Assessing Steady-state Fluorescence and PRI from Hyperspectral Proximal Sensing as Early Indicators of Plant Stress: The Case of Ozone Exposure


Abstract
:1. Introduction
2. RS of energy dissipation pathways
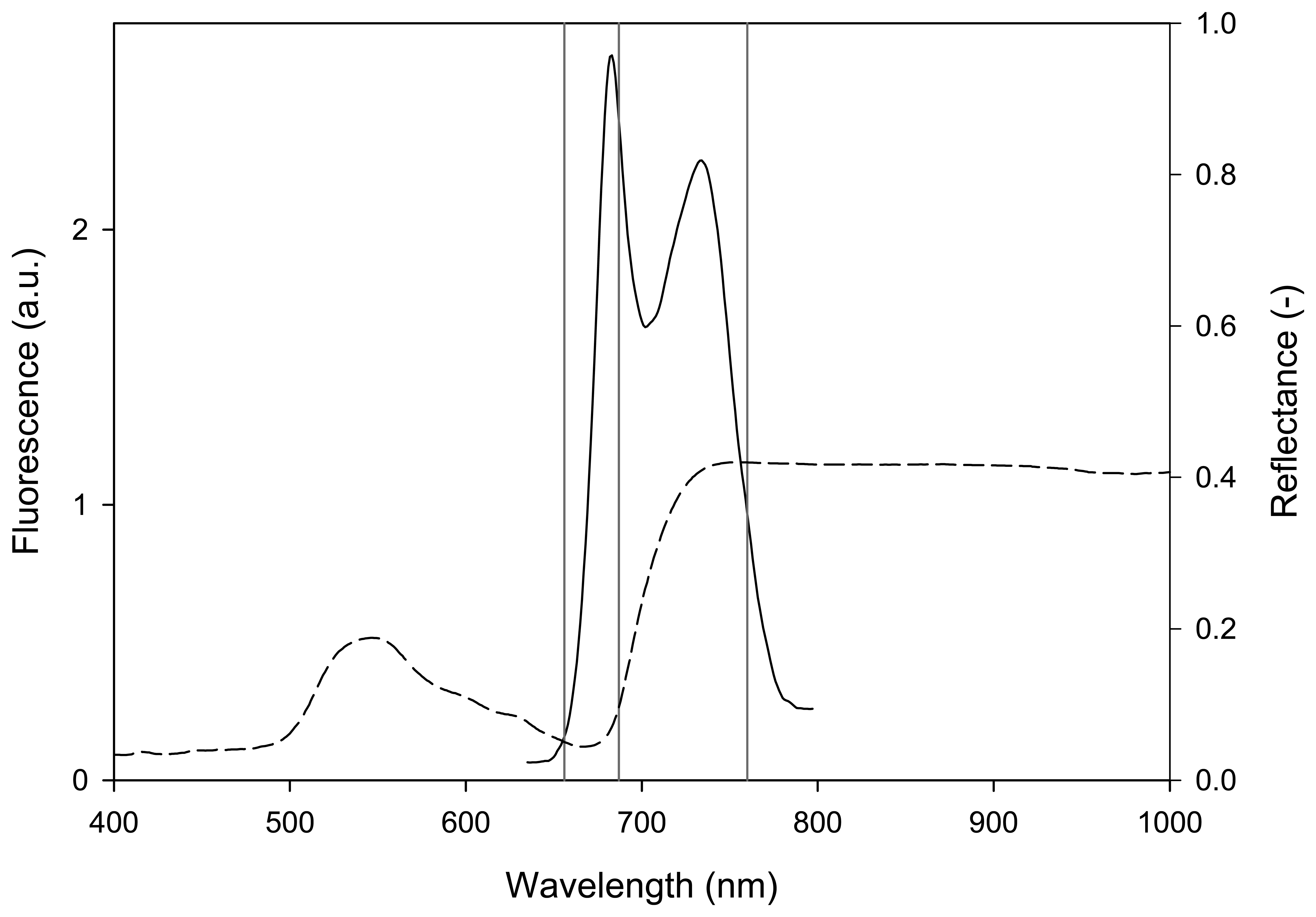
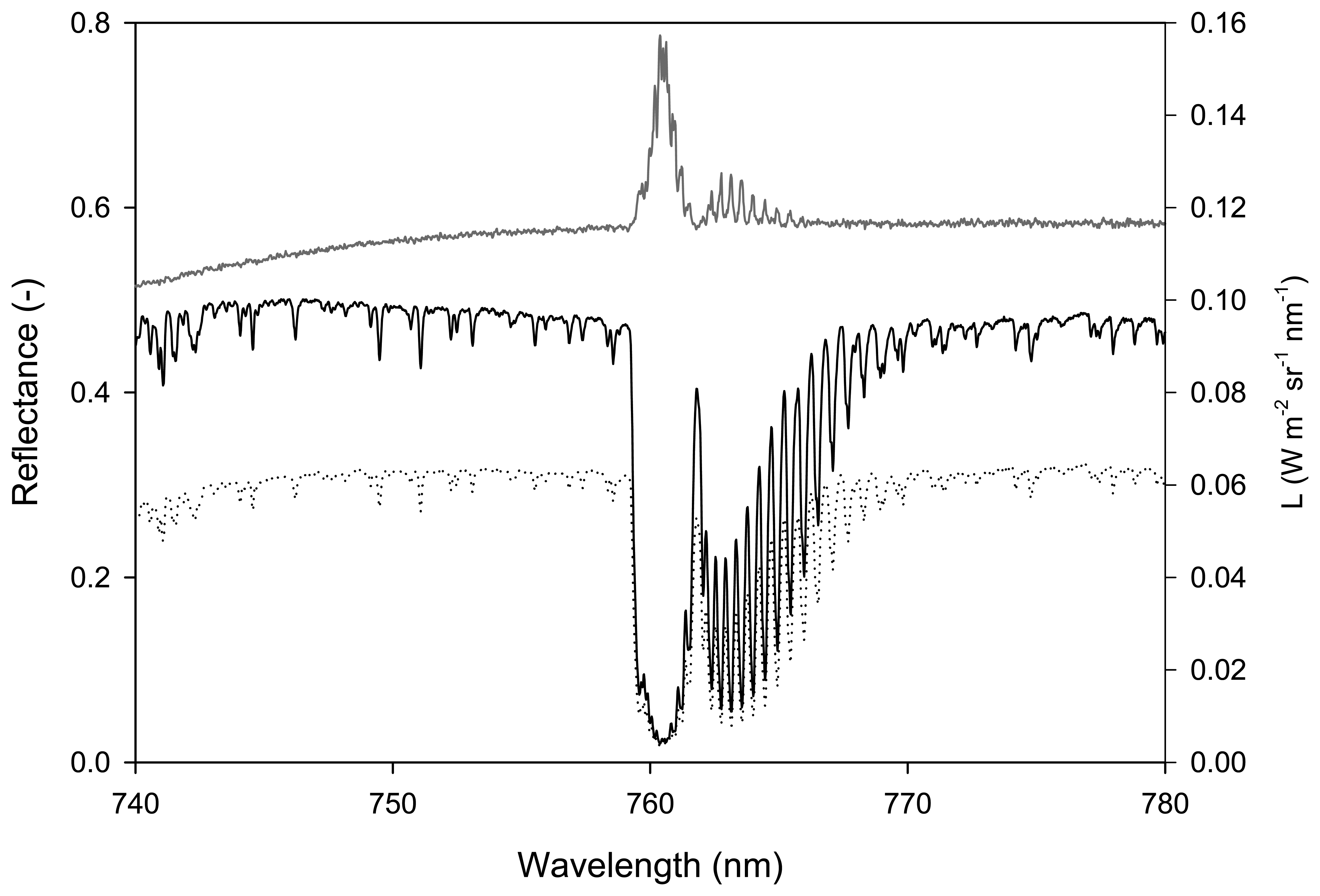
2.1 Steady-state fluorescence
2.2 Photochemical Reflectance Index (PRI)
3. Materials and methods
3.1 Plant material and ozone exposure
3.2 Data collection
3.2.1 Spectral measurements
3.2.2 Physiological measurements and meteorological dataset
3.3 Statistical analysis
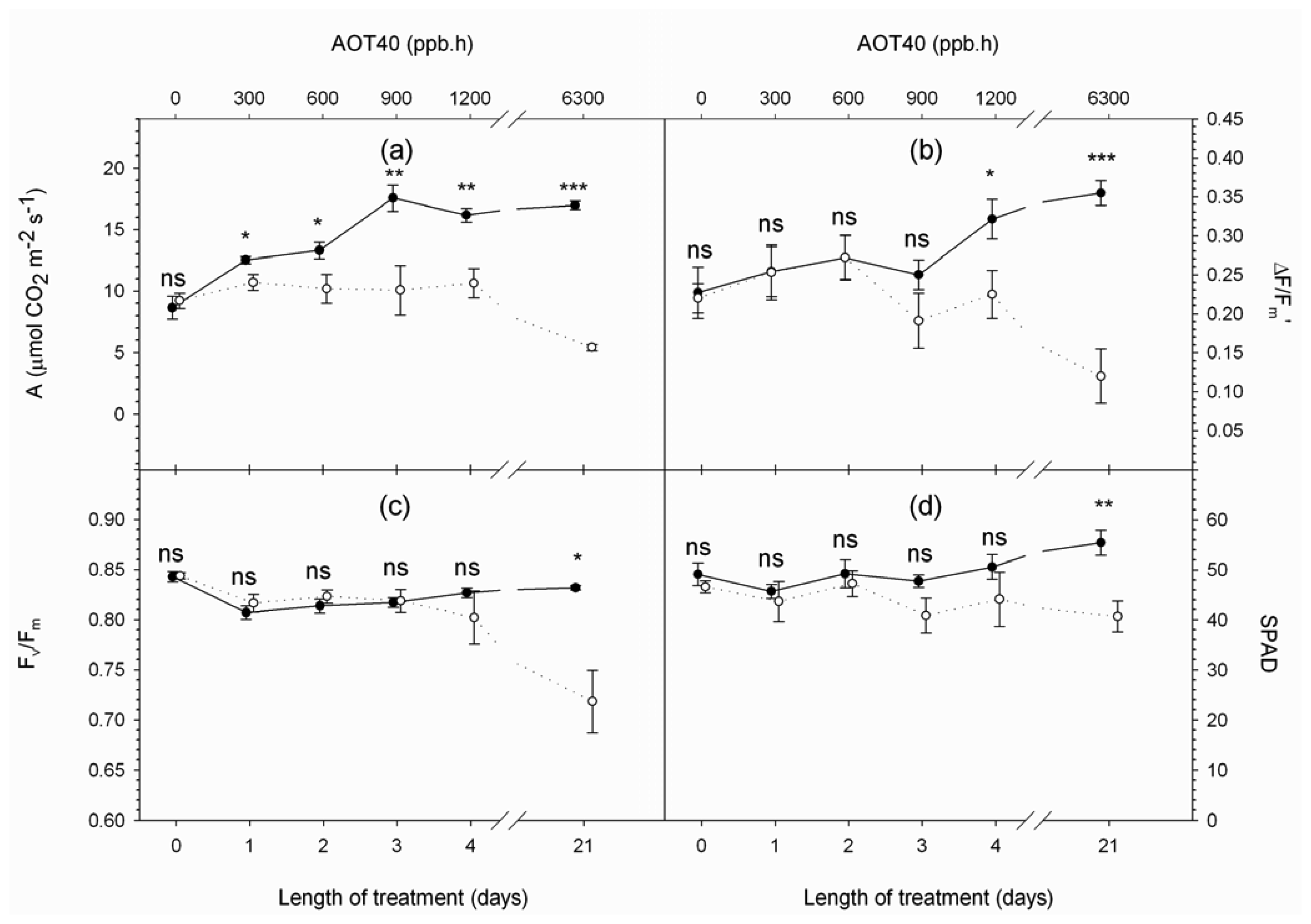
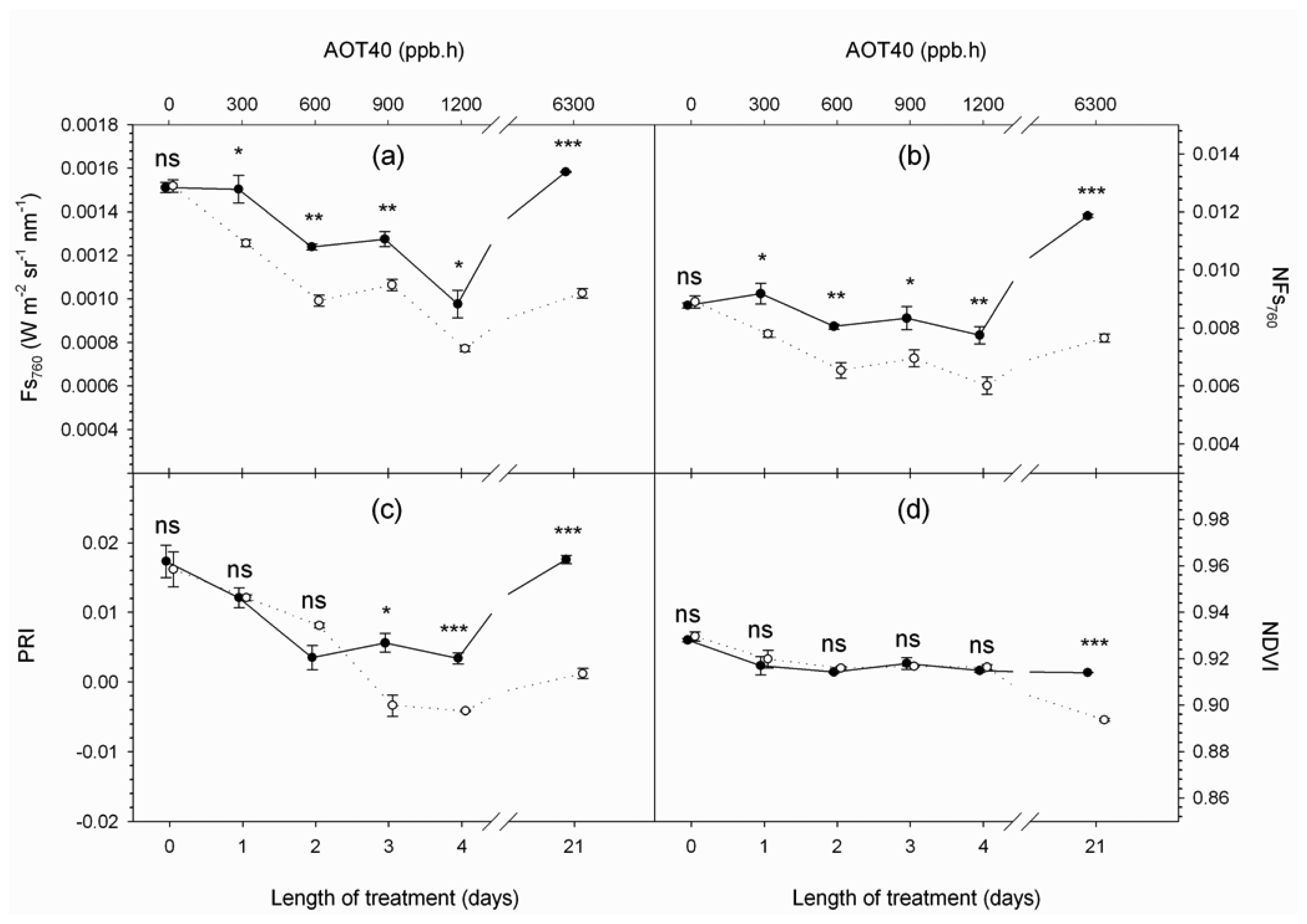
4. Results and Discussion
5. Conclusions
Acknowledgments
References and Notes
- Lichtenthaler, H.K. The stress concept in plants: an introduction. In Stress Of Life: From Molecules To Man; Csermely, P., Ed.; Ann. NY Acad. Sci; 1998; pp. 187–198. [Google Scholar]
- Zarco-Tejada, P.J.; Miller, J.R.; Mohammed, G.H.; Noland, T.L.; Sampson, P.H. Vegetation stress detection through chlorophyll a + b estimation and fluorescence effects on hyperspectral imagery. J. Environ. Qual. 2002, 31, 1433–1441. [Google Scholar]
- Sampson, P.H.; Zarco-Tejada, P.J.; Mohammed, G.H.; Miller, J.R.; Noland, T.L.; Fleming, R.L. Hyperspectral remote sensing of forest condition: estimation of chlorophyll content in tolerant hardwoods. For. Sci. 2003, 49, 381–391. [Google Scholar]
- Coops, N.C.; Stone, C.; Culvenor, D.S.; Chisholm, L. Assessment of crown condition in eucalypt vegetation by remotely sensed optical indices. J. Environ. Qual. 2004, 33, 956–964. [Google Scholar]
- Rossini, M.; Panigada, C.; Meroni, M.; Colombo, R. Assessment of oak forest condition based on leaf biochemical variables and chlorophyll fluorescence. Tree Physiol. 2006, 26, 1487–1496. [Google Scholar]
- Baret, F.; Houlès, V.; Guérif, M. Quantification of plant stress using remote sensing observations and crop models: the case of nitrogen management. J. Exp. Bot. 2007, 58, 869–880. [Google Scholar]
- Björkman, O.; Demmig-Adams, B. Regulation of photosynthetic light energy capture, conversion, and dissipation in leaves of higher plants. In Ecophysiology of Photosynthesis; Schulze, E.D., Caldwell, M.M., Eds.; Springer-Verlag: Berlin, 1995; pp. 17–47. [Google Scholar]
- Govindjee Chlorophyll a fluorescence: a bit of basics and history. In Chlorophyll a Fluorescence: A Signature of Photosynthesis. Advances in Photosynthesis and Respiration; Papageorgiou, G.C.; Govindjee (Eds.) Springer: Dordrecht, 2004; Vol. 19, pp. 1–42.
- Apel, K.; Hirt, H. Reactive oxygen species: oxidative stress and signal transduction. Ann. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar]
- Iriti, M.; Faoro, F. Oxidative Stress, the Paradigm of Ozone Toxicity in Plants and Animals. Water Air Soil Poll. 2008, 187, 285–301. [Google Scholar]
- Plascyk, J.A. The MK II Fraunhofer line discriminator (FLD-II) for airborne and orbital remote sensing of solar-stimulated luminescence. Opt. Eng. 1975, 14, 339–346. [Google Scholar]
- Moya, I.; Camenen, L.; Evain, S.; Goulas, Y.; Cerovic, Z.G.; Latouche, G.; Flexas, J.; Ounis, A. A new instrument for passive remote sensing 1. Measurements of sunlight-induced chlorophyll fluorescence. Remote Sens. Environ. 2004, 91, 186–197. [Google Scholar]
- Meroni, M.; Colombo, R. Leaf level detection of solar induced chlorophyll fluorescence by means of a subnanometer resolution spectroradiometer. Remote Sens. Environ. 2006, 103, 438–448. [Google Scholar]
- Elachi, C. Introduction to the Physics and Techniques of Remote Sensing; John Wiley and Sons: New York, 1987; p. 63. [Google Scholar]
- Plascyk, J.A.; Gabriel, F.C. The Fraunhofer Line Discriminator MKII- an airborne instrument for precise and standardized ecological luminescence measurements. IEEE Trans. Instrum. Meas. 1975, 24, 306–313. [Google Scholar]
- Demmig-Adams, B.; Adams, W.W. Photoprotection and other responses of plants to light stress. Ann. Rev. Plant Physiol. Mol. Biol. 1992, 43, 599–626. [Google Scholar]
- Horton, P.; Ruban, A.V.; Walters, R.G. Regulation of light harvesting in green plants. Ann. Rev. Plant Physiol. Mol. Biol. 1996, 47, 655–684. [Google Scholar]
- Li, X.P.; Björkman, O.; Shih, C.; Grossman, A.R.; Rosenquist, M.; Jansson, S.; Niyogi, K.K. A pigment-binding protein essential for regulation of photosynthetic light harvesting. Nature 2000, 403, 391–395. [Google Scholar]
- Horton, P.; Wentworth, M.; Ruban, A. Control of the light harvesting function of chloroplast membranes: The LHCII-aggregation model for non-photochemical quenching. FEBS Lett. 2005, 579, 4201–4206. [Google Scholar]
- Yamamoto, H.Y. Biochemistry of the violaxanthin in higher plants. Pure Appl. Chem. 1979, 51, 639–648. [Google Scholar]
- Bilger, W.; Björkman, O.; Thayer, S.S. Light-induced spectral absorbance changes in relation to photosynthesis and the epoxidation state of xanthophyll cycle components in cotton leaves. Plant Physiol. 1989, 91, 542–551. [Google Scholar]
- Gamon, J.A.; Field, C.B.; Bilger, W.; Björkman, O.; Fredeen, A.L.; Peñuelas, J. Remote-sensing of the xanthophyll cycle and chlorophyll fluorescence in sunflower leaves and canopies. Oecologia 1990, 85, 1–7. [Google Scholar]
- Gamon, J.A.; Peñuelas, J.; Field, C.B. A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency. Rem. Sens. Environ. 1992, 41, 35–44. [Google Scholar]
- Nali, C.; Pucciariello, C.; Mills, G.; Lorenzini, G. On the different sensitivity of white clover clones to ozone: Physiological and biochemical parameters in a multivariate approach. Water Air Soil Poll. 2005, 164, 137–153. [Google Scholar]
- Kärenlampi, L.; Skärby, L. Critical levels for ozone in Europe: testing and finalizing the concepts. In UN-ECE Workshop Report; Kärenlampi, L., Skärby, L., Eds.; University of Kuopio, Department of Ecology and Environmental Science: Kuopio, Finland, 1996. [Google Scholar]
- Amoros-Lopez, J.; Gomez-Chova, L.; Vila-Frances, J.; Calpe, J.; Alonso, L.; Moreno, J.; del Valle-Tascon, S. Study of the diurnal cycle of stressed vegetation for the improvement of fluorescence remote sensing. In Remote Sensing for Agriculture, Ecosystems, and Hydrology VIII; Owe, M., D'Urso, G., Neale, C.M., Gouweleeuw, B.T, Eds.; Proc. SPIE; 2006; Volume 6359, p. 63590R. [Google Scholar]
- Milton, E.J.; Rolling, E.M. Estimating the irradiance spectrum from measurements in a limited number of spectral bands. Rem. Sens. Environ. 2006, 100, 348–355. [Google Scholar]
- Meroni, M.; Colombo, R. 3S: A novel program for field Spectrometry at Subnanometer Spectral resolution. Comput. Geosci.-UK 2008. submitted. [Google Scholar]
- Meroni, M.; Colombo, R.; Cogliati, S. High resolution leaf spectral signature for the detection of solar induced chlorophyll fluorescence. In Proc. 2nd Int. Workshop Rem. Sens. Veget. Fluores.; ESA Publications Division,Canadian Space Agency: Montreal, QC, Canada, 2004. [Google Scholar]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. BBA 1989, 990, 87–92. [Google Scholar]
- Hayes, F.; Mills, G.; Harmens, H.; Novak, K.; Williams, P. ICP Vegetation experimental protocol for monitoring the incidences of ozone injury on vegetation; Natural Environment Research Council: Bangor, U.K., 2006; Available from icpvegetation.ceh.ac.uk. [Google Scholar]
- Guidi, L.; Nali, C.; Ciompi, S.; Lorenzini, G.; Franco, G. The use of chlorophyll fluorescence and leaf gas exchange as methods for studying the different responses to ozone of two bean cultivars. J. Exp. Bot. 1997, 48, 173–179. [Google Scholar]
- Calatayud, A.; Ramirez, J.W.; Iglesias, D.J.; Barreno, E. Effects of ozone on photosynthetic CO2 exchange, chlorophyll a fluorescence and antioxidant systems in lettuce leaves. Physiol. Planta. 2002, 116, 308–316. [Google Scholar]
- Calatayud, A.; Iglesias, D.J.; Talón, M.; Barreno, E. Effects of 2-month ozone exposure in spinach leaves on photosynthesis, antioxidant systems and lipid peroxidation. Plant Physiol. Biochem. 2003, 4, 839–845. [Google Scholar]
- Guidi, L.; Degl'Innocenti, E.; Genovesi, S.; Soldatini, G.F. Photosynthetic process and activities of enzymes involved in the phenylpropanoid pathway in resistant and sensitive genotypes of Lycopersicon esculentum L. exposed to ozone. Plant Sci. 2005, 168, 153–160. [Google Scholar]
- Degl'Innocenti, E.; Guidi, L.; Soldatini, G.F. Effects of elevated ozone on chlorophyll a fluorescence in symptomatic and asymptomatic leaves of two tomato genotypes. Biol. Planta. 2007, 51, 313–321. [Google Scholar]
- Paoletti, E. Ozone impacts on forests. CAB Rev.: Persp. Agric. Vet. Sci. Nutr. Nat. Resour. 2007, 2, 1–13. [Google Scholar]
- Krause, GH. Photo-inhibition of photosynthesis. An evaluation of damaging and protective mechanisms. Physiol. Planta. 1988, 74, 566–574. [Google Scholar]
- Reichenauer, T.G.; Goodman, B.A.; Kostecki, P.; Soja, G. Ozone sensitivity in Triticum durum and Triticum aestivum with respect to leaf injury, photosynthetic activity and free radical content. Physiol. Planta. 1998, 104, 681–686. [Google Scholar]
- Soldatini, G.F.; Lorenzini, G.; Filippi, F.; Nali, C.; Guidi, L. Photosynthesis of two poplar clones under long-term exposure to ozone. Physiol. Planta. 1998, 104, 707–712. [Google Scholar]
- Shavnin, S.; Maurer, S.; Matyssek, R.; Bilger, W.; Sheidegger, C. The impact of ozone fumigation and fertilization on chlorophyll fluorescence of birch leaves (Betula pendula). Trees 1999, 14, 10–16. [Google Scholar]
- Castagna, A.; Nali, C.; Ciompi, S.; Lorenzini, G.; Soldatini, G.F.; Ranieri, A. Ozone exposure affects photosynthesis of pumpkin (Cucurbita pepo) plants. New Phytol. 2001, 152, 223–229. [Google Scholar]
- Elvira, S.; Alonso, R.; Castillo, F.J.; Gimeno, B.S. On the responses of pigments and antioxidants of Pinus halepensis seedlings to Mediterranean climatic factors and long-term ozone exposure. New Phytol. 1998, 138, 419–432. [Google Scholar]
- Ranieri, A.; Giuntini, D.; Ferraro, F.; Nali, C.; Baldan, B.; Lorenzini, G.; Soldatini, G.F. Chronic ozone fumigation induces alterations in thylakoid functionality and composition in two poplar clones. Plant Physiol. Biochem. 2001, 39, 999–1008. [Google Scholar]
- Rudorff, B.F.T.; Mulchi, C.L.; Daughtry, C.S.T.; Lee, E.B. Growth and radiation use efficiency of wheat and corn grown under elevated ozone and carbon dioxide atmospheres. Remote Sens. Environ. 1996, 55, 163–173. [Google Scholar]
- Kraft, M.; Weigel, H.J.; Mejer, G.J.; Brandes, F. Reflectance measurements of leaves for detecting visible and non-visible ozone damage to crops. J. Plant Physiol. 1996, 148, 148–154. [Google Scholar]
- Meroni, M.; Picchi, V.; Rossini, M.; Cogliati, S.; Panigada, C.; Nali, C.; Lorenzini, G.; Colombo, R. Leaf level early assessment of ozone injuries by passive fluorescence and PRI. Int. J. Remote Sens. 2008, in press. [Google Scholar]
- Richardson, A.D.; Berlyn, G.P.; Gregoire, T.G. Spectral reflectance of Picea rubens (Pinaceae) and Abies balsamea (Pinaceae) needles along an elevational gradient. Mt. Moosilauke: New Hampshire, USA; In Am. J. Bot.; 2001; Volume 88, pp. 667–676. [Google Scholar]
- Dobrowski, S.Z.; Pushnik, J.C.; Zarco-Tejada, P.J.; Ustin, S.L. Simple reflectance indices track heat and water stress-induced changes in steady-state chlorophyll fluorescence at the canopy scale. Remote Sens. Environ. 2005, 97, 403–414. [Google Scholar]
- Pérez-Priego, O.; Zarco-Tejada, P.J.; Miller, J.R.; Sepulcre-Cantó, G.; Fereres, E. Detection of water stress in orchard trees with a high-resolution spectrometer through chlorophyll fluorescence in-filling of the O-2-A band. IEEE T. Geosci. Remote 2005, 43, 2860–2869. [Google Scholar]
- Thorhaug, A.; Richardson, A.D.; Berlyn, G.P. Spectral reflectance of Thalassia testudinum (Hydrocharitaceae) seagrass: low salinity effects. Am. J. Bot. 2006, 93, 110–117. [Google Scholar]
- Suárez, L.; Zarco-Tejada, P.J.; Sepulcre-Cantó, G.; Pérez-Priego, O.; Miller, J.R.; Jiménez-Muñoz, J.C.; Sobrino, J. Assessing canopy PRI for water stress detection with diurnal airborne imagery. Remote Sens. Environ. 2008, 112, 560–575. [Google Scholar]
- Stoll, M.-P.; Buschmann, C.; Court, A.; Laurila, T.; Moreno, J.; Moya, I. The FLEX-Fluorescence Explorer mission project: motivations and present status of preparatory activities. IGARSS03 2003, 1, 585–587. [Google Scholar]
- Guanter, L.; Alonso, L.; Gómez-Chova, L.; Amorós, J.; Vila, J.; Moreno, J. Estimation of solar-induced vegetation fluorescence from space measurements. Geophys. Res. Lett. 2007, 34, L08401. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Miller, J.R.; Pedrós, R.; Verhoef, W.; Berger, M. FluorMODgui V3.0 - A Graphic User Interface for the Leaf and Canopy Simulation of Chlorophyll Fluorescence. Comput. Geosci.-UK. 2006, 32(5), 577–591. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FWHM (nm) | Sampling interval (nm) | Spectral range (nm) | |
|---|---|---|---|
| Spectrometer 1 | 0.13 | 0.02 | 707-805 |
| Spectrometer 2 | 2.8 | 0.24 | 350-1050 |
© 2008 by MDPI Reproduction is permitted for noncommercial purposes.
Share and Cite
Meroni, M.; Rossini, M.; Picchi, V.; Panigada, C.; Cogliati, S.; Nali, C.; Colombo, R. Assessing Steady-state Fluorescence and PRI from Hyperspectral Proximal Sensing as Early Indicators of Plant Stress: The Case of Ozone Exposure. Sensors 2008, 8, 1740-1754. https://doi.org/10.3390/s8031740
Meroni M, Rossini M, Picchi V, Panigada C, Cogliati S, Nali C, Colombo R. Assessing Steady-state Fluorescence and PRI from Hyperspectral Proximal Sensing as Early Indicators of Plant Stress: The Case of Ozone Exposure. Sensors. 2008; 8(3):1740-1754. https://doi.org/10.3390/s8031740
Chicago/Turabian StyleMeroni, Michele, Micol Rossini, Valentina Picchi, Cinzia Panigada, Sergio Cogliati, Cristina Nali, and Roberto Colombo. 2008. "Assessing Steady-state Fluorescence and PRI from Hyperspectral Proximal Sensing as Early Indicators of Plant Stress: The Case of Ozone Exposure" Sensors 8, no. 3: 1740-1754. https://doi.org/10.3390/s8031740
APA StyleMeroni, M., Rossini, M., Picchi, V., Panigada, C., Cogliati, S., Nali, C., & Colombo, R. (2008). Assessing Steady-state Fluorescence and PRI from Hyperspectral Proximal Sensing as Early Indicators of Plant Stress: The Case of Ozone Exposure. Sensors, 8(3), 1740-1754. https://doi.org/10.3390/s8031740