
In Vitro Anti-Influenza Virus Activity of Non-Polar Primula veris subsp. veris Extract

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Extraction of P. veris subsp. veris Flowering Parts
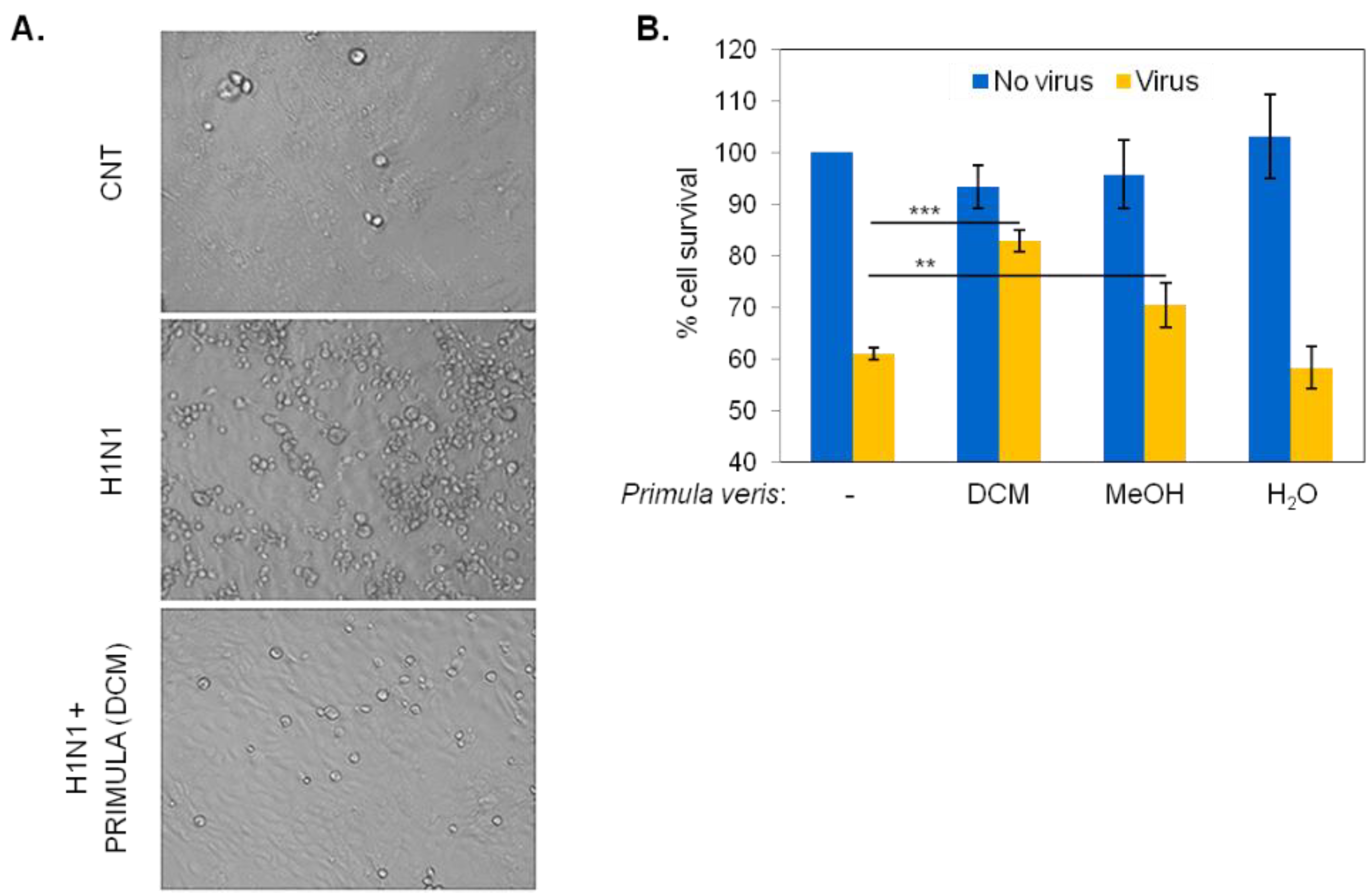
2.2. Identification of Primula veris Extracts with Virucidal Activity
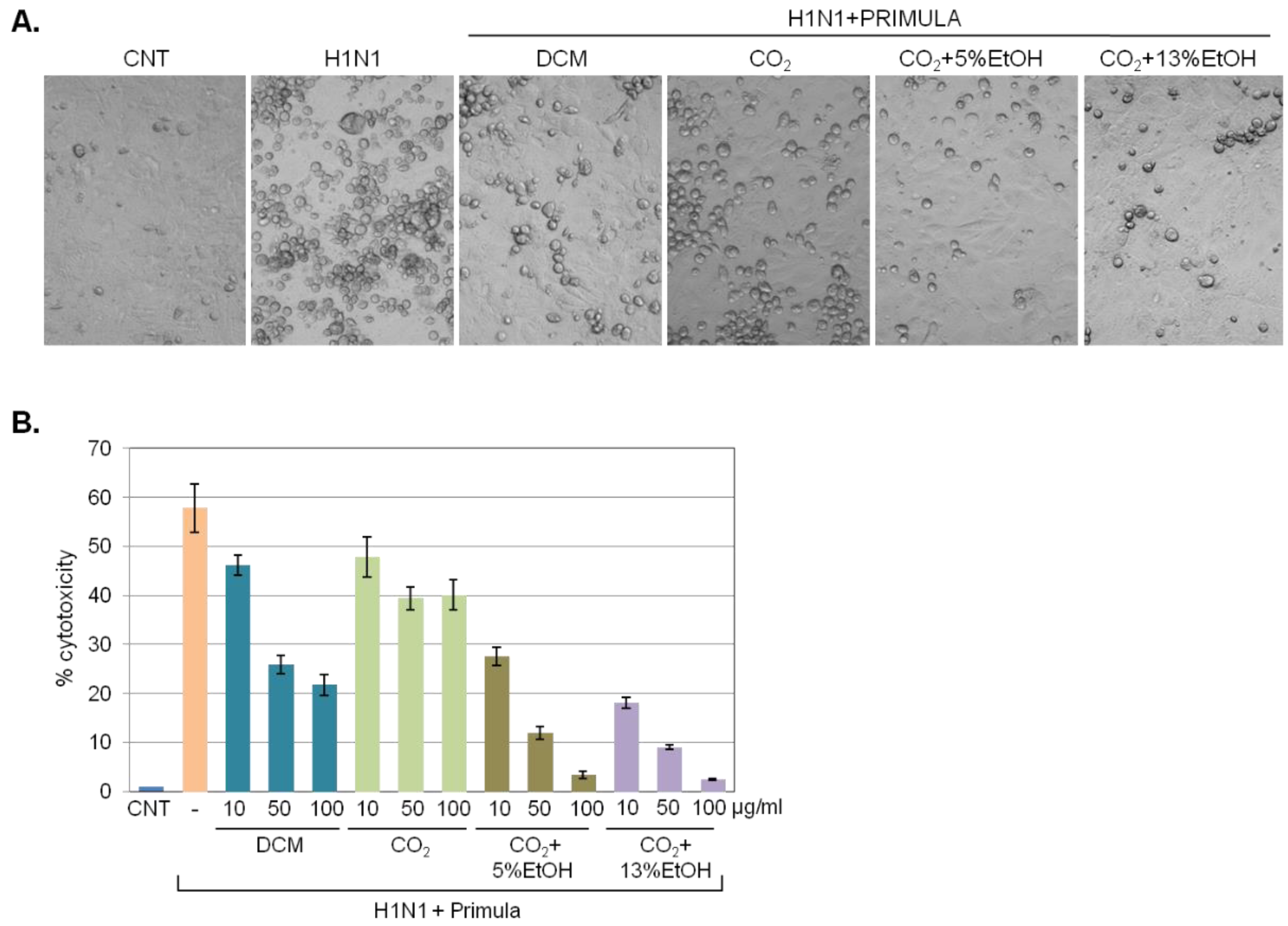
2.3. Prophylactic Effects of Primula Plant Extracts against Influenza Virus-Induced Cell Lysis
2.4. Therapeutic Effects of Primula Extract against Influenza Virus-Induced Cell Lysis
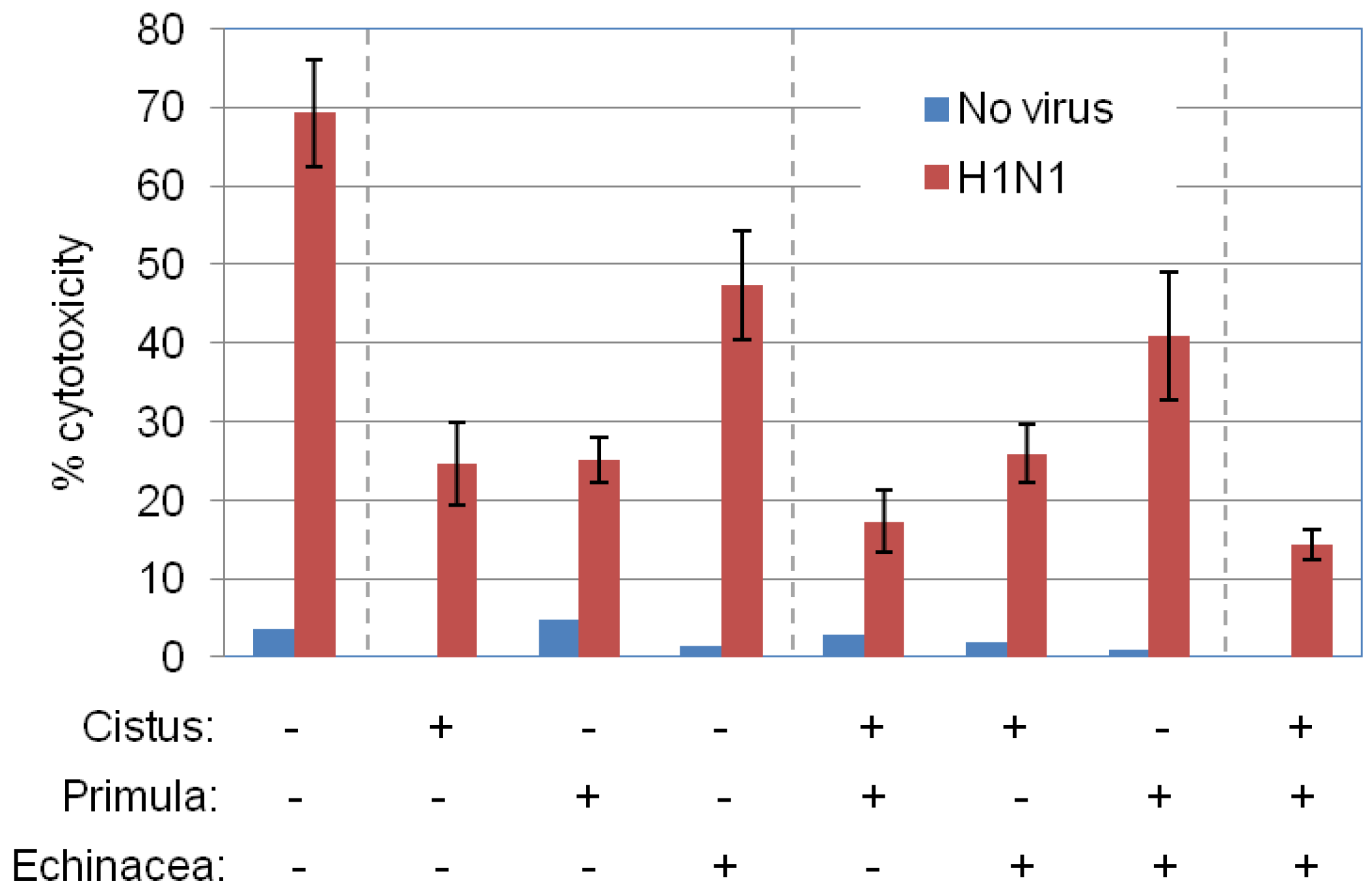
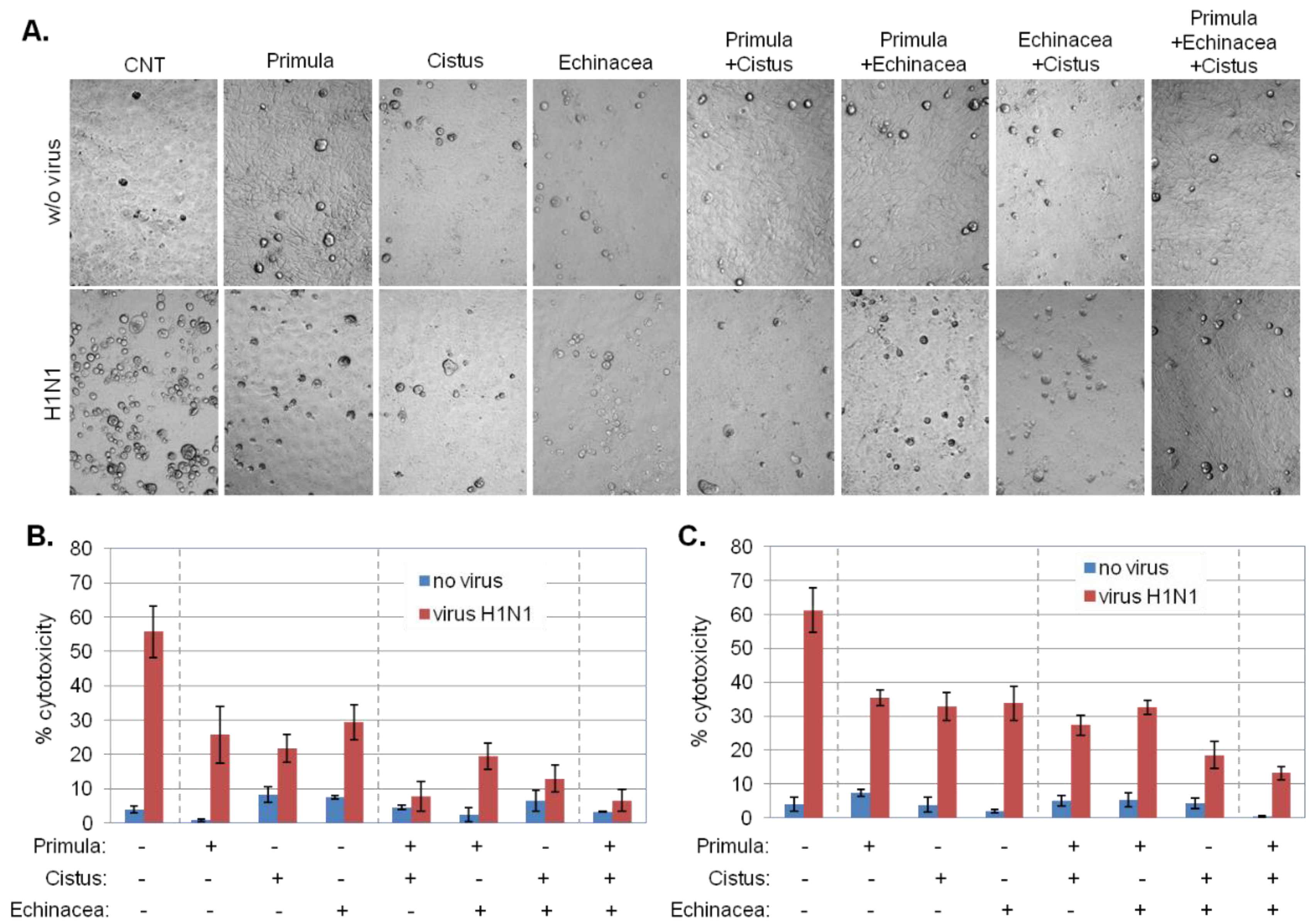
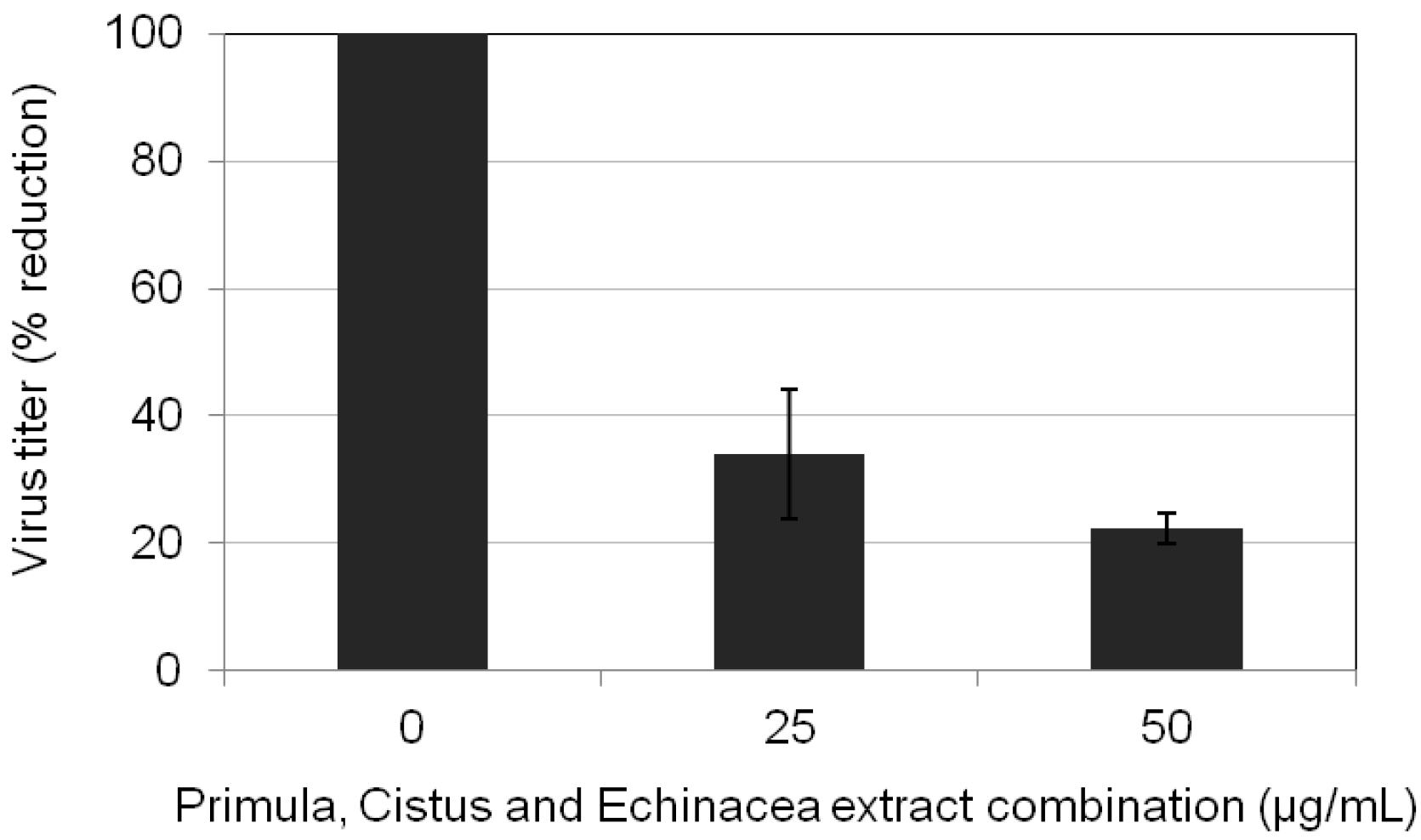
2.5. Primula, Cistus, and Echinacea Combination Treatment Results in Reduced Progeny Virus Production
2.6. Chemical Composition of Primula veris Non-Polar Extracts
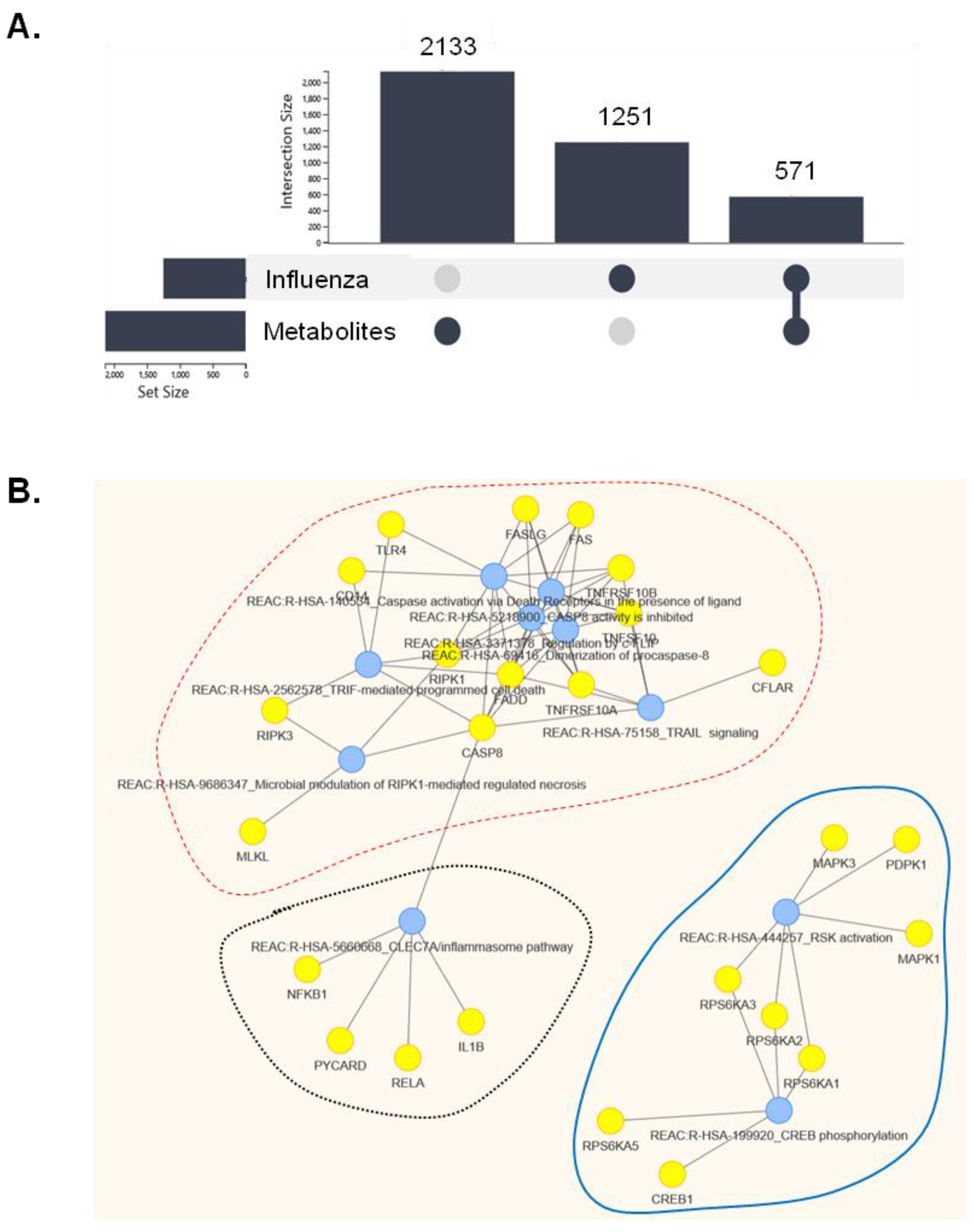
2.7. In Silico Prediction of Molecular Pathways That May Intersect the Cellular Effects of Primula veris Non-Polar Extract on Influenza Virus Infection
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plant Material
4.3. Extraction
4.4. Fractionation of P. veris subsp. veris Non-Polar Extract
4.5. TLC and HPLC Analysis
4.6. GC-MS and UPLC-HRMS Analysis
4.7. Cell Culture, Treatments and Cytotoxicity Assays
4.8. Virus and Infections
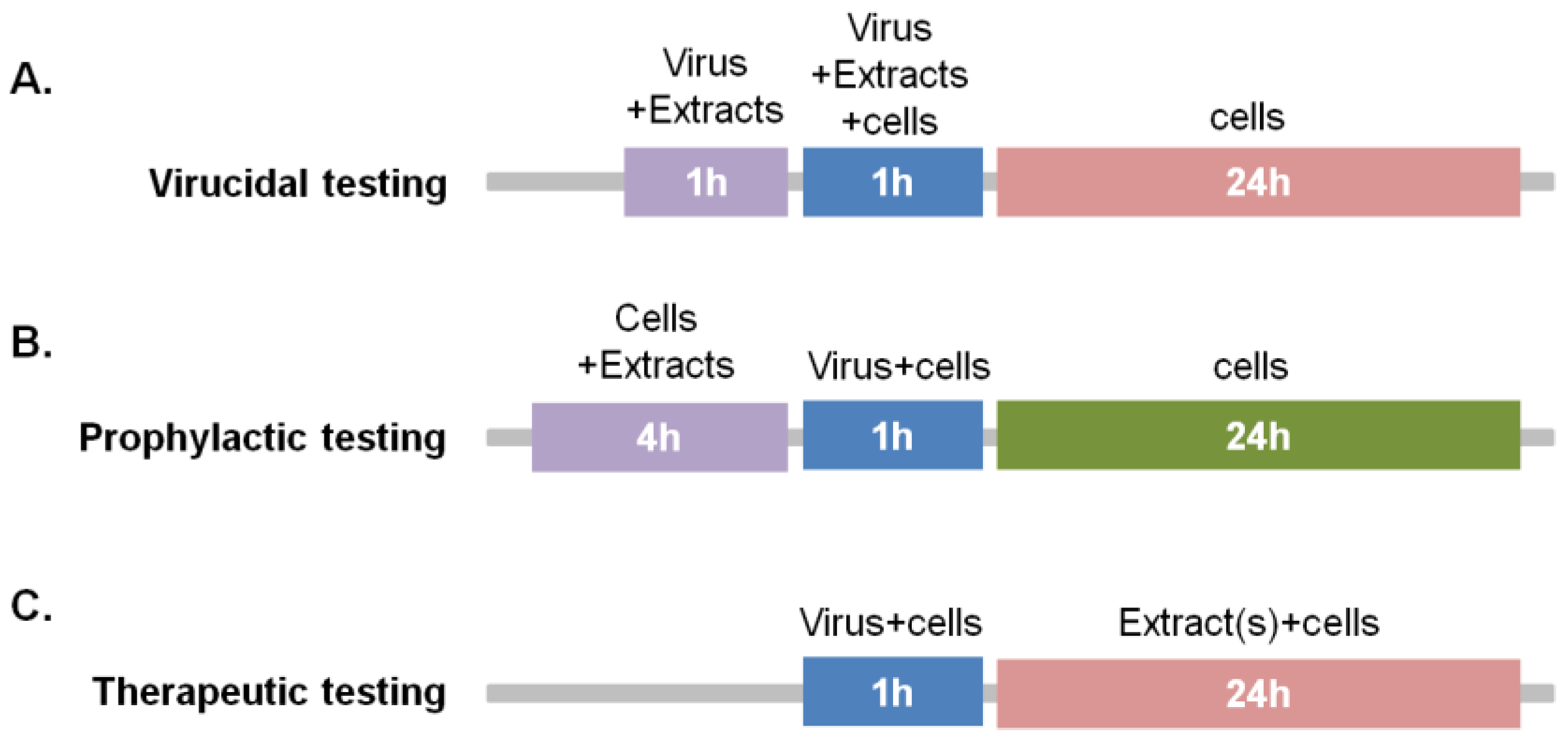
4.9. Time-of-Addition Assays
4.10. Plaque Titrations
4.11. In-Silico Predictions of Pathways That May Mediate the Anti-Viral Effects of Primula
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Keilman, L.J. Seasonal Influenza (Flu). Nurs. Clin. N. Am. 2019, 54, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Moscona, A. Neuraminidase inhibitors for influenza. N. Engl. J. Med. 2005, 353, 1363–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, A.; Bouvier, D.; Crepin, T.; McCarthy, A.A.; Hart, D.J.; Baudin, F.; Cusack, S.; Ruigrok, R.W. The cap-snatching endonuclease of influenza virus polymerase resides in the PA subunit. Nature 2009, 458, 914–918. [Google Scholar] [CrossRef] [PubMed]
- Fiore, A.E.; Fry, A.; Shay, D.; Gubareva, L.; Bresee, J.S.; Uyeki, T.M.; Centers for Disease Control and Prevention. Antiviral agents for the treatment and chemoprophylaxis of influenza: Recommendations of the Advisory Committee on Immunization Practices (ACIP). MMWR Recomm. Rep. 2011, 60, 1–24. [Google Scholar]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug. Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. Natural products as sources of new drugs over the period 1981–2002. J. Nat. Prod. 2003, 66, 1022–1037. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug. Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Glatthaar-Saalmuller, B.; Rauchhaus, U.; Rode, S.; Haunschild, J.; Saalmuller, A. Antiviral activity in vitro of two preparations of the herbal medicinal product Sinupret(R) against viruses causing respiratory infections. Phytomedicine 2011, 19, 1–7. [Google Scholar] [CrossRef]
- Pleschka, S.; Stein, M.; Schoop, R.; Hudson, J.B. Anti-viral properties and mode of action of standardized Echinacea purpurea extract against highly pathogenic avian influenza virus (H5N1, H7N7) and swine-origin H1N1 (S-OIV). Virol. J. 2009, 6, 197. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wang, X.; Li, C.; Khutsishvili, M.; Fayvush, G.; Atha, D.; Zhang, Y.; Borris, R.P. Unusual Flavones from Primula macrocalyx as Inhibitors of OAT1 and OAT3 and as Antifungal Agents against Candida rugosa. Sci. Rep. 2019, 9, 9230. [Google Scholar] [CrossRef] [Green Version]
- Seifert, S.; Kopeinig, B.; Bauer, R.; Pahl, A.; Haunschild, J. In vitro anti-inflammatory and anti-microbial activity of cowslip flowers (Primula veris L.). Planta Med. 2012, 78, PD101. [Google Scholar] [CrossRef]
- Tiwari, B.K. Ultrasound: A clean, green extraction technology. TrAC Trends Anal. Chem. 2015, 71, 100–109. [Google Scholar] [CrossRef]
- Herrero, M.; Mendiola, J.A.; Cifuentes, A.; Ibanez, E. Supercritical fluid extraction: Recent advances and applications. J. Chromatogr. A 2010, 1217, 2495–2511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vimalanathan, S.; Kang, L.; Amiguet, V.T.; Livesey, J.; Arnason, J.T.; Hudson, J. Echinacea purpurea. Aerial Parts Contain Multiple Antiviral Compounds. Pharm. Biol. 2005, 43, 740–745. [Google Scholar] [CrossRef] [Green Version]
- Ehrhardt, C.; Hrincius, E.R.; Korte, V.; Mazur, I.; Droebner, K.; Poetter, A.; Dreschers, S.; Schmolke, M.; Planz, O.; Ludwig, S. A polyphenol rich plant extract, CYSTUS052, exerts anti influenza virus activity in cell culture without toxic side effects or the tendency to induce viral resistance. Antivir. Res. 2007, 76, 38–47. [Google Scholar] [CrossRef]
- Budzianowski, J.; Morozowska, M.; Wesolowska, M. Lipophilic flavones of Primula veris L from field cultivation and in vitro cultures. Phytochemistry 2005, 66, 1033–1039. [Google Scholar] [CrossRef]
- Xynos, N.; Zervos, M.; Angelis, A.; Aligiannis, N.; Skaltsounis, A.-L. A single-step isolation of squalene from olive oil deodorizer distillates by using centrifugal partition chromatography. Sep. Sci. Technol. 2016, 51, 830–835. [Google Scholar] [CrossRef]
- Liakakou, A.; Angelis, A.; Papachristos, D.P.; Fokialakis, N.; Michaelakis, A.; Skaltsounis, L.A. Isolation of Volatile Compounds with Repellent Properties against Aedes albopictus (Diptera: Culicidae) Using CPC Technology. Molecules 2021, 26, 3072. [Google Scholar] [CrossRef]
- Kosenkova, Y.S.; Polovinka, M.P.; Komarova, N.I.; Korchagina, D.V.; Morozov, S.V.; Vyalkov, A.I.; Kurochkina, N.Y.; Cheremushkina, V.A.; Salakhutdinov, N.F. Fatty-acid composition and secondary metabolites from slightly polar extracts of the aerial part of Primula macrocalyx. Chem. Nat. Compd. 2008, 44, 564–568. [Google Scholar] [CrossRef]
- Budzianowski, J.; Wollenweber, E. Rare Flavones from the Glandular Leaf Exudate of the Oxlip, Primula Elatior L. Nat. Prod. Commun. 2007, 2, 1934578X0700200308. [Google Scholar] [CrossRef]
- Thanati, F.; Karatzas, E.; Baltoumas, F.A.; Stravopodis, D.J.; Eliopoulos, A.G.; Pavlopoulos, G.A. FLAME: A Web Tool for Functional and Literature Enrichment Analysis of Multiple Gene Lists. Biology 2021, 10, 665. [Google Scholar] [CrossRef]
- Karatzas, E.; Baltoumas, F.A.; Kasionis, I.; Sanoudou, D.; Eliopoulos, A.G.; Theodosiou, T.; Iliopoulos, I.; Pavlopoulos, G.A. Darling: A Web Application for Detecting Disease-Related Biomedical Entity Associations with Literature Mining. Biomolecules 2022, 12, 520. [Google Scholar] [CrossRef] [PubMed]
- Pinero, J.; Queralt-Rosinach, N.; Bravo, A.; Deu-Pons, J.; Bauer-Mehren, A.; Baron, M.; Sanz, F.; Furlong, L.I. DisGeNET: A discovery platform for the dynamical exploration of human diseases and their genes. Database 2015, 2015, bav028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillespie, M.; Jassal, B.; Stephan, R.; Milacic, M.; Rothfels, K.; Senff-Ribeiro, A.; Griss, J.; Sevilla, C.; Matthews, L.; Gong, C.; et al. The reactome pathway knowledgebase 2022. Nucleic Acids Res. 2022, 50, D687–D692. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Zhang, Y.; Wang, X.Z.; Zhao, J.; Yang, Z.J.; Lin, Y.P.; Sun, L.; Lu, Q.Y.; Fan, G.J. Flavonoids for Treating Viral Acute Respiratory Tract Infections: A Systematic Review and Meta-Analysis of 30 Randomized Controlled Trials. Front. Public Health 2022, 10, 814669. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wang, L.; Pan, X.; Liang, Y.; Zhang, Y.; Li, J.; Zhou, B. 5-Methoxyflavone-induced AMPKalpha activation inhibits NF-kappaB and P38 MAPK signaling to attenuate influenza A virus-mediated inflammation and lung injury in vitro and in vivo. Cell. Mol. Biol. Lett. 2022, 27, 82. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.T.; Tung, B.T.; Nguyen, P.H.; Thuong, P.T.; Yoo, S.S.; Kim, E.H.; Kim, S.K.; Oh, W.K. C-Methylated Flavonoids from Cleistocalyx operculatus and Their Inhibitory Effects on Novel Influenza A (H1N1) Neuraminidase. J. Nat. Prod. 2010, 73, 1636–1642. [Google Scholar] [CrossRef]
- Xie, C.F.; Qu, J.B.; Wu, X.Z.; Liu, N.; Ji, M.; Lou, H.X. Antifungal macrocyclic bis(bibenzyls) from the Chinese liverwort Ptagiochasm intermedlum L. Nat. Prod. Res. 2010, 24, 515–520. [Google Scholar] [CrossRef]
- Ortiz-Lopez, T.; Borges-Argaez, R.; Ayora-Talavera, G.; Canto-Ramirez, E.; Cetina-Montejo, L.; May-May, A.; Escalante-Erosa, F.; Caceres-Farfan, M. Bioassay-Guided Fractionation of Erythrostemon yucatanensis (Greenm.) Gagnon & GP Lewis Components with Anti-hemagglutinin Binding Activity against Influenza A/H1N1 Virus. Molecules 2022, 27, 5494. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, J.; Ke, C.; Zhang, H.; Zhang, S.; Tang, W.; Liu, C.; Liu, G.; Chen, S.; Hu, A.; et al. Ethanol Extract of Caesalpinia decapetala Inhibits Influenza Virus Infection In Vitro and In Vivo. Viruses 2020, 12, 557. [Google Scholar] [CrossRef]
- Galabov, A.S.; Mileva, M.; Simeonova, L.; Gegova, G. Combination activity of neuraminidase inhibitor oseltamivir and alpha-tocopherol in influenza virus A (H3N2) infection in mice. Antivir. Chem. Chemother. 2015, 24, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Juarez-Mendez, M.T.; Borges-Argaez, R.; Ayora-Talavera, G.; Escalante-Rebolledo, S.E.; Escalante-Erosa, F.; Caceres-Farfan, M. Diospyros anisandra phytochemical analysis and anti-hemagglutinin-neuraminidase activity on influenza AH1N1pdm09 virus. Nat. Prod. Res. 2022, 36, 2666–2672. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.B.; Kim, S.; Song, K. Neuraminidase Inhibitors from Mushroom Microphorus affinis. J. Microbiol. Biotechnol. 2003, 13, 778–782. [Google Scholar]
- Kim, M.; Choi, H.; Kim, S.; Kang, L.W.; Kim, Y.B. Elucidating the Effects of Curcumin against Influenza Using In Silico and In Vitro Approaches. Pharmaceuticals 2021, 14, 880. [Google Scholar] [CrossRef] [PubMed]
- Gui, R.; Chen, Q. Molecular Events Involved in Influenza A Virus-Induced Cell Death. Front. Microbiol. 2021, 12, 797789. [Google Scholar] [CrossRef] [PubMed]
- Wurzer, W.J.; Ehrhardt, C.; Pleschka, S.; Berberich-Siebelt, F.; Wolff, T.; Walczak, H.; Planz, O.; Ludwig, S. NF-kappaB-dependent induction of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) and Fas/FasL is crucial for efficient influenza virus propagation. J. Biol. Chem. 2004, 279, 30931–30937. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.; Xi, C.; Hu, K.; Gao, W.; Cai, X.; Qin, J.; Lv, S.; Du, C.; Wei, Y. Inhibition of enterovirus 71 replication and viral 3C protease by quercetin. Virol. J. 2018, 15, 116. [Google Scholar] [CrossRef] [Green Version]
- Pandey, K.P.; Zhou, Y. Influenza A Virus Infection Activates NLRP3 Inflammasome through Trans-Golgi Network Dispersion. Viruses 2022, 14, 88. [Google Scholar] [CrossRef]
- Gkouskou, K.; Vasilogiannakopoulou, T.; Andreakos, E.; Davanos, N.; Gazouli, M.; Sanoudou, D.; Eliopoulos, A.G. COVID-19 enters the expanding network of apolipoprotein E4-related pathologies. Redox Biol. 2021, 41, 101938. [Google Scholar] [CrossRef]
- Kumar, N.; Xin, Z.T.; Liang, Y.; Ly, H.; Liang, Y. NF-kappaB signaling differentially regulates influenza virus RNA synthesis. J. Virol. 2008, 82, 9880–9889. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.N.; Yu, C.H.; Cai, W.; Hua, J.; Li, S.Q.; Wang, W. Protective effect of a polyphenolic rich extract from Magnolia officinalis bark on influenza virus-induced pneumonia in mice. J. Ethnopharmacol. 2011, 134, 191–194. [Google Scholar] [CrossRef]
- Yan, H.; Wang, H.; Ma, L.; Ma, X.; Yin, J.; Wu, S.; Huang, H.; Li, Y. Cirsimaritin inhibits influenza A virus replication by downregulating the NF-kappaB signal transduction pathway. Virol. J. 2018, 15, 88. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Bao, L.; Wang, Y.; Chen, M.; Zhang, Y.; Geng, Z.; Zhao, R.; Sun, J.; Bao, Y.; Shi, Y.; et al. An Integrated Analysis Reveals Geniposide Extracted From Gardenia jasminoides J.Ellis Regulates Calcium Signaling Pathway Essential for Influenza A Virus Replication. Front. Pharm. 2021, 12, 755796. [Google Scholar] [CrossRef] [PubMed]
- Raus, K.; Pleschka, S.; Klein, P.; Schoop, R.; Fisher, P. Effect of an Echinacea-Based Hot Drink Versus Oseltamivir in Influenza Treatment: A Randomized, Double-Blind, Double-Dummy, Multicenter, Noninferiority Clinical Trial. Curr. Ther. Res. Clin. Exp. 2015, 77, 66–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodinet, C.; Mentel, R.; Wegner, U.; Lindequist, U.; Teuscher, E.; Freudenstein, J. Effect of oral application of an immunomodulating plant extract on Influenza virus type A infection in mice. Planta Med. 2002, 68, 896–900. [Google Scholar] [CrossRef]
- Kalus, U.; Grigorov, A.; Kadecki, O.; Jansen, J.P.; Kiesewetter, H.; Radtke, H. Cistus incanus (CYSTUS052) for treating patients with infection of the upper respiratory tract. A prospective, randomised, placebo-controlled clinical study. Antivir. Res. 2009, 84, 267–271. [Google Scholar] [CrossRef]
- Gaush, C.R.; Smith, T.F. Replication and plaque assay of influenza virus in an established line of canine kidney cells. Appl. Microbiol. 1968, 16, 588–594. [Google Scholar] [CrossRef]
- Lin, S.C.; Kappes, M.A.; Chen, M.C.; Lin, C.C.; Wang, T.T. Distinct susceptibility and applicability of MDCK derivatives for influenza virus research. PLoS ONE 2017, 12, e0172299. [Google Scholar] [CrossRef] [Green Version]
- Vardouli, L.; Lindqvist, C.; Vlahou, K.; Loskog, A.S.; Eliopoulos, A.G. Adenovirus delivery of human CD40 ligand gene confers direct therapeutic effects on carcinomas. Cancer Gene. Ther. 2009, 16, 848–860. [Google Scholar] [CrossRef]
- Galani, I.E.; Triantafyllia, V.; Eleminiadou, E.E.; Koltsida, O.; Stavropoulos, A.; Manioudaki, M.; Thanos, D.; Doyle, S.E.; Kotenko, S.V.; Thanopoulou, K.; et al. Interferon-lambda Mediates Non-redundant Front-Line Antiviral Protection against Influenza Virus Infection without Compromising Host Fitness. Immunity 2017, 46, 875–890 e876. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eliopoulos, A.G.; Angelis, A.; Liakakou, A.; Skaltsounis, L.A. In Vitro Anti-Influenza Virus Activity of Non-Polar Primula veris subsp. veris Extract. Pharmaceuticals 2022, 15, 1513. https://doi.org/10.3390/ph15121513
Eliopoulos AG, Angelis A, Liakakou A, Skaltsounis LA. In Vitro Anti-Influenza Virus Activity of Non-Polar Primula veris subsp. veris Extract. Pharmaceuticals. 2022; 15(12):1513. https://doi.org/10.3390/ph15121513
Chicago/Turabian StyleEliopoulos, Aristides G., Apostolis Angelis, Anastasia Liakakou, and Leandros A. Skaltsounis. 2022. "In Vitro Anti-Influenza Virus Activity of Non-Polar Primula veris subsp. veris Extract" Pharmaceuticals 15, no. 12: 1513. https://doi.org/10.3390/ph15121513
APA StyleEliopoulos, A. G., Angelis, A., Liakakou, A., & Skaltsounis, L. A. (2022). In Vitro Anti-Influenza Virus Activity of Non-Polar Primula veris subsp. veris Extract. Pharmaceuticals, 15(12), 1513. https://doi.org/10.3390/ph15121513