
Dipeptidyl Peptidase (DPP)-IV Inhibitors with Antioxidant Potential Isolated from Natural Sources: A Novel Approach for the Management of Diabetes



Abstract
:1. Introduction
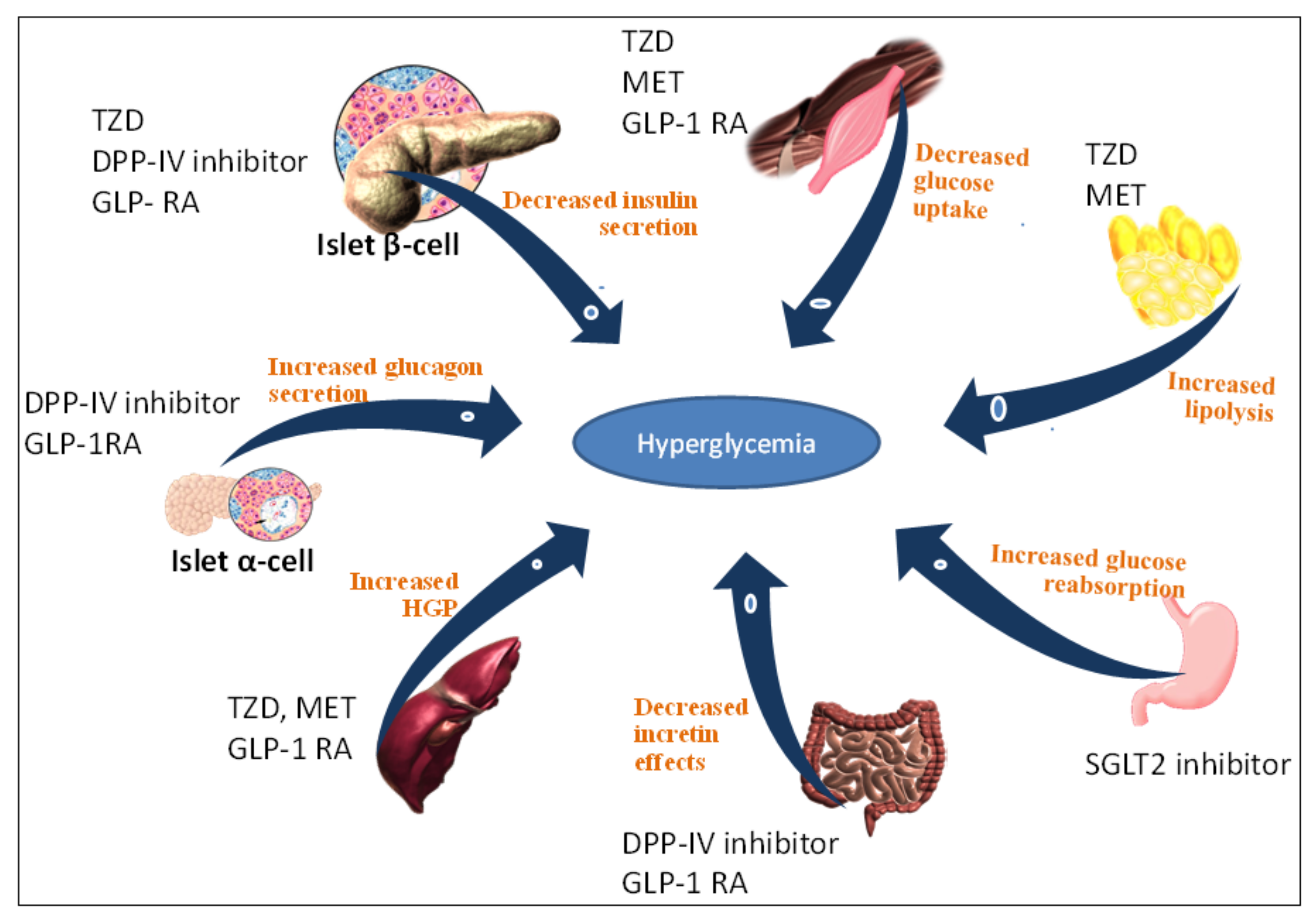
2. T2DM Treatment Strategy Based on DPP-IV Serine Protease Inhibition
3. DPP-IV Structure and Background
 Proline (P) Polypeptide Chain X- P + Polypeptide Chain
Proline (P) Polypeptide Chain X- P + Polypeptide Chain
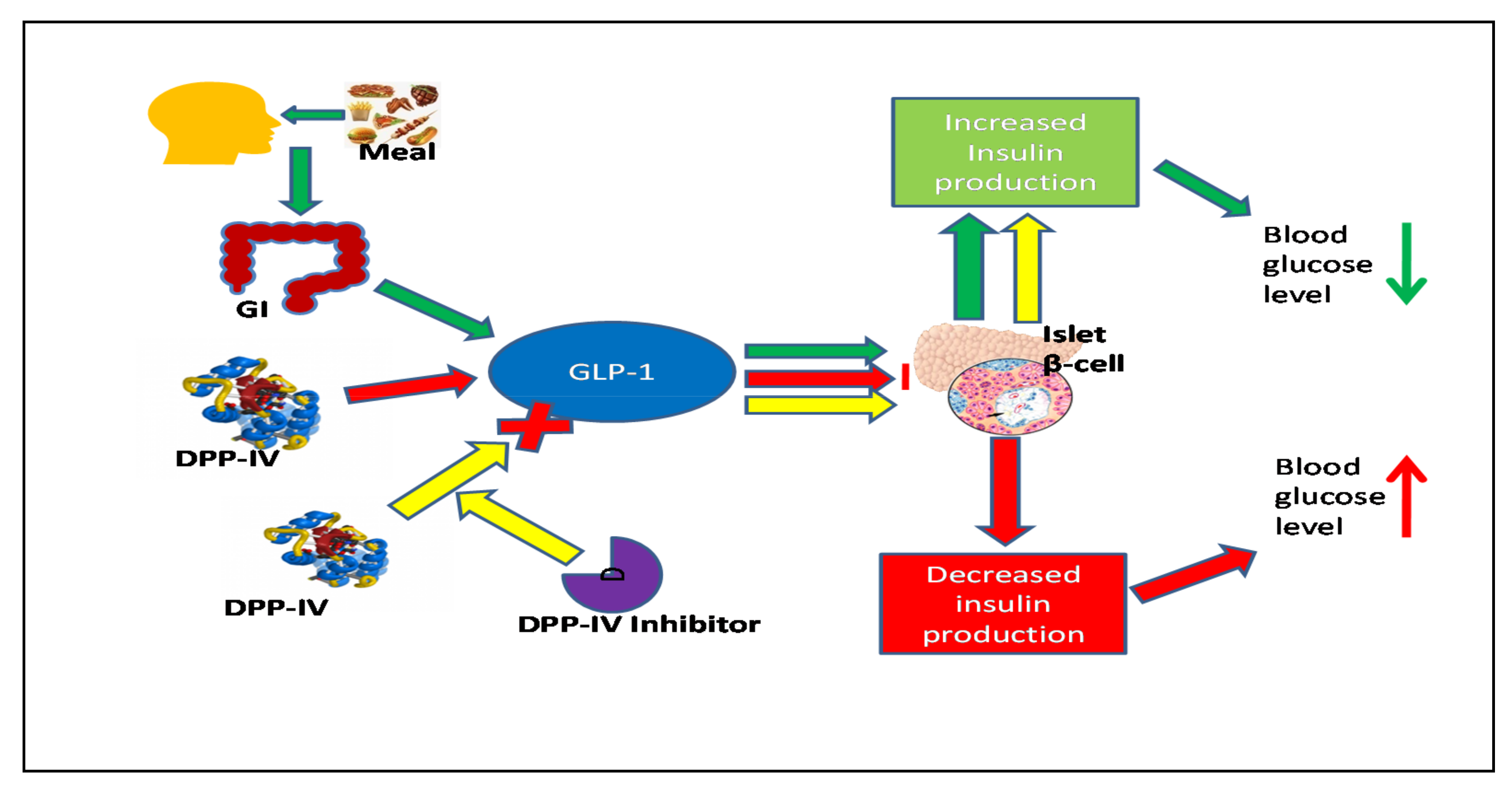
4. Distribution and Expression of the DPP-IV Enzyme & Molecular Mechanisms of the GLP-1 Receptor on the Pancreatic Endocrine System
5. Importance of Antioxidants in Overcoming Oxidative Stress in Disease and Ageing
6. Biological Actions of Incretin Hormone and Peripheral Glucose Sensors
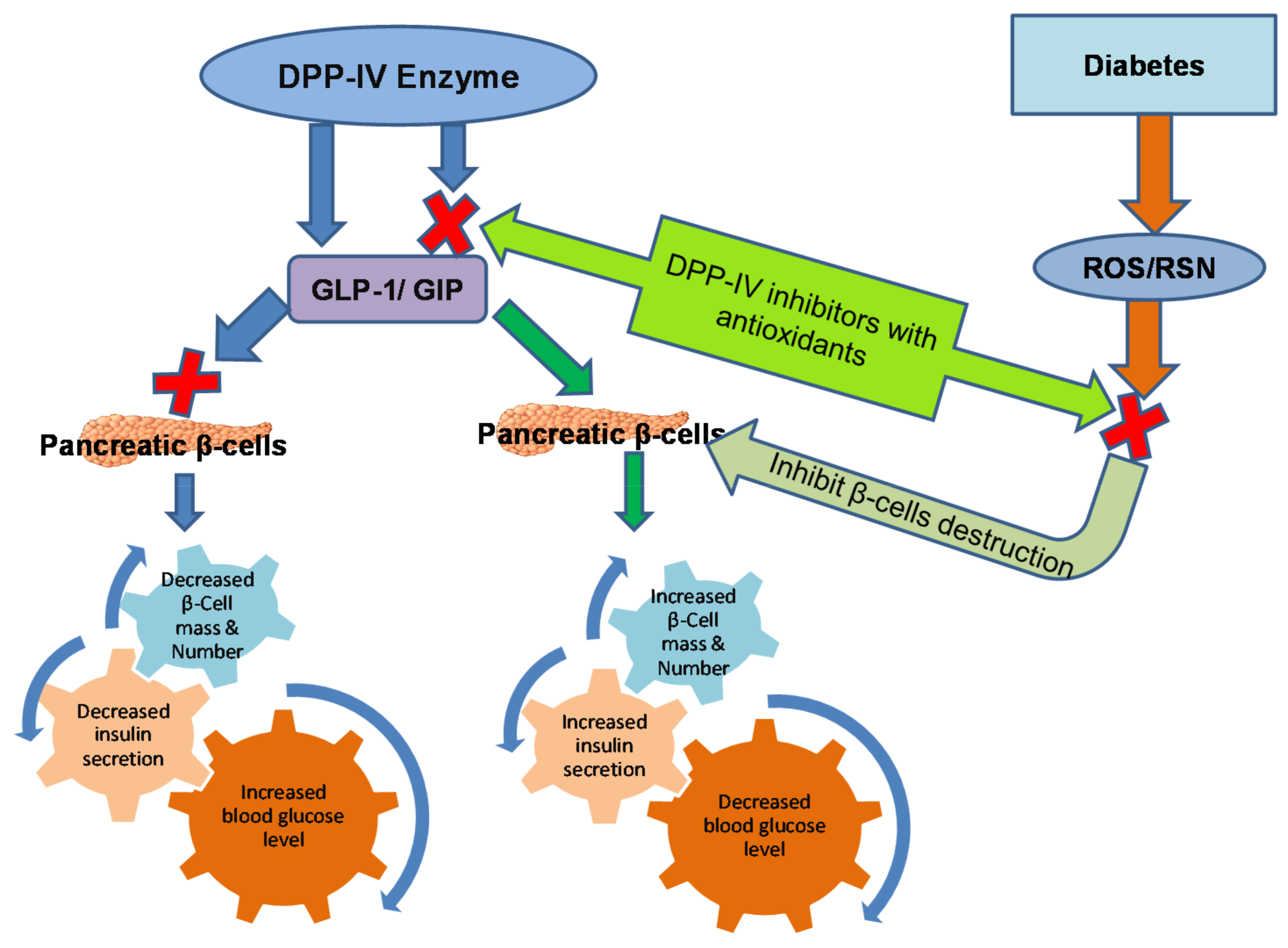
7. Mechanism of DPP-IV Inhibitors with Antioxidant Potential
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- White, J.R. A brief history of the development of diabetes medications. Diabetes Spectr. 2014, 27, 82–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiasson, J.-L.; Josse, R.G.; Gomis, R.; Hanefeld, M.; Karasik, A.; Laakso, M.; The STOP-NIDDM Trial Research Group. Acarbose treatment and the risk of cardiovascular disease and hypertension in patients with impaired glucose tolerance. JAMA 2003, 290, 486–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, D.; Tiwari, A.; Mishra, M.; Subramanian, S.S.; Baghel, U.S.; Mahajan, S.; Bisen, P.S.; Prasad, G.B. Anti-hyperglycemic and anti-hyperlipidemic potential of a polyherbal preparation “diabegon” in metabolic syndrome subject with type 2 diabetes. Afr. J. Tradit. Complement Altern. Med. 2014, 11, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Krentz, A.J.; Bailey, C.J. Oral antidiabetic agents: Current role in type 2 diabetes mellitus. Drugs 2005, 65, 385–411. [Google Scholar] [CrossRef] [PubMed]
- Pinchevsky, Y.; Butkow, N.; Raal, F.J.; Chirwa, T.; Rothberg, A. Demographic and clinical factors associated with development of type 2 diabetes: A review of the literature. Int. J. Gen. Med. 2020, 13, 121–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Diabetes Federation. IDF Diabetes Atlas, 9th ed.; International Diabetes Federation: Brussels, Belgium, 2019. [Google Scholar]
- Bodmer, M.; Meier, C.; Krähenbühl, S.; Jick, S.S.; Meier, C.R. Metformin, sulfonylureas, or other anti-diabetes drugs and the risk of lactic acidosis or hypoglycemia: A nested case-control analysis. Diabetes Care 2008, 31, 2086–2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forst, T.; Bramlage, P. Vildagliptin, a dpp-4 inhibitor for the twice-daily treatment of type 2 diabetes mellitus with or without metformin. Expert Opin. Pharmacother. 2014, 15, 1299–1313. [Google Scholar] [CrossRef]
- Heine, R.J.; Van Gaal, L.F.; Johns, D.; Mihm, M.J.; Widel, M.H.; Brodows, R.G.; The GWAA Study Group. Exenatide versus insulin glargine in patients with suboptimally controlled type 2 diabetes: A randomized trial. Ann. Intern. Med. 2005, 143, 559–569. [Google Scholar] [CrossRef]
- Maritim, A.C.; Sanders, R.A.; Watkins, J.B. Diabetes, oxidative stress, and antioxidants: A review. J. Biochem. Mol. Toxicol. 2003, 17, 24–38. [Google Scholar] [CrossRef]
- Abedini, A.; Schmidt, A.M. Mechanisms of islet amyloidosis toxicity in type 2 diabetes. FEBS Lett. 2013, 587, 1119–1127. [Google Scholar] [CrossRef] [Green Version]
- Ram, H.; Singh, A.-K. Improvements in insulin resistance and β-cells dysfunction by ddp-4 inhibition potential of Withania Somnifera (L.) dunal root extract in type 2 diabetic rat. Biointerface Res. Appl. Chem. 2020, 11, 8141–8155. [Google Scholar] [CrossRef]
- Müller, T.; Finan, B.; Bloom, S.; D’Alessio, D.; Drucker, D.; Flatt, P.; Fritsche, A.; Gribble, F.; Grill, H.; Habener, J.; et al. Glucagon-like peptide 1 (glp-1). Mol. Metab. 2019, 30, 72–130. [Google Scholar] [CrossRef]
- Gribble, F.M.; Reimann, F. Metabolic messengers: Glucagon-like peptide 1. Nat. Metab. 2021, 3, 142–148. [Google Scholar] [CrossRef]
- Silveira, S.T.; Martínez-Maqueda, D.; Recio, I.; Hernández-Ledesma, B. Dipeptidyl peptidase-iv inhibitory peptides generated by tryptic hydrolysis of a whey protein concentrate rich in β-lactoglobulin. Food Chem. 2013, 141, 1072–1077. [Google Scholar] [CrossRef]
- Ng, V.W.S.; Glasg, F. Dipeptidyl peptidase (dpp) -iv inhibitor: A novel class of oral anti-hyperglycemic agents. Clin. Pharmacol. Ther. 2007, 11, 33–34. [Google Scholar]
- Paliwal, G.; Sharma, A.; Upadhyay, N.; Das, M.; Tiwari, A. Therapeutic stimulation of glp-1 protein by implementing in silico to in vitro approach for type-2 diabetes treatment. Middle East J. Sci. Res. 2015, 23, 1005–1011. [Google Scholar] [CrossRef]
- Lammi, C.; Zanoni, C.; Arnoldi, A.; Vistoli, G. Peptides derived from soy and lupin protein as dipeptidyl-peptidase iv inhibitors: In vitro biochemical screening and in silico molecular modeling study. J. Agric. Food Chem. 2016, 64, 9601–9606. [Google Scholar] [CrossRef] [Green Version]
- Mowla, A.; Alauddin, M.; Rahman, M.A.; Ahmed, K. Antihyperglycemic effect of trigonella foenum-graecum (fenugreek) seed extract in alloxan-induced diabetic rats and its use in diabetes mellitus: A brief qualitative phytochemical and acute toxicity test on the extract. Afr. J. Tradit. Complement Altern. Med. 2009, 6, 255–261. [Google Scholar] [CrossRef] [Green Version]
- Bharti, S.K.; Krishnan, S.; Kumar, A.; Rajak, K.K.; Murari, K.; Bharti, B.K.; Gupta, A.K. Antihyperglycemic activity with dpp-iv inhibition of alkaloids from seed extract of castanospermum australe: Investigation by experimental validation and molecular docking. Phytomedicine 2012, 20, 24–31. [Google Scholar] [CrossRef]
- Jadav, P.; Bahekar, R.; Shah, S.R.; Patel, D.; Joharapurkar, A.; Kshirsagar, S.; Jain, M.; Shaikh, M.; Sairam, K.V.V.M. Long-acting peptidomimetics based dpp-iv inhibitors. Bioorg. Med. Chem. Lett. 2012, 22, 3516–3521. [Google Scholar] [CrossRef]
- Hopsu-Havu, V.K.; Glenner, G.G. A new dipeptide naphthylamidase hydrolyzing glycyl-prolyl-beta-naphthylamide. Histochemie 1966, 7, 197–201. [Google Scholar] [CrossRef]
- Barnett, A. Dpp-4 inhibitors and their potential role in the management of type 2 diabetes. Int. J. Clin. Pract. 2006, 60, 1454–1470. [Google Scholar] [CrossRef]
- Varona, A.; Blanco, L.; Perez, I.; Gil, J.; Irazusta, J.; López, J.I.; Candenas, M.L.; Pinto, F.M.; Larrinaga, G. Expression and activity profiles of DPP IV/CD26 and NEP/CD10 glycoproteins in the human renal cancer are tumor-type dependent. BMC Cancer 2010, 10, 193. [Google Scholar] [CrossRef] [Green Version]
- Leiting, B.; Pryor, K.D.; Wu, J.K.; Marsilio, F.; Patel, R.A.; Craik, C.S.; Ellman, J.A.; Cummings, R.T.; Thornberry, N.A. Catalytic properties and inhibition of proline-specific dipeptidyl peptidases ii, iv and vii. Biochem. J. 2003, 371, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.W.; Jin, L.; Piao, S.G.; Chung, B.H.; Yang, C.W. Inhibition of dipeptidyl peptidase iv protects tacrolimus-induced kidney injury. Lab. Invest. 2015, 95, 1174–1185. [Google Scholar] [CrossRef]
- Püschel, G.; Mentlein, R.; Heymann, E. Isolation and characterization of dipeptidyl peptidase iv from human placenta. Eur. J. Biochem. 1982, 126, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Davy, A.; Thomsen, K.K.; Juliano, M.A.; Alves, L.C.; Svendsen, I.; Simpson, D.J. Purification and characterization of barley dipeptidyl peptidase iv. Plant Physiol. 2000, 122, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.B.; FitzGerald, R.J. Inhibition of dipeptidyl peptidase iv (dpp-iv) by proline containing casein-derived peptides. J. Funct. Foods 2013, 5, 1909–1917. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.B.; FitzGerald, R.J. Susceptibility of milk protein-derived peptides to dipeptidyl peptidase iv (dpp-iv) hydrolysis. Food Chem. 2014, 145, 845–852. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-S.; Lee, E.-S.; Alam, M.; Jang, J.-H.; Lee, H.-S.; Oh, H.; Kim, Y.-C.; Manzoor, Z.; Koh, Y.-S.; Kang, D.-G.; et al. Soluble dpp-4 up-regulates toll-like receptors and augments inflammatory reactions, which are ameliorated by vildagliptin or mannose-6-phosphate. Metabolism 2016, 65, 89–101. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Matsui, T.; Maeda, S.; Higashimoto, Y.; Yamagishi, S.-I. Advanced glycation end products evoke endothelial cell damage by stimulating soluble dipeptidyl peptidase-4 production and its interaction with mannose 6-phosphate/insulin-like growth factor ii receptor. Cardiovasc. Diabetol. 2013, 12, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, T.; Kumagai, E.; Iwata, S.; Yamakawa, A. Soluble cd26/dipeptidyl peptidase iv enhances the transcription of il-6 and tnf-α in thp-1 cells and monocytes. PLoS ONE 2013, 8, e66520. [Google Scholar] [CrossRef] [PubMed]
- Deacon, C.F.; Ahrén, B.; Holst, J.J. Inhibitors of dipeptidyl peptidase iv: A novel approach for the prevention and treatment of type 2 diabetes? Expert Opin. Investig. Drugs 2004, 13, 1091–1102. [Google Scholar] [CrossRef] [PubMed]
- Cantini, G.; Mannucci, E.; Luconi, M. Perspectives in glp-1 research: New targets, new receptors. Trends Endocrinol. Metab. 2016, 27, 427–438. [Google Scholar] [CrossRef]
- Terrill, S.J.; Jackson, C.M.; Greene, H.E.; Lilly, N.; Maske, C.B.; Vallejo, S.; Williams, D.L. Role of lateral septum glucagon-like peptide 1 receptors in food intake. Am. J. Physiol. Integr. Comp. Physiol. 2016, 311, R124–R132. [Google Scholar] [CrossRef] [Green Version]
- Franz, M.J.; Bantle, J.P.; Beebe, C.A.; Brunzell, J.D.; Chiasson, J.-L.; Garg, A.; Holzmeister, L.A.; Hoogwerf, B.; Mayer-Davis, E.; Mooradian, A.D.; et al. Evidence-based nutrition principles and recommendations for the treatment and prevention of diabetes and related complications. Diabetes Care 2002, 25, 148–198. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V. Glucagon-like peptide-1 analogues: An overview. Indian J. Endocrinol. Metab. 2013, 17, 413–421. [Google Scholar] [CrossRef]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. IJBS 2008, 4, 89–96. [Google Scholar]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free radicals: Properties, sources, targets, and their implication in various diseases. Indian J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Kumar, H.; Lim, H.-W.; More, S.V.; Kim, B.-W.; Koppula, S.; Kim, I.S.; Choi, D.-K. The role of free radicals in the aging brain and parkinson’s disease: Convergence and parallelism. Int. J. Mol. Sci. 2012, 13, 10478–10504. [Google Scholar] [CrossRef] [Green Version]
- Yadav, D.; Mishra, M.; Joseph, A.Z.; Subramani, S.K.; Mahajan, S.; Singh, N.; Bisen, P.S.; Prasad, G.B. Status of antioxidant and lipid peroxidation in type 2 diabetic human subjects diagnosed with and without metabolic syndrome by using ncep-atpiii, idf and who criteria. Obes. Res. Clin. Pract. 2015, 9, 158–167. [Google Scholar] [CrossRef]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [Green Version]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ros and rns sources in physiological and pathological conditions. Oxid. Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef]
- Bardaweel, S.K.; Gul, M.; Alzweiri, M.; Ishaqat, A.; Alsalamat, H.A.; Bashatwah, R.M. Reactive oxygen species: The dual role in physiological and pathological conditions of the human body. Eurasian J. Med. 2018, 50, 193–201. [Google Scholar] [CrossRef]
- Guo, C.; Sun, L.; Chen, X.; Zhang, D. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen. Res. 2013, 8, 2003–2014. [Google Scholar] [CrossRef]
- Rahman, K. Studies on free radicals, antioxidants, and co-factors. Clin. Interv. Aging 2007, 2, 219–236. [Google Scholar]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef] [Green Version]
- Bashir, H.S.; Mohammed, A.M.; Magsoud, A.S.; Shaoub, A.M. Isolation of three flavonoids from withania somnifera leaves (solanaceae) and their antimicrobial activities. J. For. Prod. Ind. 2013, 2, 39–45. [Google Scholar]
- Mustafa, R.; Hamid, A.A.; Mohamed, S.; Bakar, F.A. Total phenolic compounds, flavonoids, and radical scavenging activity of 21 selected tropical plants. J. Food Sci. 2010, 75, C28–C35. [Google Scholar] [CrossRef]
- Kaurinovic, B.; Vastag, D. Flavonoids and phenolic acids as potential natural antioxidants. In Antioxidants; IntechOpen: London, UK, 2019. [Google Scholar]
- Katare, C.; Saxena, S.; Agrawal, S.; Joseph, A.Z.; Subramani, S.K.; Yadav, D.; Singh, N.; Bisen, P.S.; Prasad, G.B. Lipid-lowering and antioxidant functions of bottle gourd (lagenaria siceraria) extract in human dyslipidemia. J. Evid. Based Complementary Altern. Med. 2014, 19, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Mir, M.A.; Sawhney, S.S.; Jassal, M.M.S. Qualitative and quantitative analysis of phytochemicals of taraxacum officinale. Wudpecker J. Pharm. Pharmocology 2013, 2, 1–5. [Google Scholar]
- Raja, C.P.; Venkataraman, K. Aloe vera phytochemicals inhibits dipeptidyl peptidase iv (dpp-iv), an anti-diabetic target. Int. J. Pharm. Bio. Sci. 2016, 7, 120–128. [Google Scholar]
- Mojzer, E.B.; Hrnčič, M.K.; Škerget, M.; Knez, Ž.; Bren, U. Polyphenols: Extraction methods, antioxidative action, bioavailability and anticarcinogenic effects. Molecules 2016, 21, 901. [Google Scholar] [CrossRef]
- Li, A.-N.; Li, S.; Zhang, Y.-J.; Xu, X.-R.; Chen, Y.-M.; Li, H.-B. Resources and biological activities of natural polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef] [PubMed]
- Velarde-Salcedo, A.J.; Barrera-Pacheco, A.; Lara-Gonzalez, S.; Montero-Morán, G.M.; Díaz-Gois, A.; DE Mejia, E.; de la Rosa, A.P.B. In vitro inhibition of dipeptidyl peptidase iv by peptides derived from the hydrolysis of amaranth (amaranthus hypochondriacus L.) proteins. Food Chem. 2013, 136, 758–764. [Google Scholar] [CrossRef]
- Green, A.D.; Vasu, S.; Moffett, R.C.; Flatt, P.R. Biochimie co-culture of clonal beta cells with glp-1 and glucagon-secreting cell line impacts on beta cell insulin secretion, proliferation and susceptibility to cytotoxins. Biochimie 2016, 125, 119–125. [Google Scholar] [CrossRef]
- Janssen, P.; Rotondo, A.; Mulé, F.; Tack, J. Review article: A comparison of glucagon-like peptides 1 and 2. Aliment. Pharmacol. Ther. 2013, 37, 18–36. [Google Scholar] [CrossRef] [Green Version]
- Seino, Y.; Fukushima, M.; Yabe, D. Gip and glp-1, the two incretin hormones: Similarities and differences. J. Diabetes Investig. 2010, 1, 8–23. [Google Scholar] [CrossRef] [Green Version]
- Brubaker, P.L.; Drucker, D.J. Minireview: Glucagon-like peptides regulate cell proliferation and apoptosis in the pancreas, gut, and central nervous system. Endocrinology 2004, 145, 2653–2659. [Google Scholar] [CrossRef] [Green Version]
- Cabou, C.; Burcelin, R. GLP-1, the gut-brain, and brain-periphery axes. Rev Diabet Stud. 2011, 8, 418–431. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Q.; Grandy, S.; Hiller, J.; Kostev, K. Clinical and patient-related variables associated with initiating glp-1 receptor agonist therapy in type 2 diabetes patients in primary care in Germany. PLoS ONE 2016, 11, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kulve, J.S.T.; Veltman, D.J.; Van Bloemendaal, L.; Barkhof, F.; Deacon, C.F.; Holst, J.J.; Konrad, R.J.; Sloan, J.H.; Drent, M.L.; Diamant, M.; et al. Endogenous glp-1 mediates postprandial reductions in activation in central reward and satiety areas in patients with type 2 diabetes. Diabetologia 2015, 58, 2688–2698. [Google Scholar] [CrossRef] [Green Version]
- Fujita, Y.; Wideman, R.D.; Asadi, A.; Yang, G.; Baker, R.; Webber, T.; Zhang, T.; Wang, R.; Ao, Z.; Warnock, G.L.; et al. Glucose-dependent insulinotropic polypeptide is expressed in pancreatic islet α-cells and promotes insulin secretion. Gastroenterology 2010, 138, 1966–1975.e1. [Google Scholar] [CrossRef]
- Young, A.A.; Gedulin, B.R.; Bhavsar, S.; Bodkin, N.; Jodka, C.; Hansen, B.; Denaro, M. Glucose-lowering and insulin-sensitizing actions of exendin-4: Studies in obese diabetic (ob/ob, db/db) mice, diabetic fatty zucker rats, and diabetic rhesus monkeys (macaca mulatta). Diabetes 1999, 48, 1026–1034. [Google Scholar] [CrossRef]
- Gallwitz, B. Glp-1 agonists and dipeptidyl-peptidase iv inhibitors. Handb. Exp. Pharmacol. 2011, 203, 53–74. [Google Scholar] [CrossRef]
- Łabuzek, K.; Kozłowski, M.; Szkudłapski, D.; Sikorska, P.; Kozłowska, M.; Okopień, B. Incretin-based therapies in the treatment of type 2 diabetes—More than meets the eye? Eur. J. Intern. Med. 2013, 24, 207–212. [Google Scholar] [CrossRef]
- Mari, A.; Sallas, W.M.; He, Y.L.; Watson, C.; Ligueros-Saylan, M.; Dunning, B.E.; Deacon, C.F.; Holst, J.J.; Foley, J.E. Vildagliptin, a dipeptidyl peptidase-iv inhibitor, improves model-assessed beta-cell function in patients with type 2 diabetes. J. Clin. Endocrinol. Metab. 2005, 90, 4888–4894. [Google Scholar] [CrossRef] [Green Version]
- Trujillo, J.M.; Nuffer, W.; Ellis, S.L. Glp-1 receptor agonists: A review of head-to-head clinical studies. Ther. Adv. Endocrinol. Metab. 2015, 6, 19–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, T.P.; Vangaveti, V.N.; Malabu, U.H. Dipeptidyl peptidase-4 inhibitors and their potential role in the management of atherosclerosis--a review. Diabetes Metab. Syndr. 2015, 9, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Yogisha, S.; Raveesha, K.A. Dipeptidyl Peptidase IV inhibitory activity of Mangifera indica. J. Nat. Prod. 2010, 3, 76–79. [Google Scholar]
- Ta, N.N.; Schuyler, C.A.; Li, Y.; Lopes-Virella, M.F.; Huang, Y. Dpp-4 (cd26) inhibitor alogliptin inhibits atherosclerosis in diabetic apolipoprotein e-deficient mice. J. Cardiovasc. Pharmacol. 2011, 58, 157–166. [Google Scholar] [CrossRef]
- Hiromura, M.; Nohtomi, K.; Mori, Y.; Kataoka, H.; Sugano, M.; Ohnuma, K.; Kuwata, H.; Hirano, T. Caveolin-1, a binding protein of cd26, is essential for the anti-inflammatory effects of dipeptidyl peptidase-4 inhibitors on human and mouse macrophages. Biochem. Biophys. Res. Commun. 2018, 495, 223–229. [Google Scholar] [CrossRef]
- Terasaki, M.; Yashima, H.; Mori, Y.; Saito, T.; Matsui, T.; Hiromura, M.; Kushima, H.; Osaka, N.; Ohara, M.; Fukui, T.; et al. A dipeptidyl peptidase-4 inhibitor inhibits foam cell formation of macrophages in type 1 diabetes via suppression of cd36 and acat-1 expression. Int. J. Mol. Sci. 2020, 21, 4811. [Google Scholar] [CrossRef]
- Mardanyan, S.; Sharoyan, S.; Antonyan, A.; Zakaryan, N. Dipeptidyl peptidase iv and adenosine deaminase inhibition by Armenian plants and antidiabetic drugs. Int. J. Diabetes Metab. 2011, 19, 69–74. [Google Scholar]
- Saidu, Y.; Muhammad, S.; Yahaya, A.; Onu, A.; Mohammed, I.; Muhammad, L. In vitro screening for protein tyrosine phosphatase 1b and dipeptidyl peptidase iv inhibitors from Nigerian medicinal plants. J. Intercult. Ethnopharmacol. 2017, 6, 154–157. [Google Scholar] [CrossRef]
- Singh, A.K.; Jatwa, R.; Purohit, A.; Ram, H. Synthetic and phytocompounds based dipeptidyl peptidase-iv (dpp-iv) inhibitors for therapeutics of diabetes. J. Asian Nat. Prod. Res. 2017, 19, 1036–1045. [Google Scholar] [CrossRef]
- Mallikharjuna, P.B.; Rajanna, L.N.; Seetharam, Y.N.; Sharanabasappa, G.K. Phytochemical studies of strychnos potatorum l.F.—A medicinal plant. E J. Chem. 2007, 4, 510–518. [Google Scholar] [CrossRef]
- Singh, A.-k.; Jatwa, R.; Joshi, J. Cytoprotective and dipeptidyl peptidase-iv (dpp-iv/cd26) inhibitory roles of ocimum sanctum and momordica charantia extract. Asian J. Pharm. Clin. Res. 2014, 7, 115–120. [Google Scholar]
- Singh, A.K.; Patel, P.K.; Choudhary, K.; Joshi, J.; Yadav, D.; Jin, J.O. Quercetin and coumarin inhibit dipeptidyl peptidase-iv and exhibits antioxidant properties: In silico, in vitro, ex vivo. Biomolecules 2020, 10, 207. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.S.; Kim, H.M.; Kang, J.S.; Lee, E.Y.; Yadav, D.; Kwon, M.H.; Kim, Y.M.; Kim, H.S.; Chung, C.H. Oleanolic acid and n-acetylcysteine ameliorate diabetic nephropathy through reduction of oxidative stress and endoplasmic reticulum stress in a type 2 diabetic rat model. Nephrol. Dial. Transplant. 2016, 31, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Krishnaiah, D.; Devi, T.; Bono, A.; Sarbatly, R. Studies on phytochemical constituents of six malaysian medicinal plants. J. Med. Plants Res. 2009, 3, 67–72. [Google Scholar]
- Ameh, G.I.; Eze, C.S. Phytochemical constituents of some Nigerian plants. BioResearch 2010, 8, 685–688. [Google Scholar] [CrossRef]
- Imamura, K.; Tsuyama, Y.; Hirata, T. Identification and characterization of a novel fermented substance produced by edible aspergillus oryzae ao-1 that inhibits dpp-iv activity. J. Biosci. Bioeng. 2010, 111, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, R.; Bhavtaran, S.; Narendra, P.; Varghese, N. Journal of natural products dipeptidyl peptidase- iv inhibitory activity of Berberis aristata. Nat. Prod. 2011, 4, 158–163. [Google Scholar]
- Parmar, H.S.; Jain, P.; Chauhan, D.S.; Bhinchar, M.K.; Munjal, V.; Yusuf, M.; Choube, K.; Tawani, A.; Tiwari, V.; Manivannan, E.; et al. Dpp-iv inhibitory potential of naringin: An in silico, in vitro and in vivo study. Diabetes Res. Clin. Pract. 2012, 97, 105–111. [Google Scholar] [CrossRef]
- Estrada-Salas, P.A.; Montero-Morán, G.M.; Martínez-Cuevas, P.P.; González, C.; Barba de la Rosa, A.P. Characterization of antidiabetic and antihypertensive properties of canary seed (Phalaris canariensis L.) peptides. J. Agric. Food Chem. 2014, 62, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Joshi, J.; Jatwa, R. Dipeptidyl peptidase iv (dpp-iv/cd26) inhibitory and free radical scavenging potential of w. Somnifera and t. Foenum-graecum extract. Int. J. Phytomed. 2013, 5, 503–509. [Google Scholar] [CrossRef]
- Borde, M.K.; Mohanty, I.R.; Suman, R.K.; Deshmukh, Y.A. Dipeptidyl peptidase-iv inhibitory activities of medicinal plants: Terminalia arjuna, Commiphora mukul, Gymnema sylvestre, Morinda citrifolia, Emblica officinalis. Asian J. Pharma. Clin. Res. 2016, 9, 1–3. [Google Scholar]
- Riyanti, S.; Suganda, A.G.; Sukandar, E.Y. Dipeptidyl peptidase-iv inhibitory activity of some indonesian medicinal plants. Asian J. Pharma. Clin. Res. 2016, 9, 375–377. [Google Scholar]
- Sila, A.; Alvarez, O.M.; Haddar, A.; Frikha, F.; Dhulster, P.; Nedjar-Arroume, N.; Bougatef, A. Purification, identification and structural modelling of dpp-iv inhibiting peptides from barbel protein hydrolysate. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1008, 260–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.N.; Wang, C.J.; Lee, Y.J.; Peng, C.H. Active subfractions of abelmoschus esculentus substantially prevent free fatty acid-induced ß cell apoptosis via inhibiting dipeptidyl peptidase-4. PLoS ONE 2017, 12, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Kinghorn, A.D.; Pan, L.; Fletcher, J.N.; Chai, H. The Relevance of Higher Plants in Lead Compound Discovery Programs. J. Nat. Prod. 2011, 74, 1539–1555. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.B.; Paolella, S.; Mudgil, P.; Maqsood, S.; FitzGerald, R.J. Identification of novel dipeptidyl peptidase iv (dpp-iv) inhibitory peptides in camel milk protein hydrolysates. Food Chem. 2018, 244, 340–348. [Google Scholar] [CrossRef] [Green Version]
- Ekayanti, M.; Sauriasari, R.; Elya, B. Dipeptidyl peptidase iv inhibitory activity of fraction from white tea ethanolic extract (Camellia sinensis (L.) kuntze) ex vivo. Pharmacogn. J. 2018, 10, 190–193. [Google Scholar] [CrossRef] [Green Version]
- Setyaningsih, E.P.; Saputri, F.C.; Mun’im, A. The antidiabetic effectivity of indonesian plants extracts via dpp-iv inhibitory mechanism. J. Young Pharm. 2019, 11, 161–164. [Google Scholar] [CrossRef] [Green Version]
- Ram, H.; Jaipal, N.; Kumar, P.; Deka, P.; Kumar, S.; Kashyap, P.; Kumar, S.; Singh, B.P.; Alqarawi, A.A.; Hashem, A.; et al. Dual inhibition of dpp-4 and cholinesterase enzymes by the phytoconstituents of the ethanolic extract of prosopis cineraria pods: Therapeutic implications for the treatment of diabetes-associated neurological impairments. Curr. Alzheimer Res. 2019, 16, 1230–1244. [Google Scholar] [CrossRef]
- Kalhotra, P.; Chittepu, V.C.S.R.; Osorio-Revilla, G.; Gallardo-Velazquez, T. Phytochemicals in garlic extract inhibit therapeutic enzyme dpp-4 and induce skeletal muscle cell proliferation: A possible mechanism of action to benefit the treatment of diabetes mellitus. Biomolecules 2020, 10, 305. [Google Scholar] [CrossRef] [Green Version]
- Pino, F.R.; Gálvez, A.R.P.; Carpio, F.J.E.; Guadix, E.M. Evaluation of tenebrio molitor protein as source of peptides modulating physiological processes. Food Funct. 2020, 1, 1–26. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, Z.; Liu, G.; Wu, Y.; Ouyang, J. Inhibitory effect of chestnut (Castanea mollissima Blume) inner skin extract on the activity of α-amylase, α-glucosidase, dipeptidyl peptidase iv and in vitro digestibility of starches. Food Chem. 2020, 324, 126847. [Google Scholar] [CrossRef]
- Harnedy-Rothwell, P.A.; McLaughlin, C.M.; O’Keeffe, M.B.; Le Gouic, A.V.; Allsopp, P.J.; McSorley, E.M.; Sharkey, S.; Whooley, J.; McGovern, B.; O’Harte, F.P.; et al. Identification and characterisation of peptides from a boarfish (Capros aper) protein hydrolysate displaying in vitro dipeptidyl peptidase-iv (dpp-iv) inhibitory and insulinotropic activity. Food Res. Int. 2020, 131, 108989. [Google Scholar] [CrossRef]
- Li, Y.; Aiello, G.; Bollati, C.; Bartolomei, M.; Arnoldi, A.; Lammi, C. Phycobiliproteins from arthrospira platensis (spirulina): A new source of peptides with dipeptidyl peptidase-iv inhibitory activity. Nutrients 2020, 12, 794. [Google Scholar] [CrossRef] [Green Version]
- Jia, C.-L.; Hussain, N.; Ujiroghene, O.J.; Pang, X.-Y.; Zhang, S.-W.; Lu, J.; Liu, L.; Lv, J.-P.; Obaroakpo, J.U. Generation and characterization of dipeptidyl peptidase-iv inhibitory peptides from trypsin-hydrolyzed α-lactalbumin-rich whey proteins. Food Chem. 2020, 318, 126333. [Google Scholar] [CrossRef]
- Ansari, P.; Azam, S.; Hannan, J.M.A.; Flatt, P.R.; Abdel Wahab, Y.H.A. Anti-hyperglycaemic activity of h. Rosa-sinensis leaves is partly mediated by inhibition of carbohydrate digestion and absorption, and enhancement of insulin secretion. J. Ethnopharmacol. 2020, 253, 112647. [Google Scholar] [CrossRef]
- Prasannaraja, C.; Kamalanathan, A.S.; Vijayalakshmi, M.A.; Venkataraman, K. A dipyrrole derivative from aloe vera inhibits an anti-diabetic drug target dipeptidyl peptidase (dpp)-iv in vitro. Prep. Biochem. Biotechnol. 2020, 50, 511–520. [Google Scholar] [CrossRef]
- Yang, Y.; Shi, C.Y.; Xie, J.; Dai, J.H.; He, S.L.; Tian, Y. Identification of potential dipeptidyl peptidase (dpp)-iv inhibitors among Moringa oleifera phytochemicals by virtual screening, molecular docking analysis, adme/t-based prediction, and in vitro analyses. Molecules 2020, 25, 189. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Generic Name | Country | Brand Name |
|---|---|---|
| Sitagliptin | Europe, US, Japan | Januvia |
| Vildagliptin | Europe, US, Japan | Galvus, Equa |
| Saxagliptin | Europe, Japan | Onglyza |
| Linagliptin | Europe, US | Trajentra, Tradjenta, Trazenta |
| Alogliptin | Europe, US, Japan | Vipidia, Nesina |
| Anagliptin | Japan | Suiny |
| Teneligliptin | Japan | Tenelia |
| Gemigliptin | Korea | Zemiglo |
| Omarigliptin | Japan | Marizev |
| Gosogliptin | Russia | Pfizer |
| Denagliptin | USA, Finland | Glaxo |
| Melogliptin | Europe, US, Japan | Glenmark |
| Trelagliptin | Europe, US | Takeda |
| Retagliptin | China | --------- |
| Evogliptin | Korea | Suganon |
| Carmegliptin | Switzerland | -------- |
| Plant Name/Natural Sources | Part of Plant/Bioactive Components Used | Medicinal/as Food Use | * DPP-IV Inhibition Activity %/IC50 | References |
|---|---|---|---|---|
| Azadirachta indica | Leaves | Arthritis, anxiety, trouble sleeping, anti-diabetes | 17.78% | [86] |
| Physalis angulata L. | Leaves | Peroxidation, cardiotonic, hypotensive, anti-diabetes | 13.94% | [87] |
| Aspergillus oryzae | tetrahydroxyisoquinoline derivative(WYK-1) | Used as food | IC50-6.98 μM | [88] |
| Berberis aristata | Bark | Arthritis, anxiety, trouble sleeping, anti-diabetes | 65% | [89] |
| Helichrysum rubicundum | Flower | Anti-lipid, peroxidation, cardiotonic, hypotensive, antidegenerative, anti-diabetes | 50.2% | [79] |
| Hippophaёrhamnoides | Leaves | Excessive menstruation and diarrhea | 80.5% | |
| Origanum vulgare | Leaves/ Flower | Regulate blood sugar & lipid, diarrhea, cold | 44.9% | |
| Rubuscaesius | Leaves | Antidiabetes, Blood Disorder, Respiratory Disorder, Piles | 72.7% | |
| Zea mays | Kernels | Anti-diabetes, Arthritis, anxiety, sleeping trouble, anti-diabetes | 22.2% | |
| Melilotus officinalis | Leaves | Hypertension, antioxidants prevent aging, arteriosclerosis | 40.5% | |
| Chamomillaerecutita | Leaves | Diarrhea and excessive menstruation | 36.6% | |
| Hypericumperforatum | Seed | Hypertension, antioxidants prevent aging | 47.7% | |
| Castanospermum australe | Seed | Anti-diabetes, Arthritis, anxiety, trouble sleeping, antidiabetic | 68.0 | [20] |
| Citrus sinensis | Fruit Peel | Hyperglycimia, anti-inflammation& antioxidant | Not mention | [90] |
| Amaranthus hypochondriacus | Seed protein | Excessive menstruation and diarrhea | 50% | [60] |
| Phalariscanariensis | Seed | Hypertension, antioxidants preventing aging and arteriosclerosis | 43.5% | [91] |
| Trigonella foenum | Seed | Reproductive or sin problems, antidiabetic and ulcers. | 72.6 ± 0.8 | [92] |
| Withania somnifera | Root powder | Anxiety, sleeping problem, antioxidant, anti-diabetic | 88.35 ± 0.8 | |
| Ocimum sanctum | Leaves | Anti-cancer, heptoprotective Anti-diabetic, anti-microbial, and cardio-protective | 66.81 ± 0.05 | [83] |
| Momardica charantia | Fruits | Respiratory and blood disorder, Anti-diabetic, piles, antioxidant. | 53.25 ± 0.04 | |
| LPVPQ Peptide & IPM | Milk | Antioxidant, Anti-microbial, ACE inhibitor | IC50 43.8 μΜ & 69.3 μM | [30] |
| Aloe barbadensis | Leaves | Anti-microbial, anti-diabetic, anti-ulcer, hepatoprotactive, Anti-cancer. | IC50 2.71 mg/mL | [55] |
| Terminalia arjuna | Bark | Cardiotonic, anti-diabetic, Anti-dysenteric, anti-pyretic | 83.39% | [93] |
| Commiphora mukul | Gum-resin | Anti-inflammation, antidiabetic | 92.97% | |
| Gymnema sylvestre | Leaves | Anti-diabetic, lower blood pressure and cholesterol | 16.98% | |
| Morinda citrifolia | Fruits | Reduce blood pressure, anti-diabetic, anti-depression, anxiety | 24.64% | |
| Emblica officinalis | Fruits | Antioxidant, anti-inflammation, | 85.95% | |
| Arachishypogaea | Seed | Anti-lipid, peroxidation, cardiotonic, hypotensive, antidegenerative, anti-diabetes | 51% | [94] |
| Senna nigricans | Leaves | Skin irritations, anti-diabetes, ulcers, | 57% | |
| Solanum incanum | Fruit | Arthritis, anxiety, trouble sleeping, anti-diabetes | 68.1% | |
| Ziziphusmauritiana | Root | Anti-cancer, anti-diabetic, anti-microbial, hepatoprotective, cardioprotective, antiemetic | 56.6% | |
| FSD & WSG | Barbel muscle protein | Used as food and ornamental | IC 50 1.96 mg/mL | [95] |
| Abelmoschus esculentus | Fruits | Anti-spasmodic, diuretic, anti-diabetic, | ------ | [96] |
| Mangifera indica | Leaves | Antioxidant, cardiotonic, hypotension, anti-diabetic | 68.24% | [80,97] |
| Origanum vulgare | Leaves | Hypotension, anti-degenerative, anti-diabetic. | 44.9% | [80] |
| Menthapiperita | Leaves | Anti-diabetic, anti-cancer, hepatoprotective, anti-microbial, antiemetic | 38.2% | |
| Lagerstroemia loudonii | Leaves | Hypertension, antioxidants | 60.22% | |
| LPVP & MPVQA | Camel milk | Used as food | IC50- 87.0 μM & 93.3 μM | [98] |
| Camellia sinensis | Leaves | Hypertension, antioxidants prevent aging, arteriosclerosis | 50.47% | [99] |
| Caesapinia sappan | Heartwood | Antioxidant, anti-inflammatory, hepatoprotective, cytotoxic & hypoglycemia inhibition activity | 84.25% | [100] |
| Cinchona officinalis | Stem bark | Used as blood vessel disorder, Increase juice, increase production of digest | 62.95% | |
| Elephantopusscaber | Roots | Treatment for pain, edema, fever and cold cough. | 48.17% | |
| Muntingiacalabura | Leaves | Antispasmodic, headache, cold cough | 74.12% | |
| Foeniculum vulgare | Seeds | Treatment for respiratory, blood pressure, digestive problem diuretic. | 46.15% | |
| Morus nigra | Stem bark | Anti-bacterial, anti-diabetic, hypertension | 51% | |
| Phyllanthns niuri | Aerial parts | Treatment for liver disease, anti-cancer, anti-diabetic, anti-hypertensive | 70.48% | |
| Psidium guajava | Leaves | Treatment for diarrhea, dysentery, pain relief, Anti-diabetic, hypertension. | 66.11% | |
| Rheum palmatum | Roots | Fever and edema, anti-inflammatory | 72.67% | |
| Vernania amygdalina | Leaves | Anti-malaria, anti-diabetic kidney disease, anti-hypertensive | 50.2% | |
| Prosopis cineraria | Pod | used for treatment of asthma, bronchitis skin problem | 64.8% | [101] |
| Garlic bulb | Bulb | Antioxidant, anti-hypertensive, anti-inflammatory, anti-diabetic. | IC50- 70.88 μg/ml | [102] |
| subtilisin-flavourzyme | enebriomolitor hydrolysates | Antioxidant, anti-hypertensive. | IC50- 2.89mg/mL | [103] |
| Castanea mollissima Blume | Chestnut Inner skin | Nuts used as food and antioxidant. | IC50 1.14 μg/ml | [104] |
| Caprosaper | Protein hydrolysate IPVDM & IPV | Ornamental used only | IC50- 21.72 μM & 5.61 μM | [105] |
| Arthrospira platensis (spirulina) | C-phycocyanin (C-PC) and allophycocyanin (APC) | Anti-apoptotic, hypolipidemic, anti-inflammatory | 95.8% | [106] |
| α-lactalbumin-rich whey protein | LDQWLCEKL | Used as source of food | 131 μM | [107] |
| Hibiscus rosa-sinensis | Leaves | Detoxifier, anti-fertility, anti-cancer, anti-hypertension, and cardio-protective effect | 60% | [108] |
| Aloe vera | dipyrrole derivative | Used as anti-diabetic, skin problem, | IC50 – 8.59 nM | [109] |
| Quercetin & Coumarin | Flavonoids (bioactive compounds) | Anti-hyperglycemic and antioxidant | IC50-4.02 &54.8 nMol/ml | [84] |
| Moringa oleiferaLam. | Leaves | Anti-inflammatory, anti-diabetic, hepatoprotective | IC50- 798 nM | [110] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, A.-K.; Yadav, D.; Sharma, N.; Jin, J.-O. Dipeptidyl Peptidase (DPP)-IV Inhibitors with Antioxidant Potential Isolated from Natural Sources: A Novel Approach for the Management of Diabetes. Pharmaceuticals 2021, 14, 586. https://doi.org/10.3390/ph14060586
Singh A-K, Yadav D, Sharma N, Jin J-O. Dipeptidyl Peptidase (DPP)-IV Inhibitors with Antioxidant Potential Isolated from Natural Sources: A Novel Approach for the Management of Diabetes. Pharmaceuticals. 2021; 14(6):586. https://doi.org/10.3390/ph14060586
Chicago/Turabian StyleSingh, Anand-Krishna, Dhananjay Yadav, Neha Sharma, and Jun-O Jin. 2021. "Dipeptidyl Peptidase (DPP)-IV Inhibitors with Antioxidant Potential Isolated from Natural Sources: A Novel Approach for the Management of Diabetes" Pharmaceuticals 14, no. 6: 586. https://doi.org/10.3390/ph14060586
APA StyleSingh, A. -K., Yadav, D., Sharma, N., & Jin, J. -O. (2021). Dipeptidyl Peptidase (DPP)-IV Inhibitors with Antioxidant Potential Isolated from Natural Sources: A Novel Approach for the Management of Diabetes. Pharmaceuticals, 14(6), 586. https://doi.org/10.3390/ph14060586