Google Scholar]
- Catmull, E.; Wallace, A. Srilakshmi Creativity Inc.: Overcoming the Unseen Forces That Stand in the Way of True Inspiration; Random House: New York, NY, USA, 2015; Volume 8, ISBN 0593070100. [Google Scholar]
- Kaufman, J.C.; Sternberg, R.J. The Cambridge Handbook of Creativity; Cambridge University Press: Cambridge, UK, 2010; ISBN 978-1-139-49061-0. [Google Scholar]
- Rastelli, C.; Greco, A.; De Pisapia, N.; Finocchiaro, C. Balancing novelty and appropriateness leads to creative associations in children. PNAS Nexus 2022, 1, pgac273. [Google Scholar] [CrossRef]
- Dolan, P.; Metcalfe, R. The relationship between innovation and subjective wellbeing. Res. Policy 2012, 41, 1489–1498. [Google Scholar] [CrossRef]
- Palmiero, M.; Nori, R.; Piccardi, L. Verbal and visual divergent thinking in aging. Exp. Brain Res. 2017, 235, 1021–1029. [Google Scholar] [CrossRef]
- Schouten, K.A.; de Niet, G.J.; Knipscheer, J.W.; Kleber, R.J.; Hutschemaekers, G.J.M. The Effectiveness of Art Therapy in the Treatment of Traumatized Adults: A Systematic Review on Art Therapy and Trauma. Trauma Violence Abus. 2014, 16, 220–228. [Google Scholar] [CrossRef]
- Gajda, A.; Karwowski, M.; Beghetto, R.A. Creativity and Academic Achievement: A Meta-Analysis. J. Educ. Psychol. 2017, 109, 269–299. [Google Scholar] [CrossRef]
- Carson, S.H.; Peterson, J.B.; Higgins, D.M. Reliability, Validity, and Factor Structure of the Creative Achievement Questionnaire. Creat. Res. J. 2005, 17, 37–50. [Google Scholar] [CrossRef]
- Zaidel, D.W.; Nadal, M.; Flexas, A.; Munar, E. An Evolutionary Approach to Art and Aesthetic Experience. Psychol. Aesthet. Creat. Arts 2013, 7, 100–109. [Google Scholar] [CrossRef]
- Aubert, M.; Brumm, A.; Ramli, M.; Sutikna, T.; Saptomo, E.W.; Hakim, B.; Morwood, M.J.; Van Den Bergh, G.D.; Kinsley, L.; Dosseto, A. Pleistocene Cave Art from Sulawesi, Indonesia. Nature 2014, 514, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Morriss-Kay, G.M. The Evolution of Human Artistic Creativity. J. Anat. 2010, 216, 158–176. [Google Scholar] [CrossRef]
- Guilford, J.P. Creativity: Yesterday, Today and Tomorrow. J. Creat. Behav. 1967, 1, 3–14. [Google Scholar] [CrossRef]
- Mednick, S. The Associative Basis of the Creative Process. Psychol. Rev. 1962, 69, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A. The Neuroscience of Creativity, 1st ed.; Cambridge University Press: Cambridge, UK, 2018; ISBN 978-1-316-81698-1. [Google Scholar]
- Gabora, L. Honing Theory: A Complex Systems Framework for Creativity. arXiv 2016, arXiv:1610.02484. [Google Scholar]
- Beaty, R.E.; Seli, P.; Schacter, D.L. Network Neuroscience of Creative Cognition: Mapping Cognitive Mechanisms and Individual Differences in the Creative Brain. Curr. Opin. Behav. Sci. 2019, 27, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Jung, R.E. The Structure of Creative Cognition in the Human Brain. Front. Hum. Neurosci. 2013, 7, 330. [Google Scholar] [CrossRef]
- Ellamil, M.; Dobson, C.; Beeman, M.; Christoff, K. Evaluative and Generative Modes of Thought during the Creative Process. NeuroImage 2012, 59, 1783–1794. [Google Scholar] [CrossRef]
- Saggar, M.; Quintin, E.-M.; Kienitz, E.; Bott, N.T.; Sun, Z.; Hong, W.-C.; Chien, Y.; Liu, N.; Dougherty, R.F.; Royalty, A.; et al. Pictionary-Based FMRI Paradigm to Study the Neural Correlates of Spontaneous Improvisation and Figural Creativity. Sci. Rep. 2015, 5, 10894. [Google Scholar] [CrossRef]
- De Pisapia, N.; Bacci, F.; Parrott, D.; Melcher, D. Brain Networks for Visual Creativity: A Functional Connectivity Study of Planning a Visual Artwork. Sci. Rep. 2016, 6, 39185. [Google Scholar] [CrossRef]
- Dietrich, A. Where in the Brain Is Creativity: A Brief Account of a Wild-Goose Chase. Curr. Opin. Behav. Sci. 2019, 27, 36–39. [Google Scholar] [CrossRef]
- Chamberlain, R.; McManus, I.C.; Brunswick, N.; Rankin, Q.; Riley, H.; Kanai, R. Drawing on the Right Side of the Brain: A Voxel-Based Morphometry Analysis of Observational Drawing. NeuroImage 2014, 96, 167–173. [Google Scholar] [CrossRef]
- Shi, B.; Cao, X.; Chen, Q.; Zhuang, K.; Qiu, J. Different Brain Structures Associated with Artistic and Scientific Creativity: A Voxel-Based Morphometry Study. Sci. Rep. 2017, 7, 42911. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, A.; Alexander, P.; Fogelson, S.V.; Li, X.; Lu, Z.; Kohler, P.J.; Riley, E.; Peter, U.T.; Meng, M. The Artist Emerges: Visual Art Learning Alters Neural Structure and Function. NeuroImage 2015, 105, 440–451. [Google Scholar] [CrossRef]
- Xurui, T.; Yaxu, Y.; Qiangqiang, L.; Yu, M.; Bin, Z.; Xueming, B. Mechanisms of Creativity Differences between Art and Non-Art Majors: A Voxel-Based Morphometry Study. Front. Psychol. 2018, 9, 2319. [Google Scholar] [CrossRef]
- Sporns, O. Contributions and Challenges for Network Models in Cognitive Neuroscience. Nat. Neurosci. 2014, 17, 652–660. [Google Scholar] [CrossRef]
- Kowatari, Y.; Lee, S.H.; Yamamura, H.; Nagamori, Y.; Levy, P.; Yamane, S.; Yamamoto, M. Neural Networks Involved in Artistic Creativity. Hum. Brain Mapp. 2009, 30, 1678–1690. [Google Scholar] [CrossRef]
- Seeley, W.W.; Menon, V.; Schatzberg, A.F.; Keller, J.; Glover, G.H.; Kenna, H.; Reiss, A.L.; Greicius, M.D. Dissociable Intrinsic Connectivity Networks for Salience Processing and Executive Control. J. Neurosci. 2007, 27, 2349–2356. [Google Scholar] [CrossRef] [PubMed]
- De Pisapia, N.; Repovs, G.; Braver, T.S. Computational Models of Attention and Cognitive Control; Sun, R., Ed.; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Kenett, Y.N.; Medaglia, J.D.; Beaty, R.E.; Chen, Q.; Betzel, R.F.; Thompson-Schill, S.L.; Qiu, J. Driving the Brain towards Creativity and Intelligence: A Network Control Theory Analysis. Neuropsychologia 2018, 118, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Chrysikou, E.G. Creativity in and out of (Cognitive) Control. Curr. Opin. Behav. Sci. 2019, 27, 94–99. [Google Scholar] [CrossRef]
- Niendam, T.A.; Laird, A.R.; Ray, K.L.; Dean, Y.M.; Glahn, D.C.; Carter, C.S. Meta-Analytic Evidence for a Superordinate Cognitive Control Network Subserving Diverse Executive Functions. Cogn. Affect. Behav. Neurosci. 2012, 12, 241–268. [Google Scholar] [CrossRef]
- Raichle, M.E. The Brain’s Default Mode Network. Annu. Rev. Neurosci. 2015, 38, 433–447. [Google Scholar] [CrossRef]
- Zabelina, D.L.; Andrews-Hanna, J.R. Dynamic Network Interactions Supporting Internally-Oriented Cognition. Curr. Opin. Neurobiol. 2016, 40, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Buckner, R.L.; DiNicola, L.M. The Brain’s Default Network: Updated Anatomy, Physiology and Evolving Insights. Nat. Rev. Neurosci. 2019, 20, 593–608. [Google Scholar] [CrossRef] [PubMed]
- Park, H.R.; Kirk, I.J.; Waldie, K.E. Neural Correlates of Creative Thinking and Schizotypy. Neuropsychologia 2015, 73, 94–107. [Google Scholar] [CrossRef] [PubMed]
- Madore, K.P.; Thakral, P.P.; Beaty, R.E.; Addis, D.R.; Schacter, D.L. Neural Mechanisms of Episodic Retrieval Support Divergent Creative Thinking. Cereb. Cortex 2019, 29, 150–166. [Google Scholar] [CrossRef]
- Chen, Q.; Beaty, R.E.; Qiu, J. Mapping the Artistic Brain: Common and Distinct Neural Activations Associated with Musical, Drawing, and Literary Creativity. Hum. Brain Mapp. 2020, 41, 3403–3419. [Google Scholar] [CrossRef]
- Stevenson, C.; Baas, M.; van der Maas, H. A Minimal Theory of Creative Ability. J. Intell. 2021, 9, 9. [Google Scholar] [CrossRef]
- Pascual-Leone, A.; Amedi, A.; Fregni, F.; Merabet, L.B. The Plastic Human Brain Cortex. Annu. Rev. Neurosci. 2005, 28, 377–401. [Google Scholar] [CrossRef]
- Medaglia, J.D.; Huang, W.; Karuza, E.A.; Kelkar, A.; Thompson-Schill, S.L.; Ribeiro, A.; Bassett, D.S. Functional Alignment with Anatomical Networks Is Associated with Cognitive Flexibility. Nat. Hum. Behav. 2018, 2, 156–164. [Google Scholar] [CrossRef]
- Gu, S.; Pasqualetti, F.; Cieslak, M.; Telesford, Q.K.; Alfred, B.Y.; Kahn, A.E.; Medaglia, J.D.; Vettel, J.M.; Miller, M.B.; Grafton, S.T. Controllability of Structural Brain Networks. Nat. Commun. 2015, 6, 8414. [Google Scholar] [CrossRef]
- Hermundstad, A.M.; Bassett, D.S.; Brown, K.S.; Aminoff, E.M.; Clewett, D.; Freeman, S.; Frithsen, A.; Johnson, A.; Tipper, C.M.; Miller, M.B. Structural Foundations of Resting-State and Task-Based Functional Connectivity in the Human Brain. Proc. Natl. Acad. Sci. USA 2013, 110, 6169–6174. [Google Scholar] [CrossRef]
- Woollett, K.; Spiers, H.J.; Maguire, E.A. Talent in the Taxi: A Model System for Exploring Expertise. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 1407–1416. [Google Scholar] [CrossRef]
- Gobet, F. Expertise vs. Talent. Talent. Dev. Excell. 2013, 5, 75–86. [Google Scholar]
- Goel, V.; Grafman, J. Role of the Right Prefrontal Cortex in Ill-Structured Planning. Cogn. Neuropsychol. 2000, 17, 415–436. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.L.; Hou, C.E. Portraits of Artists: Emergence of Visual Creativity in Dementia. Arch. Neurol. 2004, 61, 842–844. [Google Scholar] [CrossRef] [PubMed]
- Drago, V.; Foster, P.S.; Okun, M.S.; Haq, I.; Sudhyadhom, A.; Skidmore, F.M.; Heilman, K.M. Artistic Creativity and DBS: A Case Report. J. Neurol. Sci. 2009, 276, 138–142. [Google Scholar] [CrossRef]
- Rankin, K.P.; Liu, A.A.; Howard, S.; Slama, H.; Hou, C.E.; Shuster, K.; Miller, B.L. A Case-Controlled Study of Altered Visual Art Production in Alzheimer’s and FTLD. Cogn. Behav. Neurol. 2007, 20, 48–61. [Google Scholar] [CrossRef]
- Sunavsky, A.; Poppenk, J. Neuroimaging Predictors of Creativity in Healthy Adults. NeuroImage 2020, 206, 116292. [Google Scholar] [CrossRef]
- Chrysikou, E.G.; Wertz, C.; Yaden, D.B.; Kaufman, S.B.; Bacon, D.; Wintering, N.A.; Jung, R.E.; Newberg, A.B. Differences in Brain Morphometry Associated with Creative Performance in High- and Average-Creative Achievers. NeuroImage 2020, 218, 116921. [Google Scholar] [CrossRef]
- Wertz, C.J.; Chohan, M.O.; Flores, R.A.; Jung, R.E. Neuroanatomy of Creative Achievement. NeuroImage 2020, 209, 116487. [Google Scholar] [CrossRef] [PubMed]
- Norman, K.A.; Polyn, S.M.; Detre, G.J.; Haxby, J.V. Beyond Mind-Reading: Multi-Voxel Pattern Analysis of FMRI Data. Trends Cogn. Sci. 2006, 10, 424–430. [Google Scholar] [CrossRef]
- Grecucci, A.; Dadomo, H.; Salvato, G.; Lapomarda, G.; Sorella, S.; Messina, I. Two Grey–White Matter Circuits Separate Borderline Personality Disorder from Controls and Mediate the Relationship between Specific Childhood Traumas and Symptoms. A mCCA+ jICA and Random Forest Approach. (Under Review, Preprint Available). Available online: https://www.preprints.org/manuscript/202302.0089/v1 (accessed on 20 January 2023).
- Ghomroudi, P.A.; Scaltritti, M.; Grecucci, A. Decoding reappraisal and suppression from neural circuits: A combined supervised and unsupervised machine learning approach. Cogn. Affect. Behav. Neurosci. 2023, 1–8. [Google Scholar] [CrossRef]
- Grecucci, A.; Lapomarda, G.; Messina, I.; Monachesi, B.; Sorella, S.; Siugzdaite, R. Structural Features Related to Affective Instability Correctly Classify Patients with Borderline Personality Disorder. A Supervised Machine Learning Approach. Front. Psychiatry 2022, 13, 804440. [Google Scholar] [CrossRef]
- Caria, A.; Grecucci, A. Neuroanatomical Predictors of Real-Time FMRI-Based Emotional Brain Regulation. Psychophys 2022, 50, 5. [Google Scholar] [CrossRef]
- Rondina, J.M.; Ferreira, L.K.; de Souza Duran, F.L.; Kubo, R.; Ono, C.R.; Leite, C.C.; Smid, J.; Nitrini, R.; Buchpiguel, C.A.; Busatto, G.F. Selecting the Most Relevant Brain Regions to Discriminate Alzheimer’s Disease Patients from Healthy Controls Using Multiple Kernel Learning: A Comparison across Functional and Structural Imaging Modalities and Atlases. NeuroImage Clin. 2018, 17, 628–641. [Google Scholar] [CrossRef] [PubMed]
- Mourao-Miranda, J.; Reinders, A.A.T.S.; Rocha-Rego, V.; Lappin, J.; Rondina, J.; Morgan, C.; Morgan, K.D.; Fearon, P.; Jones, P.B.; Doody, G.A.; et al. Individualized Prediction of Illness Course at the First Psychotic Episode: A Support Vector Machine MRI Study. Psychol. Med. 2012, 42, 1037–1047. [Google Scholar] [CrossRef]
- Melcher, D.; Bacci, F. Perception of Emotion in Abstract Artworks: A Multidisciplinary Approach. Prog. Brain Res. 2013, 204, 191–216. [Google Scholar] [CrossRef] [PubMed]
- Glazek, K. Visual and Motor Processing in Visual Artists: Implications for Cognitive and Neural Mechanisms. Psychol. Aesthet. Creat. Arts 2012, 6, 155–167. [Google Scholar] [CrossRef]
- Zabicki, A.; de Haas, B.; Zentgraf, K.; Stark, R.; Munzert, J.; Krüger, B. Subjective Vividness of Motor Imagery Has a Neural Signature in Human Premotor and Parietal Cortex. Neuroimage 2019, 197, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Bacci, F. Resonance: Snapshots of Creativity in the Brain, exh.cat; Edizioni dell’Università degli Studi di Trento: Trento, Italy, 2013. [Google Scholar]
- Penny, W.; Friston, K.; Ashburner, J.; Kiebel, S.; Nichols, T. Statistical Parametric Mapping: The Analysis of Functional Brain Images, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Gaser, C.; Dahnke, R.; Thompson, P.M.; Kurth, F.; Luders, E.; Initiative, A.D.N. CAT—A Computational Anatomy Toolbox for the Analysis of Structural MRI Data. BioRxiv 2022, 2022-06, 2022.06.11.495736. [Google Scholar]
- Ashburner, J. A Fast Diffeomorphic Image Registration Algorithm. Neuroimage 2007, 38, 95–113. [Google Scholar] [CrossRef] [PubMed]
- Orsenigo, C.; Vercellis, C. Kernel Ridge Regression for out-of-Sample Mapping in Supervised Manifold Learning. Expert Syst. Appl. 2012, 39, 7757–7762. [Google Scholar] [CrossRef]
- Dadomo, H.; Salvato, G.; Lapomarda, G.; Ciftci, Z.; Messina, I.; Grecucci, A. Structural Features Predict Sexual Trauma and Interpersonal Problems in Borderline Personality Disorder but Not in Controls: A Multi-Voxel Pattern Analysis. Front. Hum. Neurosci. 2022, 16, 773593. [Google Scholar] [CrossRef] [PubMed]
- Schrouff, J.; Rosa, M.J.; Rondina, J.M.; Marquand, A.F.; Chu, C.; Ashburner, J.; Phillips, C.; Richiardi, J.; Mourão-Miranda, J. PRoNTo: Pattern Recognition for Neuroimaging Toolbox. Neuroinformatics 2013, 11, 319–337. [Google Scholar] [CrossRef] [PubMed]
- Beaty, R.E.; Benedek, M.; Barry Kaufman, S.; Silvia, P.J. Default and Executive Network Coupling Supports Creative Idea Production. Sci. Rep. 2015, 5, 10964. [Google Scholar] [CrossRef] [PubMed]
- Tzourio-Mazoyer, N.; Landeau, B.; Papathanassiou, D.; Crivello, F.; Etard, O.; Delcroix, N.; Mazoyer, B.; Joliot, M. Automated Anatomical Labeling of Activations in SPM Using a Macroscopic Anatomical Parcellation of the MNI MRI Single-Subject Brain. Neuroimage 2002, 15, 273–289. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Yuan, Y.; Liu, C.; Luo, J. The Roles of the Temporal Lobe in Creative Insight: An Integrated Review. Think. Reason. 2017, 23, 321–375. [Google Scholar] [CrossRef]
- Jung, R.E.; Segall, J.M.; Jeremy Bockholt, H.; Flores, R.A.; Smith, S.M.; Chavez, R.S.; Haier, R.J. Neuroanatomy of Creativity. Hum. Brain Mapp. 2010, 31, 398–409. [Google Scholar] [CrossRef]
- Sawyer, R.K. Explaining Creativity: The Science of Human Innovation; Oxford University Press: Oxford, UK, 2011; ISBN 0-19-983820-8. [Google Scholar]
- Li, W.; Li, G.; Ji, B.; Zhang, Q.; Qiu, J. Neuroanatomical Correlates of Creativity: Evidence from Voxel-Based Morphometry. Front. Psychol. 2019, 10, 155. [Google Scholar] [CrossRef]
- Huang, P.; Qiu, L.; Shen, L.; Zhang, Y.; Song, Z.; Qi, Z.; Gong, Q.; Xie, P. Evidence for a Left-over-right Inhibitory Mechanism during Figural Creative Thinking in Healthy Nonartists. Hum. Brain Mapp. 2013, 34, 2724–2732. [Google Scholar] [CrossRef]
- Jung, R.E.; Flores, R.A.; Hunter, D. A New Measure of Imagination Ability: Anatomical Brain Imaging Correlates. Front. Psychol. 2016, 7, 496. [Google Scholar] [CrossRef]
- Squire, L.R.; Stark, C.E.; Clark, R.E. The Medial Temporal Lobe. Annu. Rev. Neurosci. 2004, 27, 279–306. [Google Scholar] [CrossRef] [PubMed]
- Beaty, R.E.; Benedek, M.; Silvia, P.J.; Schacter, D.L. Creative Cognition and Brain Network Dynamics. Trends Cogn. Sci. 2016, 20, 87–95. [Google Scholar] [CrossRef]
- Jauk, E.; Neubauer, A.C.; Dunst, B.; Fink, A.; Benedek, M. Gray Matter Correlates of Creative Potential: A Latent Variable Voxel-Based Morphometry Study. Neuroimage 2015, 111, 312–320. [Google Scholar] [CrossRef]
- Benedek, M.; Schües, T.; Beaty, R.E.; Jauk, E.; Koschutnig, K.; Fink, A.; Neubauer, A.C. To Create or to Recall Original Ideas: Brain Processes Associated with the Imagination of Novel Object Uses. Cortex 2018, 99, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Fink, A.; Grabner, R.H.; Benedek, M.; Reishofer, G.; Hauswirth, V.; Fally, M.; Neuper, C.; Ebner, F.; Neubauer, A.C. The Creative Brain: Investigation of Brain Activity during Creative Problem Solving by Means of EEG and FMRI. Hum. Brain Mapp. 2009, 30, 734–748. [Google Scholar] [CrossRef] [PubMed]
- McPherson, M.J.; Barrett, F.S.; Lopez-Gonzalez, M.; Jiradejvong, P.; Limb, C.J. Emotional Intent Modulates the Neural Substrates of Creativity: An FMRI Study of Emotionally Targeted Improvisation in Jazz Musicians. Sci. Rep. 2016, 6, 18460. [Google Scholar] [CrossRef]
- Bashwiner, D.M.; Wertz, C.J.; Flores, R.A.; Jung, R.E. Musical Creativity “Revealed” in Brain Structure: Interplay between Motor, Default Mode and Limbic Networks. Sci. Rep. 2016, 6, 20482. [Google Scholar] [CrossRef] [PubMed]
- Wolff, M.; Vann, S.D. The Cognitive Thalamus as a Gateway to Mental Representations. J. Neurosci. 2019, 39, 3–14. [Google Scholar] [CrossRef]
- Szameitat, A.J.; McNamara, A.; Shen, S.; Sterr, A. Neural Activation and Functional Connectivity during Motor Imagery of Bimanual Everyday Actions. PLoS ONE 2012, 7, e38506. [Google Scholar] [CrossRef]
- Takeuchi, H.; Taki, Y.; Nouchi, R.; Yokoyama, R.; Kotozaki, Y.; Nakagawa, S.; Sekiguchi, A.; Iizuka, K.; Hanawa, S.; Araki, T.; et al. Originality of Divergent Thinking Is Associated with Working Memory–Related Brain Activity: Evidence from a Large Sample Study. NeuroImage 2020, 216, 116825. [Google Scholar] [CrossRef]
- Pinho, A.L.; de Manzano, O.; Fransson, P.; Eriksson, H.; Ullen, F. Connecting to Create: Expertise in Musical Improvisation Is Associated with Increased Functional Connectivity between Premotor and Prefrontal Areas. J. Neurosci. 2014, 34, 6156–6163. [Google Scholar] [CrossRef]
- Brain Networks Underlying Mental Imagery of Auditory and Visual Information—Zvyagintsev—2013—European Journal of Neuroscience—Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/ejn.12140 (accessed on 20 January 2023).
- Chen, X.; Liang, S.; Pu, W.; Song, Y.; Mwansisya, T.E.; Yang, Q.; Liu, H.; Liu, Z.; Shan, B.; Xue, Z. Reduced Cortical Thickness in Right Heschl’s Gyrus Associated with Auditory Verbal Hallucinations Severity in First-Episode Schizophrenia. BMC Psychiatry 2015, 15, 152. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J. The Human Imagination: The Cognitive Neuroscience of Visual Mental Imagery. Nature Reviews. Neuroscience 2019, 20, 624–634. [Google Scholar] [PubMed]
- Winlove, C.I.P.; Milton, F.; Ranson, J.; Fulford, J.; MacKisack, M.; Macpherson, F.; Zeman, A. The Neural Correlates of Visual Imagery: A Co-Ordinate-Based Meta-Analysis. Cortex 2018, 105, 4–25. [Google Scholar] [CrossRef] [PubMed]
- Pidgeon, L.M.; Grealy, M.; Duffy, A.H.B.; Hay, L.; McTeague, C.; Vuletic, T.; Coyle, D.; Gilbert, S.J. Functional Neuroimaging of Visual Creativity: A Systematic Review and Meta-analysis. Brain Behav. 2016, 6, e00540. [Google Scholar] [CrossRef]
- Solso, R.L. Brain Activities in a Skilled versus a Novice Artist: An FMRI Study. Leonardo 2001, 34, 31–34. [Google Scholar] [CrossRef]
- Varoquaux, G. Cross-Validation Failure: Small Sample Sizes Lead to Large Error Bars. Neuroimage 2018, 180, 68–77. [Google Scholar] [CrossRef]
- Vabalas, A.; Gowen, E.; Poliakoff, E.; Casson, A.J. Machine Learning Algorithm Validation with a Limited Sample Size. PLoS ONE 2019, 14, e0224365. [Google Scholar] [CrossRef]
- Turner, B.O.; Santander, T.; Paul, E.J.; Barbey, A.K.; Miller, M.B. Reply to: FMRI Replicability Depends upon Sufficient Individual-Level Data. Commun. Biol. 2019, 2, 129. [Google Scholar] [CrossRef]
- Morrison, R.G.; Wallace, B. Imagery Vividness, Creativity and the Visual Arts. J. Ment. Imag. 2001, 25, 135–152. [Google Scholar]
- Haynes, J.-D.; Rees, G. Decoding Mental States from Brain Activity in Humans. Nat. Rev. Neurosci. 2006, 7, 523–534. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grecucci, A.; Rastelli, C.; Bacci, F.; Melcher, D.; De Pisapia, N. A Supervised Machine Learning Approach to Classify Brain Morphology of Professional Visual Artists versus Non-Artists. Sensors 2023, 23, 4199. https://doi.org/10.3390/s23094199
Grecucci A, Rastelli C, Bacci F, Melcher D, De Pisapia N. A Supervised Machine Learning Approach to Classify Brain Morphology of Professional Visual Artists versus Non-Artists. Sensors. 2023; 23(9):4199. https://doi.org/10.3390/s23094199
Chicago/Turabian StyleGrecucci, Alessandro, Clara Rastelli, Francesca Bacci, David Melcher, and Nicola De Pisapia. 2023. "A Supervised Machine Learning Approach to Classify Brain Morphology of Professional Visual Artists versus Non-Artists" Sensors 23, no. 9: 4199. https://doi.org/10.3390/s23094199
APA StyleGrecucci, A., Rastelli, C., Bacci, F., Melcher, D., & De Pisapia, N. (2023). A Supervised Machine Learning Approach to Classify Brain Morphology of Professional Visual Artists versus Non-Artists. Sensors, 23(9), 4199. https://doi.org/10.3390/s23094199