From Hearing to Listening: Design and Properties of an Actively Tunable Electronic Hearing Sensor

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental setup
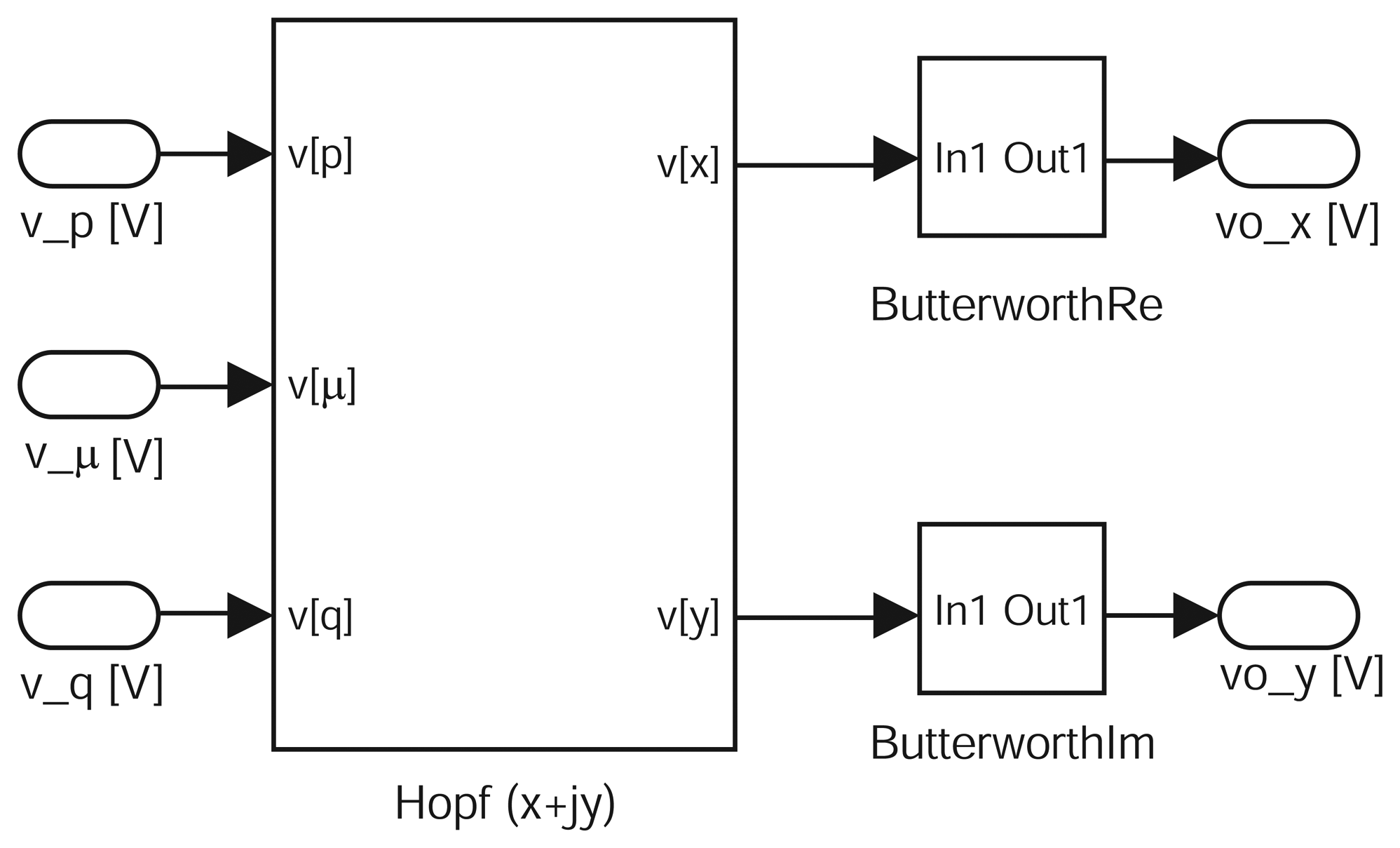
Electronic sensor design
3. Results
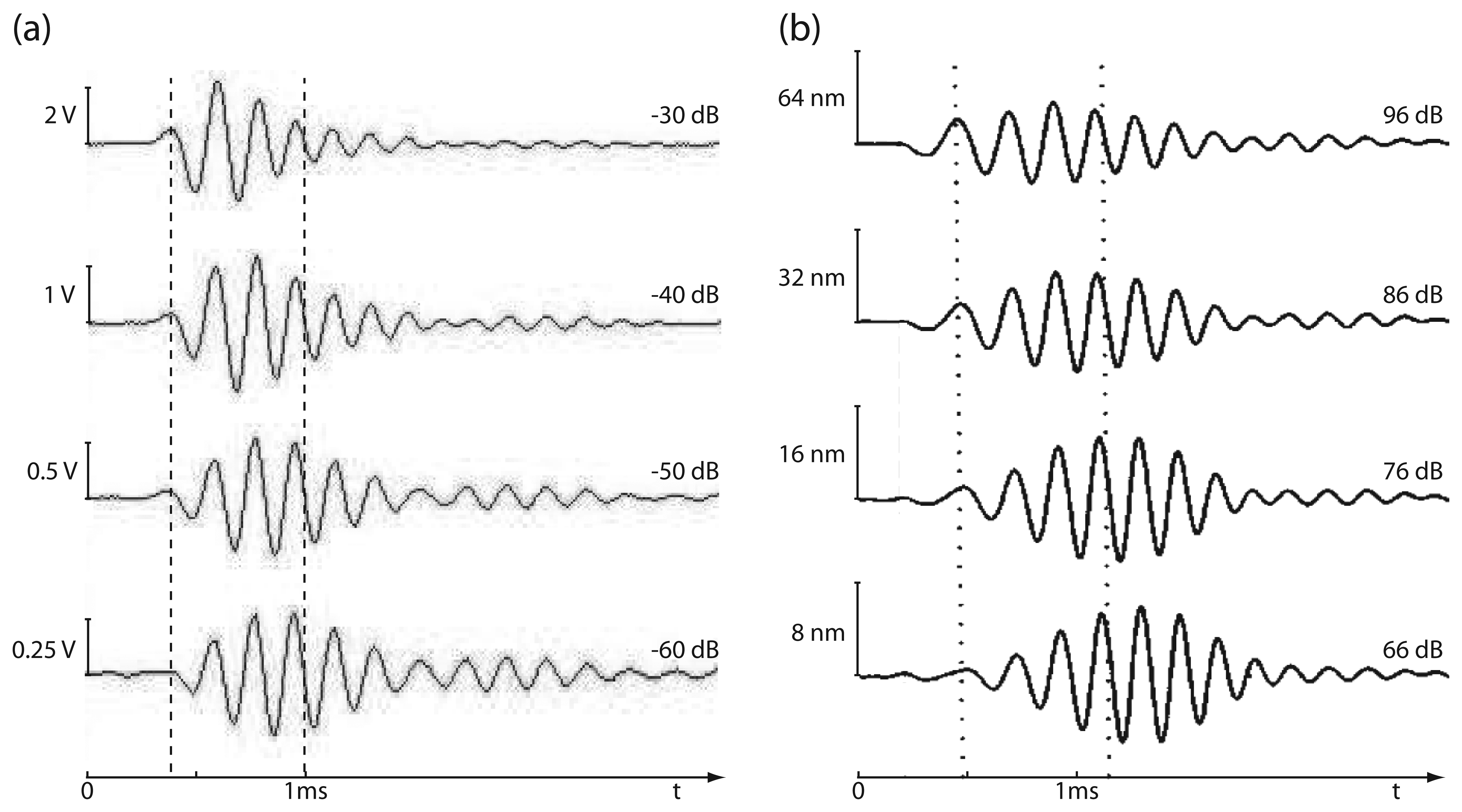
Steady-state signal response
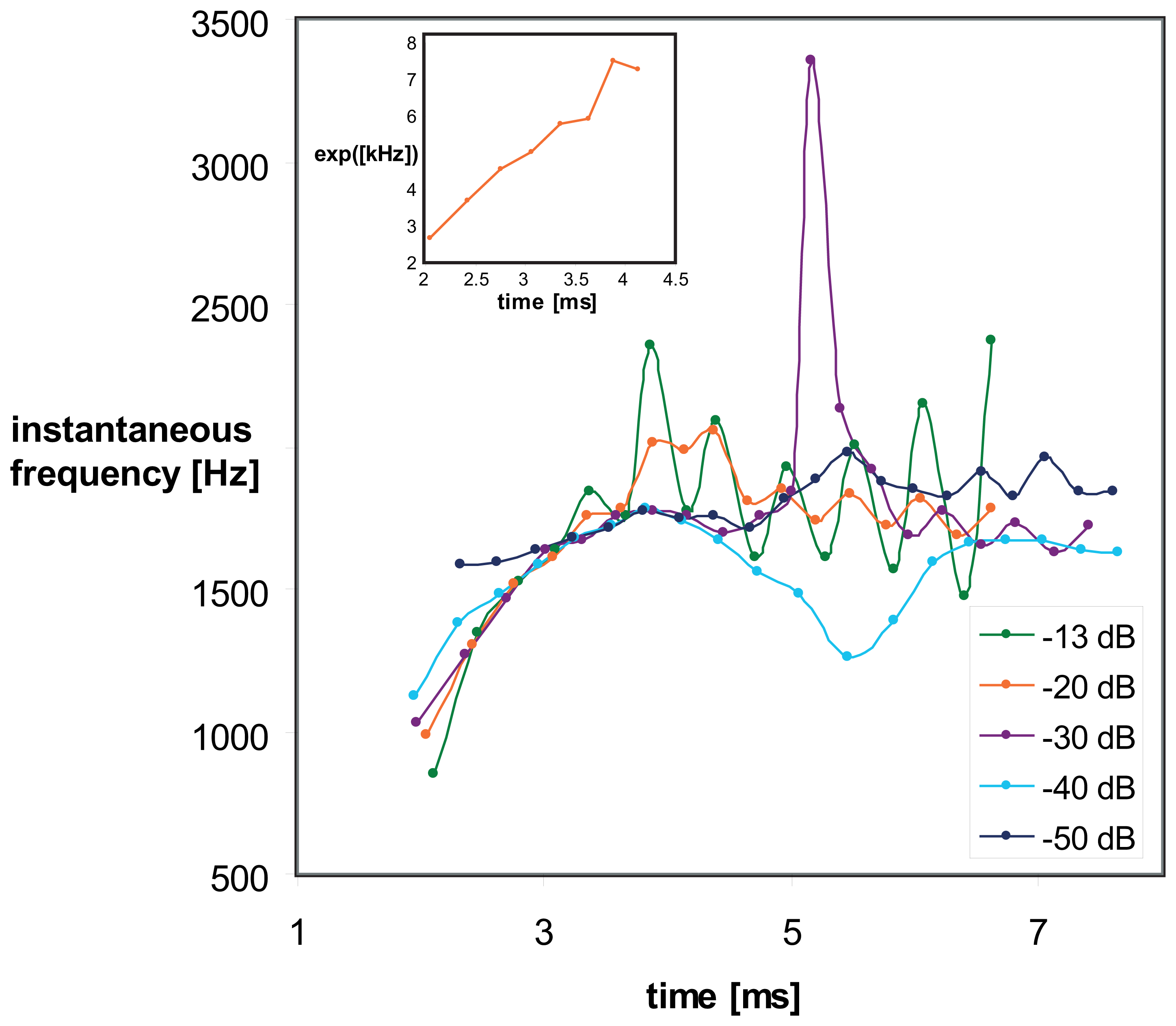
Transient signal response
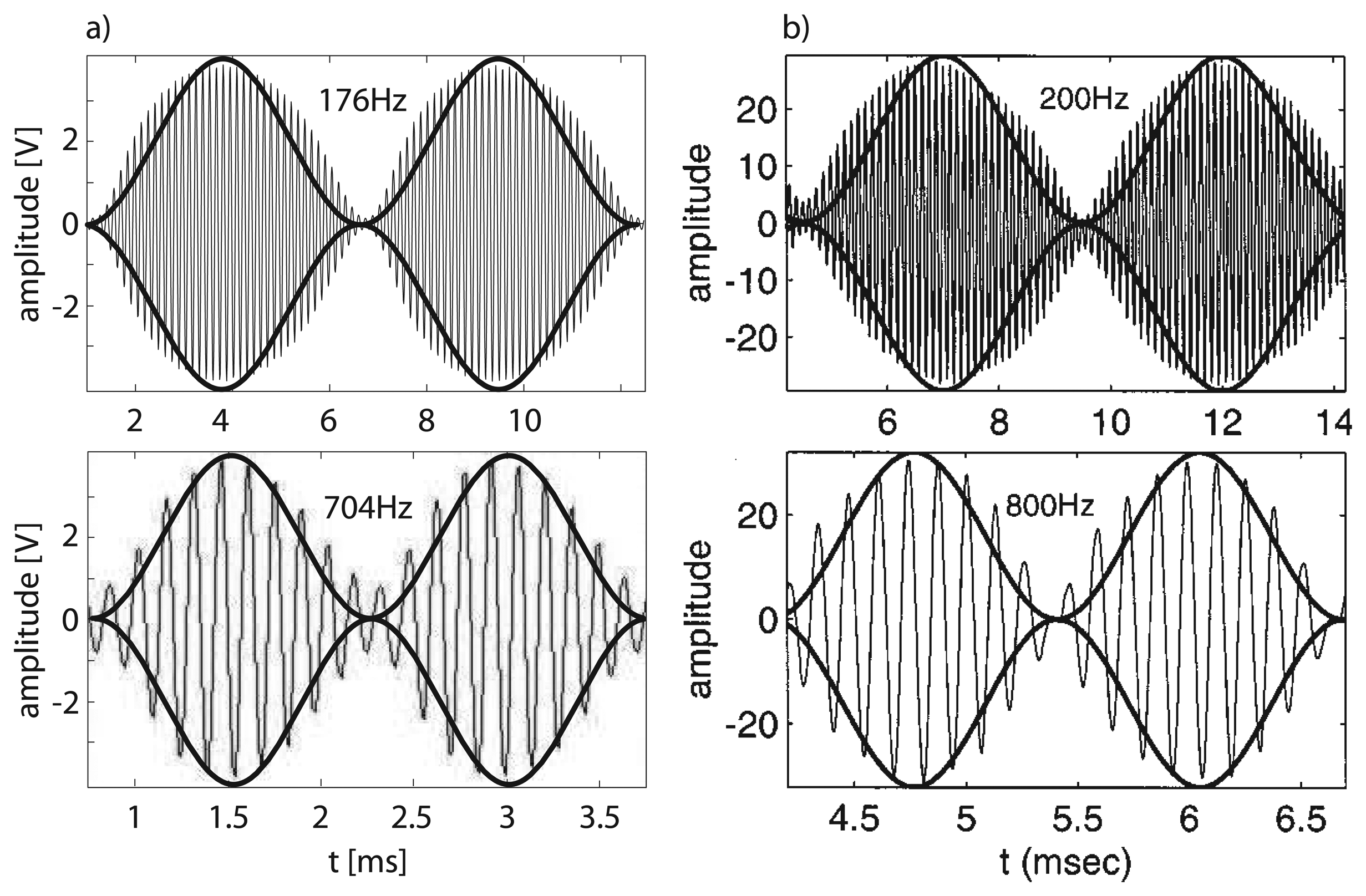
Active tuning towards signals
4. Conclusions
Acknowledgments
References
- Helmholtz, H.L.F. Die Lehre von den Tonempfindungen als physiologische Grundlage für die Theorie der Musik; Vieweg, Braunschweig: Germany, 1863. [Google Scholar]
- von Békésy, G. Zur Theorie des Hörens. Die Schwingungsform der Basilarmembran. Phys. Z. 1928, 29, 793–810. [Google Scholar]
- Gold, T. Hearing. The physical basis of the action of the cochlea. Proc. R. Soc. London Biol. Sci. 1948, 135, 492–498. [Google Scholar]
- Kemp, D.T. Stimulated acoustic emissions from within the human auditory system. J. Acoust. Soc. Am. 1978, 64, 1386–1391. [Google Scholar]
- Brownell, W.E.; Bader, C.R.; Bertrand, D.; de Ribaupierre, Y. Evoked mechanical responses of isolated cochlear outer hair cells. Science 1985, 227, 194–196. [Google Scholar]
- Robles, L.; Ruggero, M.A. Mechanics of the Mammalian Cochlea. Physiological Reviews 2001, 81, 1305–1352. [Google Scholar]
- Wiesenfeld, K.; McNamara, B. Period-doubling systems as small-signal amplifiers. Phys. Rev. Lett. 1985, 55, 13–16. [Google Scholar]
- Derighetti, B.; Ravani, M.; Stoop, R.; Meier, P.F.; Brun, E.; Badii, R. Period-doubling lasers as small-signal detectors. Phys. Rev. Lett. 1985, 55, 1746–1748. [Google Scholar]
- Ruggero, M.A.; Temchin, A.N. Unexceptional sharpness of frequency tuning in the human cochlea. Proc. Natl. Acad. Sci. U.S.A. 2005, 102, 18614–18619. [Google Scholar]
- Geisler, C.D. From Sound to Synapse; Oxford Univ. Press: Oxford, UK, 1998. [Google Scholar]
- Eguíluz, V.M.; Ospeck, M.; Choe, Y.; Hudspeth, A.J.; Magnasco, M.O. Essential nonlinearities in hearing. Phys. Rev. Lett. 2000, 84, 5232–5535. [Google Scholar]
- Camalet, S.; Duke, T.; Jülicher, F.; Prost, J. Auditory sensitivity provided by self-tuned critical oscillations of hair cells. Proc. Natl. Acad. Sci. U.S.A. 2000, 97, 3183–3188. [Google Scholar]
- Göpfert, M.C.; Humphris, A.D.L.; Albert, J.T.; Robert, D.; Hendrich, O. Power gain exhibited by motile mechanosensory neurons in Drosophila ears. Proc. Natl. Acad. Sci. U.S.A. 2005, 102, 325–330. [Google Scholar]
- Martin, P.; Hudspeth, A.J. Compressive nonlinearity in the hair bundle's active response to mechanical stimulation. Proc. Natl. Acad. Sci. U.S.A. 2001, 98, 14386–14391. [Google Scholar]
- de Boer, E. The Cochlea. Springer Handbook of Auditory Research; Dallos, P., Popper, A.N., Fay, R.R., Eds.; Springer: New York, USA, 1996; pp. 258–317. [Google Scholar]
- Peterson, B.P.; Bogert, L.C. A dynamical theory of the cochlea. J. Acoust. Soc. Am. 1950, 22, 369–381. [Google Scholar]
- Kern, A.; Stoop, R. Essential Role of Couplings between Hearing Nonlinearities. Phys. Rev. Lett. 2003, 91, 128101. [Google Scholar]
- Whitham, G.B. Linear and Nonlinear Waves; Interscience Publishers: New York, USA, 1999. [Google Scholar]
- Lighthill, J. Waves in Fluids; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Smith, R.J.; Dorf, R.C. Circuits, Devices and Systems; John Wiley & Sons: New York, USA, 1991. [Google Scholar]
- Zwislocki, J.J.; Nguyen, M. Place Code for Pitch: A Necessary Revision. Acta Otolaryngol. (Stockh.) 1999, 119, 140–145. [Google Scholar]
- Ruggero, M.A. Responses to sound of the basilar membrane of the mammalian cochlea. Curr. Opin. Neurobiol. 1992, 2, 449–456. [Google Scholar]
- Ruggero, M.A.; Narayan, S.S.; Temchin, A.N.; Recio, A. Mechanical bases of frequency tuning and neural excitation at the base of the cochlea. Proc. Natl. Acad. Sci. U.S.A. 2000, 97, 11744–11750. [Google Scholar]
- Robles, L.; Ruggero, M.A.; Rich, N.C. Two-tone distortion on the basilar membrane of the chinchilla cochlea. J. Neurophysiol. 1997, 77, 2385–2399. [Google Scholar]
- Ruggero, M.A.; Robles, L.; Rich, N.C. Two-tone suppression in the basilar membrane of the cochlea. J. Neurophysiol. 1992, 68, 1087–1099. [Google Scholar]
- Stoop, R.; Kern, A. Essential auditory contrast-sharpening is preneuronal. Proc. Natl. Acad. Sci. U.S.A. 2004, 101, 9179–9181. [Google Scholar]
- Stoop, R.; Kern, A. Two-tone suppression and combination tone generation as computations performed by the Hopf cochlea. Phys. Rev. Lett. 2004, 93, 268103. [Google Scholar]
- Recio, A.; Rhode, W.S. Basilar membrane responses to broadband stimuli. J. Acoust. Soc. Am. 2000, 108, 2281–2298. [Google Scholar]
- Rhode, W.S.; Recio, A. Basilar-membrane response to multicomponent stimuli in chinchilla. J. Acoust. Soc. Am. 2001, 110, 981–994. [Google Scholar]








© 2007 by MDPI http://www.mdpi.org
Share and Cite
Stoop, R.; Jasa, T.; Uwate, Y.; Martignoli, S. From Hearing to Listening: Design and Properties of an Actively Tunable Electronic Hearing Sensor. Sensors 2007, 7, 3287-3298. https://doi.org/10.3390/s7123287
Stoop R, Jasa T, Uwate Y, Martignoli S. From Hearing to Listening: Design and Properties of an Actively Tunable Electronic Hearing Sensor. Sensors. 2007; 7(12):3287-3298. https://doi.org/10.3390/s7123287
Chicago/Turabian StyleStoop, Ruedi, Tom Jasa, Yoko Uwate, and Stefan Martignoli. 2007. "From Hearing to Listening: Design and Properties of an Actively Tunable Electronic Hearing Sensor" Sensors 7, no. 12: 3287-3298. https://doi.org/10.3390/s7123287
APA StyleStoop, R., Jasa, T., Uwate, Y., & Martignoli, S. (2007). From Hearing to Listening: Design and Properties of an Actively Tunable Electronic Hearing Sensor. Sensors, 7(12), 3287-3298. https://doi.org/10.3390/s7123287