D-galactose/D-glucose-binding Protein from Escherichia coli as Probe for a Non-consuming Glucose Implantable Fluorescence Biosensor

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
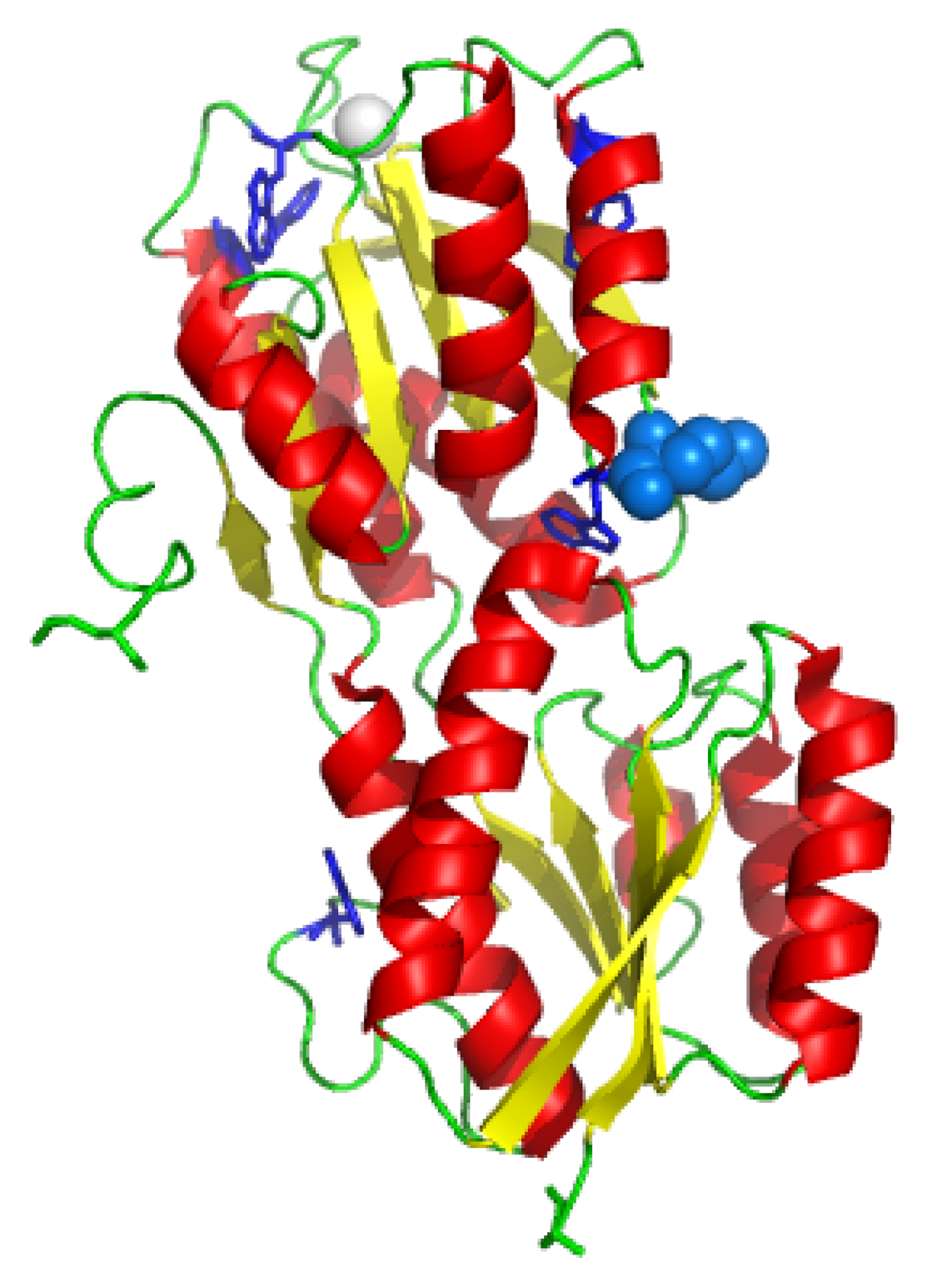
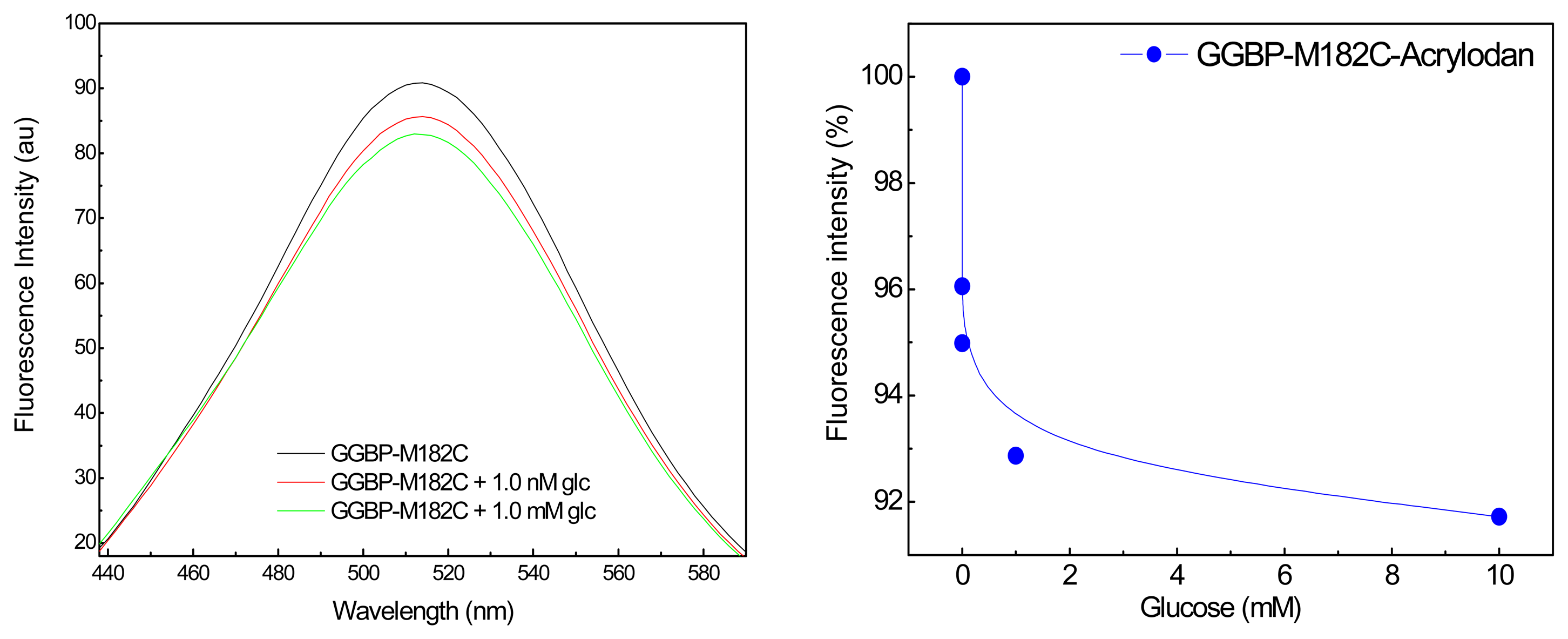
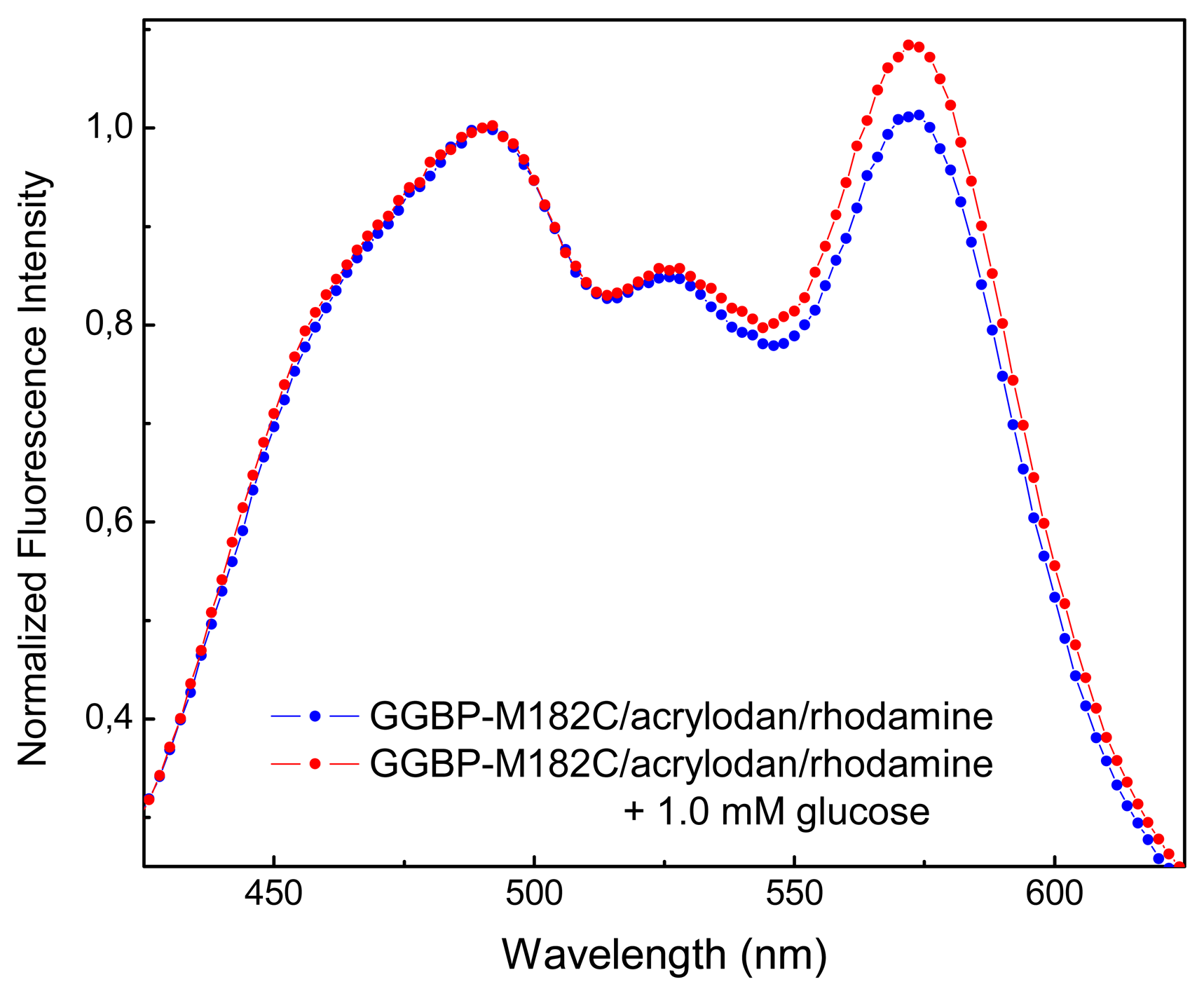
2. Results and Discussion
3. Conclusion
4. Experimental Section
4.1 Construction of the M182C Mutant of GGBP
4.2 Isolation of GGBP-M182C
4.3 Steady-State Fluorescence Spectroscopy
4.4 Fluorescence Labelling of GGBP-M182C
4.5 Frequency-domain Fluorescence Spectroscopy
Acknowledgments
References and Notes
- The Diabetes Control & Complications Trial Research Group. 1997.
- Ervin, K. R.; Kiser, E. J. Issues and implications in the selection of blood glucose monitoring technologies. Diabetes Technol. Ther. 1999, 1(1), 3–11. [Google Scholar]
- de Champdore, M.; Staiano, M.; Rossi, M.; D'Auria, S. Proteins from extremophiles as stable tools for advanced biotechnological applications of high social interest. J. R. Soc. Interface. 2007, 4(13), 183–191. [Google Scholar]
- D'Auria, S.; Alfieri, F.; Staiano, M.; Pelella, F.; Rossi, M.; Scirè, A.; Tanfani, F.; Bertoli, E.; Grycznyski, Z.; Lakowicz, J.R. Structural and thermal stability characterization of Escherichia coli D-galactose/D-glucose-binding protein. Biotechnol Prog. 2004, 20(1), 330–337. [Google Scholar]
- Vyas, N. K.; Vyas, M. N.; Quiocho, F. A. Sugar and signal-transducer binding sites of the Escherichia coli galactose chemoreceptor protein. Science 1988, 242, 1290–1295. [Google Scholar]
- Borrok, M. J.; Kiessling, L. L.; Forest, K. T. Conformational changes of D-glucose/D-galactose binding protein illuminated by apo and ultrahigh resolution ligand-bound structures; to be published.
- Quiocho, F.A. Atomic structures and function of periplasmic receptors for active transport and chemotaxis. Curr. Opin. Biol. 1991, 1, 922–933. [Google Scholar]
- Vyas, N. K.; Vyas, M. N.; Quiocho, F.A. Comparison of the periplasmic receptors for L-arabinose, D-glucose/D-galactose and D-ribose – structural and functional similarity. J Biol. Chem. 1991, 266, 5226–5237. [Google Scholar]
- Flocco, M. M.; Mowbray, S. L. The 1.9 Å X-ray structure of a closed unliganded form of the periplasmic glucose/galactose receptor from Salmonella typhimurium. J. Biol. Chem. 1994, 269, 8931–8936. [Google Scholar]
- Salins, L.L.; Ware, R.A.; Ensor, C.M.; Daunert, S. A novel reagentless sensing system for measuring glucose based on galactose/glucose-binding protein. Anal. Biochem. 2001, 294, 19–26. [Google Scholar]
- Zukin, R. S.; Strange, P. G.; Heavey, L. R.; Koshland, D. E., Jr. Properties of the galactose binding protein of Salmonella typhimurium and Escherichia coli. Biochemistry 1977, 16, 381–386. [Google Scholar]
- Scognamiglio, V.; Scirè, A.; Aurilia, V.; Staiano, M.; Crescenzo, R.; Palmucci, C.; Bertoli, E.; Rossi, M.; Tanfani, F.; D'Auria, S. A strategic fluorescence labelling of D-galactose/D-glucose-binding protein from E. coli for a better understanding of the protein structural stability and dynamics. in press.
- Neu, H. C.; Heppel, L. A. The Release of Enzymes from Kwherichia coli by Osmotic Shock and during the Formation of Spheroplasts. J. Biol. Chem. 1965, 240, 9. [Google Scholar]




© 2007 by MDPI ( http://www.mdpi.org). Reproduction is permitted for noncommercial purposes.
Share and Cite
Scognamiglio, V.; Aurilia, V.; Cennamo, N.; Ringhieri, P.; Iozzino, L.; Tartaglia, M.; Staiano, M.; Ruggiero, G.; Orlando, P.; Labella, T.; et al. D-galactose/D-glucose-binding Protein from Escherichia coli as Probe for a Non-consuming Glucose Implantable Fluorescence Biosensor. Sensors 2007, 7, 2484-2491. https://doi.org/10.3390/s7102484
Scognamiglio V, Aurilia V, Cennamo N, Ringhieri P, Iozzino L, Tartaglia M, Staiano M, Ruggiero G, Orlando P, Labella T, et al. D-galactose/D-glucose-binding Protein from Escherichia coli as Probe for a Non-consuming Glucose Implantable Fluorescence Biosensor. Sensors. 2007; 7(10):2484-2491. https://doi.org/10.3390/s7102484
Chicago/Turabian StyleScognamiglio, Viviana, Vincenzo Aurilia, Nunzio Cennamo, Paola Ringhieri, Luisa Iozzino, Micaela Tartaglia, Maria Staiano, Giuseppe Ruggiero, Pierangelo Orlando, Tullio Labella, and et al. 2007. "D-galactose/D-glucose-binding Protein from Escherichia coli as Probe for a Non-consuming Glucose Implantable Fluorescence Biosensor" Sensors 7, no. 10: 2484-2491. https://doi.org/10.3390/s7102484
APA StyleScognamiglio, V., Aurilia, V., Cennamo, N., Ringhieri, P., Iozzino, L., Tartaglia, M., Staiano, M., Ruggiero, G., Orlando, P., Labella, T., Zeni, L., Vitale, A., & D’Auria, S. (2007). D-galactose/D-glucose-binding Protein from Escherichia coli as Probe for a Non-consuming Glucose Implantable Fluorescence Biosensor. Sensors, 7(10), 2484-2491. https://doi.org/10.3390/s7102484