
PKC in Regenerative Therapy: New Insights for Old Targets




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
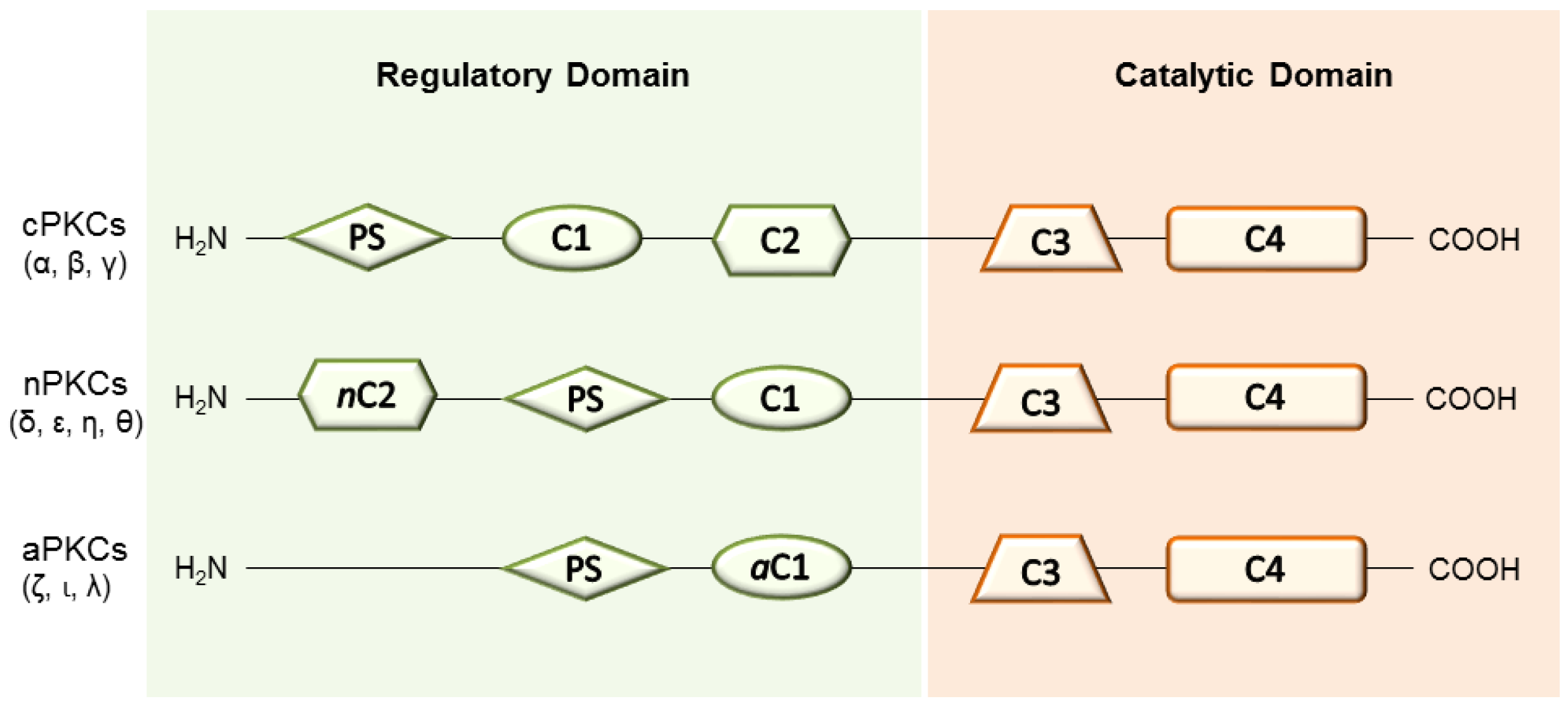
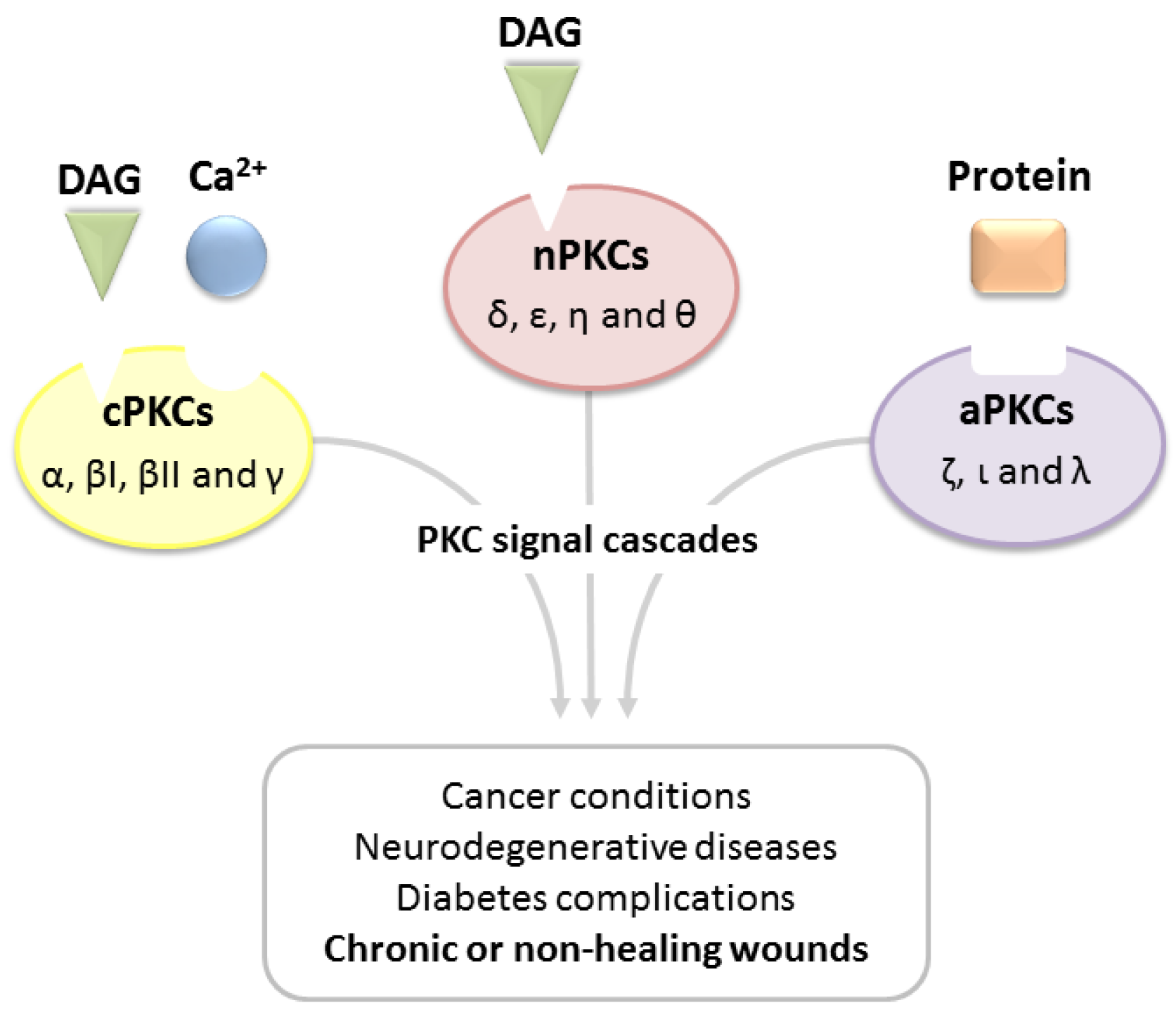
2. PKC Isoenzymes and Their Role in Tissue Regeneration
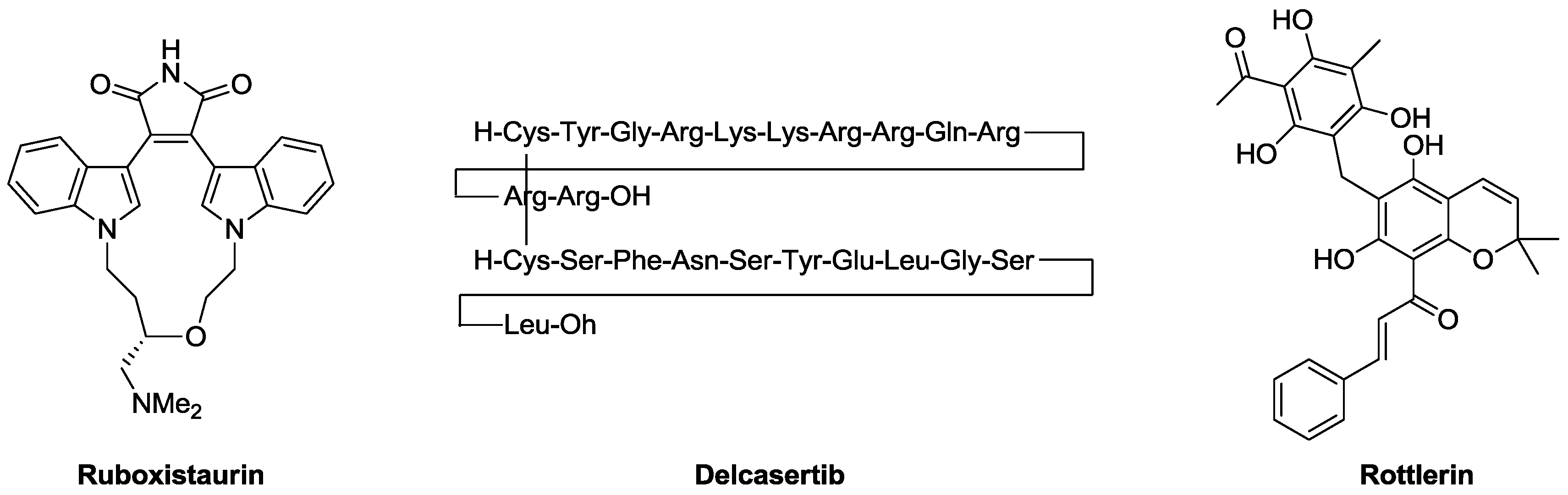
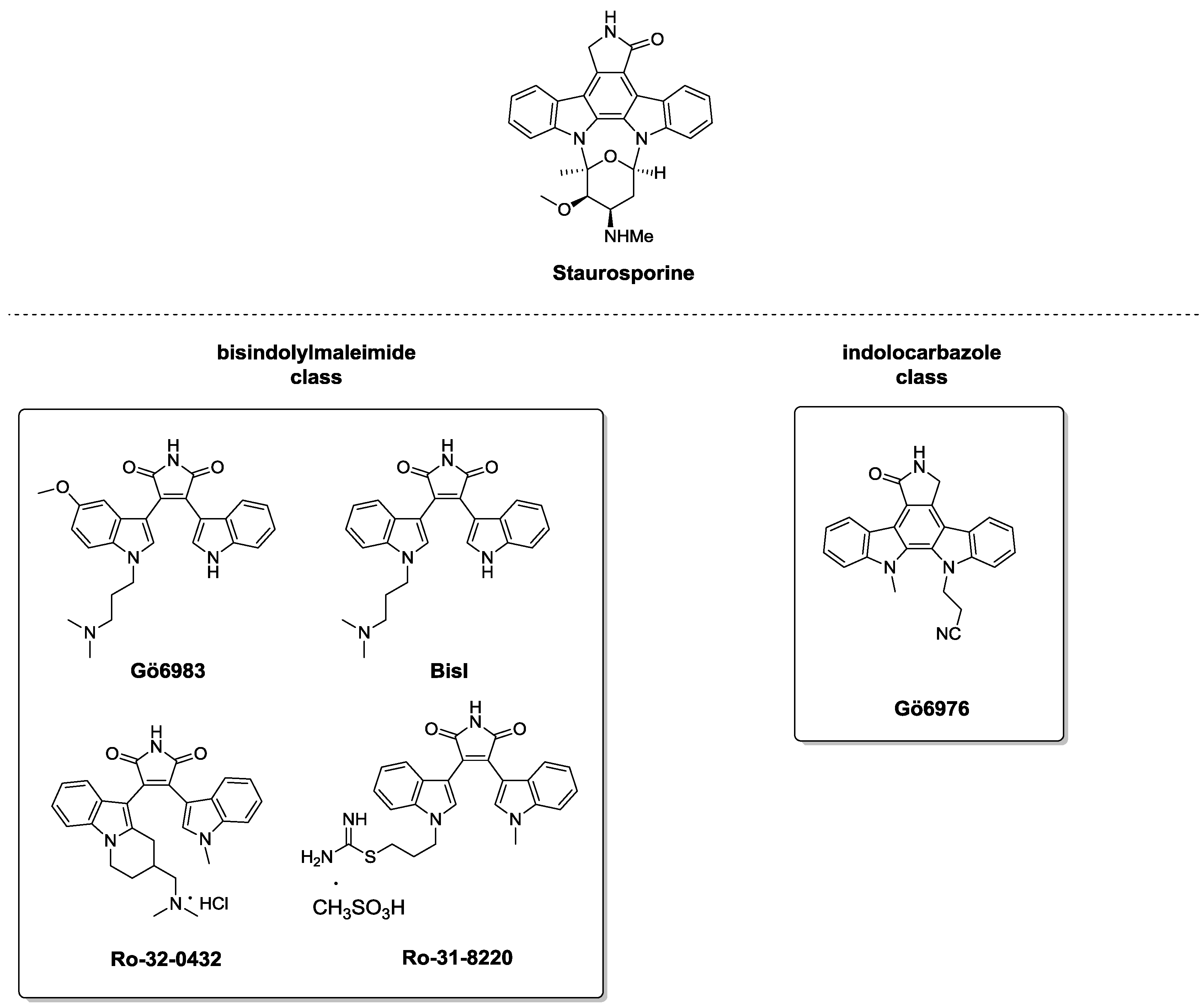
3. PKC Ligands
3.1. C3 Domain Ligands
3.2. C1 Domain Ligands
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Mason, C.; Dunnill, P. A brief definition of regenerative medicine. Regen. Med. 2008, 3, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Frykberg, R.G.; Banks, J. Challenges in the Treatment of Chronic Wounds. Adv. Wound Care 2015, 4, 560–582. [Google Scholar] [CrossRef] [PubMed]
- Pierce, G.F. Inflammation in nonhealing diabetic wounds: The space-time continuum does matter. Am. J. Pathol. 2001, 159, 399–403. [Google Scholar] [CrossRef]
- Edwards, J.; Howley, P.; Cohen, I.K. In vitro inhibition of human neutrophil elastase by oleic acid albumin formulations from derivatized cotton wound dressings. Int. J. Pharm. 2004, 284, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yannas, I.V.; Lee, E.; Orgill, D.P.; Skrabut, E.M.; Murphy, G.F. Synthesis and characterization of a model extracellular matrix that induces partial regeneration of adult mammalian skin. Proc. Natl. Acad. Sci. USA 1989, 86, 933–937. [Google Scholar] [CrossRef] [PubMed]
- Richmond, N.A.; Maderal, A.D.; Vivas, A.C. Evidence-based management of common chronic lower extremity ulcers. Dermatol. Ther. 2013, 26, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Geoffrey, C.; Gurtner, G.C.; Chapman, M.A. Regenerative Medicine: Charting a New Course in Wound Healing. Adv. Wound Care 2016, 5, 314–328. [Google Scholar]
- Kemp, P. History of regenerative medicine: Looking backwards to move forwards. Regen. Med. 2006, 1, 653–669. [Google Scholar] [CrossRef] [PubMed]
- Nunan, R.; Harding, K.G.; Martin, P. Clinical challenges of chronic wounds: Searching for an optimal animal model to recapitulate their complexity. Dis. Model Mech. 2014, 7, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Geraldes, P.; King, G.L. Activation of protein kinase C isoforms and its impact on diabetic complications. Circ. Res. 2010, 106, 1319–1331. [Google Scholar] [CrossRef] [PubMed]
- Takai, Y.; Kishimoto, A.; Inoue, M.; Nishizuka, Y. Studies on a cyclic nucleotide-independent protein kinase and its proenzyme in mammalian tissues. I. Purification and characterization of an active enzyme from bovine cerebellum. J. Biol. Chem. 1977, 252, 7603–7609. [Google Scholar] [PubMed]
- Shun-ichi, N.; Hirokei, Y.; Nishizuka, Y. Father of protein kinase C. J. Biochem. 2010, 148, 125–130. [Google Scholar]
- Van der Zee, E.A.; Luiten, P.G.; Disterhoft, J.F. Learning-Induced Alterations in Hippocampal PKC-immunoreactivity: A Review and Hypothesis of Its Functional Significance. Prog. Neuropsychopharmacol. Biol. Psychiatry 1997, 21, 531–572. [Google Scholar] [CrossRef]
- Pascale, A.; Noguès, X.; Marighetto, A.; Micheau, J.; Battaini, F.; Govoni, S.; Jaffard, R. Cytosolic hippocampal PKC and aging: Correlation with discrimination performance. NeuroReport 1998, 9, 725–729. [Google Scholar] [CrossRef]
- Newton, A.C. Regulation of the ABC kinases by phosphorylation: Protein kinase C as a paradigm. Chem. Rev. 2001, 101, 2353–2364. [Google Scholar] [CrossRef] [PubMed]
- Battaini, F.; Mochly-Rosen, D. Happy birthday protein kinase C: Past, present and future of a superfamily. Pharmacol. Res. 2007, 55, 461–466. [Google Scholar] [CrossRef] [PubMed]
- House, C.; Kemp, B.E. Protein kinase C contains a pseudosubstrate prototope in its regulatory domain. Science 1987, 238, 1726–1728. [Google Scholar] [CrossRef] [PubMed]
- Newton, A.C. Protein Kinase C: Structure, Function, and Regulation. J. Biol. Chem. 1995, 270, 28495–28498. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, S.F. Structural basis of protein kinase C isoform function. Physiol. Rev. 2008, 88, 1341–1378. [Google Scholar] [CrossRef] [PubMed]
- Newton, A.C. Protein kinase C: Poised to signal. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E395–E402. [Google Scholar] [CrossRef] [PubMed]
- Nishizuka, Y. The molecular heterogeneity of protein kinase C and its implications for cellular regulation. Nature 1988, 334, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Nishizuka, Y. Intracellular signaling by hydrolysis of phospholipids and activation of protein kinase C. Science 1992, 258, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Govoni, S.; Amadio, M.; Battaini, F.; Pascale, A. Senescence of the brain: Focus on cognitive kinases. Curr. Pharm. Des. 2010, 16, 660–671. [Google Scholar] [CrossRef] [PubMed]
- Battaini, F. Protein kinase C isoforms as therapeutic targets in nervous system disease states. Pharmacol. Res. 2001, 44, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, J. Protein kinase C isozymes as potential targets for anticancer therapy. Curr. Cancer Drug Targets 2004, 4, 125–146. [Google Scholar] [CrossRef] [PubMed]
- Blumberg, P.M.; Kedei, N.; Lewin, N.E.; Yang, D.; Czifra, G.; Pu, Y.; Peach, M.L.; Marquez, V.E. Wealth of opportunity—the C1 domain as a target for drug development. Curr. Drug Targets 2008, 9, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Bynagari-Settipalli, Y.S.; Chari, R.; Kilpatrick, L.; Kunapuli, S.P. Protein kinase C—possible therapeutic target to treat cardiovascular diseases. Cardiovasc. Hematol. Disord. Drug Targets 2010, 10, 292–308. [Google Scholar] [CrossRef] [PubMed]
- Mochly-Rosen, D.; Das, K.; Grimes, K.V. Protein kinase C, an elusive therapeutic target? Nat. Rev. Drug Discov. 2012, 11, 937–957. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.H. ABC transporters as multidrug resistance mechanisms and the development of chemosensitizers for their reversal. Cancer Cell Int. 2005, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gottesman, M.M.; Fojo, T.; Bates, S.E. Multidrug resistance in cancer: Role of ATP-dependent transporters. Nat. Rev. Cancer 2002, 2, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Martiny-Baron, G.; Fabbro, D. Classical PKC isoforms in cancer. Pharmacol. Res. 2007, 55, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Lamark, T.; Perander, M.; Outzen, H.; Kristiansen, K.; Øvertan, A.; Michaelsen, M.; Bjørkøy, G.; Johansen, T. Interaction codes within the family of mammalian Phox and Bem1p domain containing proteins. J. Biol. Chem. 2003, 278, 34568–34581. [Google Scholar] [CrossRef] [PubMed]
- Murray, N.R.; Kalari, K.R.; Fields, A.P. Protein kinase C expression and oncogenic signaling mechanisms in cancer. J. Cell Physiol. 2011, 226, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Antal, C.E.; Hudson, A.M.; Kang, E.; Zanca, C.; Wirth, C.; Stephenson, N.L.; Trotter, E.W.; Gallegos, L.L.; Miller, C.J.; Furnari, F.B.; et al. Cancer-Associated Protein Kinase C Mutations Reveal Kinase’s Role as Tumor Suppressor. Cell 2015, 160, 489–502. [Google Scholar] [CrossRef] [PubMed]
- Newton, A.C.; Brognard, J. Reversing the Paradigm: Protein Kinase C as a Tumor Suppressor. Trends Pharmacol. Sci. 2017, 38, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Kun-Sun, M.; Alkon, D.L. Pharmacology of protein kinase C activators: Cognition-enhancing and antidementic therapeutics. Pharmacol. Ther. 2010, 127, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Pascale, A.; Amadio, M.; Govoni, S.; Battaini, F. The aging brain, a key target for the future: The protein kinase C involvement. Pharmacol. Res. 2007, 55, 560–569. [Google Scholar] [CrossRef] [PubMed]
- Pascale, A.; Amadio, M.; Scampagnini, G.; Lanni, C.; Racchi, M.; Provenzani, A.; Govoni, S.; Alkon, D.L.; Quattrone, A. Neuronal ELAV proteins enhance mRNA stability by a PKCalpha-dependent pathway. PNAS 2005, 102, 12065–12070. [Google Scholar] [CrossRef] [PubMed]
- Alfonso, S.I.; Callender, J.A.; Hooli, B.; Antal, C.E.; Mullin, K.; Sherman, M.A.; Lesné, S.E.; Leitges, M.; Newton, A.C.; Tanzi, R.E.; et al. Gain-of-function mutations in protein kinase Cα (PKCα) may promote synaptic defects in Alzheimer’s disease. Sci. Signal 2016, 9, ra47. [Google Scholar] [CrossRef] [PubMed]
- Amadio, M.; Bucolo, C.; Leggio, G.M.; Drago, F.; Govoni, S.; Pascale, A. The PKCbeta/HuR/VEGF pathway in diabetic retinopathy. Biochem. Pharm. 2010, 80, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Casellini, C.M.; Barlow, P.M.; Rice, A.L.; Casey, M.; Simmons, K.; Pittenger, G.; Bastyr, E.J., 3rd; Wolka, A.M.; Vinik, A.I. A 6-month, randomized, double-masked, placebo-controlled study evaluating the effects of the protein kinase C-beta inhibitor ruboxistaurin on skin microvascular blood flow and other measures of diabetic peripheral neuropathy. Diabetes Care 2007, 30, 896–902. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.D.; Sheetz, M.J.; Aiello, L.P.; Milton, R.C.; Danis, R.P.; Zhi, X.; Girach, A.; Jimenez, M.C.; Vignati, L. Effect of ruboxistaurin on the visual acuity decline associated with long-standing diabetic macular edema. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Lincoff, A.M.; Roe, M.; Aylward, P.; Galla, J.; Rynkiewicz, A.; Guetta, V.; Zelizko, M.; Kleiman, N.; White, H.; McErlean, E.; et al. Inhibition of delta-protein kinase C by delcasertib as an adjunct to primary percutaneous coronary intervention for acute anterior ST-segment elevation myocardial infarction: Results of the PROTECTION AMI Randomized Controlled Trial. Eur. Heart J. 2014, 35, 2516–2523. [Google Scholar] [CrossRef] [PubMed]
- Joyce, N.C.; Meklir, B. Protein kinase C activation during corneal endothelial wound repair. Investig. Ophthalmol. Vis. Sci. 1992, 33, 1958–1973. [Google Scholar] [PubMed]
- Thomason, H.A.; Cooper, N.H.; Ansell, D.M.; Chiu, M.; Merrit, A.J.; Hardman, M.J.; Garrod, D.R. Direct evidence that PKCα positively regulates wound re-epithelialization: Correlation with changes in desmosomal adhesiveness. J. Pathol. 2012, 227, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Khamaisi, M.; Katagiri, S.; Keenan, H.; Park, K.; Maeda, Y.; Li, Q.; Qi, W.; Thomou, T.; Eschuk, D.; Tellechea, A.; et al. PKCδ inhibition normalizes the wound-healing capacity of diabetic human fibroblasts. J. Clin. Investig. 2016, 126, 837–853. [Google Scholar] [CrossRef] [PubMed]
- Wu-Zhang, A.X.; Newton, A.C. Protein kinase C pharmacology: Refining the toolbox. Biochem. J. 2013, 452, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Darby, I.A.; Laverdet, B.; Bonté, F.; Desmoulière, A. Fibroblasts and myofibroblasts in wound healing. Clin. Cosmet. Investig. Dermatol. 2014, 7, 301–311. [Google Scholar] [PubMed]
- Driskell, R.R.; Lichtenberger, B.M.; Hoste, E.; Kretzschmar, K.; Simons, B.D.; Charalambous, M.; Ferron, S.R.; Herault, Y.; Pavlovic, G.; Ferguson-Smith, A.C.; et al. Distinct fibroblast lineages determine dermal architecture in skin development and repair. Nature 2013, 504, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Boije, A.G.G.; Talman, V.; Yli-Kauhaluoma, J.; Tuominen, R.K.; Ekokoski, E. Current Status and Future Prospects of C1 Domain Ligands as Drug Candidates. Curr. Top. Med. Chem. 2011, 11, 1370–1392. [Google Scholar] [CrossRef]
- Davis, M.I.; Hunt, J.P.; Herrgard, S.; Ciceri, P.; Wodicka, L.M.; Pallares, G.; Hocker, M.; Treiber, D.K.; Zarrinkar, P.P. Comprehensive analysis of kinase inhibitor selectivity. Nat. Biotechnol. 2011, 29, 1046–1051. [Google Scholar] [CrossRef] [PubMed]
- Anastassiadis, T.; Deacon, S.W.; Devarajan, K.; Ma, H.; Peterson, J.R. Comprehensive assay of kinase catalytic activity reveals features of kinase inhibitor selectivity. Nat. Biotechnol. 2011, 29, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, S.E.; Parker, P.J.; Nixon, J.S. Isoenzyme specificity of bisindolylmaleimides, selective inhibitors of protein kinase C. Biochem. J. 1993, 294, 335–337. [Google Scholar] [CrossRef] [PubMed]
- Omura, S.; Iwai, Y.; Hirano, A.; Nakagawa, A.; Awaya, J.; Tsuchya, H.; Takahashi, Y.; Masuma, R. A new alkaloid AM-2282 of streptomyces origin. Taxonomy, fermentation, isolation and preliminary characterization. J. Antibiot. 1977, 30, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Martiny-Baron, G.; Kazanietz, M.G.; Mischak, H.; Blumberg, P.M.; Kochs, G.; Hug, H.; Marme, D.; Schachtele, C. Selective inhibition of protein kinase C isozymes by the indolocarbazole Gö6976. J. Biol. Chem. 1993, 268, 9194–9197. [Google Scholar] [PubMed]
- Gschwendt, M.; Dieterich, S.; Rennecke, J.; Kittstein, W.; Mueller, H.J.; Johannes, F.J. Inhibition of protein kinase Cμ by various inhibitors. Differentiation from protein kinase C isoenzymes. FEBS Lett. 1996, 392, 77–80. [Google Scholar] [CrossRef]
- Alessi, D.R. The protein kinase C inhibitors Ro 318220 and GF 109203X are equally potent inhibitors of MAPKAP kinase-1β (Rsk-2) and p70 S6 kinase. FEBS Lett. 1997, 402, 121–123. [Google Scholar] [CrossRef]
- Davies, S.P.; Reddy, H.; Caivano, M.; Cohen, P. Specificity and mechanism of action of some commonly used protein kinase inhibitors. Biochem. J. 2000, 351, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Bain, J.; Plater, L.; Elliott, M.; Shpiro, N.; Hastie, C.J.; McLauchlan, H.; Klevernic, I.; Arthur, J.S.; Alessi, D.R.; Cohen, P. The selectivity of protein kinase inhibitors: A further update. Biochem. J. 2007, 408, 297–315. [Google Scholar] [CrossRef] [PubMed]
- Takagi, T.; Naito, Y.; Uchiyama, K.; Mizuhima, K.; Suzuki, T.; Horie, R.; Hirata, I.; Tsuboi, H.; Yoshikawa, T. Carbon monoxide promotes gastric wound healing in mice via the protein kinase C pathway. Free Radic. Res. 2016, 50, 1098–1105. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.S.; Ryu, J.H.; Zuo, Z.; Yang, S.M.; Chang, H.W.; Do, S.H. Riluzole attenuates excitatory amino acid transporter type 3 activity in Xenopus oocytes via protein kinase C inhibition. Eur. J. Pharmacol. 2013, 713, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Lamanauskas, N.; Nistri, A. Riluzole blocks persistent Na+ and Ca2+ currents and modulates release of glutamate via presynaptic NMDA receptors on neonatal rat hypoglossal motoneurons in vitro. Eur. J. Neurosci. 2008, 27, 2501–2514. [Google Scholar] [CrossRef] [PubMed]
- Noh, K.M.; Hwang, J.Y.; Shin, H.C.; Koh, J.Y. A Novel Neuroprotective Mechanism of Riluzole: Direct Inhibition of Protein Kinase C. Neurobiol. Dis. 2000, 7, 375–383. [Google Scholar] [CrossRef] [PubMed]
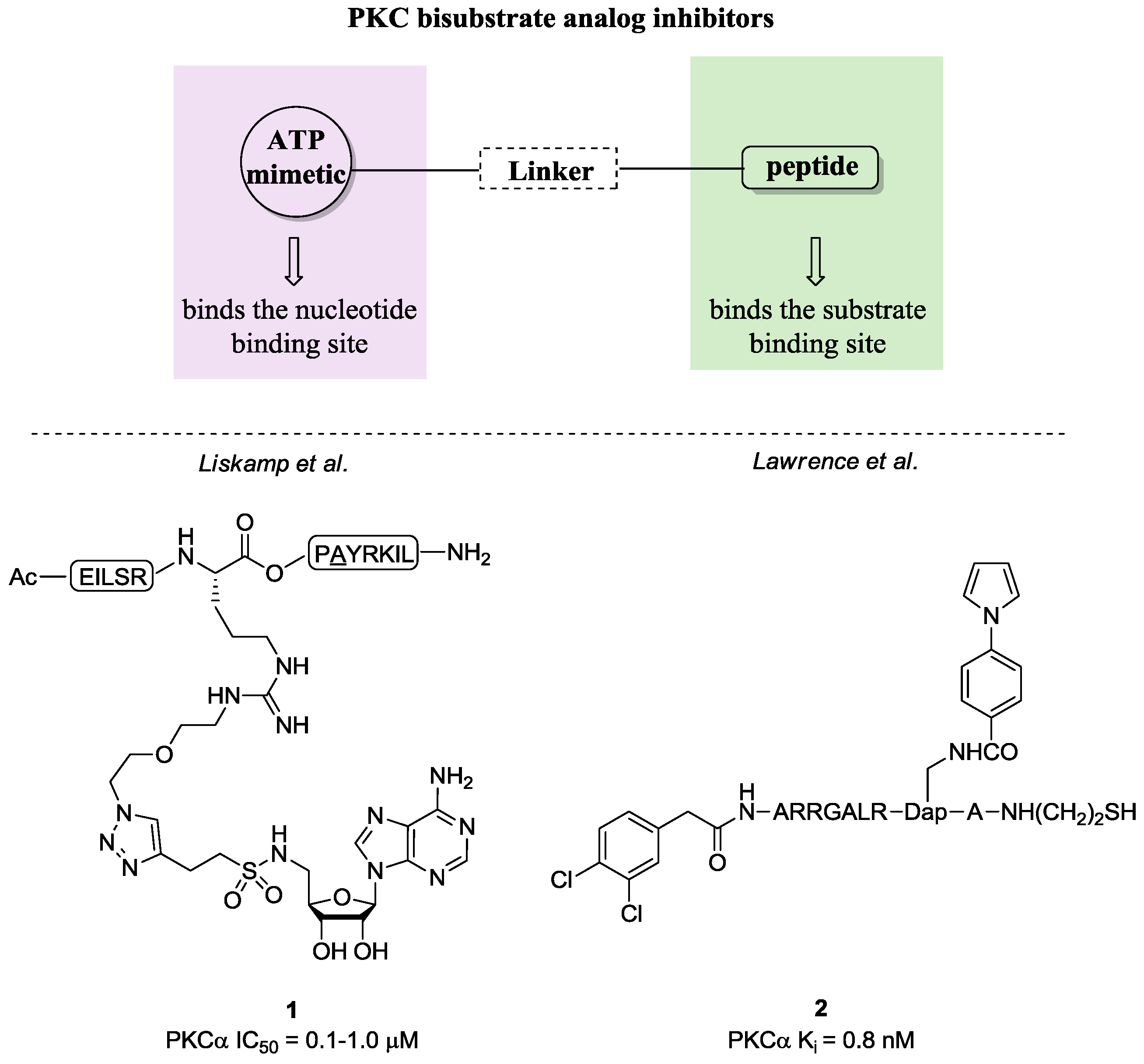
- Parang, K.; Cole, P.A. Designing bisubstrate analog inhibitors for protein kinases. Pharmacol. Ther. 2002, 93, 145–157. [Google Scholar] [CrossRef]
- Poot, A.J.; van Ameijde, J.; Slijper, M.; van den Berg, A.; Hilhorst, R.; Ruijtenbeek, R.; Rijkers, D.T.S; Liskamp, R.M.J. Development of selective bisubstrate-based inhibitors against protein kinase C (PKC) isozymes by using dynamic peptide microarrays. Chem. Biol. Chem. 2009, 10, 2042–2051. [Google Scholar] [CrossRef] [PubMed]
- Van Wandelen, L.T.M.; van Ameijde, J.; Mady, A.S.A.; Wammes, A.E.M.; Bode, A.; Poot, A.J.; Ruijtenbeek, R.; Liskamp, R.M.J. Directed modulation of protein kinase C isozyme selectivity with bisubstrate-based inhibitors. Chem. Med. Chem. 2012, 7, 2113–2121. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Nandy, S.K.; Lawrence, D.S. A highly potent and selective PKC alpha inhibitor generated via combinatorial modification of a peptide scaffold. J. Am. Chem. Soc. 2004, 126, 3394–3395. [Google Scholar] [CrossRef] [PubMed]
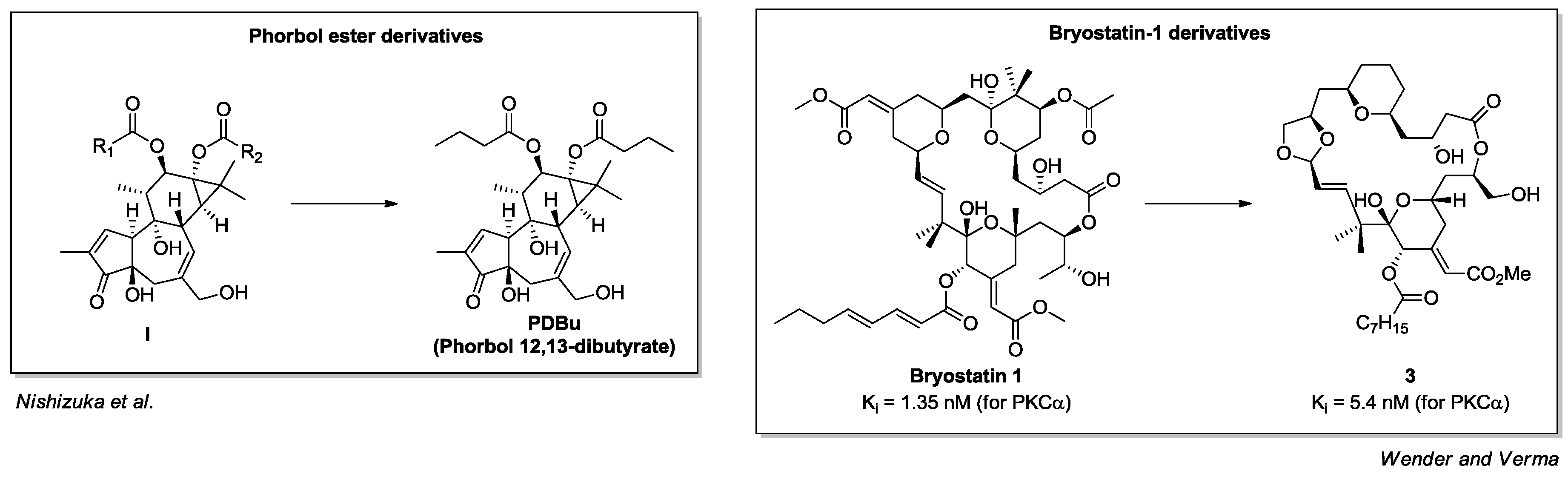
- Castagna, M.; Takai, Y.; Kaibuchi, K.; Sano, K.; Kikkawa, U.; Nishizuka, Y. Direct activation of calcium-activated, phospholipid-dependent protein kinase by tumor-promoting phorbol esters. J. Biol. Chem. 1982, 257, 7847–7851. [Google Scholar] [PubMed]
- Kortmansky, J.; Schwartz, G.K. Bryostatin-1: A novel PKC inhibitor in clinical development. Cancer Investig. 2003, 21, 924–936. [Google Scholar] [CrossRef] [PubMed]
- Driedger, P.E.; Blumberg, P.M. Specific binding of phorbol ester tumor promoters. Proc. Natl. Acad. Sci. USA 1980, 77, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Herald, C.L.; Doubek, D.L.; Herald, D.L.; Arnold, E.; Clardy, J. Isolation and structure of bryostatin 1. J. Am. Chem. Soc. 1982, 104, 6846–6848. [Google Scholar] [CrossRef]
- Wender, P.A.; Hinkle, K.W.; Koehler, M.F.; Lippa, B. The rational design of potential chemotherapeutic agents: Synthesis of bryostatin analogues. Med. Res. Rev. 1999, 19, 388–407. [Google Scholar] [CrossRef]
- Keck, G.E.; Kraft, M.B.; Truong, A.P.; Li, W.; Sanchez, C.C.; Kedei, N.; Lewin, N.E.; Blumberg, P.M. Convergent assembly of highly potent analogues of bryostatin 1 via pyran annulation: Bryostatin look-alikes that mimic phorbol ester function. J. Am. Chem. Soc. 2008, 130, 6660–6661. [Google Scholar] [CrossRef] [PubMed]
- Wender, P.A.; Verma, V.A. Design, synthesis, and biological evaluation of a potent, PKC selective, B-ring analog of bryostatin. Org. Lett. 2006, 8, 1893–1896. [Google Scholar] [CrossRef] [PubMed]
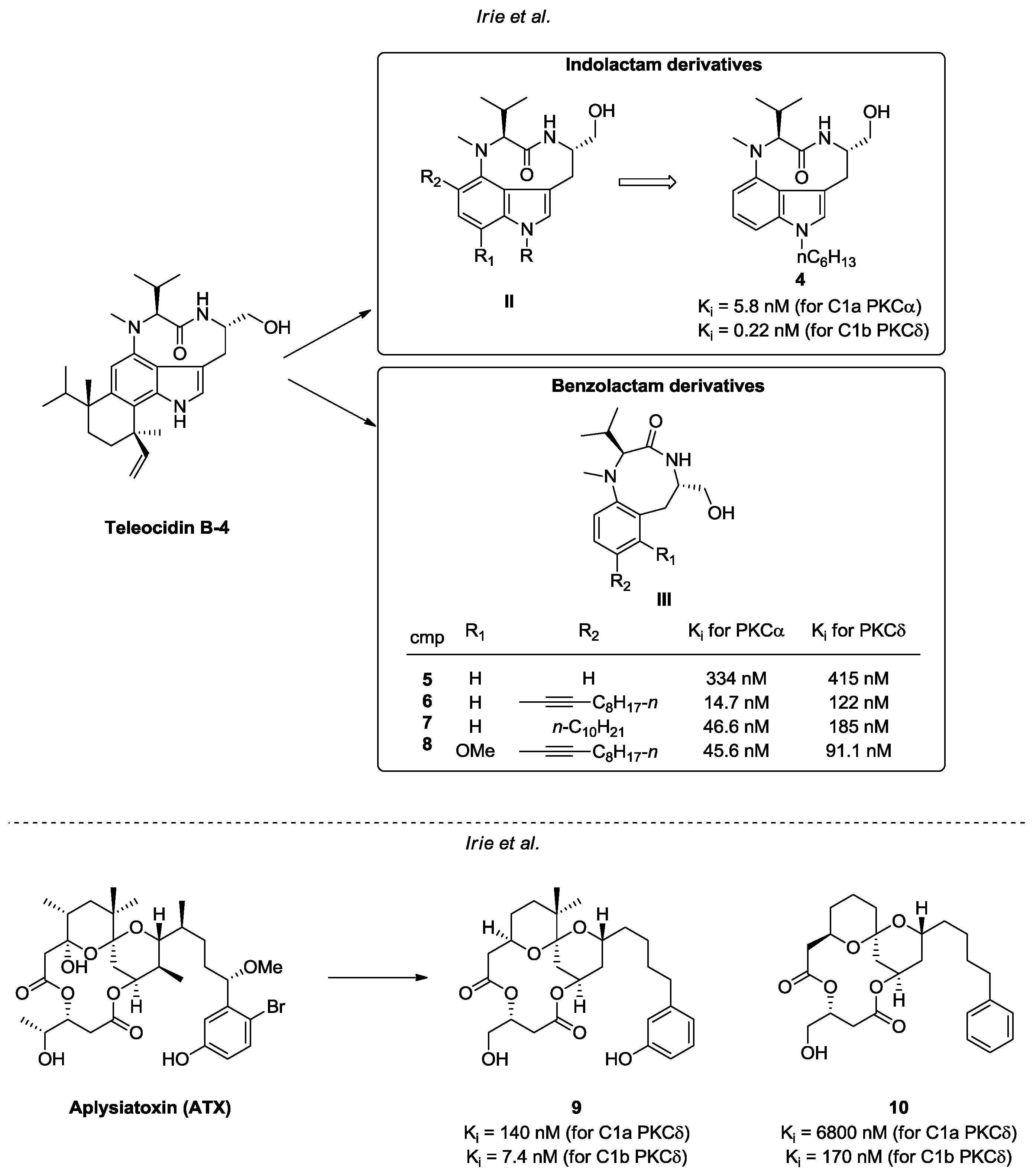
- Irie, K.; Yanagita, R.C. Synthesis and biological activities of simplified analogs of the natural PKC ligands, bryostatin-1 and aplysiatoxin. Chem. Rec. 2014, 14, 251–267. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, K.; Weinstein, I.B.; Horowitz, A.; Fujiki, H.; Matsushima, T.; Sugimura, T. Similarity of teleocidin B and phorbol ester tumour promoters in effects on membrane receptors. Nature 1981, 290, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, A.D.; Fujiki, H.; Weinstein, I.B. Comparative effects of aplysiatoxin, debromoaplysiatoxin, and teleocidin on receptor binding and phospholipid metabolism. Cancer Res. 1983, 43, 1529–1535. [Google Scholar] [PubMed]
- Fujiki, H.; Mori, M.; Nakayasu, M.; Terada, M.; Sugimura, T.; Moore, R.E. Indole alkaloids: Dihydroteleocidin B, teleocidin, and lyngbyatoxin A as members of a new class of tumor promoters. Proc. Natl. Acad. Sci. USA 1981, 78, 3872–3876. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, H.; Suganuma, M.; Nakayasu, M.; Hoshino, H.; Moore, R.E.; Sugimura, T. The third class of new tumor promoters, polyacetates (debromoaplysiatoxin and aplysiatoxin), can differentiate biological actions relevant to tumor promoters. Gann 1982, 73, 495–497. [Google Scholar] [PubMed]
- Irie, K.; Nakagawa, Y.; Ohigashi, H. Toward the Development of New Medicinal Leads with Selectivity for Protein Kinase C Isozymes. Chem. Rec. 2005, 5, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Yanagita, R.C.; Torii, K.; Nakagawa, Y.; Irie, K. Binding selectivity of 1- or 12- substituted indolactam derivatives for protein kinase C isozymes. Heterocycles 2007, 73, 289–302. [Google Scholar]
- Ma, D.; Tang, G.; Kozikowski, A.P. Synthesis of 7-Substituted Benzolactam-V8s and Their Selectivity for Protein Kinase C Isozymes. Org. Lett. 2002, 4, 2377–2380. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Zhang, T.; Wang, G.; Kozikowski, A.P.; Lewinc, N.E.; Blumbergc, P.M. Synthesis of 7,8-Disubstituted Benzolactam-V8 and Its Binding to Protein Kinase C. Bioorg. Med. Chem. Lett. 2001, 11, 99–101. [Google Scholar] [CrossRef]
- Irie, K.; Kikumori, M.; Kamachi, H.; Tanaka, K.; Murakami, A.; Yanagita, R.C.; Tokuda, H.; Suzuki, N.; Nagai, H.; Suenaga, K.; et al. Synthesis and structure–activity studies of simplified analogues of aplysiatoxin with antiproliferative activity like bryostatin-1. Pure Appl. Chem. 2012, 84, 1341–1351. [Google Scholar] [CrossRef]
- Marquez, V.E.; Blumberg, P.M. Synthetic diacylglycerols (DAG) and DAG-lactones as activators of protein kinase C (PK-C). Acc. Chem. Res. 2003, 36, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Siddiqui, M.A.; Sigano, D.M.; Krajewski, K.; Lewin, N.E.; Pu, Y.; Blumberg, P.M.; Lee, J.; Marquez, V.E. Conformationally constrained analogues of diacylglycerol. 24. Asymmetric synthesis of a chiral (R)-DAG-lactone template as a versatile precursor for highly functionalized DAG-lactones. Org. Lett. 2004, 6, 2413–2416. [Google Scholar] [CrossRef] [PubMed]
- El Kazzouli, S.; Lewin, N.E.; Blumberg, P.M.; Marquez, V.E. Conformationally constrained analogues of diacylglycerol. 30. An investigation of diacylglycerol-lactones contained heteroaryl groups reveals compounds with high selectivity for Ras guanyl nucleotide-releasing proteins. J. Med. Chem. 2008, 51, 5371–5386. [Google Scholar] [CrossRef] [PubMed]
- Duan, D.; Sigano, D.M.; Kelley, J.A.; Lai, C.C.; Lewin, N.E.; Kedei, N.; Peach, M.L.; Lee, J.; Abeyweera, T.P.; Rotenberg, S.A.; et al. Conformationally Constrained Analogues of Diacylglycerol. 29. Cells Sort Diacylglycerol-Lactone Chemical Zip Codes to Produce Diverse and Selective Biological Activities. J. Med. Chem. 2008, 51, 5198–5220. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, J.H.; Kim, S.Y.; Perry, N.A.; Lewin, N.E.; Ayres, J.A.; Blumberg, P.M. 2-Benzyl and 2-phenyl-3-hydroxypropyl pivalates as protein kinase C ligands. Bioorg. Med. Chem. 2006, 14, 2022–2031. [Google Scholar] [CrossRef] [PubMed]
- Rossi, D.; Talman, V.; Gennäs, G.B.A.; Marra, A.; Picconi, P.; Nasti, R.; Serra, M.; Ann, J.; Amadio, M.; Pascale, A.; et al. Beyond the affinity for protein kinase C: Exploring 2-phenyl-3-hydroxypropyl pivalate analogues as C1 domain-targeting ligands. Med. Chem. Commun. 2015, 6, 547–554. [Google Scholar] [CrossRef]
- Boije, A.G.G.; Talman, V.; Aitio, O.; Ekokoski, E.; Finel, M.; Tuominen, R.K.; Yli-Kauhaluoma, J. Design, Synthesis, and Biological Activity of Isophthalic Acid Derivatives Targeted to the C1 Domain of Protein Kinase C. J. Med. Chem. 2009, 52, 3969–3981. [Google Scholar]
- Talmana, V.; Amadio, M.; Osera, C.; Sorvari, S.; Gennäs, G.B.A.; Yli-Kauhaluoma, J.; Rossi, D.; Govoni, S.; Collina, S.; Ekokoski, E.; et al. The C1 domain-targeted isophthalate derivative HMI-1b11 promotes neurite outgrowth and GAP-43 expression through PKC activation in SH-SY5Y cells. Pharmacol. Res. 2013, 73, 44–54. [Google Scholar] [CrossRef] [PubMed]









© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rui, M.; Nasti, R.; Bignardi, E.; Della Volpe, S.; Rossino, G.; Rossi, D.; Collina, S. PKC in Regenerative Therapy: New Insights for Old Targets. Pharmaceuticals 2017, 10, 46. https://doi.org/10.3390/ph10020046
Rui M, Nasti R, Bignardi E, Della Volpe S, Rossino G, Rossi D, Collina S. PKC in Regenerative Therapy: New Insights for Old Targets. Pharmaceuticals. 2017; 10(2):46. https://doi.org/10.3390/ph10020046
Chicago/Turabian StyleRui, Marta, Rita Nasti, Emanuele Bignardi, Serena Della Volpe, Giacomo Rossino, Daniela Rossi, and Simona Collina. 2017. "PKC in Regenerative Therapy: New Insights for Old Targets" Pharmaceuticals 10, no. 2: 46. https://doi.org/10.3390/ph10020046
APA StyleRui, M., Nasti, R., Bignardi, E., Della Volpe, S., Rossino, G., Rossi, D., & Collina, S. (2017). PKC in Regenerative Therapy: New Insights for Old Targets. Pharmaceuticals, 10(2), 46. https://doi.org/10.3390/ph10020046