
In Vitro Effects of Low Doses of β-Caryophyllene, Ascorbic Acid and d-Glucosamine on Human Chondrocyte Viability and Inflammation


 ,
,  ,
,  and
and
Abstract
:
1. Introduction
2. Results
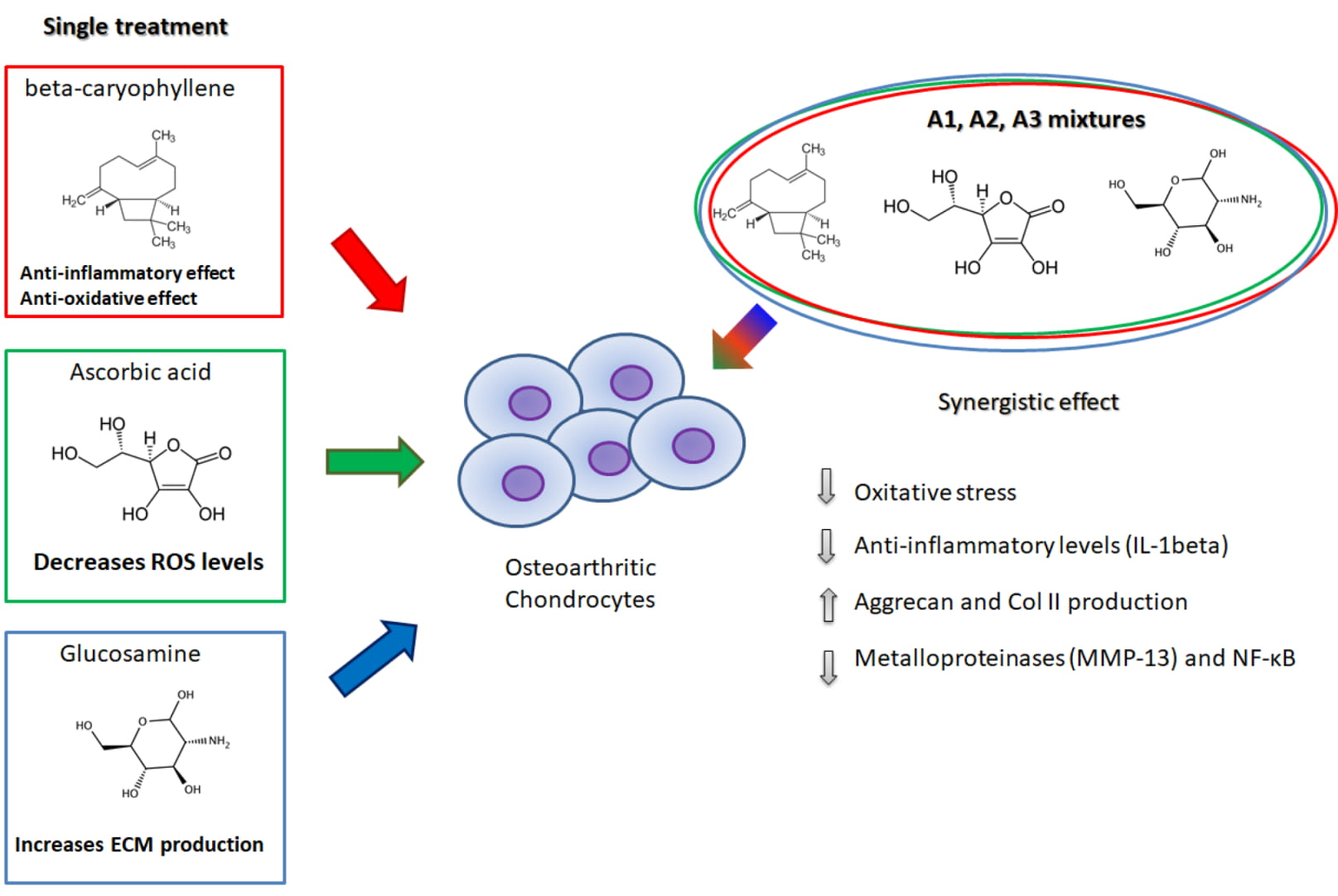
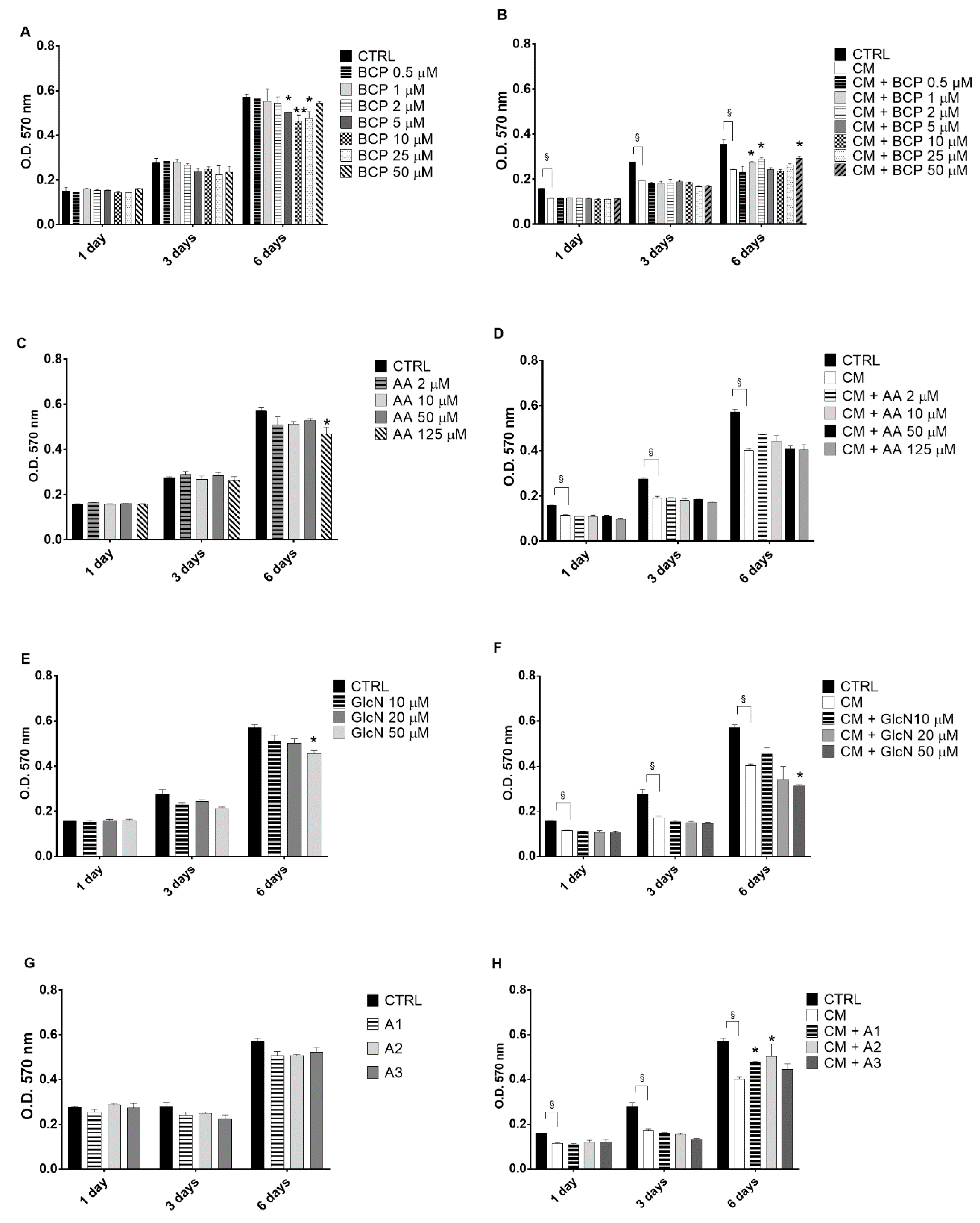
2.1. The Evaluation of the Effects of BCP, Alone or in Combination with AA and GlcN on CM-Induced Chondrocyte Toxicity
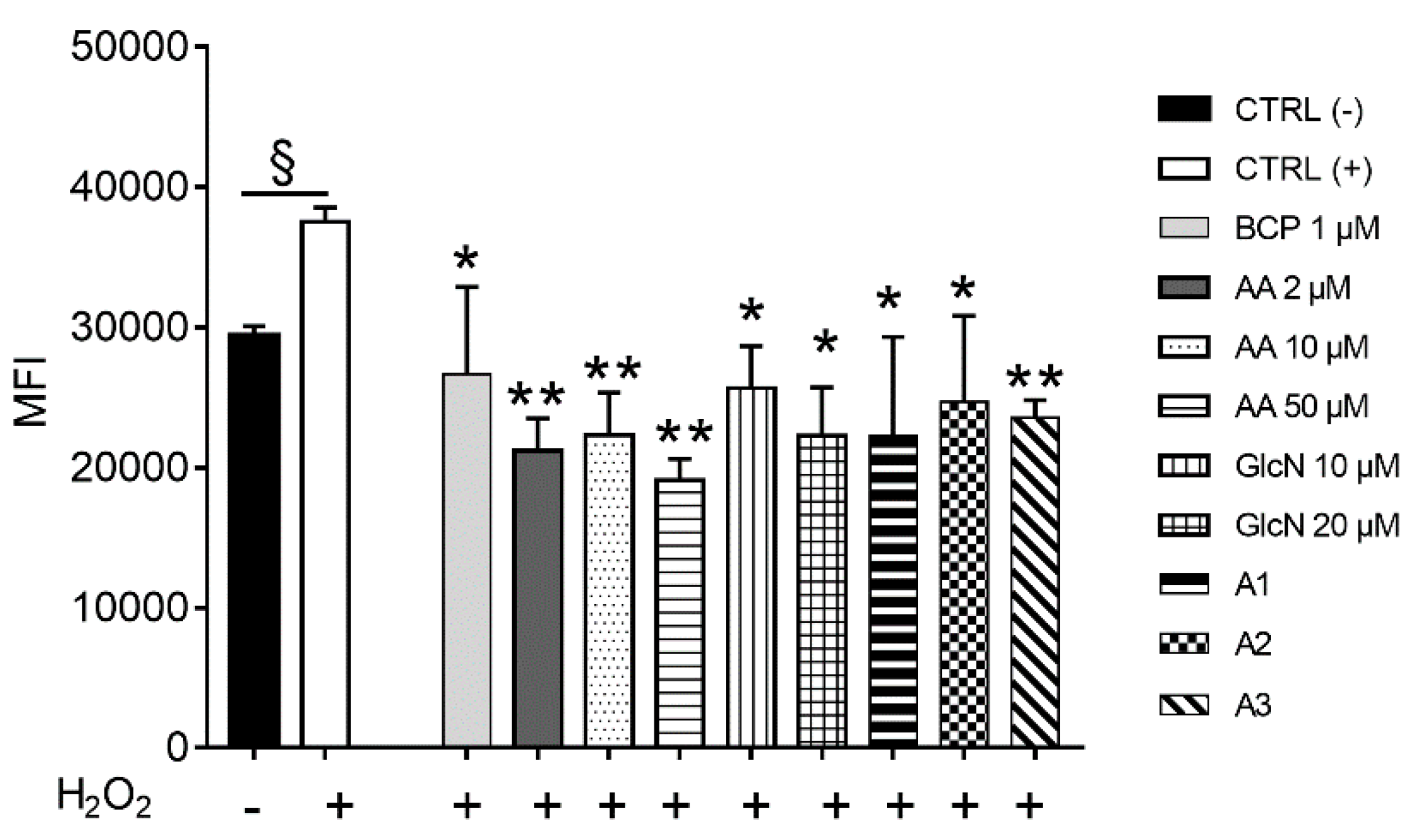
2.2. BCP, AA and d-Glucosamine Mixture Induced a Reduction of ROS Formation in Human Chondrocytes Exposed to H2O2
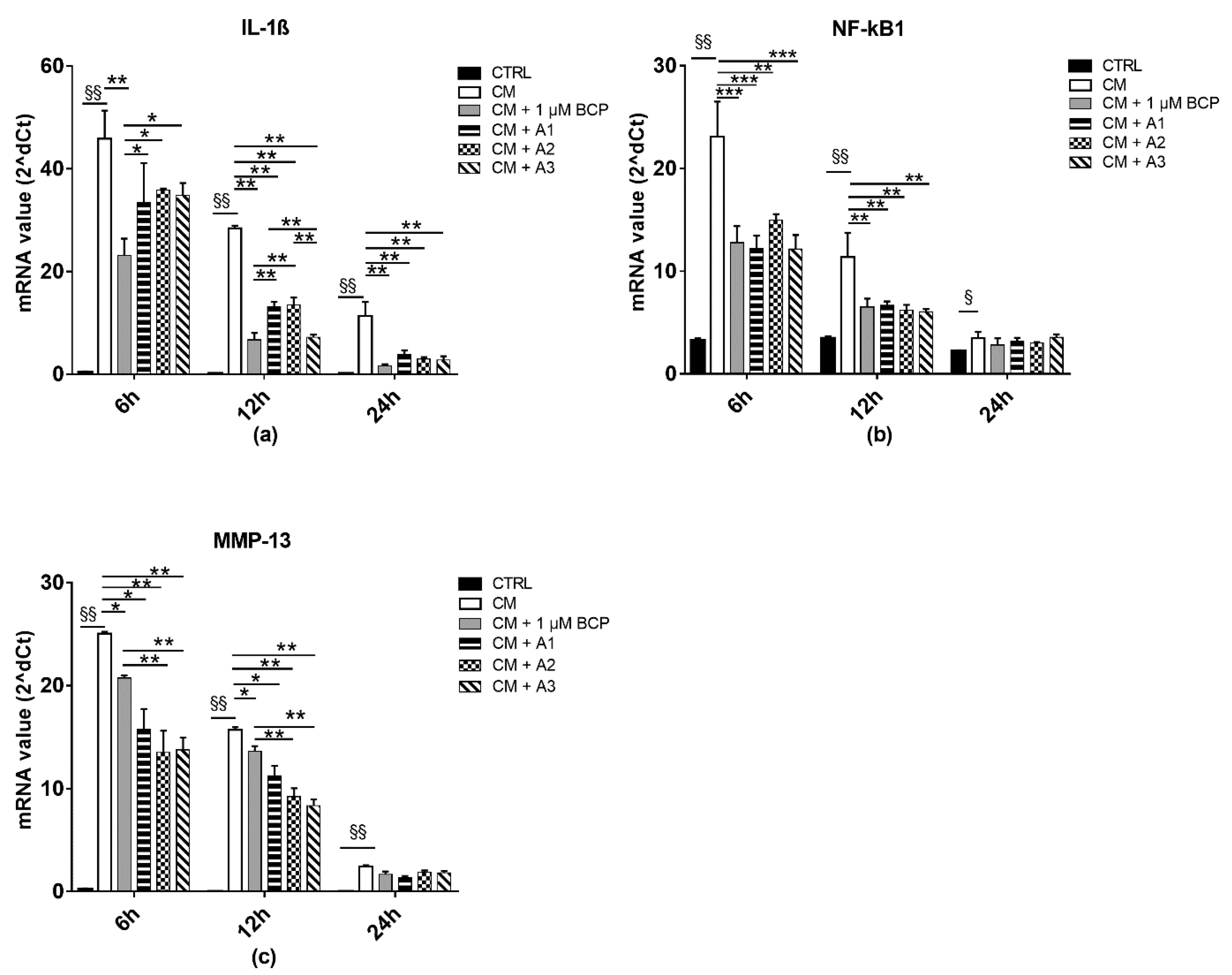
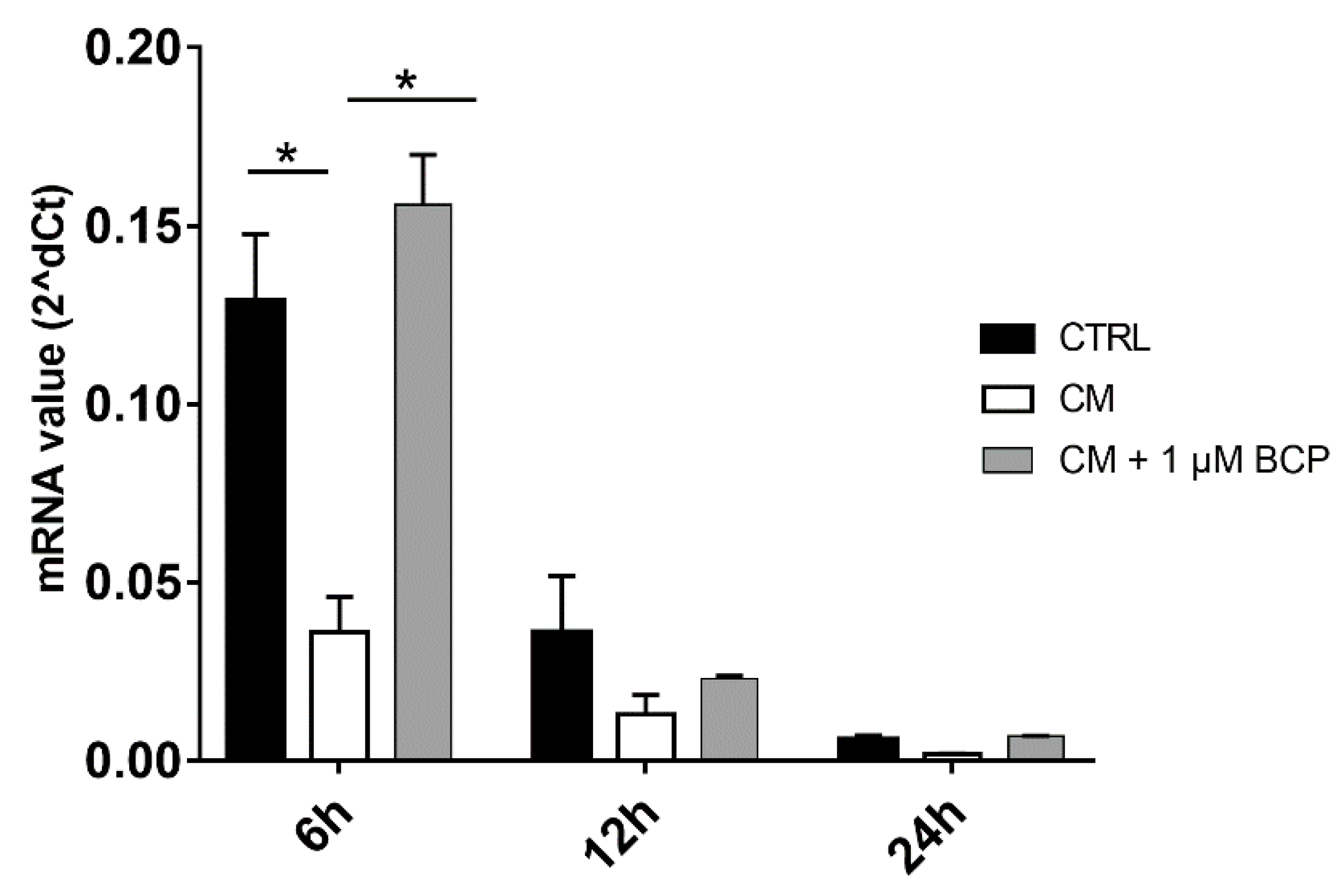
2.3. BCP, AA and GlcN Mixtures Reduce Chondrocyte Expression of IL-1beta, NF-κB and MMP-13
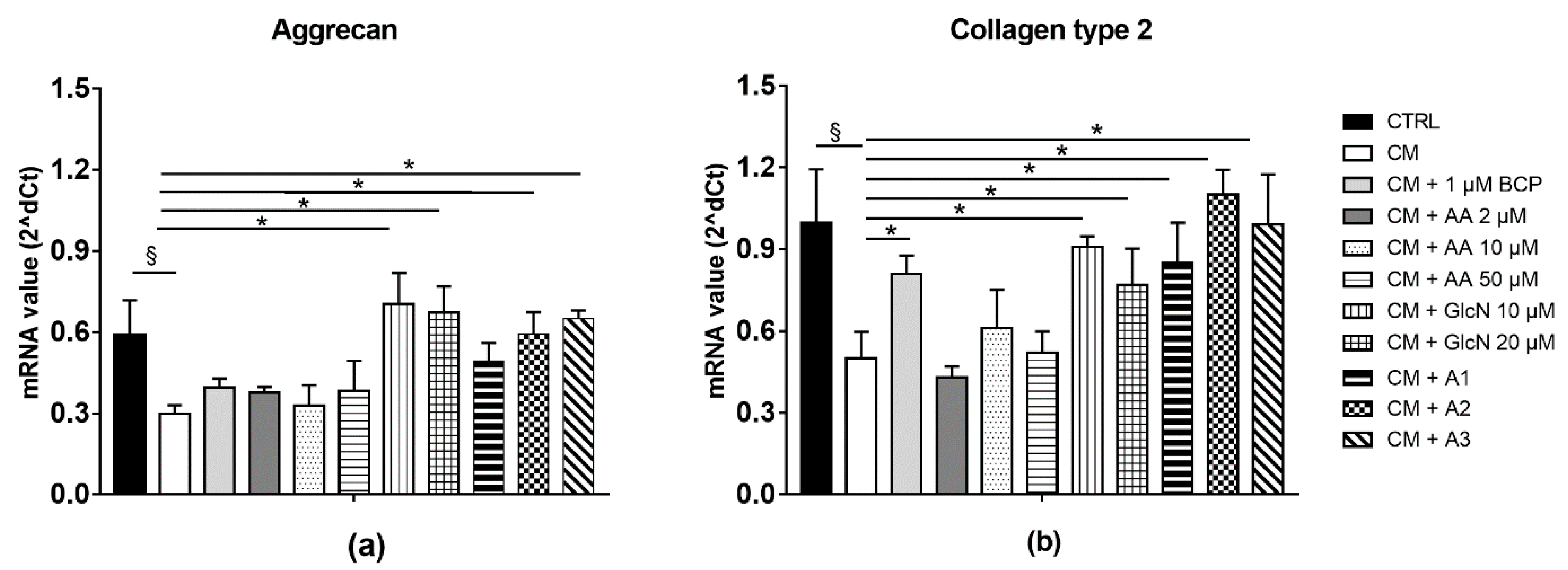
2.4. BCP, AA and GlcN Mixture Upregulate the Expression of Collagen Type II and Aggrecan
2.5. The Effect of BCP Is PPAR-Mediated
3. Discussion
4. Materials and Methods
4.1. Drugs and Chemicals
4.2. Primary Human Chondrocyte Culture
4.3. Activated U937 Monocyte Conditioned Medium
4.4. Evaluation of the Effects of BCP, AA and GlcN on Human Chondrocyte Viability
4.5. Analysis of Antioxidative, Anti-Inflammatory and Regenerative Effects Induced by BCP, AA and GlcN
4.5.1. Detection of Intracellular ROS Generation
4.5.2. RNA Isolation and qPCR Analysis
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Glyn-Jones, S.; Palmer, A.J.; Agricola, R.; Price, A.J.; Vincent, T.L.; Weinans, H.; Carr, A.J. Osteoarthritis. Lancet 2015, 386, 376–387. [Google Scholar] [CrossRef]
- Loeser, R.F.; Goldring, S.R.; Scanzello, C.R.; Goldring, M.B. Osteoarthritis: A disease of the joint as an organ. Arthritis Rheum. 2012, 64, 1697–1707. [Google Scholar] [CrossRef] [Green Version]
- Kraus, V.B.; McDaniel, G.; Huebner, J.L.; Stabler, T.V.; Pieper, C.F.; Shipes, S.W.; Petry, N.A.; Low, P.S.; Shen, J.; McNearney, T.A.; et al. Direct in vivo evidence of activated macrophages in human osteoarthritis. Osteoarthr. Cartil. 2016, 24, 1613–1621. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, M.; Nam, J. The role of changes in extracellular matrix of cartilage in the presence of inflammation on the pathology of osteoarthritis. BioMed Res. Int. 2013, 2013, 284873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivotto, E.; Otero, M.; Marcu, K.B.; Goldring, M.B. Pathophysiology of osteoarthritis: Canonical NF-κB/IKKβ-dependent and kinase-independent effects of IKKα in cartilage degradation and chondrocyte differentiation. RMD Open 2015, 1, e000061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mengshol, J.A.; Vincenti, M.P.; Brinckerhoff, C.E. IL-1 induces collagenase-3 (MMP-13) promoter activity in stably transfected chondrocytic cells: Requirement for Runx-2 and activation by p38 MAPK and JNK pathways. Nucleic Acids Res. 2001, 29, 4361–4372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakibaei, M.; John, T.; Schulze-Tanzil, G.; Lehmann, I.; Mobasheri, A. Suppression of NF-kappaB activation by curcumin leads to inhibition of expression of cyclo-oxygenase-2 and matrix metalloproteinase-9 in human articular chondrocytes: Implications for the treatment of osteoarthritis. Biochem. Pharmacol. 2007, 73, 1434–1445. [Google Scholar] [CrossRef]
- Douglas, R.J. Corticosteroid injection into the osteoarthritic knee: Drug selection, dose, and injection frequency. Int. J. Clin. Pract. 2012, 66, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Godwin, M.; Dawes, M. Intra-articular steroid injections for painful knees. Systematic review with meta-analysis. Can. Fam. Physician Med. Fam. Can. 2004, 50, 241–248. [Google Scholar]
- Stellavato, A.; Restaino, O.F.; Vassallo, V.; Finamore, R.; Ruosi, C.; Cassese, E.; De Rosa, M.; Schiraldi, C. Comparative Analyses of Pharmaceuticals or Food Supplements Containing Chondroitin Sulfate: Are Their Bioactivities Equivalent? Adv. Ther. 2019, 36, 3221–3237. [Google Scholar] [CrossRef] [Green Version]
- Brun, P.; Zavan, B.; Vindigni, V.; Schiavinato, A.; Pozzuoli, A.; Iacobellis, C.; Abatangelo, G. In vitro response of osteoarthritic chondrocytes and fibroblast-like synoviocytes to a 500–730 kDa hyaluronan amide derivative. J. Biomed. Mater. Res. Part B Appl. Biomater. 2012, 100, 2073–2081. [Google Scholar] [CrossRef]
- Tarricone, E.; Mattiuzzo, E.; Belluzzi, E.; Elia, R.; Benetti, A.; Venerando, R.; Vindigni, V.; Ruggieri, P.; Brun, P. Anti-Inflammatory Performance of Lactose-Modified Chitosan and Hyaluronic Acid Mixtures in an In Vitro Macrophage-Mediated Inflammation Osteoarthritis Model. Cells 2020, 9, 1328. [Google Scholar] [CrossRef]
- Krustev, E.; Reid, A.; McDougall, J.J. Tapping into the endocannabinoid system to ameliorate acute inflammatory flares and associated pain in mouse knee joints. Arthritis Res. Ther. 2014, 16, 437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irrera, N.; D’Ascola, A.; Pallio, G.; Bitto, A.; Mazzon, E.; Mannino, F.; Squadrito, V.; Arcoraci, V.; Minutoli, L.; Campo, G.M.; et al. β-Caryophyllene Mitigates Collagen Antibody Induced Arthritis (CAIA) in Mice Through a Cross-Talk between CB2 and PPAR-γ Receptors. Biomolecules 2019, 9, 326. [Google Scholar] [CrossRef] [Green Version]
- Burston, J.J.; Sagar, D.R.; Shao, P.; Bai, M.; King, E.; Brailsford, L.; Turner, J.M.; Hathway, G.J.; Bennett, A.J.; Walsh, D.A.; et al. Cannabinoid CB2 receptors regulate central sensitization and pain responses associated with osteoarthritis of the knee joint. PLoS ONE 2013, 8, e80440. [Google Scholar] [CrossRef] [Green Version]
- Dunn, S.L.; Wilkinson, J.M.; Crawford, A.; Bunning, R.A.D.; Le Maitre, C.L. Expression of Cannabinoid Receptors in Human Osteoarthritic Cartilage: Implications for Future Therapies. Cannabis Cannabinoid Res. 2016, 1, 3–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roche, R.; Hoareau, L.; Bes-Houtmann, S.; Gonthier, M.P.; Laborde, C.; Baron, J.F.; Haffaf, Y.; Cesari, M.; Festy, F. Presence of the cannabinoid receptors, CB1 and CB2, in human omental and subcutaneous adipocytes. Histochem. Cell Biol. 2006, 126, 177–187. [Google Scholar] [CrossRef]
- Turcotte, C.; Blanchet, M.R.; Laviolette, M.; Flamand, N. The CB(2) receptor and its role as a regulator of inflammation. Cell. Mol. Life Sci. CMLS 2016, 73, 4449–4470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Sullivan, S.E. An update on PPAR activation by cannabinoids. Br. J. Pharmacol. 2016, 173, 1899–1910. [Google Scholar] [CrossRef] [Green Version]
- Richardson, D.; Pearson, R.G.; Kurian, N.; Latif, M.L.; Garle, M.J.; Barrett, D.A.; Kendall, D.A.; Scammell, B.E.; Reeve, A.J.; Chapman, V. Characterisation of the cannabinoid receptor system in synovial tissue and fluid in patients with osteoarthritis and rheumatoid arthritis. Arthritis Res. Ther. 2008, 10, R43. [Google Scholar] [CrossRef] [Green Version]
- Gouze, J.N.; Gouze, E.; Popp, M.P.; Bush, M.L.; Dacanay, E.A.; Kay, J.D.; Levings, P.P.; Patel, K.R.; Saran, J.P.; Watson, R.S.; et al. Exogenous glucosamine globally protects chondrocytes from the arthritogenic effects of IL-1beta. Arthritis Res. Ther. 2006, 8, R173. [Google Scholar] [CrossRef] [Green Version]
- Sandy, J.D.; Gamett, D.; Thompson, V.; Verscharen, C. Chondrocyte-mediated catabolism of aggrecan: Aggrecanase-dependent cleavage induced by interleukin-1 or retinoic acid can be inhibited by glucosamine. Biochem. J. 1998, 335 Pt 1, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Chiu, P.-R.; Hu, Y.-C.; Huang, T.-C.; Hsieh, B.-S.; Yeh, J.-P.; Cheng, H.-L.; Huang, L.-W.; Chang, K.-L. Vitamin C Protects Chondrocytes against Monosodium Iodoacetate-Induced Osteoarthritis by Multiple Pathways. Int. J. Mol. Sci. 2016, 18, 38. [Google Scholar] [CrossRef] [Green Version]
- Chang, Z.; Huo, L.; Li, P.; Wu, Y.; Zhang, P. Ascorbic acid provides protection for human chondrocytes against oxidative stress. Mol. Med. Rep. 2015, 12, 7086–7092. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Ma, W.; Du, J. β-Caryophyllene (BCP) ameliorates MPP+ induced cytotoxicity. Biomed. Pharmacother. Biomed. Pharmacother. 2018, 103, 1086–1091. [Google Scholar] [CrossRef] [PubMed]
- Assis, L.C.; Straliotto, M.R.; Engel, D.; Hort, M.A.; Dutra, R.C.; de Bem, A.F. β-Caryophyllene protects the C6 glioma cells against glutamate-induced excitotoxicity through the Nrf2 pathway. Neuroscience 2014, 279, 220–231. [Google Scholar] [CrossRef]
- Askari, V.R.; Shafiee-Nick, R. Promising neuroprotective effects of β-caryophyllene against LPS-induced oligodendrocyte toxicity: A mechanistic study. Biochem. Pharmacol. 2019, 159, 154–171. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, D.; Levy, R.; Carroll, B. Toxicological Evaluation of β-Caryophyllene Oil: Subchronic Toxicity in Rats. Int. J. Toxicol. 2016, 35, 558–567. [Google Scholar] [CrossRef] [PubMed]
- Sharma, C.; Al Kaabi, J.M.; Nurulain, S.M.; Goyal, S.N.; Kamal, M.A.; Ojha, S. Polypharmacological Properties and Therapeutic Potential of β-Caryophyllene: A Dietary Phytocannabinoid of Pharmaceutical Promise. Curr. Pharm. Des. 2016, 22, 3237–3264. [Google Scholar] [CrossRef]
- Agiba, A.M. Nutraceutical formulations containing glucosamine and chondroitin sulphate in the treatment of osteoarthritis: Emphasis on clinical efficacy and formulation challenges. Int. J. Curr. Pharm. Res. 2017, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Ragle, R.L.; Sawitzke, A.D. Nutraceuticals in the management of osteoarthritis: A critical review. Drugs Aging 2012, 29, 717–731. [Google Scholar] [CrossRef]
- Navarro, V.J.; Khan, I.; Björnsson, E.; Seeff, L.B.; Serrano, J.; Hoofnagle, J.H. Liver injury from herbal and dietary supplements. Hepatology 2017, 65, 363–373. [Google Scholar] [CrossRef] [Green Version]
- Ronis, M.J.J.; Pedersen, K.B.; Watt, J. Adverse Effects of Nutraceuticals and Dietary Supplements. Annu. Rev. Pharmacol. Toxicol. 2018, 58, 583–601. [Google Scholar] [CrossRef]
- Klauke, A.L.; Racz, I.; Pradier, B.; Markert, A.; Zimmer, A.M.; Gertsch, J.; Zimmer, A. The cannabinoid CB₂ receptor-selective phytocannabinoid beta-caryophyllene exerts analgesic effects in mouse models of inflammatory and neuropathic pain. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2014, 24, 608–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, Y.; Wang, W.; Zhang, C.; Wu, Y.; Liu, Y.; Zhou, X. Cannabinoid WIN-55,212-2 mesylate inhibits ADAMTS-4 activity in human osteoarthritic articular chondrocytes by inhibiting expression of syndecan-1. Mol. Med. Rep. 2016, 13, 4569–4576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pahl, H.L. Activators and target genes of Rel/NF-κB transcription factors. Oncogene 1999, 18, 6853–6866. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.P.; Gan, Y.H.; Zhang, C.G.; Zhou, C.Y.; Ma, K.T.; Meng, J.H.; Ma, X.C. Requirement of the NF-κB pathway for induction of Wnt-5A by interleukin-1β in condylar chondrocytes of the temporomandibular joint: Functional crosstalk between the Wnt-5A and NF-κB signaling pathways. Osteoarthr. Cartil. 2011, 19, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Fan, Z.; Yang, H.; Bau, B.; Söder, S.; Aigner, T. Role of mitogen-activated protein kinases and NFkappaB on IL-1beta-induced effects on collagen type II, MMP-1 and 13 mRNA expression in normal articular human chondrocytes. Rheumatol. Int. 2006, 26, 900–903. [Google Scholar] [CrossRef]
- Wu, C.; Jia, Y.; Lee, J.H.; Jun, H.J.; Lee, H.S.; Hwang, K.Y.; Lee, S.J. trans-Caryophyllene is a natural agonistic ligand for peroxisome proliferator-activated receptor-α. Bioorganic Med. Chem. Lett. 2014, 24, 3168–3174. [Google Scholar] [CrossRef]
- Youssef, D.A.; El-Fayoumi, H.M.; Mahmoud, M.F. Beta-caryophyllene alleviates diet-induced neurobehavioral changes in rats: The role of CB2 and PPAR-γ receptors. Biomed. Pharmacother. Biomed. Pharmacother. 2019, 110, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Bento, A.F.; Marcon, R.; Dutra, R.C.; Claudino, R.F.; Cola, M.; Leite, D.F.P.; Calixto, J.B. β-Caryophyllene inhibits dextran sulfate sodium-induced colitis in mice through CB2 receptor activation and PPARγ pathway. Am. J. Pathol. 2011, 178, 1153–1166. [Google Scholar] [CrossRef] [PubMed]
- Picciolo, G.; Pallio, G.; Altavilla, D.; Vaccaro, M.; Oteri, G.; Irrera, N.; Squadrito, F. β-Caryophyllene Reduces the Inflammatory Phenotype of Periodontal Cells by Targeting CB2 Receptors. Biomedicines 2020, 8, 164. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Z.; Raghunath, M. Focus on collagen: In vitro systems to study fibrogenesis and antifibrosis state of the art. Fibrogenesis Tissue Repair 2009, 2, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brun, P.; Abatangelo, G.; Radice, M.; Zacchi, V.; Guidolin, D.; Gordini, D.D.; Cortivo, R. Chondrocyte aggregation and reorganization into three-dimensional scaffolds. J. Biomed. Mater. Res. 1999, 46, 337–346. [Google Scholar] [CrossRef]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival. Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Accession Number) | Name | Primer Sequences |
|---|---|---|
| ACAN (NM_001135.3) | Aggrecan | Fw 5′-TGCGGGTCAACAGTGCCTATC-3′ Rv 5′-CACGATGCCTTTCACCACGAC-3′ |
| Col2A1 (NM_001844.5) | Collagen Type II Alpha 1 Chain | Fw 5′-CGCTGGTGCTGCTGACGCTGCTCGT-3′ Rv 5′-GGCACCTTTTTCACCTTTGTCAC-3′ |
| IL-6 (JQ250825.1) | Interleukin 6 | Fw 5′-CACGCCTTGGACAGAATCCA-3′ Rv 5′-CCTCCAGCAACCAGGAATGT-3′ |
| IL-1β (NM_000576.3) | Interleukin 1 beta | Fw 5′-GAATCTCCGACCACCACTACAG-3′ Rv 5′-TGATCGTACAGGTGCATCGTG-3′ |
| LSGALS1 (NM_002305.4) | Galectin 1 | Fw 5′-TCTCGGGTGGAGTCTTCTGA-3′ Rv 5′-GTTCAGCACGAAGCTCTTAGC-3′ |
| LSGALS3 (NM_002306.4) | Galectin 3 | Fw 5′-CTGCTGGGGCACTGATTGT-3′ Rv 5′-TGTTTGCATTGGGCTTCACC-3′ |
| MMP-13 (AY741163.1) | Matrix metalloproteinase 13 | Fw 5′-AACGCCAGACAAATGTGACC-3′ Rv 5′-AGGTCATGAGAAGGGTGCTC-3′ |
| NF-κB-1 (NM_001243985.2) | nuclear factor kappa B subunit 1 | Fw 5′-CCGGGATGGCTTCTATGAGG-3′ Rv 5′- GGGGTTGTTGTTGGTCTGGA-3′ |
| PPAR γ (NM_001330615.2) | peroxisome proliferator activated receptor gamma | Fw 5′-ACCCAGAAAGCGATTCCTTCA-3′ Rv 5′-AGTGGTCTTCCATTACGGAGAGATC-3′ |
| PPIA (NM_021130.5) | Peptidylprolyl Isomerase A | Fw 5′-GGGCTTTAGGCTGTAGGTCAA-3′ Rv 5′-AACCAAAGCTAGGGAGAGGC-3′ |
| TNF-α (NM_000594.3) | TNF- alpha | Fw 5′-AAGCCTGTAGCCCATGTTGT-3′ Rv 5′-GGACCTGGGAGTAGATGAGGT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mattiuzzo, E.; Faggian, A.; Venerando, R.; Benetti, A.; Belluzzi, E.; Abatangelo, G.; Ruggieri, P.; Brun, P. In Vitro Effects of Low Doses of β-Caryophyllene, Ascorbic Acid and d-Glucosamine on Human Chondrocyte Viability and Inflammation. Pharmaceuticals 2021, 14, 286. https://doi.org/10.3390/ph14030286
Mattiuzzo E, Faggian A, Venerando R, Benetti A, Belluzzi E, Abatangelo G, Ruggieri P, Brun P. In Vitro Effects of Low Doses of β-Caryophyllene, Ascorbic Acid and d-Glucosamine on Human Chondrocyte Viability and Inflammation. Pharmaceuticals. 2021; 14(3):286. https://doi.org/10.3390/ph14030286
Chicago/Turabian StyleMattiuzzo, Elena, Alessia Faggian, Rina Venerando, Andrea Benetti, Elisa Belluzzi, Giovanni Abatangelo, Pietro Ruggieri, and Paola Brun. 2021. "In Vitro Effects of Low Doses of β-Caryophyllene, Ascorbic Acid and d-Glucosamine on Human Chondrocyte Viability and Inflammation" Pharmaceuticals 14, no. 3: 286. https://doi.org/10.3390/ph14030286
APA StyleMattiuzzo, E., Faggian, A., Venerando, R., Benetti, A., Belluzzi, E., Abatangelo, G., Ruggieri, P., & Brun, P. (2021). In Vitro Effects of Low Doses of β-Caryophyllene, Ascorbic Acid and d-Glucosamine on Human Chondrocyte Viability and Inflammation. Pharmaceuticals, 14(3), 286. https://doi.org/10.3390/ph14030286