The Cannabinoid-Like Compound, VSN16R, Acts on Large Conductance, Ca2+-Activated K+ Channels to Modulate Hippocampal CA1 Pyramidal Neuron Firing

 ,
, 
Abstract
:1. Introduction
2. Methods
2.1. Ethical Approval
2.2. Preparation of Acute Hippocampus Slices
2.3. Electrophysiology
2.4. Pharmacology
2.5. Data Analysis
3. Results
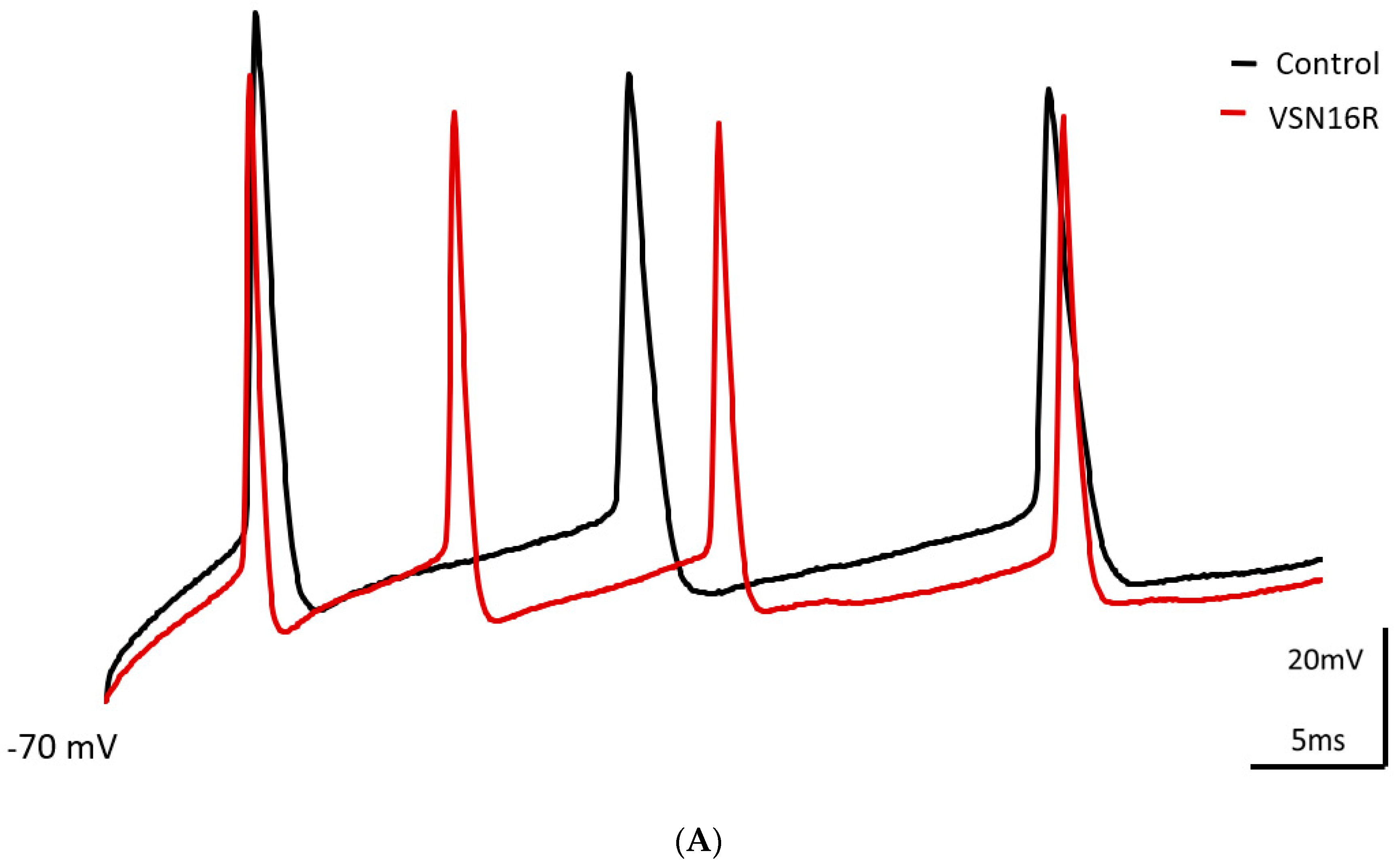
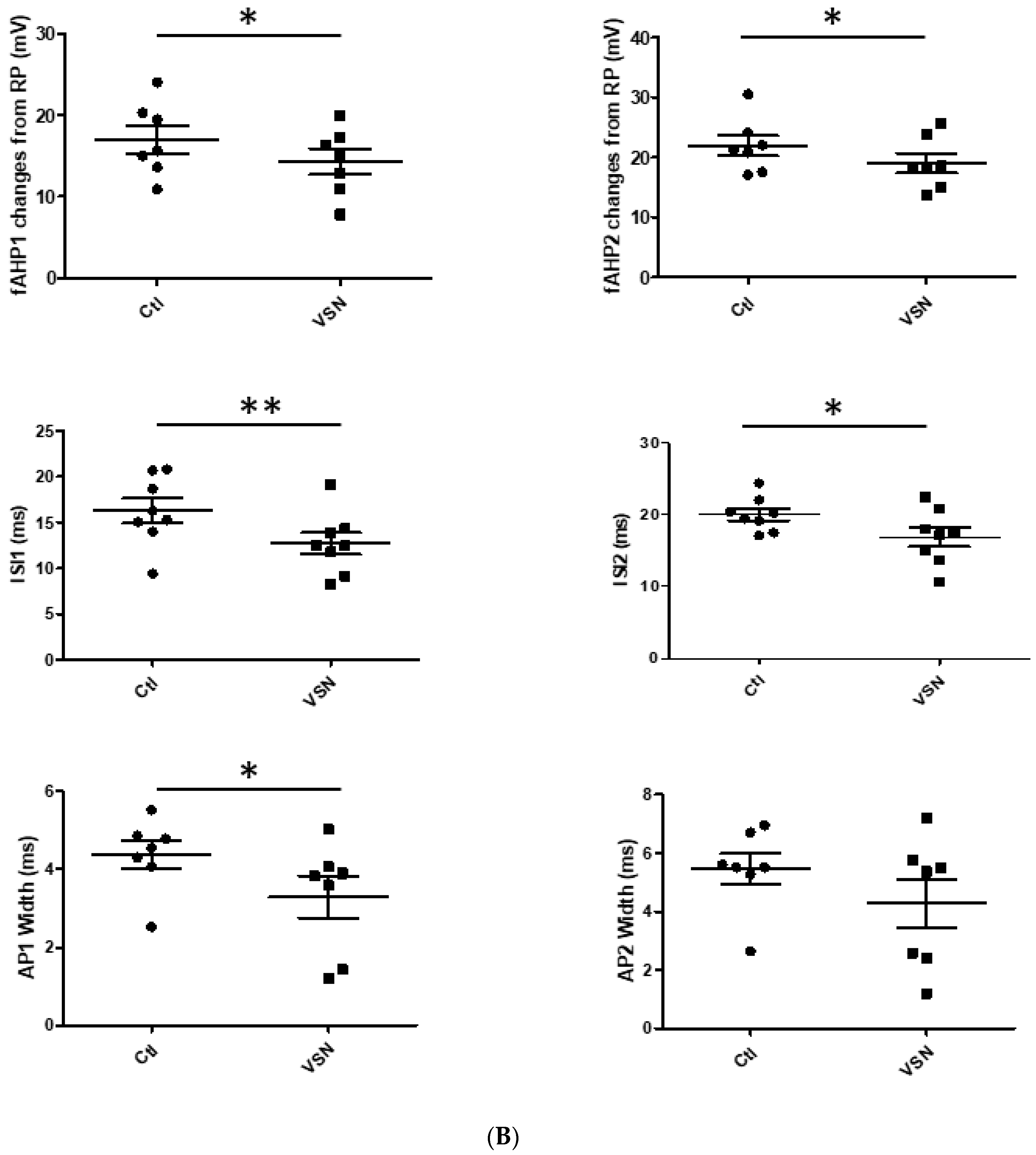
3.1. VSN16R Affects AP Waveform during High Frequency Stimulation in Hippocampal CA1 Pyramidal Neurons
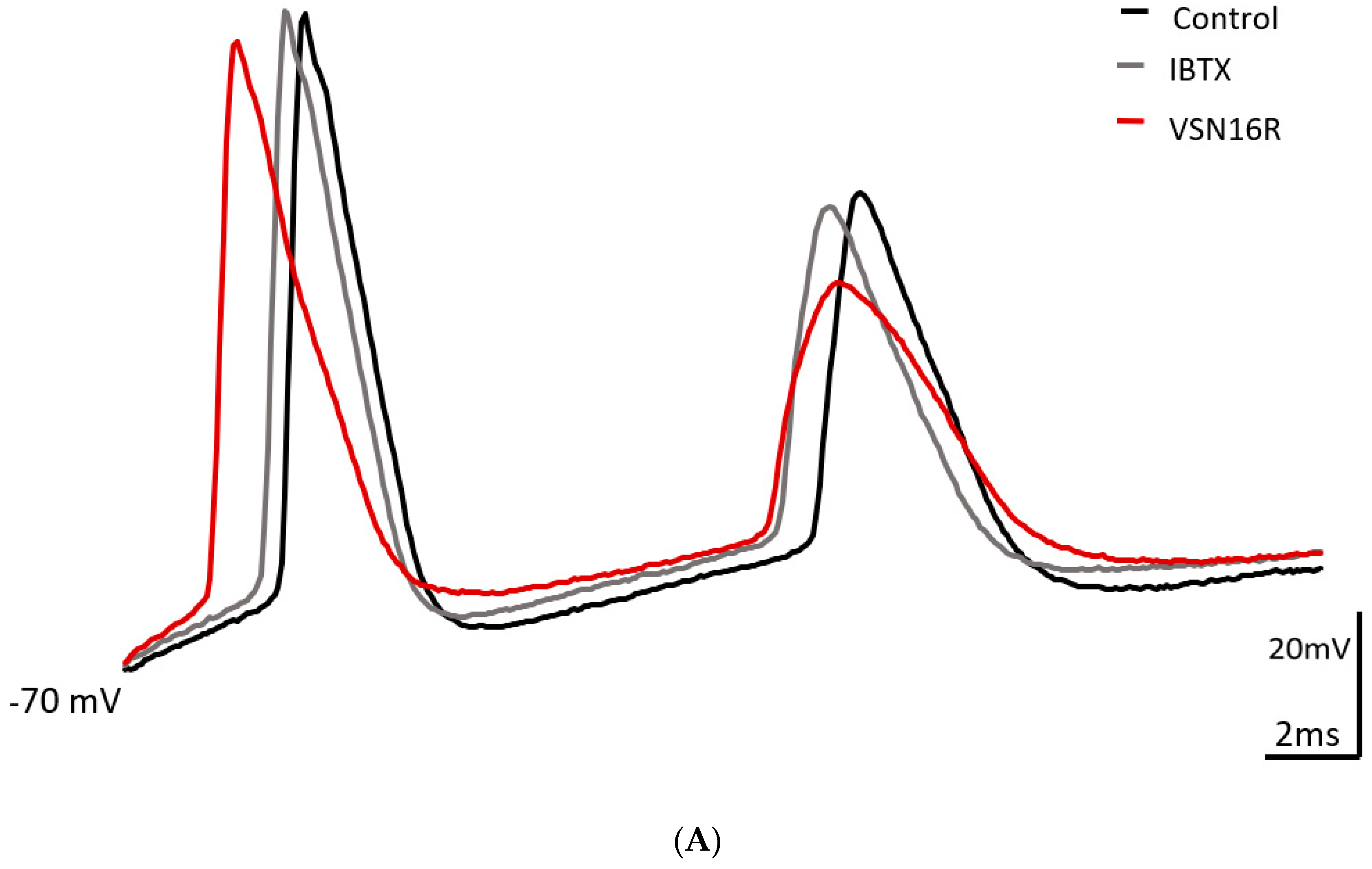
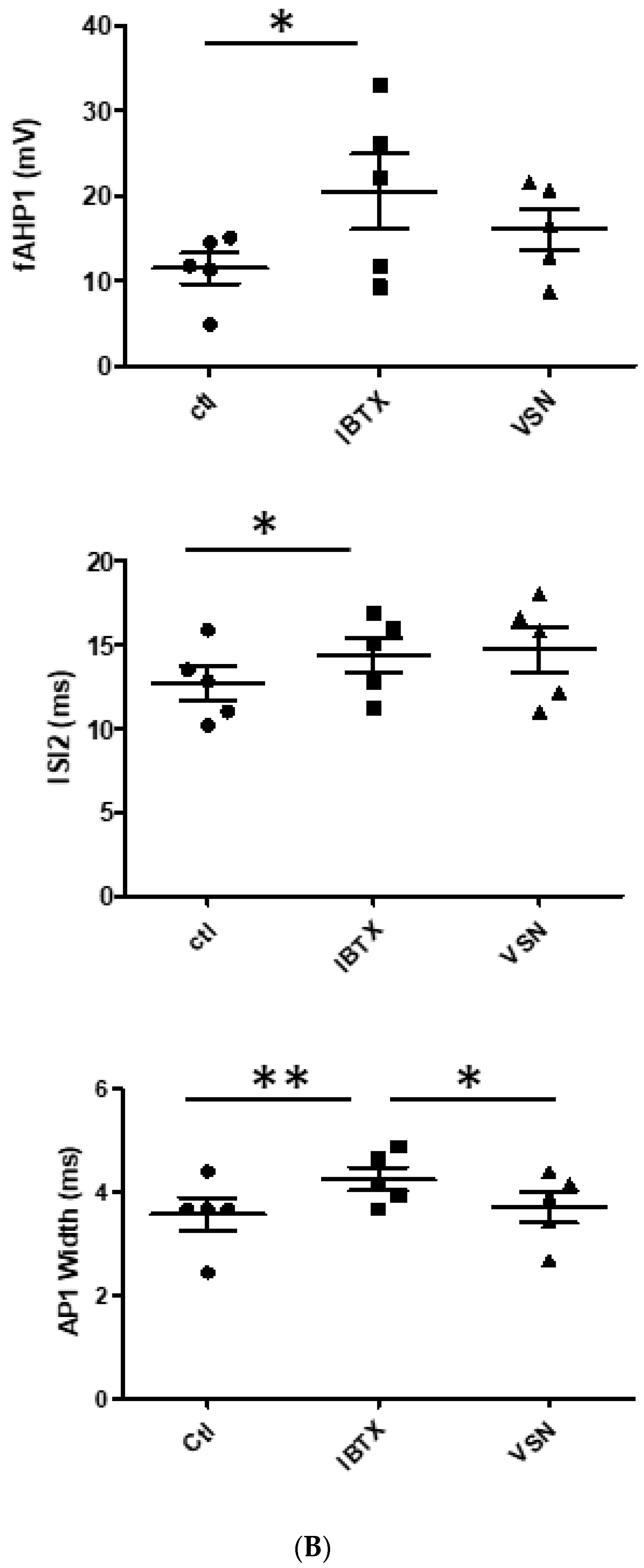
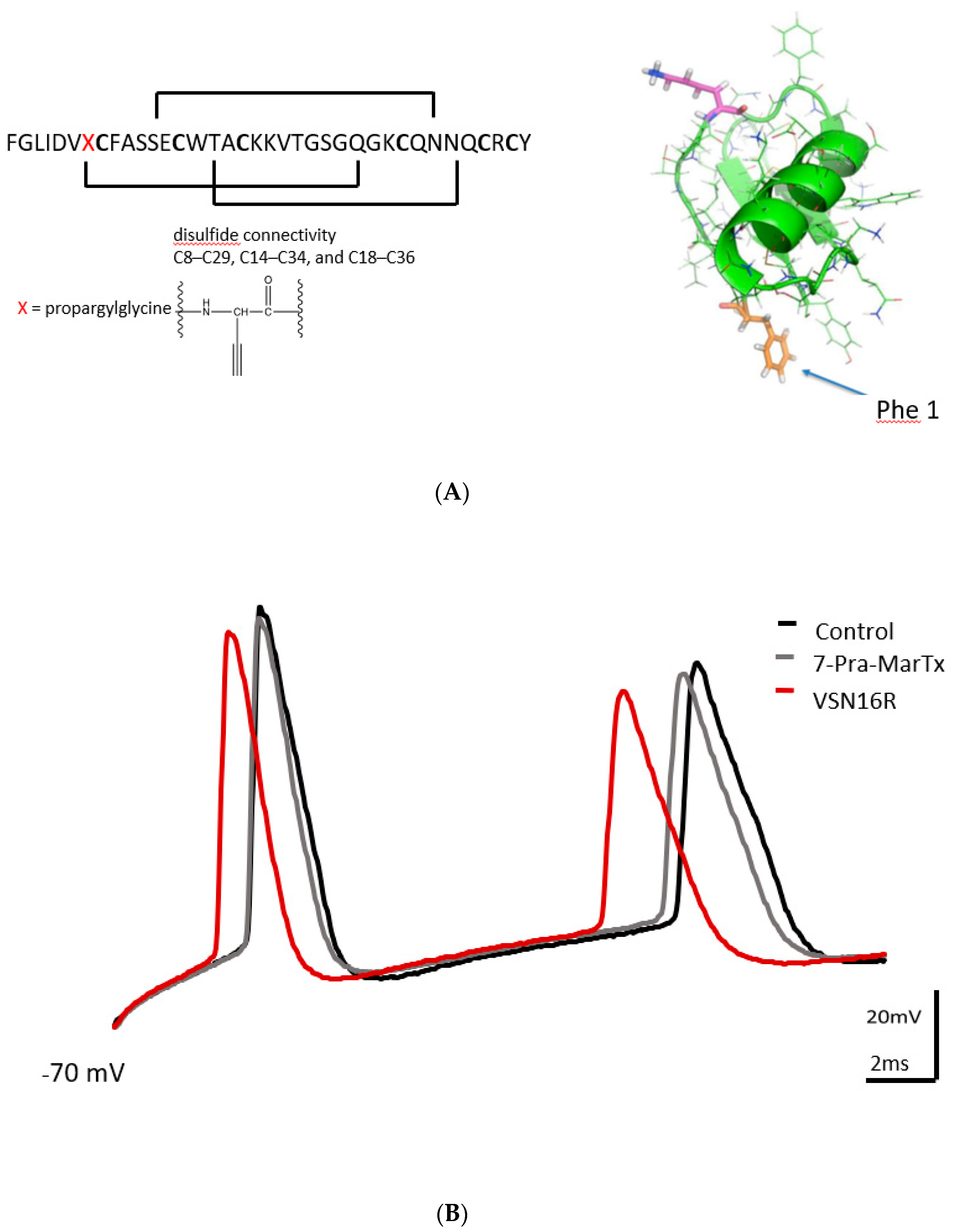
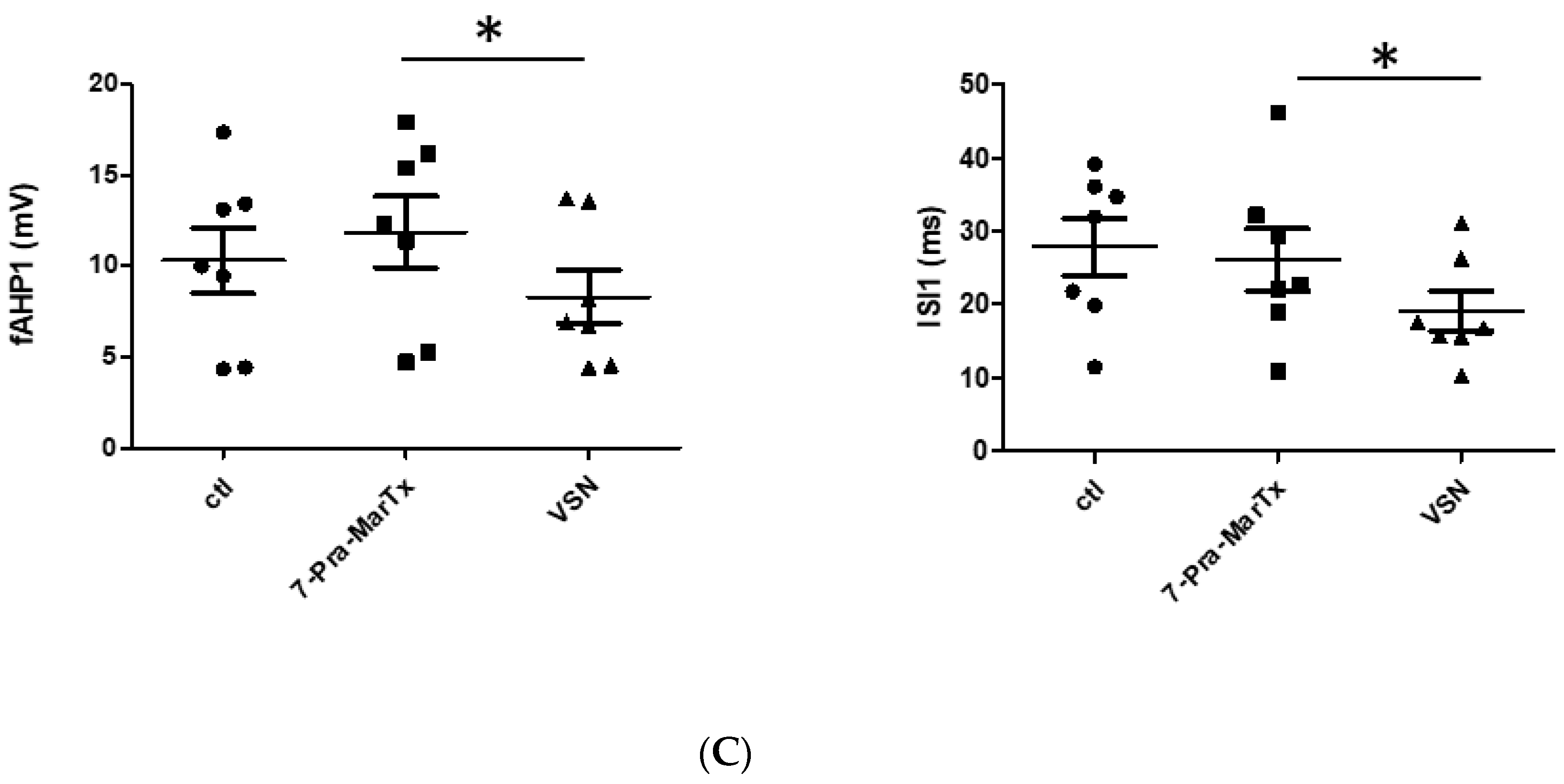
3.2. VSN16R Effects Are Blocked by IBTX, but Effects Persist, or Are Uncovered, in the Presence of 7-Pra-MarTx
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hoi, P.M.; Visintin, C.; Okuyama, M.; Gardiner, S.M.; Kaup, S.S.; Bennett, T.; Baker, D.; Selwood, D.L.; Hiley, C.R. Vascular pharmacology of a novel cannabinoid-like compound, 3-(5-dimethylcarbamoyl-pent-1-enyl)-N-(2-hydroxy-1-methyl-ethyl) benzamide (VSN16) in the rat. Br. J. Pharmacol. 2007, 152, 751–764. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.; Pryce, G.; Visintin, C.; Sisay, S.; Bondarenko, A.I.; Ho, W.S.V.; Jackson, S.J.; Williams, T.E.; Al-Izki, S.; Sevastou, I.; et al. Big conductance calcium-activated potassium channel openers control spasticity without sedation. Br. J. Pharmacol. 2017, 174, 2662–2681. [Google Scholar] [CrossRef] [Green Version]
- Latorre, R.; Castillo, K.; Carrasquel-Ursulaez, W.; Sepulveda, R.V.; Gonzalez-Nilo, F.; Gonzalez, C.; Alvarez, O. Molecular determinants of BK channel functional diversity and functioning. Physiol. Rev. 2017, 97, 39–87. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Perez, V.; Lingle, C.J. Regulation of BK channels by beta and gamma subunits. Annu Rev. Physiol. 2019, 81, 113–137. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V.; De Petrocellis, L. Why do cannabinoid receptors have more than one endogenous ligand? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 3216–3228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondarenko, A.I. Endothelial atypical cannabinoid receptor: Do we have enough evidence? Br. J. Pharmacol. 2014, 171, 5573–5588. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.P.; Safo, P.K.; Regehr, W.G. Endocannabinoids inhibit transmission at granule cell to Purkinje cell synapses by modulating three types of presynaptic calcium channels. J. Neurosci. 2004, 24, 5623–5631. [Google Scholar] [CrossRef]
- Gu, N.; Vervaeke, K.; Storm, J.F. BK potassium channels facilitate high-frequency firing and cause early spike frequency adaptation in rat CA1 hippocampal pyramidal cells. J. Physiol. 2007, 580, 859–882. [Google Scholar] [CrossRef]
- Ji, Y.H.; Wang, W.X.; Ye, J.G.; He, L.L.; Li, Y.J.; Yan, Y.P.; Zhou, Z. Martentoxin, a novel K+-channel-blocking peptide: Purification, cDNA and genomic cloning, and electrophysiological and pharmacological characterization. J. Neurochem. 2003, 84, 325–335. [Google Scholar] [CrossRef]
- Shi, J.; He, H.Q.; Zhao, R.; Duan, Y.H.; Chen, J.; Chen, Y.; Yang, J.; Zhang, J.W.; Shu, X.Q.; Zheng, P.; et al. Inhibition of martentoxin on neuronal BK channel subtype (alpha+beta4): implications for a novel interaction model. Biophys. J. 2008, 94, 3706–3713. [Google Scholar] [CrossRef]
- Golding, N.L.; Jung, H.Y.; Mickus, T.; Spruston, N. Dendritic calcium spike initiation and repolarization are controlled by distinct potassium channel subtypes in CA1 pyramidal neurons. J. Neurosci. 1999, 19, 8789–8798. [Google Scholar] [CrossRef]
- Wang, B.; Jaffe, D.B.; Brenner, R. Current understanding of iberiotoxin-resistant BK channels in the nervous system. Front Physiol. 2014, 5, 382. [Google Scholar] [CrossRef] [PubMed]
- Brenner, R.; Jegla, T.J.; Wickenden, A.; Liu, Y.; Aldrich, R.W. Cloning and functional characterization of novel large conductance calcium-activated potassium channel beta subunits, hKCNMB3 and hKCNMB4. J. Biol. Chem. 2000, 275, 6453–6461. [Google Scholar] [CrossRef] [PubMed]
- Wallner, M.; Meera, P.; Toro, L. Molecular basis of fast inactivation in voltage and Ca2+-activated K+ channels: A transmembrane beta-subunit homolog. Proc. Natl. Acad. Sci. USA 1999, 96, 4137–4142. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.M.; Ding, J.P.; Lingle, C.J. Molecular basis for the inactivation of Ca2+- and voltage-dependent BK channels in adrenal chromaffin cells and rat insulinoma tumor cells. J. Neurosci. 1999, 19, 5255–5264. [Google Scholar] [CrossRef]
- Meera, P.; Wallner, M.; Toro, L. A neuronal beta subunit (KCNMB4) makes the large conductance, voltage- and Ca2+-activated K+ channel resistant to charybdotoxin and iberiotoxin. Proc. Natl. Acad. Sci. USA 2000, 97, 5562–5567. [Google Scholar] [CrossRef]
- McLarnon, J.G. Inactivation of a high conductance calcium dependent potassium current in rat hippocampal neurons. Neurosci. Lett. 1995, 193, 5–8. [Google Scholar] [CrossRef]
- Tao, J.; Shi, J.; Liu, Z.R.; Ji, Y.H. Martentoxin: A unique ligand of BK channels. Sheng Li Xue Bao 2012, 64, 355–364. [Google Scholar]
- Tao, J.; Shi, J.; Yan, L.; Chen, Y.; Duan, Y.H.; Ye, P.; Feng, Q.; Zhang, J.W.; Shu, X.Q.; Ji, Y.H. Enhancement effects of martentoxin on glioma BK channel and BK channel (alpha+beta1) subtypes. PLoS ONE 2011, 6, e15896. [Google Scholar] [CrossRef]
- Brenner, R.; Chen, Q.H.; Vilaythong, A.; Toney, G.M.; Noebels, J.L.; Aldrich, R.W. BK channel beta4 subunit reduces dentate gyrus excitability and protects against temporal lobe seizures. Nat. Neurosci. 2005, 8, 1752–1759. [Google Scholar] [CrossRef]
- Petrik, D.; Brenner, R. Regulation of STREX exon large conductance, calcium-activated potassium channels by the beta4 accessory subunit. Neuroscience 2007, 149, 789–803. [Google Scholar] [CrossRef] [PubMed]
- Piwonska, M.; Wilczek, E.; Szewczyk, A.; Wilczynski, G.M. Differential distribution of Ca2+-activated potassium channel beta4 subunit in rat brain: Immunolocalization in neuronal mitochondria. Neuroscience 2008, 153, 446–460. [Google Scholar] [CrossRef] [PubMed]
- Alle, H.; Kubota, H.; Geiger, J.R. Sparse but highly efficient Kv3 outpace BKCa channels in action potential repolarization at hippocampal mossy fiber boutons. J. Neurosci. 2011, 31, 8001–8012. [Google Scholar] [CrossRef] [PubMed]
- Benton, M.D.; Lewis, A.H.; Bant, J.S.; Raman, I.M. Iberiotoxin-sensitive and -insensitive BK currents in Purkinje neuron somata. J. Neurophysiol. 2013, 109, 2528–2541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitmire, L.E.; Ling, L.; Bugay, V.; Carver, C.M.; Timilsina, S.; Chuang, H.H.; Jaffe, D.B.; Shapiro, M.S.; Cavazos, J.E.; Brenner, R. Downregulation of KCNMB4 expression and changes in BK channel subtype in hippocampal granule neurons following seizure activity. PLoS ONE 2017, 12, e0188064. [Google Scholar] [CrossRef] [PubMed]
- Verma-Ahuja, S.; Evans, M.S.; Pencek, T.L. Evidence for decreased calcium dependent potassium conductance in hippocampal CA3 neurons of genetically epilepsy-prone rats. Epilepsy. Res. 1995, 22, 137–144. [Google Scholar] [CrossRef]
- N’Gouemo, P. Targeting BK (big potassium) channels in epilepsy. Expert Opin. Ther. Targets 2011, 15, 1283–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentzen, B.H.; Olesen, S.-P.; Rønn, L.B.; Grunnet, M. BK channel activators and their therapeutic perspectives. Front. Physiol. 2014, 5, 389. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, S.; Feng, Y.; Xiao, Q.; Cheng, J.; Tao, J. The Yin and Yang of BK channels in epilepsy. CNS Neurol. Disord. Drug Targets 2018, 17, 272–279. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AP Waveform Parameter | High Frequency Stimuli | Low Frequency Stimuli | ||
|---|---|---|---|---|
| Control | VSN16R | Control | VSN16R | |
| fAHP1 (mV) | 17.0 ± 1.7 | 14.3 ± 1.6 * | 16.5 ± 1.7 | 17.2 ± 1.4 |
| fAHP2 (mV) | 21.9 ± 1.7 | 19.0 ± 1.6 * | 20.5 ± 1.6 | 20.2 ± 2.1 |
| ISI1 (ms) | 16.3 ± 1.3 | 12.7 ± 1.2 ** | 30.4 ± 4.3 | 26.4 ± 4.4 |
| ISI2 (ms) | 20.1 ± 0.8 | 16.9 ± 1.3 * | 31.2 ± 2.1 | 29.2 ± 2.1 |
| AP1 width (ms) | 4.4 ± 0.4 | 3.3 ± 0.5 * | 4.4 ± 0.3 | 4.0 ± 0.4 |
| AP2 width (ms) | 5.5 ± 0.5 | 4.3 ± 0.8 | 5.2 ± 0.5 | 5.0 ± 0.5 |
| AP Waveform Parameter | High Frequency Stimuli | Low Frequency Stimuli | ||||
|---|---|---|---|---|---|---|
| Control | IBTX | VSN16R | Control | IBTX | VSN16R | |
| fAHP1(mV) | 11.5 ± 1.8 | 20.5 ± 4.4 * | 16.1 ± 12.4 | 11.6 ± 2.2 | 13.6 ± 2.4 | 14.3 ± 2.5 |
| fAHP2(mV) | 18.7 ± 1.5 | 22.9 ± 3.0 | 21.1 ± 2.1 | 17.6 ± 2.5 | 22.7 ± 4.3 | 18.0 ± 2.4 |
| ISI1(ms) | 10.0 ± 0.7 | 10.9 ± 0.9 | 10.9 ± 0.9 | 12.2 ± 1.0 | 13.2 ± 1.1 | 12.2 ± 1.1 |
| ISI2(ms) | 12.7 ± 1.0 | 14.4 ± 1.1 * | 14.7 ± 1.3 | 17.0 ± 1.1 | 17.9 ± 1.4 | 17.3 ± 1.8 |
| AP1 width(ms) | 3.6 ± 0.3 | 4.2 ** ± 0.2 | 3.7 * ± 0.3 | 3.8 ± 0.4 | 4.2 * ± 0.3 | 4.0 ± 0.3 |
| AP2 width(ms) | 4.7 ± 0.4 | 5.7 ± 0.5 | 5.3 ± 0.4 | 4.9 ± 0.4 | 5.5 * ± 0.5 | 5.1 ± 0.4 |
| AP Waveform Parameter | High Frequency Stimuli | Low Frequency Stimuli | ||||
|---|---|---|---|---|---|---|
| Control | 7-Pra-MarTx | VSN16R | Control | 7-Pra-MarTx | VSN16R | |
| fAHP1(mV) | 10.8 ± 1.8 | 13.2 ± 1.7 | 10.0 ± 1.2 | 10.3 ± 1.8 | 11.9 ± 2.0 | 8.3 ± 1.5 ** |
| fAHP2(mV) | 18.4 ± 1.5 | 18.1 ± 1.9 | 15.5 ± 1.0* | 14.8 ± 2.0 | 15.4 ± 2.0 | 12.3 ± 1.7 ** |
| ISI1(ms) | 14.8 ± 1.2 | 14.1 ± 1.3 | 12.3 ± 0.8 * | 27.9 ± 3.9 | 26.1 ± 4.3 | 19.1 ± 2.7 * |
| ISI2(ms) | 20.5 ± 1.7 | 17.6 ± 1.3 | 17.8 ± 1.6 | 37.3 ± 2.6 | 40.2 ± 3.2 | 31.3 ± 3.0 * |
| AP1 width(ms) | 4.0 ± 0.2 | 3.7 * ± 0.2 | 3.3 * ± 0.2 | 4.1 ± 0.3 | 3.8 ± 0.3 | 3.7 ± 0.3 |
| AP2 width(ms) | 5.2 ± 0.6 | 4.9 * ± 0.5 | 5.0 ± 0.5 | 5.3 ± 0.5 | 4.7 * ± 0.4 | 5.2 ± 0.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabatabaee, S.; Baker, D.; Selwood, D.L.; Whalley, B.J.; Stephens, G.J. The Cannabinoid-Like Compound, VSN16R, Acts on Large Conductance, Ca2+-Activated K+ Channels to Modulate Hippocampal CA1 Pyramidal Neuron Firing. Pharmaceuticals 2019, 12, 104. https://doi.org/10.3390/ph12030104
Tabatabaee S, Baker D, Selwood DL, Whalley BJ, Stephens GJ. The Cannabinoid-Like Compound, VSN16R, Acts on Large Conductance, Ca2+-Activated K+ Channels to Modulate Hippocampal CA1 Pyramidal Neuron Firing. Pharmaceuticals. 2019; 12(3):104. https://doi.org/10.3390/ph12030104
Chicago/Turabian StyleTabatabaee, Setareh, David Baker, David L. Selwood, Benjamin J. Whalley, and Gary J. Stephens. 2019. "The Cannabinoid-Like Compound, VSN16R, Acts on Large Conductance, Ca2+-Activated K+ Channels to Modulate Hippocampal CA1 Pyramidal Neuron Firing" Pharmaceuticals 12, no. 3: 104. https://doi.org/10.3390/ph12030104
APA StyleTabatabaee, S., Baker, D., Selwood, D. L., Whalley, B. J., & Stephens, G. J. (2019). The Cannabinoid-Like Compound, VSN16R, Acts on Large Conductance, Ca2+-Activated K+ Channels to Modulate Hippocampal CA1 Pyramidal Neuron Firing. Pharmaceuticals, 12(3), 104. https://doi.org/10.3390/ph12030104