

MK2 Inhibitors as a Potential Crohn’s Disease Treatment Approach for Regulating MMP Expression, Cleavage of Checkpoint Molecules and T Cell Activity


Abstract
:1. Introduction
2. Results
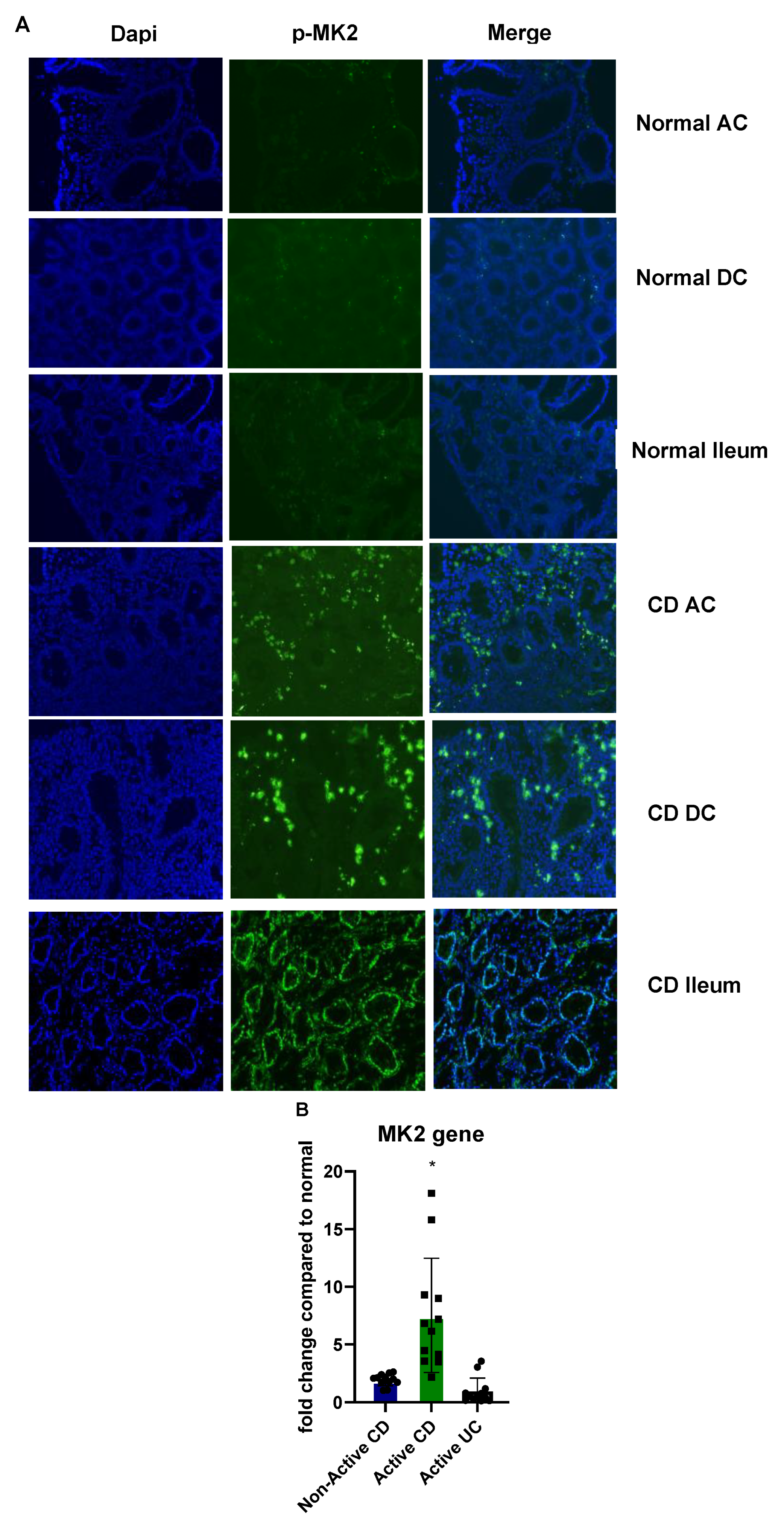
2.1. MK2 Expression and Activity Are Increased in CD
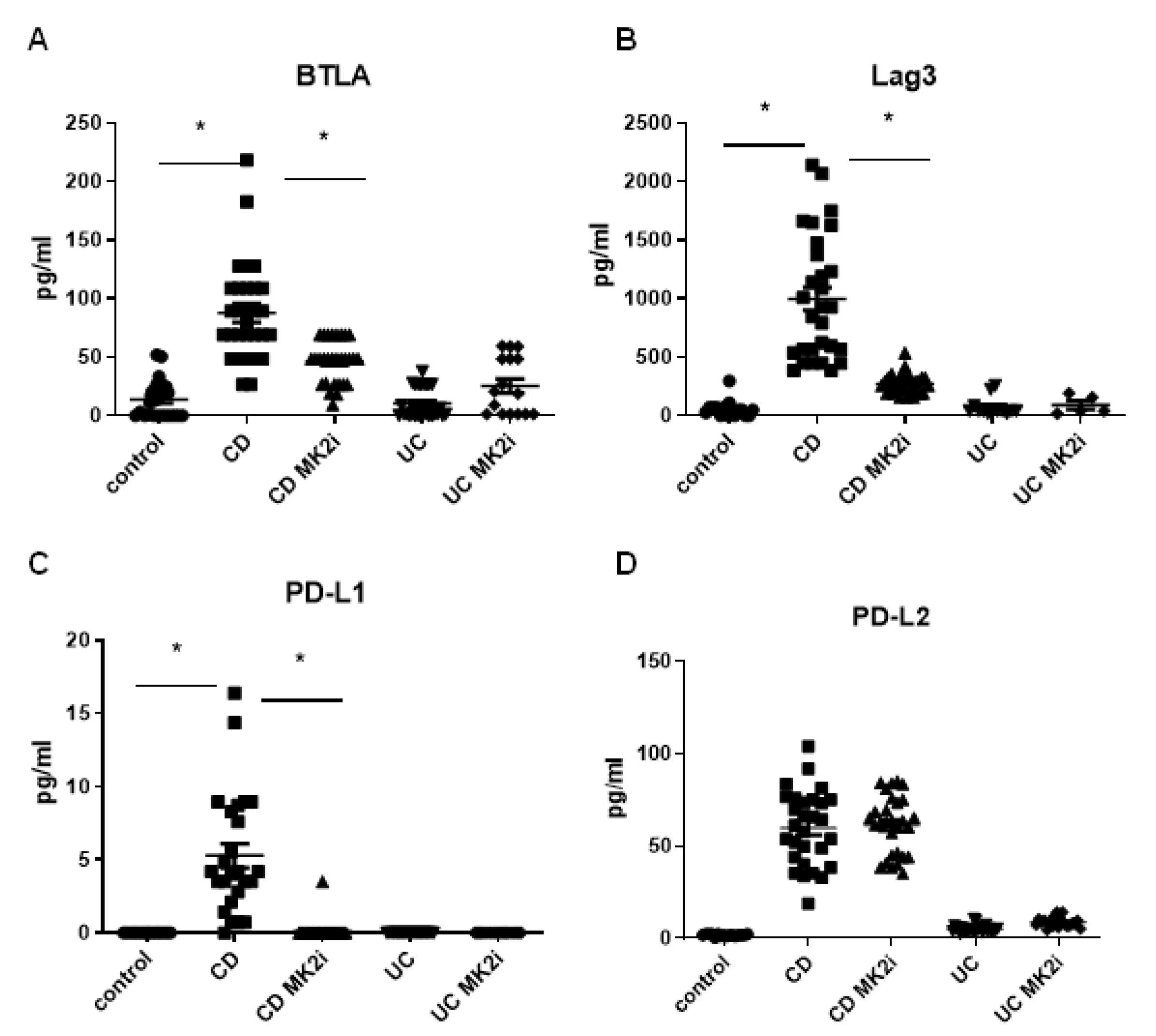
2.2. Checkpoint Molecules Are Cleaved from CD Tissues in an MK2-Dependent Manner
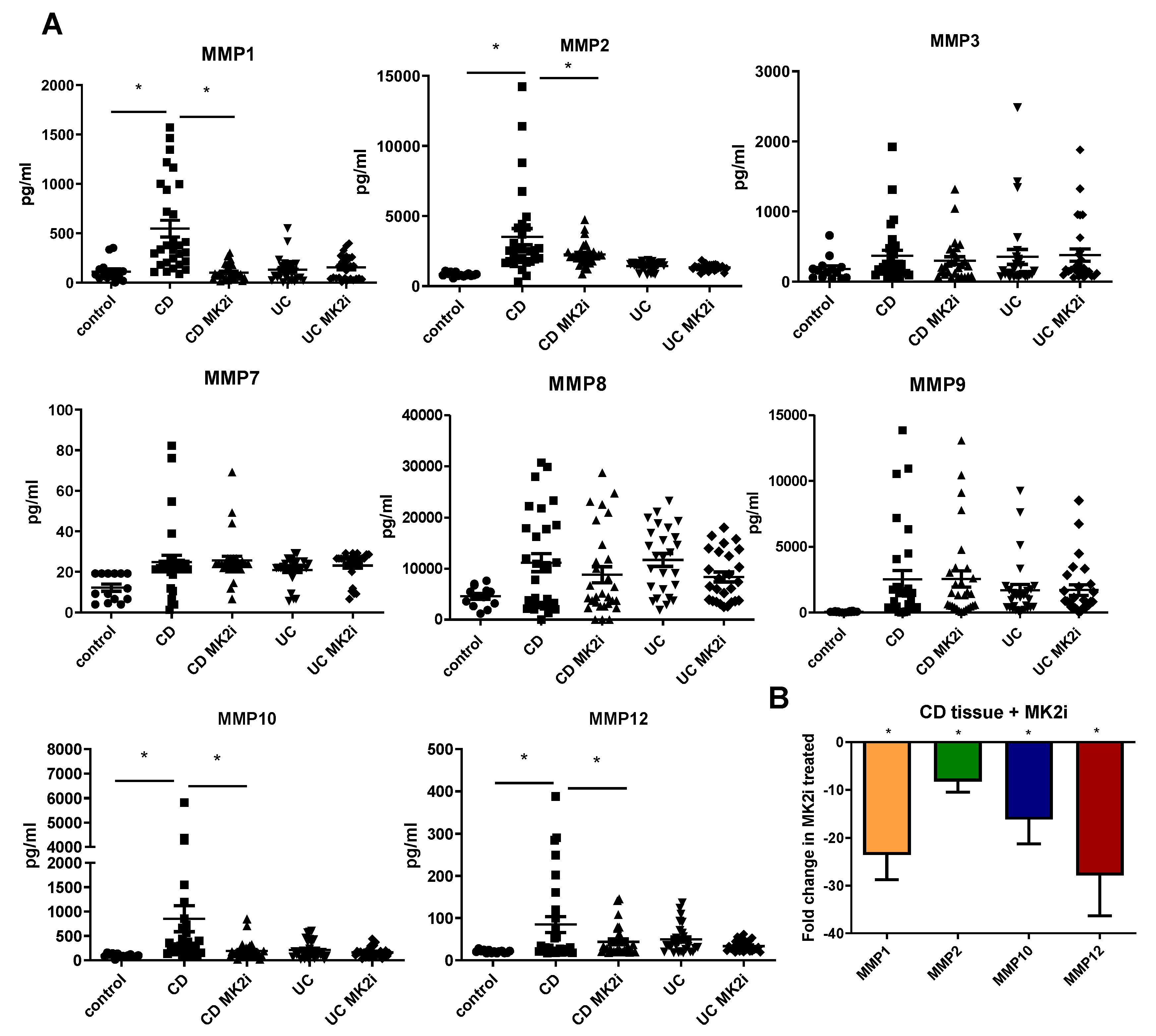
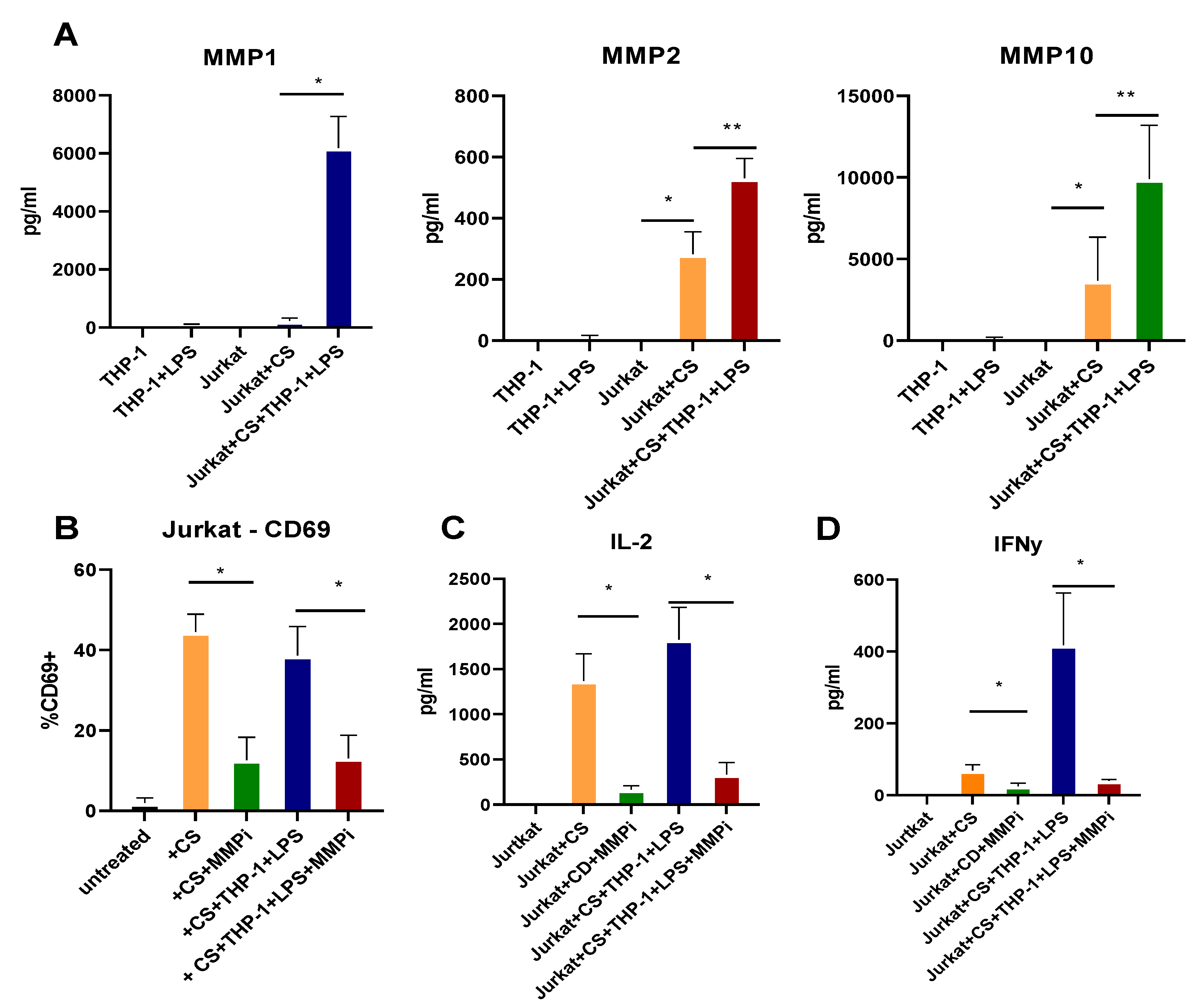
2.3. MMP Production Is Increased by CD Tissues in an MK2-Dependent Manner
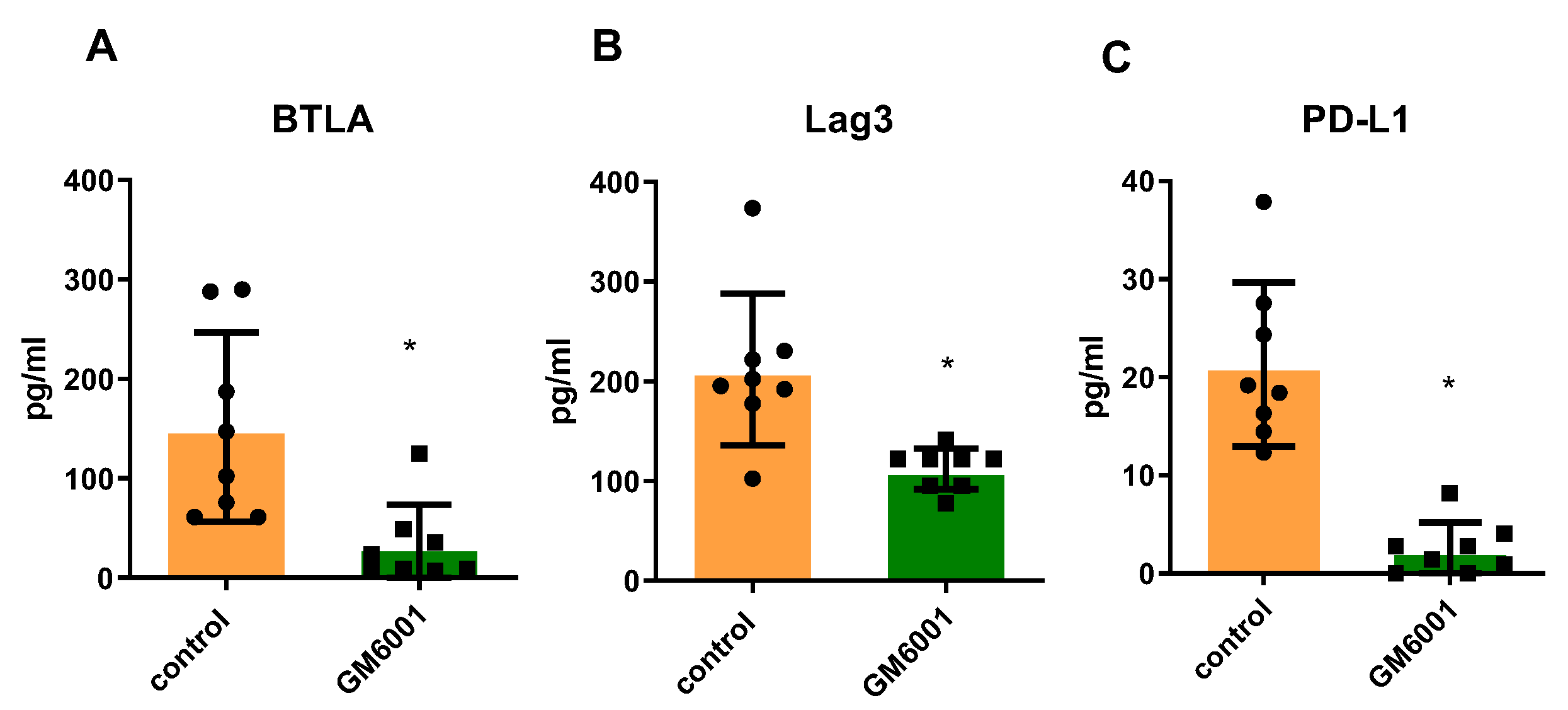
2.4. Checkpoint Molecules Are Cleaved from CD Tissues in an MMP-Dependent Manner
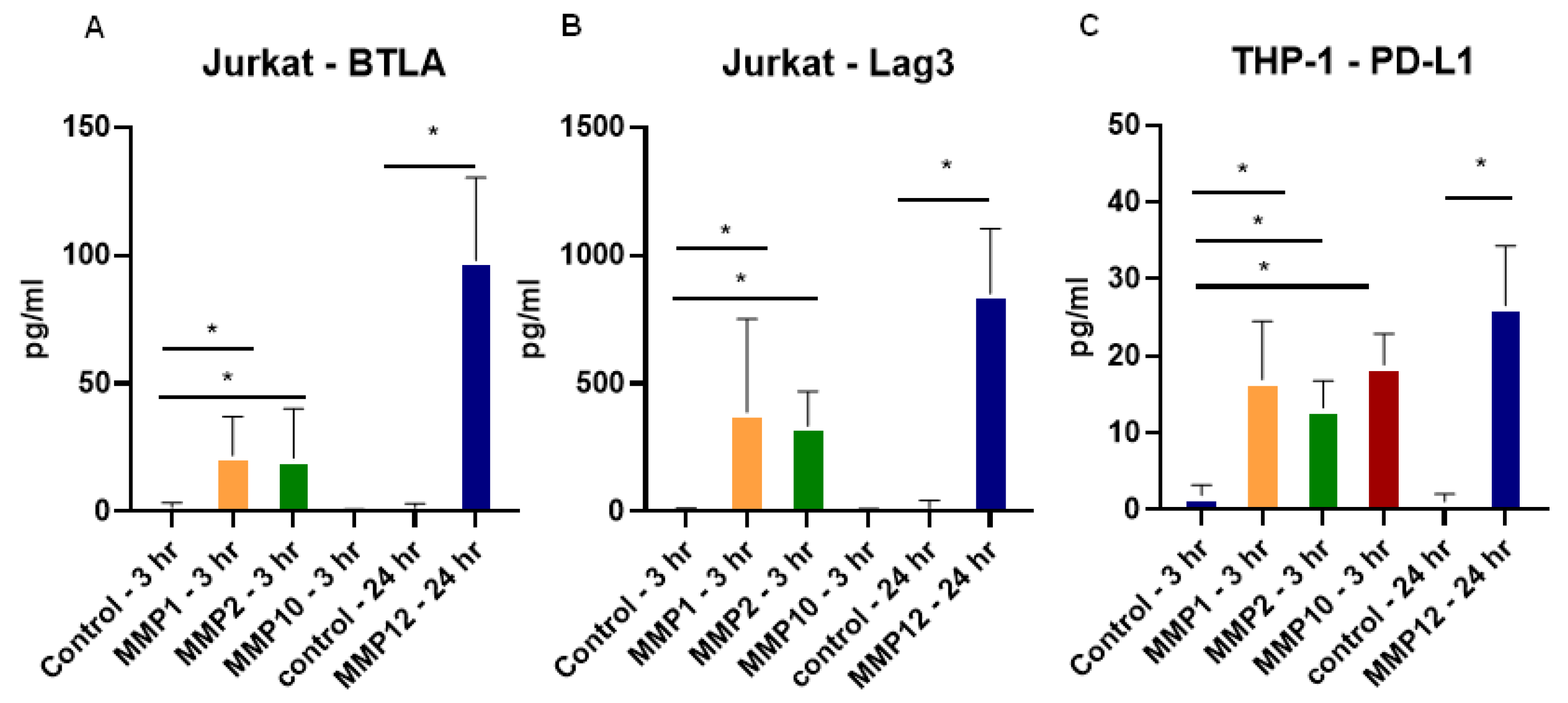
2.5. Recombinant MMPs Cleave Checkpoint Molecules from T Cells and Monocytes
2.6. T Cell Activation and Cytokine Production Are Dependent on MMPs
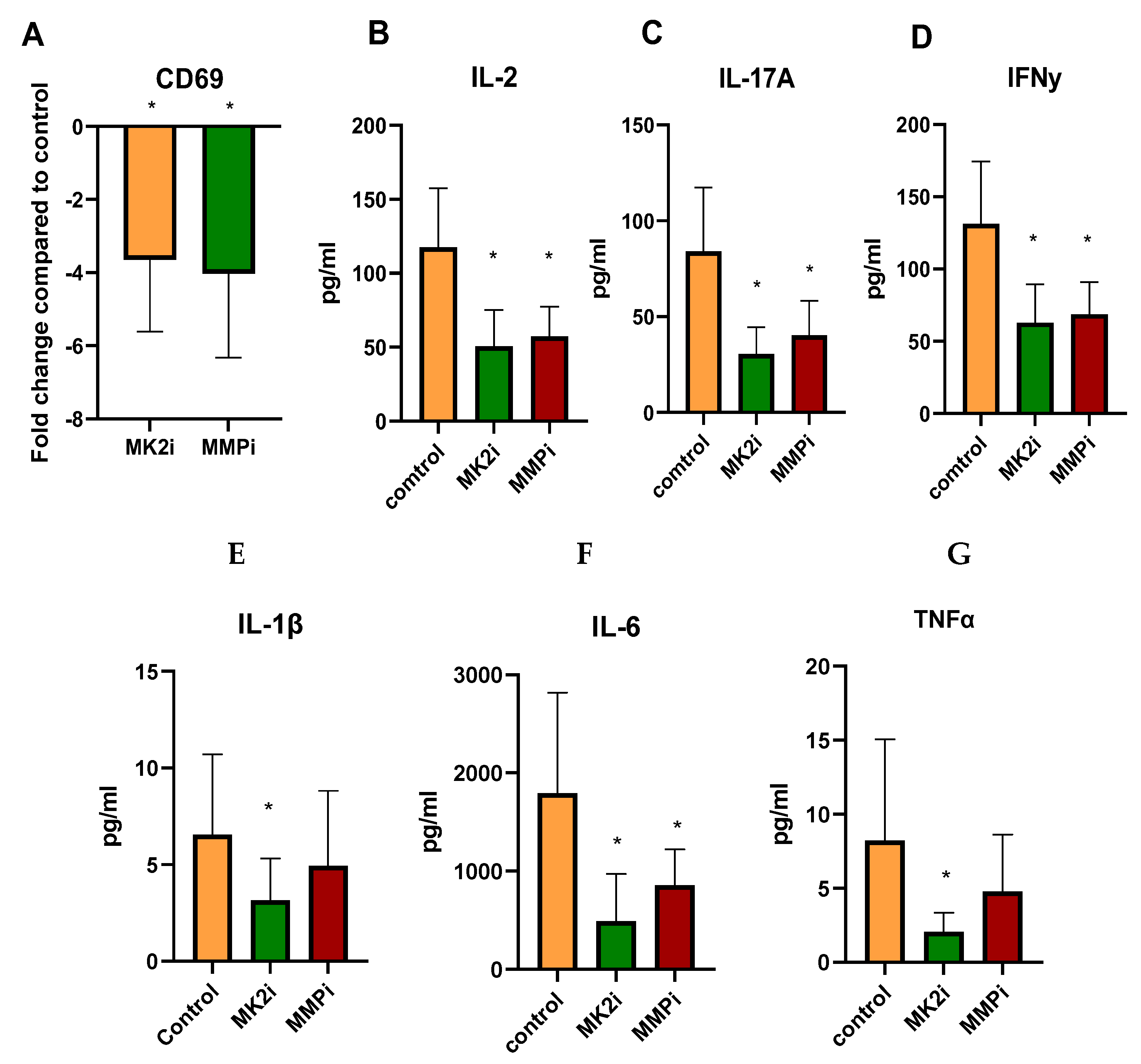
2.7. CD Tissues Treated with MK2i or MMPi Have Decreased T Cell Activation Markers
3. Discussion
4. Materials and Methods
4.1. Tissue Collection and Processing
4.2. Cell Lines
4.3. Immunofluorescence
4.4. Multiplex Arrays
4.5. Quantitative Real Time PCR
4.6. MMP Cleavage Assays
4.7. Jurkat and THP-1 Co-Culture Assays
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2017, 390, 2769–2778. [Google Scholar] [CrossRef]
- Imam, T.; Park, S.; Kaplan, M.H.; Olson, M.R. Effector T Helper Cell Subsets in Inflammatory Bowel Diseases. Front. Immunol. 2018, 9, 1212. [Google Scholar] [CrossRef]
- Chen, M.L.; Sundrud, M.S. Cytokine Networks and T-Cell Subsets in Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2016, 22, 1157–1167. [Google Scholar] [CrossRef] [Green Version]
- Maruhashi, T.; Sugiura, D.; Okazaki, I.-M.; Shimizu, K.; Maeda, T.K.; Ikubo, J.; Yoshikawa, H.; Maenaka, K.; Ishimaru, N.; Kosako, H.; et al. Binding of LAG-3 to stable peptide-MHC class II limits T cell function and suppresses autoimmunity and anti-cancer immunity. Immunity 2022, 55, 912–924.e8. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-T.; Workman, C.J.; Flies, D.; Pan, X.; Marson, A.L.; Zhou, G.; Hipkiss, E.L.; Ravi, S.; Kowalski, J.; Levitsky, H.I.; et al. Role of LAG-3 in Regulatory T Cells. Immunity 2004, 21, 503–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauché, D.; Joyce-Shaikh, B.; Jain, R.; Grein, J.; Ku, K.S.; Blumenschein, W.M.; Ganal-Vonarburg, S.C.; Wilson, D.C.; McClanahan, T.K.; Malefyt, R.D.W.; et al. LAG3+ Regulatory T Cells Restrain Interleukin-23-Producing CX3CR1+ Gut-Resident Macrophages during Group 3 Innate Lymphoid Cell-Driven Colitis. Immunity 2018, 49, 342–352.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Zheng, Y.; Mao, R.; Su, Z.; Zhang, J. BTLA/HVEM Signaling: Milestones in Research and Role in Chronic Hepatitis B Virus Infection. Front. Immunol. 2019, 10, 617. [Google Scholar] [CrossRef]
- Steinberg, M.W.; Turovskaya, O.; Shaikh, R.B.; Kim, G.; McCole, D.F.; Pfeffer, K.; Murphy, K.M.; Ware, C.F.; Kronenberg, M. A crucial role for HVEM and BTLA in preventing intestinal inflammation. J. Exp. Med. 2008, 205, 1463–1476. [Google Scholar] [CrossRef] [Green Version]
- Chulkina, M.; Beswick, E.J.; Pinchuk, I.V. Role of PD-L1 in Gut Mucosa Tolerance and Chronic Inflammation. Int. J. Mol. Sci. 2020, 21, 9165. [Google Scholar] [CrossRef]
- Aguirre, J.E.; Beswick, E.J.; Grim, C.; Uribe, G.; Tafoya, M.; Chacon Palma, G.; Samedi, V.; McKee, R.; Villeger, R.; Fofanov, Y.; et al. Matrix metalloproteinases cleave membrane-bound PD-L1 on CD90+ (myo-)fibroblasts in Crohn’s disease and regulate Th1/Th17 cell responses. Int. Immunol. 2020, 32, 57–68. [Google Scholar] [CrossRef]
- Rohani, M.G.; Parks, W.C. Matrix remodeling by MMPs during wound repair. Matrix Biol. 2015, 44, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Muller-Quernheim, J. MMPs are regulatory enzymes in pathways of inflammatory disorders, tissue injury, malignancies and remodelling of the lung. Eur. Respir. J. 2011, 38, 12–14. [Google Scholar] [CrossRef]
- O’Sullivan, S.; Gilmer, J.F.; Medina, C. Matrix Metalloproteinases in Inflammatory Bowel Disease: An Update. Mediat. Inflamm. 2015, 2015, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, G.; Saermark, T.; Kirkegaard, T.; Brynskov, J. Spontaneous and cytokine induced expression and activity of matrix metalloproteinases in human colonic epithelium. Clin. Exp. Immunol. 2008, 155, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Schuppan, D.; Freitag, T. Fistulising Crohn’s disease: MMPs gone awry. Gut 2004, 53, 622–624. [Google Scholar] [CrossRef] [Green Version]
- Rogler, G.; Hausmann, M. Factors Promoting Development of Fibrosis in Crohn’s Disease. Front. Med. 2017, 4, 96. [Google Scholar] [CrossRef]
- Liu, X.; Wu, T.; Chi, P. Inhibition of MK2 shows promise for preventing postoperative ileus in mice. J. Surg. Res. 2013, 185, 102–112. [Google Scholar] [CrossRef]
- Ray, A.L.; Castillo, E.F.; Morris, K.T.; Nofchissey, R.A.; Weston, L.L.; Samedi, V.G.; Hanson, J.A.; Gaestel, M.; Pinchuk, I.V.; Beswick, E.J. Blockade of MK2 is protective in inflammation-associated colorectal cancer development. Int. J. Cancer 2015, 138, 770–775. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.Y.; Yuece, B.; MH, C.; Lv, S.; CJ, C.; Ochs, S.; Sibaev, A.; Deindl, E.; Schaefer, C.; Storr, M. Inhibition of p38/Mk2 signaling pathway improves the anti-inflammatory effect of WIN55 on mouse experimental colitis. Lab. Investig. 2013, 93, 322–333. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Jiang, J.; Liu, J.; Xu, L.; Duan, S.; Sun, L.; Zhao, W.; Qian, F. MK2 Is Required for Neutrophil-Derived ROS Production and Inflammatory Bowel Disease. Front. Med. 2020, 7, 207. [Google Scholar] [CrossRef]
- Dezutter-Dambuyant, C.; Durand, I.; Alberti, L.; Bendriss-Vermare, N.; Valladeau-Guilemond, J.; Duc, A.; Magron, A.; Morel, A.-P.; Sisirak, V.; Rodriguez, C.; et al. A novel regulation of PD-1 ligands on mesenchymal stromal cells through MMP-mediated proteolytic cleavage. OncoImmunology 2015, 5, e1091146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tindemans, I.; Joosse, M.E.; Samsom, J.N. Dissecting the Heterogeneity in T-Cell Mediated Inflammation in IBD. Cells 2020, 9, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinchuk, I.V.; Saada, J.I.; Beswick, E.J.; Boya, G.; Qiu, S.M.; Mifflin, R.C.; Raju, G.S.; Reyes, V.E.; Powell, D.W. PD-1 ligand expression by human colonic myofibroblasts/fibroblasts regulates CD4+ T-cell activity. Gastroenterology 2008, 135, 1228–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faleiro, R.; Liu, J.; Karunarathne, D.; Edmundson, A.; Winterford, C.; Nguyen, T.H.; Simms, L.A.; Radford-Smith, G.; Wykes, M. Crohn’s disease is facilitated by a disturbance of programmed death-1 ligand 2 on blood dendritic cells. Clin. Transl. Immunol. 2019, 8, e01071. [Google Scholar] [CrossRef]
- Nakazawa, A.; Dotan, I.; Brimnes, J.; Allez, M.; Shao, L.; Tsushima, F.; Azuma, M.; Mayer, L. The expression and function of costimulatory molecules B7H and B7-H1 on colonic epithelial cells. Gastroenterology 2004, 126, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Macon-Lemaitre, L.; Triebel, F. The negative regulatory function of the lymphocyte-activation gene-3 co-receptor (CD223) on human T cells. Immunology 2005, 115, 170–178. [Google Scholar] [CrossRef]
- Wang, M.; Du, Q.; Jin, J.; Wei, Y.; Lu, Y.; Li, Q. LAG3 and its emerging role in cancer immunotherapy. Clin. Transl. Med. 2021, 11, e365. [Google Scholar] [CrossRef]
- Lange, A.; Sundén-Cullberg, J.; Magnuson, A.; Hultgren, O. Soluble B and T Lymphocyte Attenuator Correlates to Disease Severity in Sepsis and High Levels Are Associated with an Increased Risk of Mortality. PLoS ONE 2017, 12, e0169176. [Google Scholar] [CrossRef] [Green Version]
- Das, K.; Prasad, R.; Ansari, S.A.; Roy, A.; Mukherjee, A.; Sen, P. Matrix metalloproteinase-2: A key regulator in coagulation proteases mediated human breast cancer progression through autocrine signaling. Biomed. Pharmacother. 2018, 105, 395–406. [Google Scholar] [CrossRef]
- Xu, L.; Bergan, R.C. Genistein inhibits matrix metalloproteinase type 2 activation and prostate cancer cell invasion by blocking the transforming growth factor beta-mediated activation of mitogen-activated protein kinase-activated protein kinase 2-27-kDa heat shock protein pathway. Mol. Pharmacol. 2006, 70, 869–877. [Google Scholar]
- Kumar, B.; Koul, S.; Petersen, J.; Khandrika, L.; Hwa, J.S.; Meacham, R.B.; Wilson, S.; Koul, H.K. p38 Mitogen-Activated Protein Kinase–Driven MAPKAPK2 Regulates Invasion of Bladder Cancer by Modulation of MMP-2 and MMP-9 Activity. Cancer Res. 2010, 70, 832–841. [Google Scholar] [CrossRef] [PubMed]
- Derkacz, A.; Olczyk, P.; Olczyk, K.; Komosinska-Vassev, K. The Role of Extracellular Matrix Components in Inflammatory Bowel Diseases. J. Clin. Med. 2021, 10, 1122. [Google Scholar] [CrossRef] [PubMed]
- Marônek, M.; Marafini, I.; Gardlík, /.R.; Link, R.; Troncone, E.; Monteleone, G. Metalloproteinases in Inflammatory Bowel Diseases. J. Inflamm. Res. 2021, 14, 1029–1041. [Google Scholar] [CrossRef]
- Schreiber, S.; Siegel, C.A.; Friedenberg, K.A.; Younes, Z.H.; Seidler, U.; Bhandari, B.R.; Wang, K.; Wendt, E.; McKevitt, M.; Zhao, S.; et al. A Phase 2, Randomized, Placebo-Controlled Study Evaluating Matrix Metalloproteinase-9 Inhibitor, Andecaliximab, in Patients With Moderately to Severely Active Crohn’s Disease. J. Crohn’s Colitis 2018, 12, 1014–1020. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.; Hellriegel, E.T.; Hope, H.R.; Burt, D.; Monahan, J.B. Safety, Tolerability, Pharmacokinetics, and Pharmacodynamics of the MK2 Inhibitor ATI-450 in Healthy Subjects: A Placebo-Controlled, Randomized Phase 1 Study. Clin. Pharmacol. Adv. Appl. 2021, 13, 123–134. [Google Scholar] [CrossRef]
- Brown, D.I.; Cooley, B.C.; Quintana, M.T.; Lander, C.; Willis, M.S. Nebulized Delivery of the MAPKAP Kinase 2 Peptide Inhibitor MMI-0100 Protects Against Ischemia-Induced Systolic Dysfunction. Int. J. Pept. Res. Ther. 2016, 22, 317–324. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal | UC | CD | |
|---|---|---|---|
| Ileum | 6 | 0 | 10 |
| AC/transverse | 6 | 7 | 6 |
| DC/sigmoid | 6 | 6 | 3 |
| Rectum | 5 | 5 | 6 |
| Mixed | 0 | 9 | 5 |
| Total | 23 | 27 | 29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lebish, E.J.; Morgan, N.J.; Valentine, J.F.; Beswick, E.J. MK2 Inhibitors as a Potential Crohn’s Disease Treatment Approach for Regulating MMP Expression, Cleavage of Checkpoint Molecules and T Cell Activity. Pharmaceuticals 2022, 15, 1508. https://doi.org/10.3390/ph15121508
Lebish EJ, Morgan NJ, Valentine JF, Beswick EJ. MK2 Inhibitors as a Potential Crohn’s Disease Treatment Approach for Regulating MMP Expression, Cleavage of Checkpoint Molecules and T Cell Activity. Pharmaceuticals. 2022; 15(12):1508. https://doi.org/10.3390/ph15121508
Chicago/Turabian StyleLebish, Eric J., Natalie J. Morgan, John F. Valentine, and Ellen J. Beswick. 2022. "MK2 Inhibitors as a Potential Crohn’s Disease Treatment Approach for Regulating MMP Expression, Cleavage of Checkpoint Molecules and T Cell Activity" Pharmaceuticals 15, no. 12: 1508. https://doi.org/10.3390/ph15121508
APA StyleLebish, E. J., Morgan, N. J., Valentine, J. F., & Beswick, E. J. (2022). MK2 Inhibitors as a Potential Crohn’s Disease Treatment Approach for Regulating MMP Expression, Cleavage of Checkpoint Molecules and T Cell Activity. Pharmaceuticals, 15(12), 1508. https://doi.org/10.3390/ph15121508