
The Potential Advantage of Targeting Both PD-L1/PD-L2/PD-1 and IL-10–IL-10R Pathways in Acute Myeloid Leukemia

 , , ,
, , ,  and
and 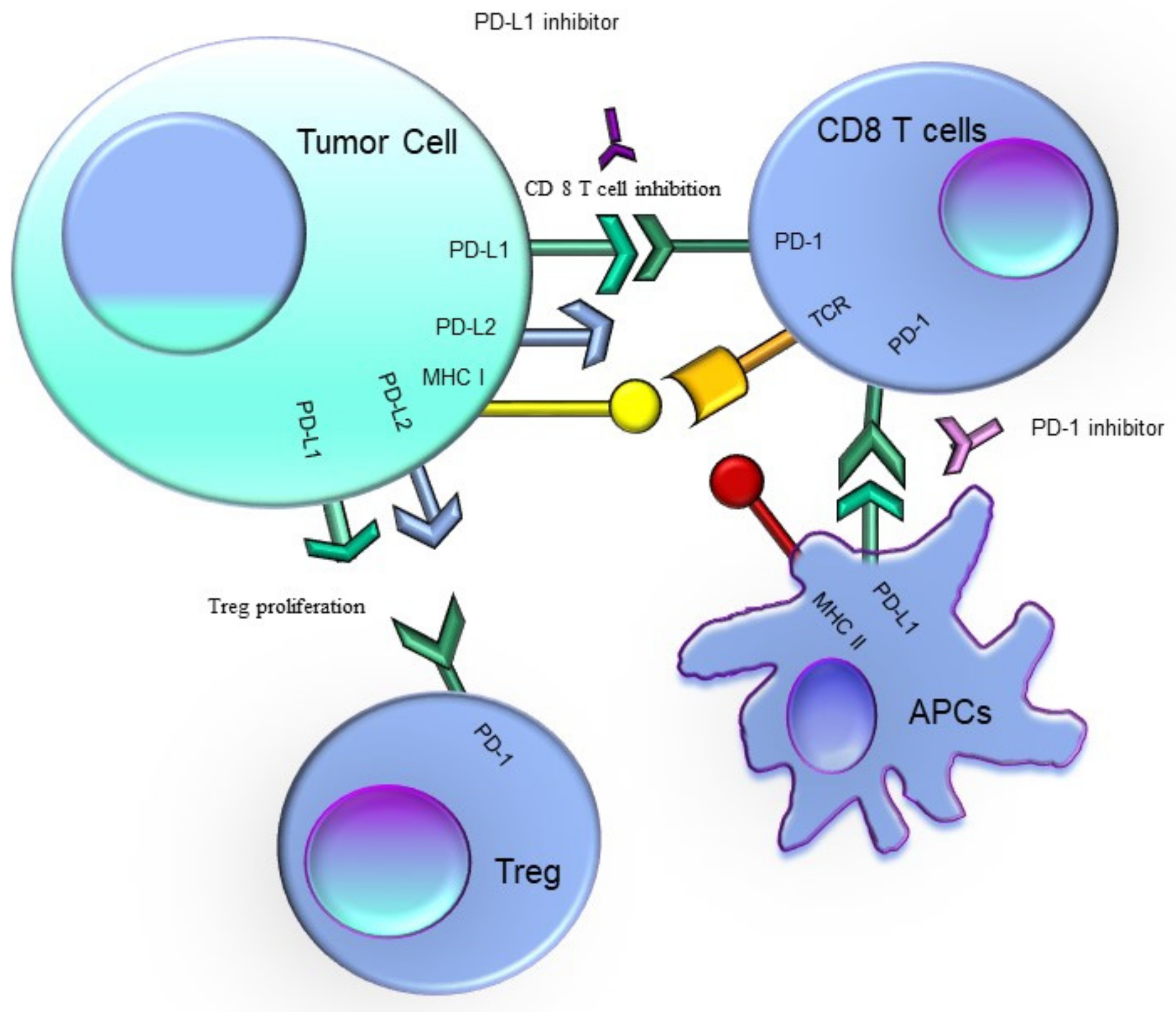{kind=link}
Abstract
:1. Introduction
2. IL-10
2.1. IL-10 and Tumor Progression
2.2. IL-10 as an Anti-Tumor Cytokine
2.3. IL-10 and AML
3. PD-1–PD-L1 Pathway Suppresses the Type 1 T-Cell Responses Which Fight Cancer
4. IL-10 Increases PD-L1 on Malignant Cells
5. PD-1/PD-L1 in AML
6. Targeting the PD-1 Signaling Pathway in Cancer Restored Tumor-Specific T Cell Effector Functions
7. Consequences of the Hypothesis and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Okazaki, T.; Honjo, T. The Pd-1-Pd-L Pathway in Immunological Tolerance. Trends Immunol. 2006, 27, 195–201. [Google Scholar] [CrossRef]
- Sharpe, A.H.; Wherry, E.J.; Ahmed, R.; Freeman, G.J. The Function of Programmed Cell Death 1 and Its Ligands in Regulating Autoimmunity and Infection. Nat. Immunol. 2007, 8, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. Pd-1 and Its Ligands in Tolerance and Immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiner, B.; Mitsdoerffer, B.C.; Kieseier, L.; Chen, H.P.; Hartung, M.W.; Wiendl, H. Interferon-Beta Enhances Monocyte and Dendritic Cell Expression of B7-H1 (Pd-L1), a Strong Inhibitor of Autologous T-Cell Activation: Relevance for the Immune Modulatory Effect in Multiple Sclerosis. J. Neuroimmunol. 2004, 155, 172–182. [Google Scholar] [CrossRef]
- Kim, J.; Myers, A.C.; Chen, L.; Pardoll, D.M.; Truong-Tran, Q.A.; Lane, A.P.; McDyer, J.F.; Fortuno, L.; Schleimer, R.P. Constitutive and Inducible Expression of B7 Family of Ligands by Human Airway Epithelial Cells. Am. J. Respir. Cell Mol. 2005, 33, 280–299. [Google Scholar] [CrossRef] [PubMed]
- Stanciu, L.A.; Bellettato, C.M.; Laza-Stanca, V.; Coyle, A.J.; Papi, A.; Johnston, S.L. Expression of Programmed Death-1 Ligand (Pd-L) 1, Pd-L2, B7-H3, and Inducible Costimulator Ligand on Human Respiratory Tract Epithelial Cells and Regulation by Respiratory Syncytial Virus and Type 1 and 2 Cytokines. J. Infect. Dis. 2006, 193, 404–412. [Google Scholar] [CrossRef]
- Boasso, A.; Hardy, A.W.; Landay, A.L.; Martinson, J.L.; Anderson, S.A.; Dolan, M.J.; Clerici, M.; Shearer, G.M. Pdl-1 Upregulation on Monocytes and T Cells by Hiv Via Type I Interferon: Restricted Expression of Type I Interferon Receptor by Ccr5-Expressing Leukocytes. Clin. Immunol. 2008, 129, 132–144. [Google Scholar] [CrossRef] [Green Version]
- Haile, S.T.; Bosch, J.J.; Agu, N.I.; Zeender, A.M.; Somasundaram, P.; Srivastava, M.K.; Britting, S.; Wolf, J.B.; Ksander, B.R.; Ostrand-Rosenberg, S. Tumor Cell Programmed Death Ligand 1-Mediated T Cell Suppression Is Overcome by Coexpression of Cd80. J. Immunol. 2011, 186, 6822–6829. [Google Scholar] [CrossRef]
- Terawaki, S.S.; Chikuma, S.; Shibayama, T.; Hayashi, T.; Yoshida, T.O.; Honjo, T. Ifn-Alpha Directly Promotes Programmed Cell Death-1 Transcription and Limits the Duration of T Cell-Mediated Immunity. J. Immunol. 2011, 186, 2772–2779. [Google Scholar] [CrossRef] [PubMed]
- Telcian, A.G.; Laza-Stanca, V.; Edwards, M.R.; Harker, J.A.; Wang, H.; Bartlett, N.W.; Mallia, P.; Zdrenghea, M.T.; Kebadze, T.; Coyle, A.J.; et al. RSV-Induced Bronchial Epithelial Cell PD-L1 Expression Inhibits CD8+ T Cell Nonspecific Antiviral Activity. J. Infect. Dis. 2011, 203, 85–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latchman, Y.E.; Wood, C.R.; Chernova, T.; Chaudhary, D.; Borde, M.; Chernova, I.; Iwai, Y.; Long, A.J.; Brown, J.A.; Nunes, R.J.; et al. PD-L2 is a second ligand for PD-1 and inhibits T cell activation. Nat. Immunol. 2001, 2, 261–268. [Google Scholar] [CrossRef]
- Loke, P.; Allison, J.P. PD-L1 and PD-L2 are differentially regulated by Th1 and Th2 cells. Proc. Natl. Acad. Sci. USA 2003, 100, 5336–5341. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.A.; Dorfman, D.M.; Ma, F.-R.; Sullivan, E.L.; Munoz, O.; Wood, C.R.; Greenfield, E.A.; Freeman, G.J. Blockade of Programmed Death-1 Ligands on Dendritic Cells Enhances T Cell Activation and Cytokine Production. J. Immunol. 2003, 170, 1257–1266. [Google Scholar] [CrossRef]
- Wang, S.; Bajorath, J.; Flies, D.B.; Dong, H.; Honjo, T.; Chen, L. Molecular Modeling and Functional Mapping of B7-H1 and B7-DC Uncouple Costimulatory Function from PD-1 Interaction. J. Exp. Med. 2003, 197, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Ishiwata, K.; Watanabe, N.; Guo, M.; Tomihara, K.; Brumlik, M.J.; Yagita, H.; Pardoll, E.; Chen, L.; Shin, T. Costimulator B7-DC attenuates strong Th2 responses induced by Nippostrongylus brasiliensis. J. Immunol. 2010, 184, 2086–2094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messal, N.; Serriari, N.-E.; Pastor, S.; Nunès, J.A.; Olive, D. PD-L2 is expressed on activated human T cells and regulates their function. Mol. Immunol. 2011, 48, 2214–2219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesterhuis, W.J.; Steer, H.; Lake, R.A. PD-L2 is predominantly expressed by Th2 cells. Mol. Immunol. 2011, 49, 1–3. [Google Scholar] [CrossRef]
- Rodriguez-Garcia, M.; Porichis, F.; de Jong, O.G.; Levi, K.; Diefenbach, T.J.; Lifson, J.D.; Freeman, G.J.; Walker, B.D.; Kaufmann, D.E.; Kavanagh, D.G. Expression of Pd-L1 and Pd-L2 on Human Macrophages Is up-Regulated by Hiv-1 and Differentially Modulated by Il-10. J. Leukoc. Biol. 2011, 89, 507–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norde, W.J.; Maas, F.; Hobo, W.; Korman, A.; Quigley, M.; Kester, M.G.; Hebeda, K.; Falkenburg, J.F.; Schaap, N.; de Witte, T.M.; et al. PD-1/PD-L1 Interactions Contribute to Functional T-Cell Impairment in Patients Who Relapse with Cancer After Allogeneic Stem Cell Transplantation. Cancer Res. 2011, 71, 5111–5122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, G.J.; Long, A.J.; Iwai, Y.; Bourque, K.; Chernova, T.; Nishimura, H.; Fitz, L.J.; Malenkovich, N.; Okazaki, T.; Byrne, M.C.; et al. Engagement of the Pd-1 Immunoinhibitory Receptor by a Novel B7 Family Member Leads to Negative Regulation of Lymphocyte Activation. J. Exp. Med. 2000, 192, 1027–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, S.-Y.; Otsuji, M.; Gorski, K.; Huang, X.; Slansky, J.E.; Pai, S.I.; Shalabi, A.; Shin, T.; Pardoll, D.M.; Tsuchiya, H. B7-Dc, a New Dendritic Cell Molecule with Potent Costimulatory Properties for T Cells. J. Exp. Med. 2001, 193, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Youngnak, P.; Kozono, Y.; Kozono, H.; Iwai, H.; Otsuki, N.; Jin, H.; Omura, K.; Yagita, H.; Pardoll, D.M.; Chen, L.; et al. Differential binding properties of B7-H1 and B7-DC to programmed death-1. Biochem. Biophys. Res. Commun. 2003, 307, 672–677. [Google Scholar] [CrossRef]
- Ghiotto, M.; Gauthier, L.; Serriari, N.; Pastor, S.; Truneh, A.; Nunes, J.; Olive, D. PD-L1 and PD-L2 differ in their molecular mechanisms of interaction with PD-1. Int. Immunol. 2010, 22, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, H.; Nose, M.; Hiai, H.; Minato, N.; Honjo, T. Development of Lupus-like Autoimmune Diseases by Disruption of the PD-1 Gene Encoding an ITIM Motif-Carrying Immunoreceptor. Immunity 1999, 11, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Carter, L.; Fouser, L.A.; Jussif, J.; Fitz, L.; Deng, B.; Wood, C.R.; Collins, M.; Honjo, T.; Freeman, G.J.; Carreno, B.M. Pd-1:Pd-L Inhibitory Pathway Affects Both Cd4(+) and Cd8(+) T Cells and Is Overcome by Il-2. Eur. J. Immunol. 2002, 32, 634–643. [Google Scholar] [CrossRef]
- Mazanet, M.M.; Hughes, C.C. B7-H1 Is Expressed by Human Endothelial Cells and Suppresses T Cell Cytokine Synthesis. J. Immunol. 2002, 169, 3581–3588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, J.L. PD-1 signaling in primary T cells. Immunol. Rev. 2009, 229, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.J.; Ni, L.; Zhang, Y.; Zhang, C.L.; Wu, X.Y.; Atia, A.N.; Thayer, P.; Moorman, J.P.; Yao, Z.Q. Pd-1 Negatively Regulates Interleukin-12 Expression by Limiting Stat-1 Phosphorylation in Monocytes/Macrophages During Chronic Hepatitis C Virus Infection. Immunology 2011, 132, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ma, C.J.; Ni, L.; Zhang, C.L.; Wu, X.Y.; Kumaraguru, U.; Li, C.F.; Moorman, J.P.; Yao, Z.Q. Cross-Talk between Programmed Death-1 and Suppressor of Cytokine Signaling-1 in Inhibition of IL-12 Production by Monocytes/Macrophages in Hepatitis C Virus Infection. J. Immunol. 2011, 186, 3093–3103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azuma, T.; Yao, S.; Zhu, G.; Flies, A.; Flies, S.J.; Chen, L. B7-H1 is a ubiquitous antiapoptotic receptor on cancer cells. Blood 2008, 111, 3635–3643. [Google Scholar] [CrossRef] [PubMed]
- Butte, M.; Keir, M.E.; Phamduy, T.B.; Sharpe, A.H.; Freeman, G.J. Programmed Death-1 Ligand 1 Interacts Specifically with the B7-1 Costimulatory Molecule to Inhibit T Cell Responses. Immunity 2007, 27, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-J.; Omiya, R.; Matsumura, Y.; Sakoda, Y.; Kuramasu, A.; Augustine, M.M.; Yao, S.; Tsushima, F.; Narazaki, H.; Anand, S.; et al. B7-H1/CD80 interaction is required for the induction and maintenance of peripheral T-cell tolerance. Blood 2010, 116, 1291–1298. [Google Scholar] [CrossRef] [PubMed]
- Flies, D.B.; Wang, S.; Xu, H.; Chen, L. Cutting Edge: A Monoclonal Antibody Specific for the Programmed Death-1 Homolog Prevents Graft-versus-Host Disease in Mouse Models. J. Immunol. 2011, 187, 1537–1541. [Google Scholar] [CrossRef]
- Zhu, Y.; Yao, S.; Chen, L. Cell Surface Signaling Molecules in the Control of Immune Responses: A Tide Model. Immunity 2011, 34, 466–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, T.; Kennedy, G.; Gorski, K.; Tsuchiya, H.; Koseki, H.; Azuma, M.; Yagita, H.; Chen, L.; Powell, J.; Pardoll, D.; et al. Cooperative B7-1/2 (Cd80/Cd86) and B7-Dc Costimulation of Cd4+ T Cells Independent of the Pd-1 Receptor. J. Exp. Med. 2003, 198, 31–38. [Google Scholar] [CrossRef]
- Shin, T.; Yoshimura, K.; Shin, T.; Crafton, E.B.; Tsuchiya, H.; Housseau, F.; Koseki, H.; Schulick, R.D.; Chen, L.; Pardoll, D.M. In vivo costimulatory role of B7-DC in tuning T helper cell 1 and cytotoxic T lymphocyte responses. J. Exp. Med. 2005, 201, 1531–1541. [Google Scholar] [CrossRef]
- Fukushima, A.; Yamaguchi, T.; Azuma, M.; Yagita, H.; Ueno, H. Involvement of programmed death-ligand 2 (PD-L2) in the development of experimental allergic conjunctivitis in mice. Br. J. Ophthalmol. 2006, 90, 1040–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, K.; Fukuyama, S.; Eguchi-Tsuda, M.; Nakano, T.; Matsumoto, T.; Matsumura, M.; Moriwaki, A.; Kan-o, K.; Wada, Y.; Yagita, H.; et al. B7-DC induced by IL-13 works as a feedback regulator in the effector phase of allergic asthma. Biochem. Biophys. Res. Commun. 2008, 365, 170–175. [Google Scholar] [CrossRef]
- Zhang, J.M.; An, J. Cytokines, Inflammation, and Pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couper, K.; Blount, D.G.; Riley, E.M. IL-10: The Master Regulator of Immunity to Infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef] [PubMed]
- Jofra, T.; Galvani, G.; Cosorich, I.; de Giorgi, L.; Annoni, A.; Vecchione, A.; Sorini, C.; Falcone, M.; Fousteri, G. Experimental Colitis in Il-10-Deficient Mice Ameliorates in the Absence of Ptpn22. Clin. Exp. Immunol. 2019, 197, 263–275. [Google Scholar] [PubMed] [Green Version]
- De Smedt, T.; van Mechelen, M.; de Becker, G.; Urbain, J.; Leo, O.; Moser, M. Effect of interleukin-10 on dendritic cell maturation and function. Eur. J. Immunol. 1997, 27, 1229–1235. [Google Scholar] [CrossRef]
- O'Farrell, A.; Liu, Y.; Moore, K.W.; Mui, A.L. IL-10 inhibits macrophage activation and proliferation by distinct signaling mechanisms: Evidence for Stat3-dependent and -independent pathways. EMBO J. 1998, 17, 1006–1018. [Google Scholar] [CrossRef]
- Mannino, M.H.; Zhu, Z.; Xiao, H.; Bai, Q.; Wakefield, M.; Fang, Y. The paradoxical role of IL-10 in immunity and cancer. Cancer Lett. 2015, 367, 103–107. [Google Scholar] [CrossRef]
- Chen, W.F.; Zlotnik, A. IL-10: A novel cytotoxic T cell differentiation factor. J. Immunol. 1991, 147, 528–534. [Google Scholar]
- Wang, Z.; Guan, D.; Huo, J.; Biswas, S.K.; Huang, Y.; Yang, Y.; Xu, S.; Lam, K.-P. IL-10 Enhances Human Natural Killer Cell Effector Functions via Metabolic Reprogramming Regulated by mTORC1 Signaling. Front. Immunol. 2021, 12, 619195. [Google Scholar] [CrossRef]
- Polukort, S.H.; Rovatti, J.; Carlson, L.; Thompson, C.; Ser-Dolansky, J.; Kinney, S.R.; Schneider, S.S.; Mathias, C.B. Il-10 Enhances Ige-Mediated Mast Cell Responses and Is Essential for the Development of Experimental Food Allergy in Il-10-Deficient Mice. J. Immunol. 2016, 196, 4865–4876. [Google Scholar] [CrossRef] [Green Version]
- Rafiq, K.; Charitidou, L.; Bullens, D.M.A.; Kasran, A.; Lorre, K.; Ceuppens, J.L.; van Gool, S.W. Regulation of the Il-10 Production by Human T Cells. Scand. J. Immunol. 2001, 53, 139–147. [Google Scholar] [CrossRef]
- Berti, F.C.B.; Oliveira, K.B. Il-10 in Cancer: Just a Classical Immunosuppressive Factor or Also an Immunostimulating One? AIMS Allergy Immunol. 2018, 2, 88–97. [Google Scholar] [CrossRef]
- Chen, L.; Shi, Y.; Zhu, X.; Guo, W.; Zhang, M.; Che, Y.; Tang, L.; Yang, X.; You, Q.; Liu, Z. IL-10 secreted by cancer-associated macrophages regulates proliferation and invasion in gastric cancer cells via c-Met/STAT3 signaling. Oncol. Rep. 2019, 42, 595–604. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, Y.; Shou, L.; Shi, Y.; Shen, H.; Zhu, M.; Ye, X.; Jin, J.; Xie, W. Increased Serum Level of Interleukin-10 Predicts Poor Survival and Early Recurrence in Patients with Peripheral T-Cell Lymphomas. Front. Oncol. 2020, 10, 584261. [Google Scholar] [CrossRef]
- Zeng, L.; O’Connor, C.; Zhang, J.; Kaplan, A.M.; Cohen, D.A. IL-10 promotes resistance to apoptosis and metastatic potential in lung tumor cell lines. Cytokine 2010, 49, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Zeni, E.; Mazzetti, L.; Miotto, D.; Cascio, N.L.; Maestrelli, P.; Querzoli, P.; Pedriali, M.; de Rosa, E.; Fabbri, L.M.; Mapp, C.E.; et al. Macrophage expression of interleukin-10 is a prognostic factor in nonsmall cell lung cancer. Eur. Respir. J. 2007, 30, 627–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, L.; Qi, Q.; Wang, P.; Chen, H.; Chen, Z.; Meng, Z.; Liu, L. Serum levels of IL-6, IL-8, and IL-10 are indicators of prognosis in pancreatic cancer. J. Int. Med. Res. 2018, 46, 5228–5236. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Wu, D.; Wu, P.; Wang, Z.; Huang, J. Serum IL-10 Predicts Worse Outcome in Cancer Patients: A Meta-Analysis. PLoS ONE 2015, 10, e0139598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soria, J.-C.; Moon, C.; Kemp, B.L.; Liu, D.D.; Feng, L.; Tang, X.; Chang, Y.S.; Mao, L.; Khuri, F.R. Lack of interleukin-10 expression could predict poor outcome in patients with stage I non-small cell lung cancer. Clin. Cancer Res. 2003, 9, 1785–1791. [Google Scholar]
- Toiyama, Y.; Miki, C.; Inoue, Y.; Minobe, S.; Urano, H.; Kusunoki, M. Loss of tissue expression of interleukin-10 promotes the disease progression of colorectal carcinoma. Surg. Today 2009, 40, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Choucair, K.; Kelso, J.D.; Duff, J.R.; Cassidy, C.S.; Albrethsen, M.; Ashraf, M.; Verghese, C.; Oft, M.; Brunicardi, F.C.; Dworkin, L.; et al. Interleukin 10-Mediated Response and Correlated Anemia in a Patient with Advanced Non-Small Cell Lung Carcinoma. Case Rep. Oncol. 2019, 12, 297–303. [Google Scholar] [CrossRef]
- Emmerich, J.; Mumm, J.B.; Chan, I.; LaFace, D.; Truong, H.; McClanahan, T.K.; Gorman, D.M.; Oft, M. IL-10 Directly Activates and Expands Tumor-Resident CD8+ T Cells without De Novo Infiltration from Secondary Lymphoid Organs. Cancer Res. 2012, 72, 3570–3581. [Google Scholar] [CrossRef] [Green Version]
- Tanikawa, T.; Wilke, C.M.; Kryczek, I.; Chen, G.Y.; Kao, J.; Núñez, G.; Zou, W. Interleukin-10 Ablation Promotes Tumor Development, Growth, and Metastasis. Cancer Res. 2012, 72, 420–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naing, A.; Infante, J.R.; Papadopoulos, K.P.; Chan, I.H.; Shen, C.; Ratti, N.P.; Rojo, B.; Autio, K.A.; Wong, D.J.; Patel, M.R.; et al. Pegylated Il-10 (Pegilodecakin) Induces Systemic Immune Activation, Cd8(+) T Cell Invigoration and Polyclonal T Cell Expansion in Cancer Patients. Cancer Cell 2018, 34, 775–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecht, J.R.; Lonardi, S.; Bendell, J.C.; Sim, H.-W.; Macarulla, T.; Lopez, C.D.; van Cutsem, E.; Martin, A.J.M.; Park, J.O.; Greil, R.; et al. Randomized Phase III Study of FOLFOX Alone and with Pegilodecakin as Second-line Therapy in Patients with Metastatic Pancreatic Cancer (SEQUOIA). J. Clin. Oncol. 2020, 38, 637. [Google Scholar] [CrossRef]
- Spigel, D.; Jotte, R.; Nemunaitis, J.; Shum, M.; Schneider, J.; Goldschmidt, J.; Eisenstein, J.; Berz, D.; Seneviratne, L.; Socoteanu, M.; et al. Randomized Phase 2 Studies of Checkpoint Inhibitors Alone or in Combination with Pegilodecakin in Patients with Metastatic NSCLC (CYPRESS 1 and CYPRESS 2). J. Thorac. Oncol. 2020, 16, 327–333. [Google Scholar] [CrossRef]
- Bruserud, Ø.; Gjertsen, T.; Brustugun, O.T.; Bassøe, C.F.; Nesthus, I.; Akselsen, E.; Bühring, H.J.; Pawelec, G. Effects of Interleukin 10 on Blast Cells Derived from Patients with Acute Myelogenous Leukemia. Leukemia 1999, 11, 1910–1920. [Google Scholar]
- Bruserud, O. IL-4, IL-10 and IL-13 in acute myelogenous leukemia. Cytokines Cell. Mol. Ther. 1998, 4, 187–198. [Google Scholar]
- Westermann, F.; Kube, D.; Haier, B.; Bohlen, H.; Engert, A.; Zuehlsdorf, M.; Diehl, V.; Tesch, H. Interleukin 10 inhibits cytokine production of human AML cells. Ann. Oncol. 1996, 7, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Asano, Y.; Shibata, S.; Kobayashi, S.; Okamura, S.; Niho, Y. Interleukin-10 inhibits the autocrine growth of leukemic blast cells from patients with acute myeloblastic leukemia. Int. J. Hematol. 1997, 66, 445–450. [Google Scholar] [CrossRef]
- Sanchez-Correa, B.; Bergua, J.M.; Campos, C.; Gayoso, I.; Arcos, M.J.; Bañas, H.; Morgado, S.; Casado, J.G.; Solana, R.; Tarazona, R. Cytokine profiles in acute myeloid leukemia patients at diagnosis: Survival is inversely correlated with IL-6 and directly correlated with IL-10 levels. Cytokine 2013, 61, 885–891. [Google Scholar] [CrossRef]
- Kornblau, S.M.; McCue, D.; Singh, N.; Chen, W.; Estrov, Z.; Coombes, K. Recurrent expression signatures of cytokines and chemokines are present and are independently prognostic in acute myelogenous leukemia and myelodysplasia. Blood 2010, 116, 4251–4261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Guardia, R.D.; Lopez-Millan, B.; Lavoie, J.R.; Bueno, C.; Castaño, J.; Gómez-Casares, M.; Vives, S.; Palomo, L.; Juan, M.; Delgado, J.; et al. Detailed Characterization of Mesenchymal Stem/Stromal Cells from a Large Cohort of Aml Patients Demonstrates a Definitive Link to Treatment Outcomes. Stem Cell Rep. 2017, 8, 1573–1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Q.; Li, G.; Fozza, C.; Wang, S.; Yang, S.; Sang, Y.; Liu, X.; Chen, C. Levels and Clinical Significance of Regulatory B Cells and T Cells in Acute Myeloid Leukemia. BioMed Res. Int. 2020, 2020, 7023168. [Google Scholar] [CrossRef]
- Musuraca, G.; de Matteis, S.; Napolitano, R.; Papayannidis, C.; Guadagnuolo, V.; Fabbri, F.; Cangini, D.; Ceccolini, M.; Giannini, M.B.; Lucchesi, A.; et al. Il-17/Il-10 Double-Producing T Cells: New Link between Infections, Immunosuppression and Acute Myeloid Leukemia. J. Transl. Med. 2015, 13, 229. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Wan, Y.; Liu, J.; Wang, Y.; Li, S.; Xing, H.; Tang, K.; Tian, Z.; Rao, Q.; Wang, M.; et al. Regulatory T Cells Promote the Stemness of Acute Myeloid Leukemia Cells through Il10 Cytokine Related Signaling Pathway. Blood 2017, 130, 2727. [Google Scholar]
- Park, H.H.; Kim, M.; Lee, B.-H.; Lim, J.; Kim, Y.; Lee, E.J.; Min, W.S.; Kang, C.S.; Kim, W.I.; Shim, S.I.; et al. Intracellular IL-4, IL-10, and IFN-gamma levels of leukemic cells and bone marrow T cells in acute leukemia. Ann. Clin. Lab. Sci. 2006, 36, 7–15. [Google Scholar]
- Kotenko, S.V.; Krause, C.D.; Izotova, L.S.; Pollack, B.P.; Wu, W.; Pestka, S. Identification and functional characterization of a second chain of the interleukin-10 receptor complex. EMBO J. 1997, 16, 5894–5903. [Google Scholar] [CrossRef] [Green Version]
- Deng, J.; Theodore, C.; Standiford, J. Interleukin-10. In Encyclopedia of Respiratory Medicine, 2nd ed.; Sam, M.J., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 295–302. [Google Scholar]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. Interleukin-10 and the Interleukin-10 Receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef]
- Chen, N.; Xu, Y.; Mou, J.; Rao, Q.; Xing, H.; Tian, Z.; Tang, K.; Wang, M.; Wang, J. Targeting of IL-10R on acute myeloid leukemia blasts with chimeric antigen receptor-expressing T cells. Blood Cancer J. 2021, 11, 144. [Google Scholar] [CrossRef] [PubMed]
- Zou, W.; Chen, L. Inhibitory B7-family molecules in the tumour microenvironment. Nat. Rev. Immunol. 2008, 8, 467–477. [Google Scholar] [CrossRef] [PubMed]
- De Monte, L.; Reni, M.; Tassi, E.; Clavenna, D.; Papa, I.; Recalde, H.; Braga, M.; di Carlo, V.; Doglioni, C.; Protti, M.P. Intratumor T Helper Type 2 Cell Infiltrate Correlates with Cancer-Associated Fibroblast Thymic Stromal Lymphopoietin Production and Reduced Survival in Pancreatic Cancer. J. Exp. Med. 2011, 208, 469–478. [Google Scholar] [CrossRef]
- Rozali, E.N.; Hato, S.V.; Robinson, B.W.; Lake, R.A.; Lesterhuis, W.J. Programmed Death Ligand 2 in Cancer-Induced Immune Suppression. Clin. Dev. Immunol. 2012, 2012, 656340. [Google Scholar] [CrossRef] [PubMed]
- Pardoll, D.M. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef] [Green Version]
- Iwai, Y.; Ishida, M.; Tanaka, Y.; Okazaki, T.; Honjo, T.; Minato, N. Involvement of PD-L1 on tumor cells in the escape from host immune system and tumor immunotherapy by PD-L1 blockade. Proc. Natl. Acad. Sci. USA 2002, 99, 12293–12297. [Google Scholar] [CrossRef] [Green Version]
- Rodig, N.; Ryan, T.; Allen, J.A.; Pang, H.; Grabie, N.; Chernova, T.; Greenfield, E.A.; Liang, S.C.; Sharpe, A.H.; Lichtman, A.H.; et al. Endothelial expression of PD-L1 and PD-L2 down-regulates CD8+ T cell activation and cytolysis. Eur. J. Immunol. 2003, 33, 3117–3126. [Google Scholar] [CrossRef]
- Schreiner, B.; Bailey, S.L.; Shin, T.; Chen, L.; Miller, S.D. PD-1 ligands expressed on myeloid-derived APC in the CNS regulate T-cell responses in EAE. Eur. J. Immunol. 2008, 38, 2706–2717. [Google Scholar] [CrossRef] [Green Version]
- Keir, M.E.; Liang, S.C.; Guleria, I.; Latchman, Y.E.; Qipo, A.; Albacker, L.; Koulmanda, M.; Freeman, G.J.; Sayegh, M.H.; Sharpe, A.H. Tissue expression of PD-L1 mediates peripheral T cell tolerance. J. Exp. Med. 2006, 203, 883–895. [Google Scholar] [CrossRef]
- Dong, H.; Strome, S.E.; Salomao, D.R.; Tamura, H.; Hirano, F.; Flies, D.B.; Roche, P.C.; Lu, J.; Zhu, G.; Tamada, K.; et al. Tumor-associated B7-H1 promotes T-cell apoptosis: A potential mechanism of immune evasion. Nat. Med. 2002, 8, 793–800. [Google Scholar] [CrossRef]
- Wherry, E.J. T Cell Exhaustion. Nat. Immunol. 2011, 12, 492–499. [Google Scholar] [CrossRef]
- Wang, L.; Pino-Lagos, K.; de Vries, V.C.; Guleria, I.; Sayegh, M.H.; Noelle, R.J. Programmed death 1 ligand signaling regulates the generation of adaptive Foxp3+CD4+ regulatory T cells. Proc. Natl. Acad. Sci. USA 2008, 105, 9331–9336. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Lau, R.; Yu, D.; Zhu, W.; Korman, A.; Weber, J. Pd1 Blockade Reverses the Suppression of Melanoma Antigen-Specific Ctl by Cd4+ Cd25(Hi) Regulatory T Cells. Int. Immunol. 2009, 21, 1065–1077. [Google Scholar] [CrossRef] [Green Version]
- Fife, B.T.; Pauken, K.E.; Eagar, T.N.; Obu, T.; Wu, J.; Tang, Q.; Azuma, M.; Krummel, M.F.; Bluestone, J.A. Interactions between PD-1 and PD-L1 promote tolerance by blocking the TCR–induced stop signal. Nat. Immunol. 2009, 10, 1185–1192. [Google Scholar] [CrossRef]
- Hino, R.; Kabashima, K.; Kato, Y.; Yagi, H.; Nakamura, M.; Honjo, T.; Okazaki, T.; Tokura, Y. Tumor cell expression of programmed cell death-1 ligand 1 is a prognostic factor for malignant melanoma. Cancer 2010, 116, 1757–1766. [Google Scholar] [CrossRef]
- Taube, J.M.; Anders, R.A.; Young, G.D.; Xu, H.; Sharma, R.; McMiller, T.L.; Chen, S.; Klein, A.P.; Pardoll, D.M.; Topalian, S.L.; et al. Colocalization of Inflammatory Response with B7-H1 Expression in Human Melanocytic Lesions Supports an Adaptive Resistance Mechanism of Immune Escape. Sci. Transl. Med. 2012, 4, 127. [Google Scholar] [CrossRef] [Green Version]
- Butte, M.; Peña-Cruz, V.; Kim, M.-J.; Freeman, G.J.; Sharpe, A.H. Interaction of human PD-L1 and B7-1. Mol. Immunol. 2008, 45, 3567–3572. [Google Scholar] [CrossRef] [Green Version]
- Frigola, X.; Inman, B.; Krco, C.J.; Liu, X.; Harrington, S.M.; Bulur, P.A.; Dietz, A.; Dong, H.; Kwon, E.D. Soluble B7-H1: Differences in production between dendritic cells and T cells. Immunol. Lett. 2011, 142, 78–82. [Google Scholar] [CrossRef] [Green Version]
- Frigola, X.; Inman, B.; Lohse, C.M.; Krco, C.J.; Cheville, J.C.; Thompson, R.H.; Leibovich, B.; Blute, M.L.; Dong, H.; Kwon, E.D. Identification of a Soluble Form of B7-H1 That Retains Immunosuppressive Activity and Is Associated with Aggressive Renal Cell Carcinoma. Clin. Cancer Res. 2011, 17, 1915–1923. [Google Scholar] [CrossRef] [Green Version]
- Said, E.; Dupuy, F.P.; Trautmann, L.; Zhang, Y.; Shi, Y.; El-Far, M.; Hill, B.J.; Noto, A.; Ancuta, P.; Peretz, Y.; et al. Programmed death-1–induced interleukin-10 production by monocytes impairs CD4+ T cell activation during HIV infection. Nat. Med. 2010, 16, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Selenko-Gebauer, N.; Majdic, O.; Szekeres, A.; Höfler, G.; Guthann, E.; Korthäuer, U.; Zlabinger, G.; Steinberger, P.; Pickl, W.F.; Stockinger, H.; et al. B7-H1 (programmed death-1 ligand) on dendritic cells is involved in the induction and maintenance of T cell anergy. J. Immunol. 2003, 170, 3637–3644. [Google Scholar] [CrossRef] [Green Version]
- Kuang, D.-M.; Zhao, Q.; Peng, C.; Xu, J.; Zhang, J.-P.; Wu, C.; Zheng, L. Activated monocytes in peritumoral stroma of hepatocellular carcinoma foster immune privilege and disease progression through PD-L1. J. Exp. Med. 2009, 206, 1327–1337. [Google Scholar] [CrossRef]
- Kuang, D.-M.; Wu, Y.; Chen, N.; Cheng, J.; Zhuang, S.-M.; Zheng, L. Tumor-derived hyaluronan induces formation of immunosuppressive macrophages through transient early activation of monocytes. Blood 2007, 110, 587–595. [Google Scholar] [CrossRef]
- Huber, S.; Hoffmann, R.; Muskens, F.; Voehringer, D. Alternatively activated macrophages inhibit T-cell proliferation by Stat6-dependent expression of PD-L2. Blood 2010, 116, 3311–3320. [Google Scholar] [CrossRef] [Green Version]
- Bögels, M.; Braster, R.; Nijland, P.G.; Gül, N.; van de Luijtgaarden, W.; Fijneman, R.J.; Meijer, G.A.; Jimenez, C.R.; Beelen, R.H.; van Egmond, M. Carcinoma origin dictates differential skewing of monocyte function. OncoImmunology 2012, 1, 798–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darrah, P.A.; Hegde, S.T.; Patel, D.T.; Lindsay, R.W.B.; Chen, L.; Roederer, M.; Seder, R.A. IL-10 production differentially influences the magnitude, quality, and protective capacity of Th1 responses depending on the vaccine platform. J. Exp. Med. 2010, 207, 1421–1433. [Google Scholar] [CrossRef] [Green Version]
- Rigopoulou, E.I.; Abbott, W.G.; Haigh, P.; Naoumov, N.V. Blocking of interleukin-10 receptor—A novel approach to stimulate T-helper cell type 1 responses to hepatitis C virus. Clin. Immunol. 2005, 117, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Urbani, S.; Amadei, B.; Tola, D.; Massari, M.; Schivazappa, S.; Missale, G.; Ferrari, C. PD-1 Expression in Acute Hepatitis C Virus (HCV) Infection Is Associated with HCV-Specific CD8 Exhaustion. J. Virol. 2006, 80, 11398–11403. [Google Scholar] [CrossRef] [Green Version]
- Radziewicz, H.; Ibegbu, C.C.; Fernandez, M.L.; Workowski, K.A.; Obideen, K.; Wehbi, M.; Hanson, H.L.; Steinberg, J.P.; Masopust, D.; Wherry, E.J.; et al. Liver-Infiltrating Lymphocytes in Chronic Human Hepatitis C Virus Infection Display an Exhausted Phenotype with High Levels of PD-1 and Low Levels of CD127 Expression. J. Virol. 2007, 81, 2545–2553. [Google Scholar] [CrossRef] [Green Version]
- Brooks, D.G.; Ha, S.-J.; Elsaesser, H.; Sharpe, A.H.; Freeman, G.J.; Oldstone, M.B.A. IL-10 and PD-L1 operate through distinct pathways to suppress T-cell activity during persistent viral infection. Proc. Natl. Acad. Sci. USA 2008, 105, 20428–20433. [Google Scholar] [CrossRef] [Green Version]
- Day, C.L.; Kaufmann, D.E.; Kiepiela, P.; Brown, J.A.; Moodley, E.S.; Reddy, S.; Mackey, E.W.; Miller, J.D.; Leslie, A.; DePierres, C.; et al. PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature 2006, 443, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Velu, V.; Titanji, K.; Zhu, B.; Husain, S.; Pladevega, A.; Lai, L.; Vanderford, T.H.; Chennareddi, L.; Silvestri, G.; Freeman, G.J.; et al. Enhancing SIV-specific immunity in vivo by PD-1 blockade. Nature 2008, 458, 206–210. [Google Scholar] [CrossRef]
- Díaz-Valdés, N.; Manterola, L.; Belsúe, V.; Riezu-Boj, J.-I.; Larrea, E.; Echeverria, I.; Llopiz, D.; López-Sagaseta, J.; Lerat, H.; Pawlotsky, J.; et al. Improved dendritic cell-based immunization against hepatitis C virus using peptide inhibitors of interleukin 10. Hepatology 2010, 53, 23–31. [Google Scholar] [CrossRef]
- Kantarjian, H. Acute myeloid leukemia—Major progress over four decades and glimpses into the future. Am. J. Hematol. 2015, 91, 131–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Gajewski, T.F.; Kline, J. PD-1/PD-L1 interactions inhibit antitumor immune responses in a murine acute myeloid leukemia model. Blood 2009, 114, 1545–1552. [Google Scholar] [CrossRef] [Green Version]
- Hallett, W.H.; Jing, W.; Drobyski, W.R.; Johnson, B.D. Immunosuppressive Effects of Multiple Myeloma Are Overcome by PD-L1 Blockade. Biol. Blood Marrow Transplant. 2011, 17, 1133–1145. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Munger, M.; Highfill, S.L.; Tolar, J.; Weigel, B.J.; Riddle, M.; Sharpe, A.H.; Vallera, D.A.; Azuma, M.; Levine, B.L.; et al. Program death-1 signaling and regulatory T cells collaborate to resist the function of adoptively transferred cytotoxic T lymphocytes in advanced acute myeloid leukemia. Blood 2010, 116, 2484–2493. [Google Scholar] [CrossRef]
- Salih, H.R.; Wintterle, S.; Krusch, M.; Kroner, A.; Huang, Y.-H.; Chen, L.; Wiendl, H. The role of leukemia-derived B7-H1 (PD-L1) in tumor–T-cell interactions in humans. Exp. Hematol. 2006, 34, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, S.; Wang, L.; Zhang, W.G.; Ji, Y.; Ma, X. Clinical Significance of B7-H1 (Pd-L1) Expression in Human Acute Leukemia. Cancer Biol. Ther. 2008, 7, 622–627. [Google Scholar] [CrossRef] [Green Version]
- Berthon, C.; Driss, V.; Liu, J.; Kuranda, K.; Leleu, X.; Jouy, N.; Hetuin, D.; Quesnel, B. In acute myeloid leukemia, B7-H1 (PD-L1) protection of blasts from cytotoxic T cells is induced by TLR ligands and interferon-gamma and can be reversed using MEK inhibitors. Cancer Immunol. Immunother. 2010, 59, 1839–1849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, W.; Ma, X.; Li, X.; Wang, Y.; Li, C.; Meng, H.; Liu, X.; Yu, Z.; You, S.; Qiu, L. B7-H1 up-regulation on dendritic-like leukemia cells suppresses T cell immune function through modulation of IL-10/IL-12 production and generation of Treg cells. Leuk. Res. 2009, 33, 948–957. [Google Scholar] [CrossRef]
- Annibali, O.; Crescenzi, A.; Tomarchio, V.; Pagano, A.; Bianchi, A.; Grifoni, A.; Avvisati, G. PD-1 /PD-L1 checkpoint in hematological malignancies. Leuk. Res. 2018, 67, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-F.; Zhang, Q.-T.; Xin, H.-Z.; Gan, S.-L.; Ma, J.; Liu, Y.-F.; Xie, X.-S.; Sun, H. Expression of Programmed Death Ligand-1 (PD-L1) in Human Acute Leukemia and Its Clinical Significance. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2015, 23, 930–934. [Google Scholar] [PubMed]
- Yang, H.; Bueso-Ramos, C.; Dinardo, C.D.; Estecio, M.; Davanlou, M.; Geng, Q.-R.; Fang, Z.; Nguyen, M.; Pierce, S.; Wei, Y.; et al. Expression of PD-L1, PD-L2, PD-1 and CTLA4 in myelodysplastic syndromes is enhanced by treatment with hypomethylating agents. Leukemia 2013, 28, 1280–1288. [Google Scholar] [CrossRef]
- Zajac, M.; Zaleska, J.; Dolnik, A.; Bullinger, L.; Giannopoulos, K. Expression of CD 274 (PD-L1) is associated with unfavourable recurrent mutations in AML. Br. J. Haematol. 2017, 183, 822–825. [Google Scholar] [CrossRef] [Green Version]
- Rotte, A.; Jin, J.; Lemaire, V. Mechanistic overview of immune checkpoints to support the rational design of their combinations in cancer immunotherapy. Ann. Oncol. 2017, 29, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Gray, C.P.; Arosio, P.; Hersey, P. Heavy chain ferritin activates regulatory T cells by induction of changes in dendritic cells. Blood 2002, 99, 3326–3334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francisco, L.M.; Salinas, V.; Brown, K.E.; Vanguri, V.K.; Freeman, G.J.; Kuchroo, V.K.; Sharpe, A.H. PD-L1 regulates the development, maintenance, and function of induced regulatory T cells. J. Exp. Med. 2009, 206, 3015–3029. [Google Scholar] [CrossRef]
- Ustun, C.; Miller, J.S.; Munn, D.H.; Weisdorf, D.J.; Blazar, B.R. Regulatory T Cells in Acute Myelogenous Leukemia: Is It Time for Immunomodulation? Blood 2011, 118, 5084–5095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczepanski, M.J.; Szajnik, M.; Czystowska-Kuźmicz, M.; Mandapathil, M.; Strauss, L.; Welsh, A.; Foon, K.A.; Whiteside, T.L.; Boyiadzis, M. Increased Frequency and Suppression by Regulatory T Cells in Patients with Acute Myelogenous Leukemia. Clin. Cancer Res. 2009, 15, 3325–3332. [Google Scholar] [CrossRef] [Green Version]
- Ersvaer, E.; Liseth, K.; Skavland, J.; Gjertsen, B.T.; Bruserud, Ø. Intensive Chemotherapy for Acute Myeloid Leukemia Differentially Affects Circulating Tc1, Th1, Th17 and Treg Cells. BMC Immunol. 2010, 11, 38. [Google Scholar] [CrossRef] [Green Version]
- Shenghui, Z.; Yixiang, H.; Jianbo, W.; Kang, Y.; Laixi, B.; Yan, Z.; Xi, X. Elevated frequencies of CD4+CD25+CD127lo regulatory T cells is associated to poor prognosis in patients with acute myeloid leukemia. Int. J. Cancer 2010, 129, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Tsimberidou, A.M.; Estey, E.; Wen, S.; Pierce, S.; Kantarjian, H.; Albitar, M.; Kurzrock, R. The prognostic significance of cytokine levels in newly diagnosed acute myeloid leukemia and high-risk myelodysplastic syndromes. Cancer 2008, 113, 1605–1613. [Google Scholar] [CrossRef]
- Giannopoulos, K. Targeting Immune Signaling Checkpoints in Acute Myeloid Leukemia. J. Clin. Med. 2019, 8, 236. [Google Scholar] [CrossRef] [Green Version]
- Ørskov, A.D.; Treppendahl, M.B.; Skovbo, A.; Holm, M.S.; Friis, L.S.; Hokland, M.; Grønbæk, K. Hypomethylation and up-regulation ofPD-1in T cells by azacytidine in MDS/AML patients: A rationale for combined targeting of PD-1 and DNA methylation. Oncotarget 2015, 6, 9612–9626. [Google Scholar] [CrossRef] [Green Version]
- Dinardo, C.D.; Jonas, B.A.; Pullarkat, V.; Thirman, M.J.; Garcia, J.S.; Wei, A.H.; Konopleva, M.; Döhner, H.; Letai, A.; Fenaux, P.; et al. Azacitidine and Venetoclax in Previously Untreated Acute Myeloid Leukemia. N. Engl. J. Med. 2020, 383, 617–629. [Google Scholar] [CrossRef]
- Zeidan, A.M.; Boddu, P.; Wood, B.L.; Zelterman, D.; Little, R.F.; Ivy, S.P.; Caldwell, A.; Sanchez-Espiridion, B.; Alatrash, G.; Sharon, E.; et al. Blast Mrd Aml-2: Blockade of Pd-1 Added to Standard Therapy to Target Measurable Residual Disease (Mdr) in Acute Myeloid Leukemia (Aml) 2—A Randomized Phase 2 Study of the Venetoclax, Azacitidine, and Pembrolizumab Versus Venetoclax and Azacitidine as First Line Therapy in Older Patients with Aml Who Are Ineligible or Who Refuse Intensive Chemotherapy. Blood 2020, 136, 11–12. [Google Scholar]
- Daver, N.; Garcia-Manero, G.; Basu, S.; Boddu, P.C.; Alfayez, M.; Cortes, J.E.; Konopleva, M.; Ravandi-Kashani, F.; Jabbour, E.; Kadia, T.M.; et al. Efficacy, Safety, and Biomarkers of Response to Azacitidine and Nivolumab in Relapsed/Refractory Acute Myeloid Leukemia: A Nonrandomized, Open-Label, Phase II Study. Cancer Discov. 2018, 9, 370–383. [Google Scholar] [CrossRef] [Green Version]
- Gojo, I.; Stuart, R.K.; Webster, J.; Blackford, A.; Varela, J.C.; Morrow, J.; DeZern, A.E.; Foster, M.C.; Levis, M.J.; Coombs, C.C.; et al. Multi-Center Phase 2 Study of Pembroluzimab (Pembro) and Azacitidine (AZA) in Patients with Relapsed/Refractory Acute Myeloid Leukemia (AML) and in Newly Diagnosed (≥65 Years) AML Patients. Blood 2019, 134, 832. [Google Scholar] [CrossRef]
- Zeidan, A.M.; Cavenagh, J.; Voso, M.T.; Taussig, D.; Tormo, M.; Boss, I.; Copeland, W.B.; Gray, V.E.; Previtali, A.; O’Connor, T.; et al. Efficacy and Safety of Azacitidine (AZA) in Combination with the Anti-PD-L1 Durvalumab (durva) for the Front-Line Treatment of Older Patients (pts) with Acute Myeloid Leukemia (AML) Who Are Unfit for Intensive Chemotherapy (IC) and Pts with Higher-Risk Myelodysplastic Syndromes (HR-MDS): Results from a Large, International, Randomized Phase 2 Study. Blood 2019, 134, 829. [Google Scholar]
- Zeidan, A.M.; Boddu, P.; Wood, B.L.; Zelterman, D.; Little, R.F.; Ivy, S.P.; Caldwell, A.; Sanchez-Espiridion, B.; Alatrash, G.; Sharon, E.; et al. Blast Mrd Aml-1 Trial: Blockade of Pd-1 Added to Standard Therapy to Target Measurable Residual Disease in Acute Myeloid Leukemia (Aml) 1—An Investigator-Initiated, Ctep-Sponsored, Randomized Phase 2 Study of the Anti-Pd-1 Antibody Pembrolizumab in Combination with Conventional Intensive Chemotherapy (Ic) as Frontline Therapy in Patients with Acute Myeloid Leukemia (Aml). Blood 2020, 136, 15. [Google Scholar]
- Lindblad, K.E.; Thompson, J.; Gui, G.; Valdez, J.; Worthy, T.; Tekleab, H.; Hughes, T.; Goswami, M.; Oetjen, K.; Kim, N.-Y.; et al. Pembrolizumab and Decitabine for Refractory or Relapsed Acute Myeloid Leukemia. Blood 2018, 132, 1437. [Google Scholar] [CrossRef]
- Liao, D.; Wang, M.; Liao, Y.; Li, J.; Niu, T. A Review of Efficacy and Safety of Checkpoint Inhibitor for the Treatment of Acute Myeloid Leukemia. Front. Pharmacol. 2019, 10, 609. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Cui, L.; Zhao, X.; Bai, H.; Cai, S.; Wang, G.; Zhao, Z.; Zhao, J.; Chen, S.; Song, J.; et al. Use of Immunotherapy with Programmed Cell Death 1 Vs Programmed Cell Death Ligand 1 Inhibitors in Patients with Cancer: A Systematic Review and Meta-Analysis. JAMA Oncol. 2020, 6, 375–384. [Google Scholar] [CrossRef]
- Ijaz, A.; Khan, A.Y.; Malik, S.U.; Faridi, W.; Fraz, M.A.; Usman, M.; Tariq, M.J.; Durer, S.; Russ, A.; Parr, N.N.C.; et al. Significant Risk of Graft-versus-Host Disease with Exposure to Checkpoint Inhibitors before and after Allogeneic Transplantation. Biol. Blood Marrow Transplant. 2018, 25, 94–99. [Google Scholar] [CrossRef] [Green Version]
- Albring, J.C.; Inselmann, S.; Sauer, T.; Schliemann, C.; Altvater, B.; Kailayangiri, S.; Rössig, C.; Hartmann, W.; Knorrenschild, J.R.; Sohlbach, K.; et al. PD-1 checkpoint blockade in patients with relapsed AML after allogeneic stem cell transplantation. Bone Marrow Transplant. 2016, 52, 317–320. [Google Scholar] [CrossRef]
- Nair, R.; Salinas-Illarena, A.; Baldauf, H.-M. New strategies to treat AML: Novel insights into AML survival pathways and combination therapies. Leukemia 2020, 35, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Blank, C.; Brown, I.; Peterson, A.C.; Spiotto, M.; Iwai, Y.; Honjo, T.; Gajewski, T.F. PD-L1/B7H-1 Inhibits the Effector Phase of Tumor Rejection by T Cell Receptor (TCR) Transgenic CD8+ T Cells. Cancer Res. 2004, 64, 1140–1145. [Google Scholar] [CrossRef] [Green Version]
- Jimbu, L.; Mesaros, O.; Popescu, C.; Neaga, A.; Berceanu, I.; Dima, D.; Gaman, M.; Zdrenghea, M. Is There a Place for Pd-1-Pd-L Blockade in Acute Myeloid Leukemia? Pharmaceuticals 2021, 14, 288. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, K.; Harb, W.; Lu, N.; Ma, X.; He, Y.; Yuan, L.; Fu, M.; Lin, Y.; Xu, W.; Wang, X.; et al. Phase I study of KN035, a novel fusion Anti-PD-L1 antibody administered subcutaneously in patients with advanced solid tumors in the USA. Ann. Oncol. 2018, 29, viii405. [Google Scholar] [CrossRef]
- Xu, J.-M.; Qin, S.; Zhang, Y.; Zhang, Y.; Jia, R.; Liu, R.; Zhang, G.; Zhao, C.; Lu, N.; Liu, H.; et al. Phase I study of KN035, the first subcutaneously administered, novel fusion anti-PD-L1 antibody in patients with advanced solid tumors in China. J. Clin. Oncol. 2019, 37, 2608. [Google Scholar] [CrossRef]
- D’Angelo, S.P.; Robinson, S.I.; Lam, J.; Adams, B.J.; Freddo, J.L.; Theuer, C.P.; Maki, R.G. ENVASARC: A pivotal trial of envafolimab, and envafolimab in combination with ipilimumab, in patients with advanced or metastatic undifferentiated pleomorphic sarcoma or myxofibrosarcoma who have progressed on prior chemotherapy. J. Clin. Oncol. 2021, 39, TPS11581. [Google Scholar] [CrossRef]
- Clingan, P.; Brungs, D.; Ladwa, R.; Mant, A.; McGrath, M.; Tazbirkova, A.; Koralewski, P.; Lugowska, I.; Charoentum, C.; Dechaphunkul, A.; et al. 1084P Cosibelimab, an anti-PD-L1 antibody, in metastatic cutaneous squamous cell carcinoma (mCSCC): Preliminary safety and efficacy results from a phase I clinical trial. Ann. Oncol. 2020, 31, S736. [Google Scholar] [CrossRef]
- Lin, C.C.; Taylor, M.; Boni, V.; Brunsvig, P.F.; Geater, S.L.; Salvagni, S.; Garrido Lopez, P.; Özgüroğlu, M.; Sriuranpong, V.; Ponce Aix, P.A.S.; et al. Phase I/Ii Study of Spartalizumab (Pdr001), an Anti-Pd1 Mab, in Patients with Advanced Melanoma or Non-Small Cell Lung Cancer. Ann. Oncol. 2018, 29, viii413. [Google Scholar] [CrossRef]
- Akinleye, A.; Rasool, Z. Immune checkpoint inhibitors of PD-L1 as cancer therapeutics. J. Hematol. Oncol. 2019, 12, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curran, M.; Montalvo, W.; Yagita, H.; Allison, J.P. PD-1 and CTLA-4 combination blockade expands infiltrating T cells and reduces regulatory T and myeloid cells within B16 melanoma tumors. Proc. Natl. Acad. Sci. USA 2010, 107, 4275–4280. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jimbu, L.; Mesaros, O.; Neaga, A.; Nanut, A.M.; Tomuleasa, C.; Dima, D.; Bocsan, C.; Zdrenghea, M. The Potential Advantage of Targeting Both PD-L1/PD-L2/PD-1 and IL-10–IL-10R Pathways in Acute Myeloid Leukemia. Pharmaceuticals 2021, 14, 1105. https://doi.org/10.3390/ph14111105
Jimbu L, Mesaros O, Neaga A, Nanut AM, Tomuleasa C, Dima D, Bocsan C, Zdrenghea M. The Potential Advantage of Targeting Both PD-L1/PD-L2/PD-1 and IL-10–IL-10R Pathways in Acute Myeloid Leukemia. Pharmaceuticals. 2021; 14(11):1105. https://doi.org/10.3390/ph14111105
Chicago/Turabian StyleJimbu, Laura, Oana Mesaros, Alexandra Neaga, Ana Maria Nanut, Ciprian Tomuleasa, Delia Dima, Corina Bocsan, and Mihnea Zdrenghea. 2021. "The Potential Advantage of Targeting Both PD-L1/PD-L2/PD-1 and IL-10–IL-10R Pathways in Acute Myeloid Leukemia" Pharmaceuticals 14, no. 11: 1105. https://doi.org/10.3390/ph14111105
APA StyleJimbu, L., Mesaros, O., Neaga, A., Nanut, A. M., Tomuleasa, C., Dima, D., Bocsan, C., & Zdrenghea, M. (2021). The Potential Advantage of Targeting Both PD-L1/PD-L2/PD-1 and IL-10–IL-10R Pathways in Acute Myeloid Leukemia. Pharmaceuticals, 14(11), 1105. https://doi.org/10.3390/ph14111105