
Bioactive Compounds in Aegopodium podagraria Leaf Extracts and Their Effects against Fluoride-Modulated Oxidative Stress in the THP-1 Cell Line

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Antioxidant Capacity of Goutweed Extracts Obtained Using Different Solvents
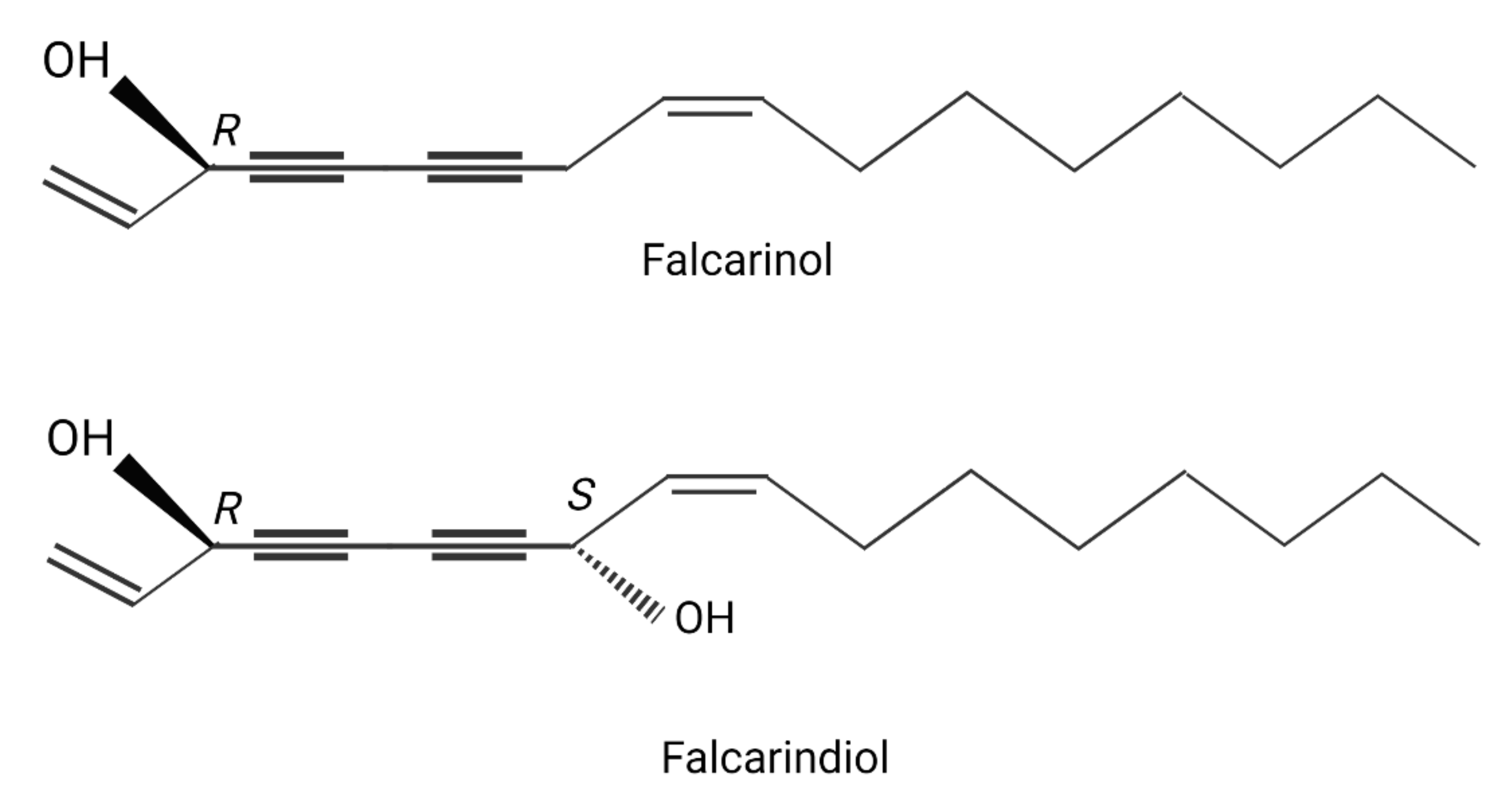
2.2. Chemical Composition of the Leaf Extracts (Polyacetylene and Essential Oil)
2.3. Cell Culture Experiments
2.3.1. Cellular SOD and CAT Activity
2.3.2. Cellular Antioxidant Capacity
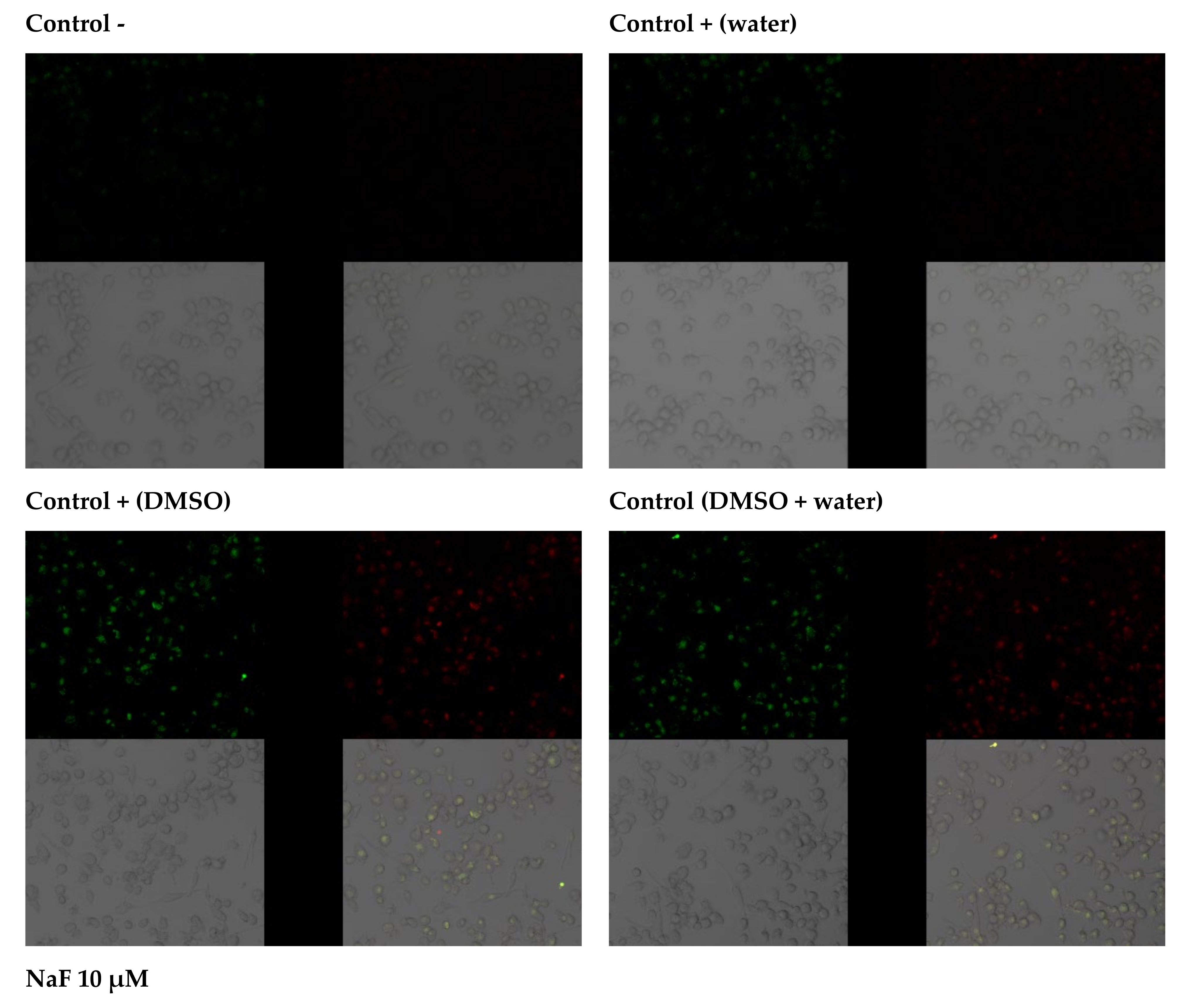
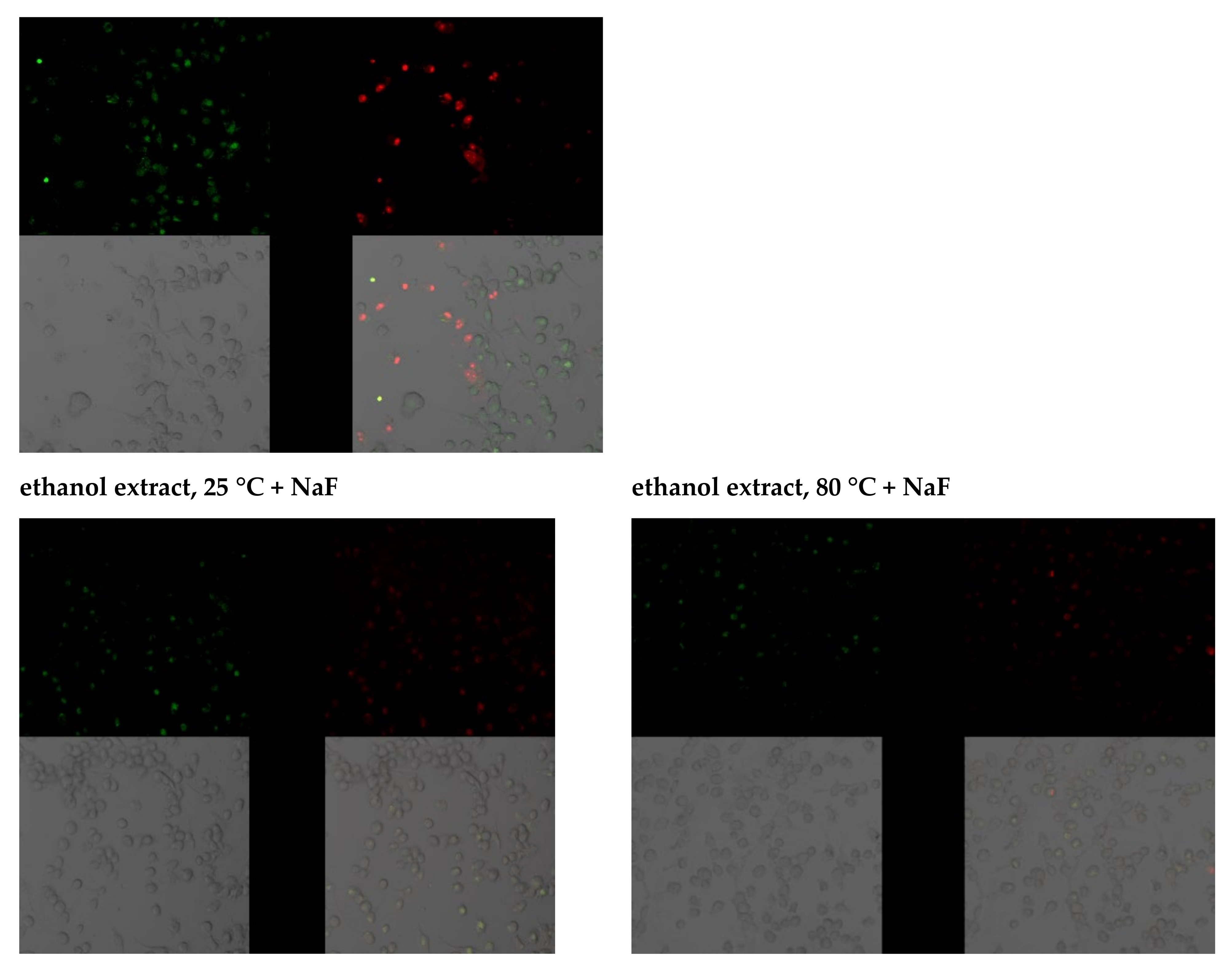
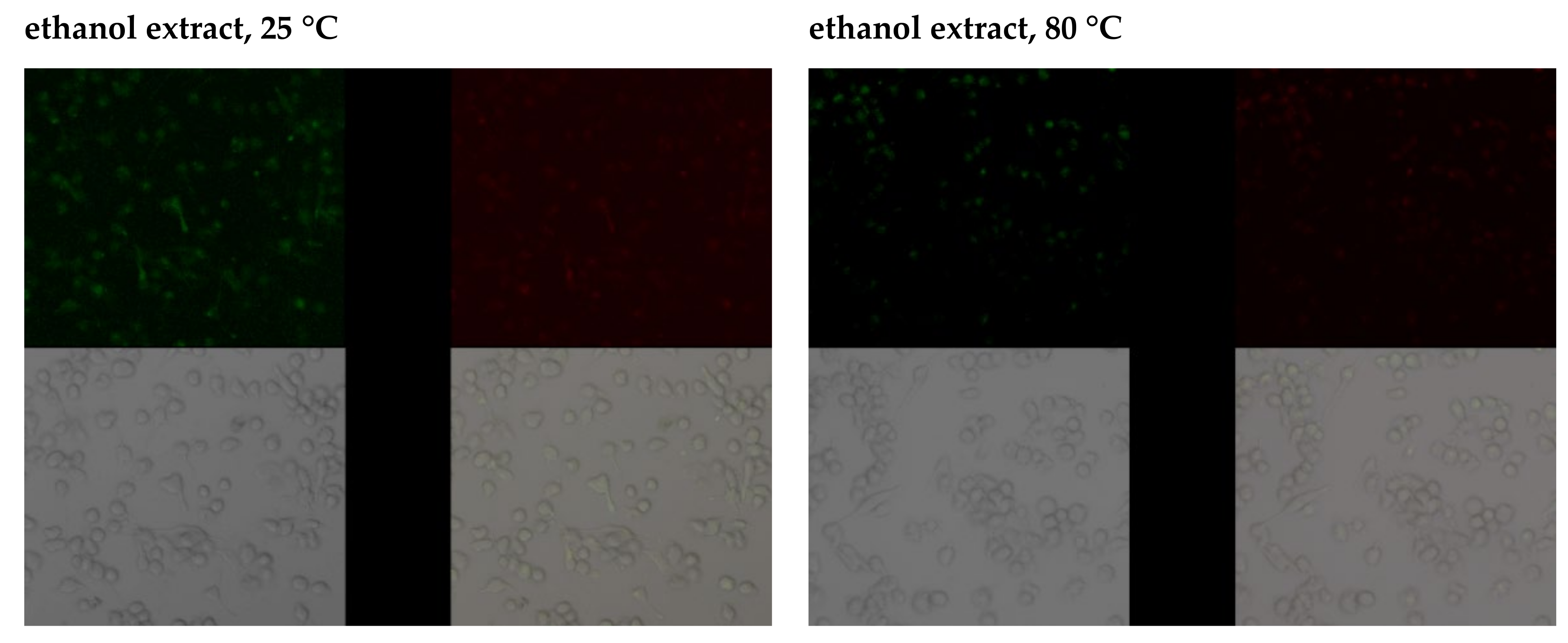
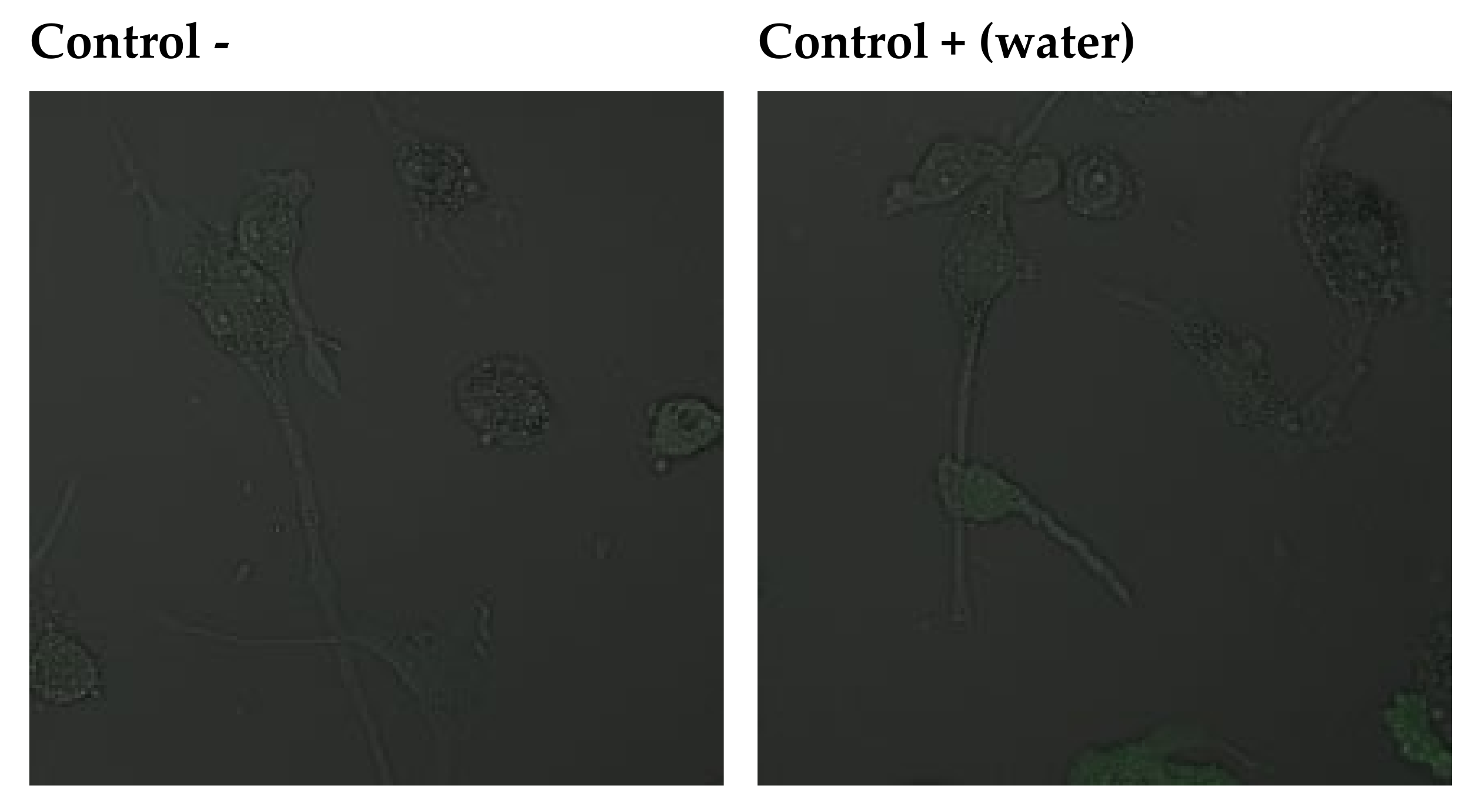
2.3.3. Anti-Apoptotic Activity of Leaf Extracts
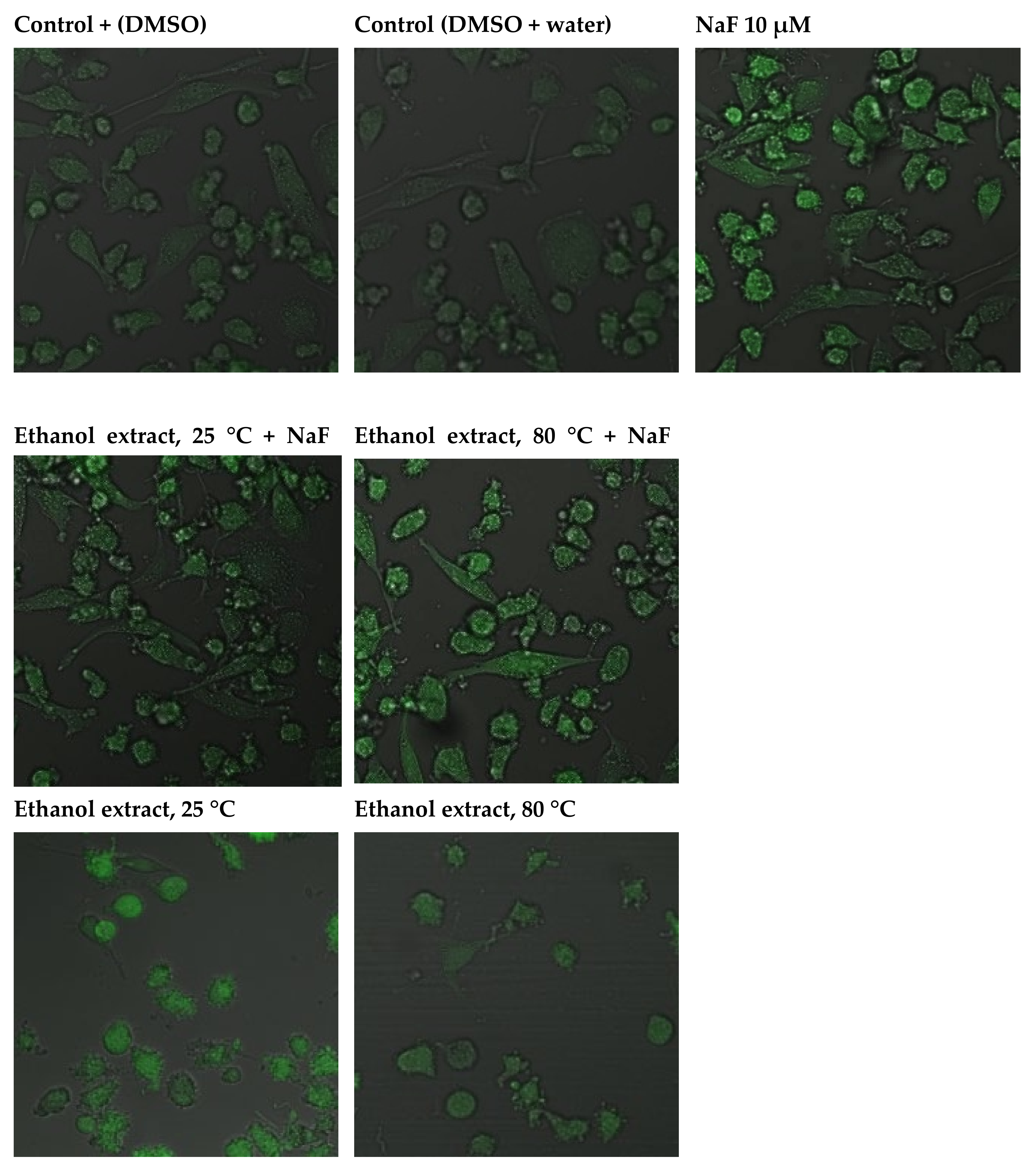
2.3.4. Capacity of Intracellular ROS Decrease
3. Discussion
3.1. Characteristics of the Source Material: Analysis of Essential Oils, Total Polyphenols and Polyacetylenes, and Antioxidant Capacity of Goutweed Extracts Obtained Using Different Solvents
3.2. Study of the Antioxidant Response of Macrophages Exposed to Goutweed Extracts
4. Materials and Methods
4.1. Plant Material
4.2. Preparation of an Aqueous Extract
4.3. Preparation of Ethanol Extract
4.4. Preparation of Extracts for Analysis
4.5. Antioxidant Activity of Extracts by the DPPH Method
4.6. Determination of the Reduction Potential of Extracts by the FRAP Method
4.7. Determination of Polyacetylenes
4.8. Determination of the Total Phenolic Content (TPC) in Extracts
4.9. Isolation of Essential Oil
4.10. Identification of Essential Oil Constituents
4.11. Analysis of Essential Oil
4.12. Cell Cultures
4.13. Measurement of the Total Antioxidant Potential (ABTS) and the Activity of Antioxidant Enzymes
4.14. Imaging of Intracellular ROS Generation
4.15. Imaging of Cytoplasmic ROS Generation
4.16. In Vitro Imaging of Apoptosis
4.17. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stefanovic, O.; Comic, L.; Stanojevic, D.; Solujic-Sukdolak, S. Antibacterial Activity of Aegopodium podagraria L. Extracts and Interaction Between Extracts and Antibiotics. Turk. J. Biol. 2009, 33, 145–150. [Google Scholar]
- Jakubczyk, K.; Kwiatkowski, P.; Sienkiewicz, M.; Janda, K. The content of polyphenols in extract from goutweed (Aegopodium podagraria L.) and their antistaphylococcal activity. Postępy Fitoter. 2018, 19, 3–9. [Google Scholar]
- Jakubczyk, K.; Janda, K.; Styburski, D.; Łukomska, A. Goutweed (Aegopodium podagraria L.)—Botanical Characteristics and Prohealthy Properties. Postepy Hig. Med. Dosw. 2020, 74, 28–35. [Google Scholar] [CrossRef]
- Christensen, L.P.; Brandt, K. Bioactive Polyacetylenes in Food Plants of the Apiaceae Family: Occurrence, Bioactivity and Analysis. J. Pharm. Biomed. Anal. 2006, 41, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Orav, A.; Viitak, A.; Vaher, M. Identification of Bioactive Compounds in the Leaves and Stems of Aegopodium podagraria by Various Analytical Techniques. Procedia Chem. 2010, 2, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Young, J.F.; Christensen, L.P.; Theil, P.K.; Oksbjerg, N. The polyacetylenes falcarinol and falcarindiol affect stress responses in myotube cultures in a biphasic manner. Dose Response 2008, 6, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Prior, R.M.; Lundgaard, N.H.; Light, M.E.; Stafford, G.I.; van Staden, J.; Jäger, A.K. The Polyacetylene Falcarindiol with COX-1 Activity Isolated from Aegopodium podagraria L. J. Ethnopharmacol. 2007, 113, 176–178. [Google Scholar] [CrossRef]
- Castaneda, O.A.; Lee, S.-C.; Ho, C.-T.; Huang, T.-C. Macrophages in Oxidative Stress and Models to Evaluate the Antioxidant Function of Dietary Natural Compounds. J. Food Drug Anal. 2017, 25, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Pogozhikh, N.I.; Tovchiga, O.V.; Evlash, V.V.; Stepanova, S.I.; Koyro, O.O. Substantiation of the Rational Drying Conditions for the Herbal Raw Material of Goutweed (Aegopodium podagraria L.) Aerial Part. South Asian Res. J. Nat. Prod. 2018, 1, 1–11. [Google Scholar]
- Wróblewska, A.; Janda, K.; Makuch, E.; Walasek, M.; Miądlicki, P.; Jakubczyk, K. Effect of Extraction Method on the Antioxidative Activity of Ground Elder (Aegopodium podagraria L.). Pol. J. Chem. Technol. 2019, 21, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Ozkan, G.; Kamiloglu, S.; Ozdal, T.; Boyacioglu, D.; Capanoglu, E. Potential Use of Turkish Medicinal Plants in the Treatment of Various Diseases. Molecules 2016, 21, 257. [Google Scholar] [CrossRef]
- Kapetanos, C.; Karioti, A.; Bojović, S.; Marin, P.; Veljić, M.; Skaltsa, H. Chemical and Principal-Component Analyses of the Essential Oils of Apioideae Taxa (Apiaceae) from Central Balkan. Chem. Biodivers. 2008, 5, 101–119. [Google Scholar] [CrossRef]
- Szczuko, M.; Zapalowska-Chwyć, M.; Drozd, R. A Low Glycemic Index Decreases Inflammation by Increasing the Concentration of Uric Acid and the Activity of Glutathione Peroxidase (GPx3) in Patients with Polycystic Ovary Syndrome (PCOS). Molecules 2019, 24, 1508. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, K. Natural Compounds Involved in Adipose Tissue Mass Control in in Vitro Studies. Postepy Hig. Med. Dosw. 2011, 65, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Tomac, I.; Šeruga, M.; Labuda, J. Evaluation of Antioxidant Activity of Chlorogenic Acids and Coffee Extracts by an Electrochemical DNA-Based Biosensor. Food Chem. 2020, 325, 126787. [Google Scholar] [CrossRef] [PubMed]
- Jakubczyk, K.; Łukomska, A.; Gutowska, I.; Kochman, J.; Janił, J.; Janda, K. Edible Flowers Extracts as a Source of Bioactive Compounds with Antioxidant Properties—In Vitro Studies. Appl. Sci. 2021, 11, 2120. [Google Scholar] [CrossRef]
- Augšpole, I.; Duma, M.; Ozola, B.; Cinkmanis, I. Phenolic Profile of Fresh and Frozen Nettle, Goutweed, Dandelion and Chickweed Leaves. FOODBALT 2017 2017, 36–39. [Google Scholar] [CrossRef]
- Zofia, N.-Ł.; Martyna, Z.-D.; Aleksandra, Z.; Tomasz, B. Comparison of the Antiaging and Protective Properties of Plants from the Apiaceae Family. Oxid. Med. Cell. Longev. 2020, 2020, 5307614. [Google Scholar] [CrossRef] [PubMed]
- Petrović, G.M.; Stamenković, J.G.; Kostevski, I.R.; Stojanović, G.S.; Mitić, V.D.; Zlatković, B.K. Chemical Composition of Volatiles; Antimicrobial, Antioxidant and Cholinesterase Inhibitory Activity of Chaerophyllum Aromaticum L. (Apiaceae) Essential Oils and Extracts. Chem. Biodivers. 2017, 14, e1600367. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, H.; Karami, A.; Maggi, F. Essential Oil Composition, Total Phenolic and Flavonoids Contents, and Antioxidant Activity of Oliveria Decumbens Vent. (Apiaceae) at Different Phenological Stages. J. Clean. Prod. 2018, 198, 91–95. [Google Scholar] [CrossRef]
- Naima, B.; Abdelkrim, R.; Ouarda, B.; Salah, N.N.; Larbi, B.A.M. Chemical Composition, Antimicrobial, Antioxidant and Anticancer Activities of Essential Oil from Ammodaucus Leucotrichus Cosson & Durieu (Apiaceae) Growing in South Algeria. Bull. Chem. Soc. Ethiop. 2019, 33, 541–549. [Google Scholar] [CrossRef]
- Aloulou, A.; Hamden, K.; Elloumi, D.; Ali, M.B.; Hargafi, K.; Jaouadi, B.; Ayadi, F.; Elfeki, A.; Ammar, E. Hypoglycemic and Antilipidemic Properties of Kombucha Tea in Alloxan-Induced Diabetic Rats. BMC Complement. Altern. Med. 2012, 12, 63. [Google Scholar] [CrossRef] [Green Version]
- Do Nascimento, K.F.; Moreira, F.M.F.; Alencar Santos, J.; Kassuya, C.A.L.; Croda, J.H.R.; Cardoso, C.A.L.; Vieira, M.D.C.; Góis Ruiz, A.L.T.; Ann Foglio, M.; de Carvalho, J.E.; et al. Antioxidant, Anti-Inflammatory, Antiproliferative and Antimycobacterial Activities of the Essential Oil of Psidium Guineense Sw. and Spathulenol. J. Ethnopharmacol. 2018, 210, 351–358. [Google Scholar] [CrossRef]
- Paramonov, E.A.; Khalilova, A.Z.; Odinokov, V.N.; Khalilov, L.M. Identification and Biological Activity of Volatile Organic Compounds Isolated from Plants and Insects. III. Chromatography-Mass Spectrometry of Volatile Compounds of Aegopodium podagraria. Chem. Nat. Compd. 2000, 36, 584–586. [Google Scholar] [CrossRef]
- Valyova, M.; Tashev, A.; Stoyanov, S.; Yordanova, G. In Vitro Free-Radical Scavenging Activity of Aegopodium podagraria L. and Orlaya Grandiflora (L.) Hoffm. (Apiaceae). J. Chem. Technol. Metall. 2016, 51, 271–274. [Google Scholar]
- Flieger, J.; Flieger, M. The [DPPH●/DPPH-H]-HPLC-DAD Method on Tracking the Antioxidant Activity of Pure Antioxidants and Goutweed (Aegopodium podagraria L.) Hydroalcoholic Extracts. Molecules 2020, 25, 6005. [Google Scholar] [CrossRef]
- Mao, L.C.; Pan, X.; Que, F.; Fang, X.H. Antioxidant Properties of Water and Ethanol Extracts from Hot Air-Dried and Freeze-Dried Daylily Flowers. Eur. Food Res. Technol. Z. Lebensm.-Unters. -Forsch. A 2006, 222, 236–241. [Google Scholar] [CrossRef]
- Spigno, G.; De Faveri, D.M. Antioxidants from Grape Stalks and Marc: Influence of Extraction Procedure on Yield, Purity and Antioxidant Power of the Extracts. J. Food Eng. 2007, 78, 793–801. [Google Scholar] [CrossRef]
- Sultana, B.; Anwar, F.; Ashraf, M. Effect of Extraction Solvent/Technique on the Antioxidant Activity of Selected Medicinal Plant Extracts. Molecules 2009, 14, 2167–2180. [Google Scholar] [CrossRef] [PubMed]
- Azmir, J.; Zaidul, I.S.M.; Rahman, M.M.; Sharif, K.M.; Mohamed, A.; Sahena, F.; Jahurul, M.H.A.; Ghafoor, K.; Norulaini, N.A.N.; Omar, A.K.M. Techniques for Extraction of Bioactive Compounds from Plant Materials: A Review. J. Food Eng. 2013, 117, 426–436. [Google Scholar] [CrossRef]
- Shabir, G.; Anwar, F.; Sultana, B.; Khalid, Z.M.; Afzal, M.; Khan, Q.M.; Ashrafuzzaman, M. Antioxidant and Antimicrobial Attributes and Phenolics of Different Solvent Extracts from Leaves, Flowers and Bark of Gold Mohar [Delonix Regia (Bojer Ex Hook.) Raf.]. Molecules 2011, 16, 7302–7319. [Google Scholar] [CrossRef]
- Jakubczyk, K.; Łukomska, A.; Baranowska-Bosiacka, I.; Goschorska, M.; Dec, K.; Wolska, J.; Janda, K.; Piotrowska, K.; Kupnicka, P.; Kupczak, P.; et al. The influence of extracts from the seeds of the common nettle (urtica dioica L.) on the activity of antioxidative enzymes in macrophages incubated with sodium fluoride. Fluoride 2018, 51, 65–76. [Google Scholar]
- Barbier, O.; Arreola-Mendoza, L.; Del Razo, L.M. Molecular Mechanisms of Fluoride Toxicity. Chem. Biol. Interact. 2010, 188, 319–333. [Google Scholar] [CrossRef] [PubMed]
- Miranda, G.H.N.; Gomes, B.A.Q.; Bittencourt, L.O.; Aragão, W.A.B.; Nogueira, L.S.; Dionizio, A.S.; Buzalaf, M.A.R.; Monteiro, M.C.; Lima, R.R. Chronic Exposure to Sodium Fluoride Triggers Oxidative Biochemistry Misbalance in Mice: Effects on Peripheral Blood Circulation. Oxid. Med. Cell. Longev. 2018, 2018, e8379123. [Google Scholar] [CrossRef]
- Gutowska, I.; Baranowska-Bosiacka, I.; Goschorska, M.; Kolasa, A.; Łukomska, A.; Jakubczyk, K.; Dec, K.; Chlubek, D. Fluoride as a Factor Initiating and Potentiating Inflammation in THP1 Differentiated Monocytes/Macrophages. Toxicol. In Vitro 2015, 29, 1661–1668. [Google Scholar] [CrossRef]
- Cevallos, A.M.; Herrera, J.; López-Villaseñor, I.; Hernández, R. Differential Effects of Two Widely Used Solvents, DMSO and Ethanol, on the Growth and Recovery of Trypanosoma Cruzi Epimastigotes in Culture. Korean J. Parasitol. 2017, 55, 81–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verheijen, M.; Lienhard, M.; Schrooders, Y.; Clayton, O.; Nudischer, R.; Boerno, S.; Timmermann, B.; Selevsek, N.; Schlapbach, R.; Gmuender, H.; et al. DMSO Induces Drastic Changes in Human Cellular Processes and Epigenetic Landscape in Vitro. Sci. Rep. 2019, 9, 4641. [Google Scholar] [CrossRef] [Green Version]
- Tunçer, S.; Gurbanov, R.; Sheraj, I.; Solel, E.; Esenturk, O.; Banerjee, S. Low Dose Dimethyl Sulfoxide Driven Gross Molecular Changes Have the Potential to Interfere with Various Cellular Processes. Sci. Rep. 2018, 8, 14828. [Google Scholar] [CrossRef] [PubMed]
- Galvao, J.; Davis, B.; Tilley, M.; Normando, E.; Duchen, M.R.; Cordeiro, M.F. Unexpected Low-Dose Toxicity of the Universal Solvent DMSO. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2014, 28, 1317–1330. [Google Scholar] [CrossRef]
- Mannan, A.; Liu, C.; Arsenault, P.R.; Towler, M.J.; Vail, D.R.; Lorence, A.; Weathers, P.J. DMSO Triggers the Generation of ROS Leading to an Increase in Artemisinin and Dihydroartemisinic Acid in Artemisia Annua Shoot Cultures. Plant Cell Rep. 2010, 29, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Pekkarinen, S.S.; Stöckmann, H.; Schwarz, K.; Heinonen, I.M.; Hopia, A.I. Antioxidant Activity and Partitioning of Phenolic Acids in Bulk and Emulsified Methyl Linoleate. J. Agric. Food Chem. 1999, 47, 3036–3043. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of Antioxidant Power: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Benzi, F.; Szeto, Y. Total Antioxidant Capacity of Teas by the Ferric Reducing/Antioxidant Power Assay. J. Agric. Food Chem. 1999, 47, 633–636. Available online: https://pubs.acs.org/doi/10.1021/jf9807768 (accessed on 5 July 2019). [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extraction | Solvent | DPPH | FRAP | TPC | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| [%] | [uM Fe(II)/L] | [mg/L] Gallic Acid (GAE)/[g/100 g DW] | ||||||||
| water 25 °Cz | water | 9.65 | ± | 1.29 *y,x,v,f,h | 177.79 | ± | 32.31 *y,x,w | 5.66 | ± | 0.45 *y,x,w,v,f,h |
| water 90 °Cy | water | 6.54 | ± | 0.96 * z,x,w,v,u,h,f | 331.03 | ± | 46.45 *z | 12.85 | ± | 0.22 *z,w,u |
| ethanol 25 °Cx | ethanol | 13.92 | ± | 1.07 *z,y,w,u,f | 333.55 | ± | 123.55 *z | 13.73 | ± | 0.38 *z,u |
| ethanol 80 °Cw | ethanol | 10.83 | ± | 1.40 *y,x,v,f,h | 416.42 | ± | 36.85 *z,v,u,f,h | 17.79 | ± | 1.02 *z,y,u,h |
| water 25 °Cv | DMSO | 9.71 | ± | 0.36 *z,y,w,u | 151.98 | ± | 14.50 *w | 4.98 | ± | 0.46 *z,u |
| water 90 °Cu | DMSO | 15.11 | ± | 0.46 *y,w,v,f,h | 259.68 | ± | 14.22 *w | 11.03 | ± | 0.49 *y,x,w,u,f,h |
| ethanol 25 °Cf | DMSO | 14.42 | ± | 2.10 *z,y,x,w,u | 293.91 | ± | 96.06 *w | 13.83 | ± | 3.81 *z,u |
| ethanol 80 °Ch | DMSO | 15.65 | ± | 0.55 *z,y,w,u | 302.86 | ± | 44.66 *w | 13.96 | ± | 0.23 *z,w,u |
| Extract | Falcarinol mg/100g | Falcarindiol mg/100g |
|---|---|---|
| ethanol 25 °C f | nd | 185.81 ± 19.21 *h |
| ethanol 80 °C h | 46.91 ± 3.47 *f | 474.51 ± 75.74 *f |
| Compound | Amount [%] | RI exp. * |
|---|---|---|
| n-Hexanal | 0.4 | 772 |
| n-Heptanal | 1.1 | 876 |
| Benzaldehyde + α-Pinene | 0.4 | 927 |
| Octane-2,3-dione | 0.1 | 959 |
| 6-Methylhept-5-ene-2-one | 0.8 | 961 |
| β-Pinene | 3.0 | 967 |
| 2-Pentylfuran | 0.5 | 976 |
| n-Octanal | 4.1 | 980 |
| 2-Phenylethanal | 0.2 | 1006 |
| m-Cymene | 1.4 | 1010 |
| Limonene | 0.4 | 1019 |
| (E)-Ocimene | 0.1 | 1025 |
| (E)-Oct-2-enal | 0.2 | 1031 |
| 3-Ethyl-2-methylhexa-1,3-diene | 0.1 | 1043 |
| γ-Terpinen | 0.4 | 1048 |
| cis-Linalol oxide (furanoid) | 0.2 | 1057 |
| (E,E)-Octa-3,5-dien-2-one | 0.3 | 1065 |
| Nonan-2-one | 1.1 | 1070 |
| n-Heptanoic acid | 0.1 | 1075 |
| (E)-6-Methylhepta-3,5-dien-2-one | 0.1 | 1078 |
| n-Nonanal | 0.8 | 1082 |
| Linalool | 1.8 | 1085 |
| Nopinone | 0.4 | 1106 |
| Non-3-en-2-one | 0.1 | 1116 |
| trans-Pinocarveol | 0.9 | 1123 |
| (E,Z)-Nona-2,6-dienal | 0.1 | 1126 |
| cis-Verbenol | 0.3 | 1128 |
| (E)-Non-2-en-1-al | 0.5 | 1135 |
| α-Pinocarvone | 0.3 | 1138 |
| trans-Verbenol | 0.1 | 1146 |
| endo Borneol | 0.2 | 1149 |
| p-Cymen-8-ol | 0.3 | 1160 |
| Terpinen-4-ol | 0.8 | 1162 |
| Myrtenal | 0.3 | 1169 |
| α-Terpineol | 0.4 | 1172 |
| Myrtenol | 0.8 | 1179 |
| Caprylicacid | 1.2 | 1184 |
| 2,6,6-Trimethylcyclohex-1-ene-1-carbaldehyde | 0.1 | 1194 |
| trans-Carveol | 0.1 | 1196 |
| Thymol methylether | 0.1 | 1209 |
| p-Cumical dehyde | 0.1 | 1211 |
| (Z)-Hex-3-enyl isovalerate | 0.1 | 1218 |
| Geraniol | 0.2 | 1236 |
| (E)-Dec-2-en-1-al | 1.0 | 1238 |
| p-Menth-4(8)-en-9-ol | 0.1 | 1249 |
| Pelargonic acid | 0.9 | 1268 |
| Carvacrol | 0.5 | 1277 |
| Dihydroedulan I | 1.3 | 1282 |
| Dihydroedulan II | 1.2 | 1286 |
| (E,E)-Deca-2,4-dienal | 0.1 | 1289 |
| Tridecane | 0.7 | 1300 |
| α-Cubebene | 0.1 | 1349 |
| Non-2-enoic acid | 0.6 | 1353 |
| Capric acid | 0.5 | 1365 |
| α-Cubebene | 0.2 | 1373 |
| α-Ylangene | 0.2 | 1377 |
| β-Elemene | 1.6 | 1389 |
| (E)-Dec-2-enoic acid | 0.3 | 1397 |
| (E)-α-Ionone | 0.1 | 1407 |
| (E-β-Caryophyllene | 0.7 | 1419 |
| β-copaene | 0.3 | 1429 |
| trans-α-Bergamotene | 0.4 | 1434 |
| (Z)-β-Farnesene | 0.3 | 1437 |
| (E)-β-Farnesene | 0.4 | 1447 |
| α-Himachalene | 0.2 | 1450 |
| α-Humulene | 0.4 | 1453 |
| 5,6-Epoxy-β-ionone | 0.7 | 1463 |
| β-Ionone | 1.0 | 1466 |
| α-Curcumene + γ-Muurolene | 0.5 | 1472 |
| Germacrene D | 1.0 | 1479 |
| (3Z,6E)-α-Farnesene | 0.3 | 1482 |
| α-Selinene | 0.3 | 1484 |
| γ-Muurolene | 0.2 | 1490 |
| 2,4-Ditert-butylphenol | 1.3 | 1493 |
| β-Bisabolene | 1.8 | 1503 |
| γ-Cadinene | 0.2 | 1508 |
| δ-Cadinene | 0.7 | 1516 |
| α-Calacorene | 0.4 | 1531 |
| Isoaromadendreneepoxide | 1.1 | 1545 |
| cis-Sesquisabinenhydrate | 0.2 | 1553 |
| 1,5-Epoxysalvial-4(14)-ene | 0.4 | 1559 |
| Spathulenol | 15.8 | 1576 |
| β-Caryophylleneepoxide | 3.6 | 1578 |
| Salvial-4(14)-en-1-one | 1.8 | 1585 |
| Humuleneepoxide II | 3.5 | 1601 |
| Aristoleneepoxide | 0.4 | 1604 |
| Isopropyllaurate | 0.3 | 1614 |
| 7-Hydroxyfarnesen | 0.8 | 1616 |
| Widdrol | 0.7 | 1630 |
| trans-Longipinocarveol | 0.5 | 1640 |
| α-Cadinol | 0.4 | 1643 |
| Ledeneoxide II | 1.8 | 1659 |
| Khusinol | 2.0 | 1673 |
| 2-Ethylhexyl benzoate | 0.4 | 1768 |
| Myristoleic acid | 0.2 | 1797 |
| diIisobutylphtalate (artifact) | 0.1 | 1827 |
| Heptadecan-2-one | 2.4 | 1832 |
| Decan-2-yl benzoate | 0.2 | 1834 |
| Methylpalmitate | 0.5 | 1908 |
| di-Butyl phthalate (artifact) | 0.2 | 1918 |
| Palmitic acid | 2.3 | 1960 |
| Phytol | 1.1 | 2101 |
| Tricosane | 0.2 | 2298 |
| Pentacosane | 0.1 | 2499 |
| Sum of identified | 87.0 |
| Sample | ABTS | SOD | CAT | ||||||
|---|---|---|---|---|---|---|---|---|---|
| [mM] | [U/mL] | [nmol/min/mL/Protein] | |||||||
| Negative control a | 14.7 | ± | 0.2 *b,c,d,f,g,h,i | 31.4 | ± | 6.4 *c,d,e,h,i | 51.3 | ± | 8.1 *b,c,d |
| Water (NaF solvent) b | 36.7 | ± | 7.8 *a,c,f,g,h,i | 30.1 | ± | 3.1 *c,d,e,h,i | 138.7 | ± | 5.0 *a,d,e,f |
| DMSO (extract solvent) c | 67.0 | ± | 6.6 *a,b,d,e,f,g,h | 11.4 | ± | 2.4 *a,b,f,g | 203.5 | ± | 30.2 *a,e,f,g,h,i |
| DMSO + water (extract and NaF solvent) d | 49.7 | ± | 5.0 * a,c,e,f,g,h,i | 12.3 | ± | 1.2 *a,b,f,g | 219.4 | ± | 55.5 *a,b,e,f,g,h,i |
| NaF 10 µM e | 13.8 | ± | 3.2 *b,c,d,f,g,h,i | 8.7 | ± | 1.5 *a,b,f,g | 46.3 | ± | 9.3 *b,c,d |
| ethanol extract 25 °C f | 113.0 | ± | 8.2 *a,b,c,d,e,g,i | 44.7 | ± | 12.4 *c,d,e,h,i | 491.0 | ± | 12.8 *a,b,c,d,e,g,h,i |
| ethanol extract 25 °C + NaF g | 84.4 | ± | 9.5 *a,b,c,d,e,f,h,i | 37.8 | ± | 10.9 *,d,e,h,i | 245.2 | ± | 22.0 *a,b,e,f,h,i |
| ethanol extract 80 °C h | 99.3 | ± | 3.1 *a,b,c,d,e,g,i | 10.0 | ± | 2.6 *f,g | 272.1 | ± | 9.6 *a,b,e,f,g,i |
| ethanol extract 80 °C + NaF i | 69.4 | ± | 5.1 *a,b,d,e,f,g,h | 12.0 | ± | 2.8 *a,b,f,g | 140.1 | ± | 31.9 *a,d,e,f |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakubczyk, K.; Łukomska, A.; Czaplicki, S.; Wajs-Bonikowska, A.; Gutowska, I.; Czapla, N.; Tańska, M.; Janda-Milczarek, K. Bioactive Compounds in Aegopodium podagraria Leaf Extracts and Their Effects against Fluoride-Modulated Oxidative Stress in the THP-1 Cell Line. Pharmaceuticals 2021, 14, 1334. https://doi.org/10.3390/ph14121334
Jakubczyk K, Łukomska A, Czaplicki S, Wajs-Bonikowska A, Gutowska I, Czapla N, Tańska M, Janda-Milczarek K. Bioactive Compounds in Aegopodium podagraria Leaf Extracts and Their Effects against Fluoride-Modulated Oxidative Stress in the THP-1 Cell Line. Pharmaceuticals. 2021; 14(12):1334. https://doi.org/10.3390/ph14121334
Chicago/Turabian StyleJakubczyk, Karolina, Agnieszka Łukomska, Sylwester Czaplicki, Anna Wajs-Bonikowska, Izabela Gutowska, Norbert Czapla, Małgorzata Tańska, and Katarzyna Janda-Milczarek. 2021. "Bioactive Compounds in Aegopodium podagraria Leaf Extracts and Their Effects against Fluoride-Modulated Oxidative Stress in the THP-1 Cell Line" Pharmaceuticals 14, no. 12: 1334. https://doi.org/10.3390/ph14121334
APA StyleJakubczyk, K., Łukomska, A., Czaplicki, S., Wajs-Bonikowska, A., Gutowska, I., Czapla, N., Tańska, M., & Janda-Milczarek, K. (2021). Bioactive Compounds in Aegopodium podagraria Leaf Extracts and Their Effects against Fluoride-Modulated Oxidative Stress in the THP-1 Cell Line. Pharmaceuticals, 14(12), 1334. https://doi.org/10.3390/ph14121334