
Plant Terpenoids as Hit Compounds against Trypanosomiasis


 , and
, and 
Abstract
:1. Introduction
2. Trypanosomiasis
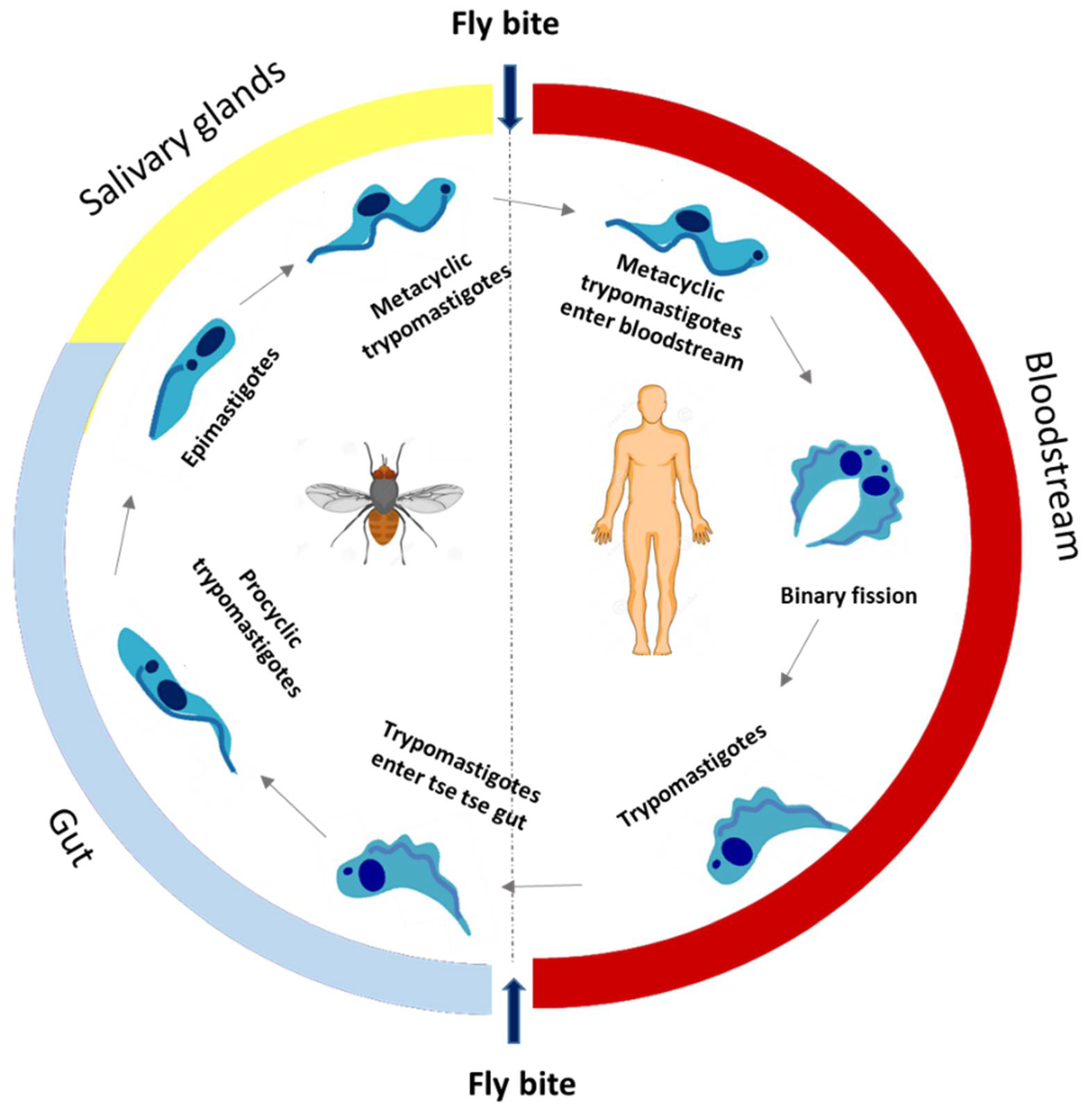
2.1. Human African Trypanosomiasis (HAT)
2.1.1. Antitrypanosomal Chemotherapy Targets and Current Drugs against HAT
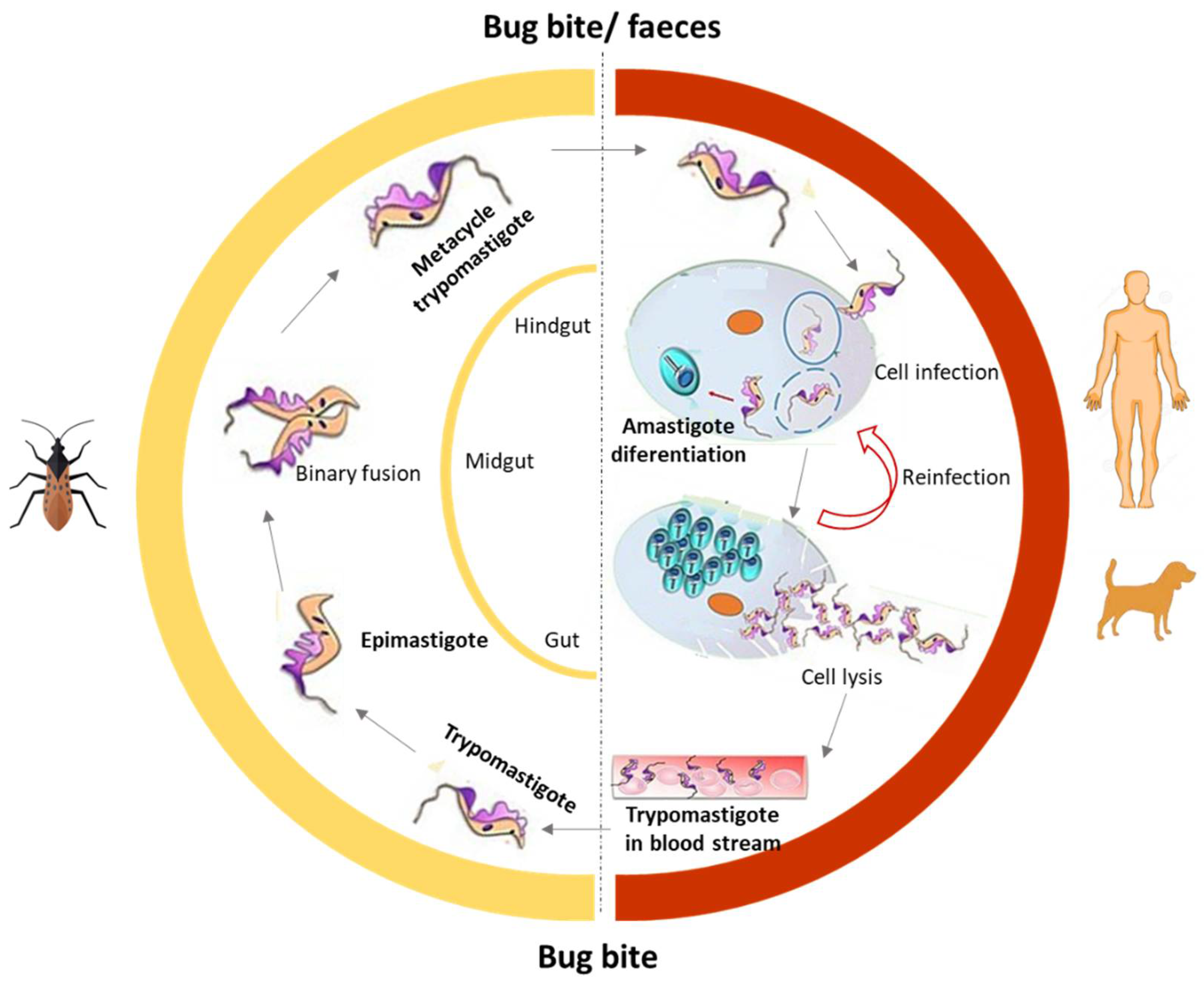
2.2. Human American Trypanosomiasis (Chagas Disease, CD)
2.2.1. Antitrypanosomal Chemotherapy Targets and Current Drugs against CD
3. Terpenic Compounds with Antitrypanosomal Activity
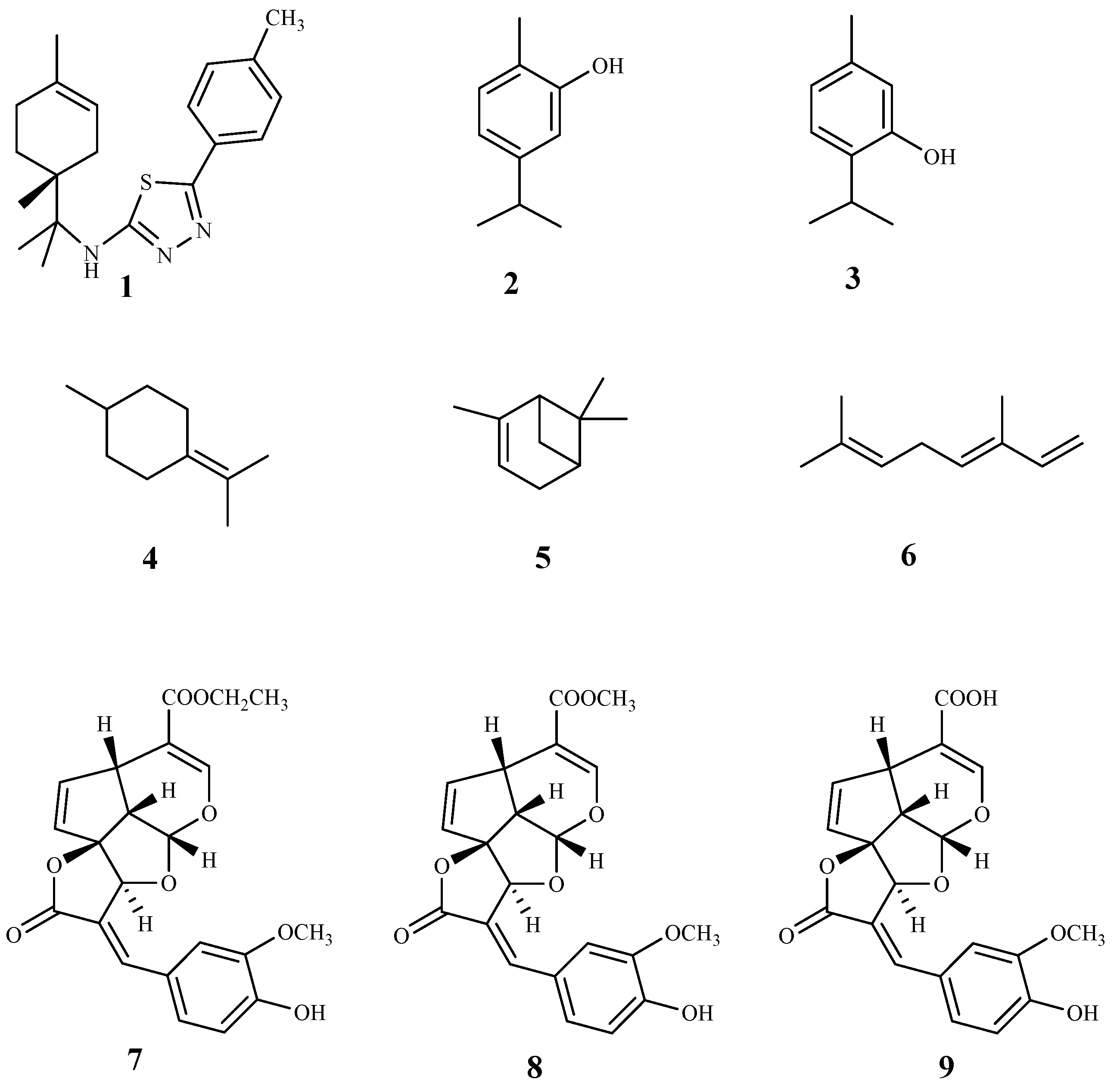
3.1. Monoterpenes and Iridoids
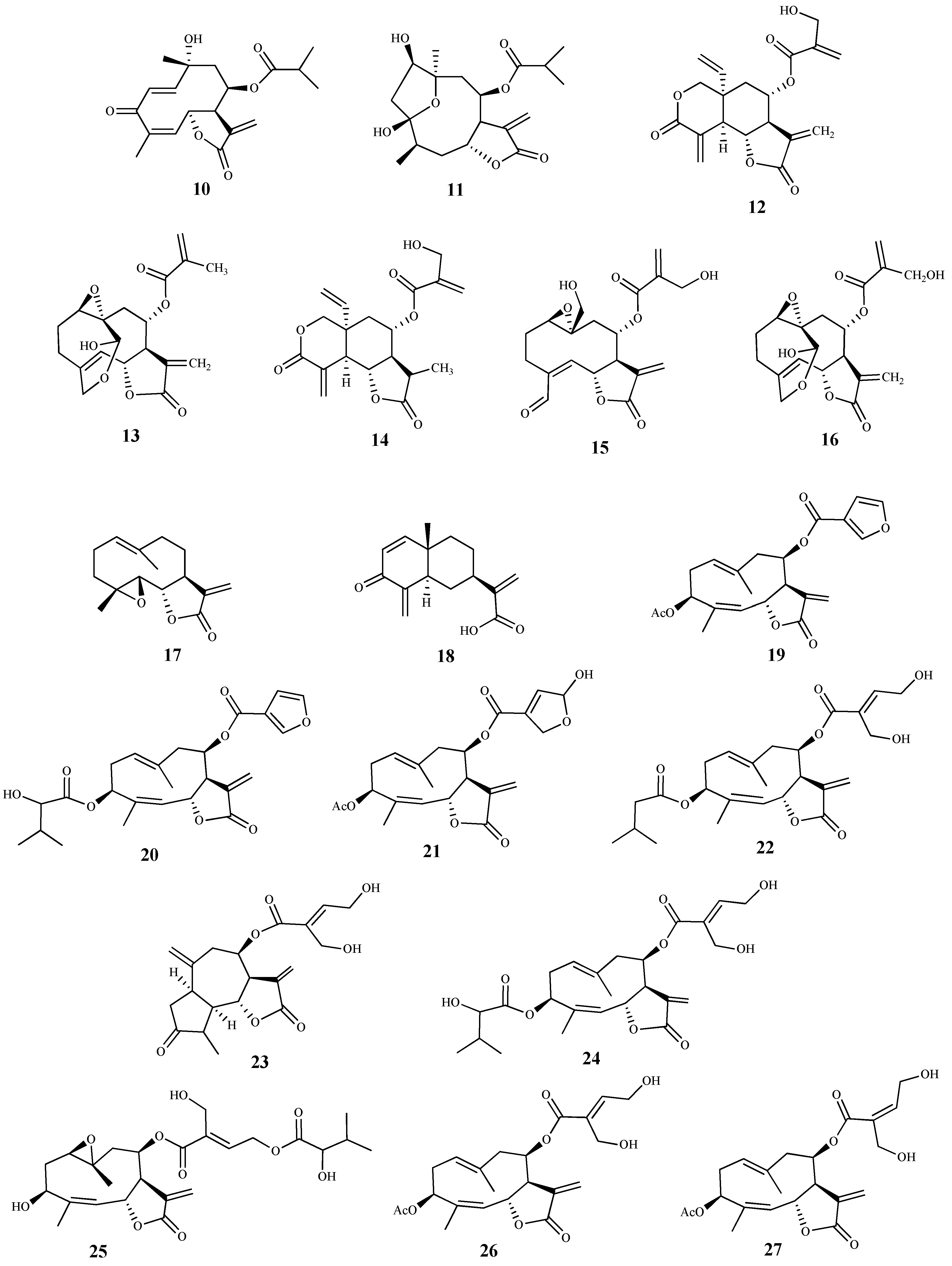
3.2. Sesquiterpenes
3.3. Diterpenes
3.4. Triterpenes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- World Health Organization. Ending the Neglect to Attain the Sustainable Development Goals. A Road Map for Neglected Tropical Diseases 2021–2030; Ntuli, M.M., Ed.; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Hotez, P.J.; Aksoy, S.; Brindley, P.J.; Kamhawi, S. What constitutes a neglected tropical disease? PLoS Negl. Trop. Dis. 2020, 14, e0008001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisuzzaman; Tsuji, N. Schistosomiasis and hookworm infection in humans: Disease burden, pathobiology and anthelmintic vaccines. Parasitol. Int. 2020, 75, 102051. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Corbeil, A.; Do Monte-Neto, R.L.; Fernandez-Prada, C. Of drugs and trypanosomatids: New tools and knowledge to reduce bottlenecks in drug discovery. Genes 2020, 11, 722. [Google Scholar] [CrossRef] [PubMed]
- Molyneux, D.H.; Asamoa-Bah, A.; Fenwick, A.; Savioli, L.; Hotez, P. The history of the neglected tropical disease movement. Trans. R. Soc. Trop. Med. Hyg. 2021, 115, 169–175. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Neglected tropical diseases: Impact of COVID-19 and WHO’s response—Maladies tropicales négligées: Impact de la COVID-19 et réponse de l’OMS. Wkly. Epidemiol. Rec. 2020, 95, 461–468. [Google Scholar]
- Cheuka, P.; Mayoka, G.; Mutai, P.; Chibale, K. The Role of Natural Products in Drug Discovery and Development against Neglected Tropical Diseases. Molecules 2017, 22, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capela, R.; Moreira, R.; Lopes, F. An overview of drug resistance in protozoal diseases. Int. J. Mol. Sci. 2019, 20, 5748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Herrera, A.; Cortez-Maya, S.; Bocanegra-Garcia, V.; Banik, B.K.; Rivera, G. Recent Advances in the Development of Broad-Spectrum Antiprotozoal Agents. Curr. Med. Chem. 2020, 28, 583–606. [Google Scholar] [CrossRef] [PubMed]
- Field, M.C.; Horn, D.; Fairlamb, A.H.; Ferguson, M.A.J.; Gray, D.W.; Read, K.D.; De Rycker, M.; Torrie, L.S.; Wyatt, P.G.; Wyllie, S.; et al. Anti-trypanosomatid drug discovery: An ongoing challenge and a continuing need. Nat. Rev. Microbiol. 2017, 15, 217–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournet, A.; Munoz, V.; Muñoz, V. Natural products as trypanocidal, antileishmanial and antimalarial drugs. Curr. Top. Med. Chem. 2002, 2, 1215–1237. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.J.; Khalid, S.A.; Romanha, A.J.; Alves, T.M.A.; Biavatti, M.W.; Brun, R.; Da Costa, F.B.; de Castro, S.L.; Ferreira, V.F.; de Lacerda, M.V.G.; et al. The Potential of Secondary Metabolites from Plants as Drugs or Leads Against Protozoan Neglected Diseases—Part II. Curr. Med. Chem. 2012, 19, 2176–2228. [Google Scholar] [CrossRef] [PubMed]
- Tullius Scotti, M.; Scotti, L.; Ishiki, H.; Fávaro Ribeiro, F.; da Cruz, R.M.D.; de Oliveira, M.P.; Jaime Bezerra Mendonça, F. Natural Products as a Source for Antileishmanial and Antitrypanosomal Agents. Comb. Chem. High Throughput Screen. 2016, 19, 537–553. [Google Scholar] [CrossRef] [PubMed]
- Isah, M.B.; Ibrahim, M.A.; Mohammed, A.; Aliyu, A.B.; Masola, B.; Coetzer, T.H.T. A systematic review of pentacyclic triterpenes and their derivatives as chemotherapeutic agents against tropical parasitic diseases. Parasitology 2016, 143, 1219–1231. [Google Scholar] [CrossRef] [PubMed]
- Muschietti, L.V.; Ulloa, J.L. Natural Sesquiterpene Lactones as Potential Trypanocidal Therapeutic Agents: A Review. Nat. Prod. Commun. 2016, 11, 1569–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cockram, P.E.; Smith, T.K. Active Natural Product Scaffolds against Trypanosomatid Parasites: A Review. J. Nat. Prod. 2018, 81, 2138–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ungogo, M.A.; Ebiloma, G.U.; Ichoron, N.; Igoli, J.O.; de Koning, H.P.; Balogun, E.O. A Review of the Antimalarial, Antitrypanosomal, and Antileishmanial Activities of Natural Compounds Isolated From Nigerian Flora. Front. Chem. 2020, 8, 1221. [Google Scholar] [CrossRef]
- Chan-Bacab, M.J.; Reyes-Estebanez, M.M.; Camacho-Chab, J.C.; Ortega-Morales, B.O. Microorganisms as a potential source of molecules to control trypanosomatid diseases. Molecules 2021, 26, 1388. [Google Scholar] [CrossRef]
- Hzounda Fokou, J.B.; Dize, D.; Etame Loe, G.M.; Nko’o, M.H.J.; Ngene, J.P.; Ngoule, C.C.; Boyom, F.F. Anti-leishmanial and anti-trypanosomal natural products from endophytes. Parasitol. Res. 2021, 120, 785–796. [Google Scholar] [CrossRef]
- Lenzi, J.; Costa, T.M.; Alberton, M.D.; Goulart, J.A.G.; Tavares, L.B.B. Medicinal fungi: A source of antiparasitic secondary metabolites. Appl. Microbiol. Biotechnol. 2018, 102, 5791–5810. [Google Scholar] [CrossRef]
- Nweze, J.A.; Mbaoji, F.N.; Li, Y.M.; Yang, L.Y.; Huang, S.S.; Chigor, V.N.; Eze, E.A.; Pan, L.X.; Zhang, T.; Yang, D.F. Potentials of marine natural products against malaria, leishmaniasis, and trypanosomiasis parasites: A review of recent articles. Infect. Dis. Poverty 2021, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Lucas, L.A.; Cândido, A.C.B.B.; Santos, M.F.C.; Caffrey, C.R.; Bastos, J.K.; Ambrósio, S.R.; Magalhães, L.G. Antiparasitic Properties of Propolis Extracts and Their Compounds. Chem. Biodivers. 2021, 18, 310. [Google Scholar] [CrossRef]
- Asfaram, S.; Fakhar, M.; Keighobadi, M.; Akhtari, J. Promising Anti-Protozoan Activities of Propolis (Bee Glue) as Natural Product: A Review. Acta Parasitol. 2021, 66, 1–12. [Google Scholar] [CrossRef]
- Simoben, C.V.; Ntie-Kang, F.; Akone, S.H.; Sippl, W. Compounds from African Medicinal Plants with Activities Against Selected Parasitic Diseases: Schistosomiasis, Trypanosomiasis and Leishmaniasis. Nat. Products Bioprospect. 2018, 8, 151–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naß, J.; Efferth, T. The activity of Artemisia spp. and their constituents against Trypanosomiasis. Phytomedicine 2018, 47, 184–191. [Google Scholar] [CrossRef] [PubMed]
- de Morais, M.C.; de Souza, J.V.; da Silva Maia Bezerra Filho, C.; Dolabella, S.S.; de Sousa, D.P. Trypanocidal essential oils: A review. Molecules 2020, 25, 4568. [Google Scholar] [CrossRef] [PubMed]
- Nair, J.J.; van Staden, J. Antiprotozoal alkaloid principles of the plant family Amaryllidaceae. Bioorg. Med. Chem. Lett. 2019, 29, 126642. [Google Scholar] [CrossRef] [PubMed]
- Montesino, N.L.; Schmidt, T.J. Salvia species as sources of natural products with antiprotozoal activity. Int. J. Mol. Sci. 2018, 19, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, J.K.; da Trindade, R.; Alves, N.S.; Figueiredo, P.L.; Maia, J.G.S.; Setzer, W.N. Essential Oils from Neotropical Piper Species and Their Biological Activities. Int. J. Mol. Sci. 2017, 18, 2571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Human African Trypanosomiasis (Sleeping Sickness). Available online: https://www.who.int/health-topics/human-african-trypanosomiasis#tab=tab_1 (accessed on 13 January 2022).
- World Health Organization. Trypanosomiasis, Human African (Sleeping Sickness). Available online: https://www.who.int/news-room/fact-sheets/detail/trypanosomiasis-human-african-(sleeping-sickness) (accessed on 4 March 2021).
- Drugs for Neglected Diseases Initiative (DNDi) Sleping Sickness. Available online: https://dndi.org/diseases/sleeping-sickness/facts/ (accessed on 7 January 2022).
- Centers for Disease Control and Prevention (CDC) Parasites—African Trypanosomiasis (also Known as Sleeping Sickness). Available online: https://www.cdc.gov/parasites/sleepingsickness/ (accessed on 6 January 2022).
- Keiser, J.; Stich, A.; Burri, C. New drugs for the treatment of human African trypanosomiasis: Research and development. Parasitol. Today 2001, 17, 42–49. [Google Scholar] [CrossRef]
- Brand, S.; Norcross, N.R.; Thompson, S.; Harrison, J.R.; Smith, V.C.; Robinson, D.A.; Torrie, L.S.; McElroy, S.P.; Hallyburton, I.; Norval, S.; et al. Lead optimization of a pyrazole sulfonamide series of trypanosoma brucei N -myristoyltransferase inhibitors: Identification and evaluation of CNS penetrant compounds as potential treatments for stage 2 human african trypanosomiasis. J. Med. Chem. 2014, 57, 9855–9869. [Google Scholar] [CrossRef] [PubMed]
- Bijlmakers, M.J. Ubiquitination and the Proteasome as Drug Targets in Trypanosomatid Diseases. Front. Chem. 2021, 8, e630888. [Google Scholar] [CrossRef] [PubMed]
- Kourbeli, V.; Chontzopoulou, E.; Moschovou, K.; Pavlos, D.; Mavromoustakos, T.; Papanastasiou, I.P. An Overview on Target-Based Drug Design against Kinetoplastid Protozoan Infections: Human African. Molecules 2021, 26, 4629. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Khan, S.A.; Alshammari, M.K.; Alqahtani, A.M.; Alanazi, T.A.; Kamal, M.; Jawaid, T.; Ghoneim, M.M.; Alshehri, S.; Shakeel, F. Discovery, Development, Inventions and Patent Review of Fexinidazole: The First All-Oral Therapy for Human African Trypanosomiasis. Pharmaceuticals 2022, 15, 128. [Google Scholar] [CrossRef] [PubMed]
- Steketee, P.C.; Giordani, F.; Vincent, I.M.; Crouch, K.; Achcar, F.; Dickens, N.J.; Morrison, L.J.; MacLeod, A.; Barrett, M.P. Transcriptional differentiation of Trypanosoma brucei during in vitro acquisition of resistance to acoziborole. PLoS Negl. Trop. Dis. 2021, 15, e0009939. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Chagas Disease (also Known as American Trypanosomiasis). Available online: https://www.who.int/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (accessed on 4 March 2021).
- Dumonteil, E.; Herrera, C. The Case for the Development of a Chagas Disease Vaccine: Why? How? When? Trop. Med. Infect. Dis. 2021, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Lidani, K.C.F.; Andrade, F.A.; Bavia, L.; Damasceno, F.S.; Beltrame, M.H.; Messias-Reason, I.J.; Sandri, T.L. Chagas disease: From discovery to a worldwide health problem. J. Phys. Oceanogr. 2019, 49, 166. [Google Scholar] [CrossRef] [PubMed]
- Mills, R.M. Chagas Disease: Epidemiology and Barriers to Treatment. Am. J. Med. 2020, 133, 1262–1265. [Google Scholar] [CrossRef] [PubMed]
- Duarte-Silva, E.; Morais, L.H.; Clarke, G.; Savino, W.; Peixoto, C. Targeting the Gut Microbiota in Chagas Disease: What Do We Know so Far? Front. Microbiol. 2020, 11, 3083. [Google Scholar] [CrossRef]
- Onyekwelu, K. Life Cycle of Trypanosoma cruzi in the Invertebrate and the Vertebrate Hosts. In Biology of Trypanosoma Cruzi; De Souza, W., Ed.; IntechOpen: London, UK, 2019; ISBN 978-1-83968-204-9. [Google Scholar]
- Francisco, A.F.; Jayawardhana, S.; Olmo, F.; Lewis, M.D.; Wilkinson, S.R.; Taylor, M.C.; Kelly, J.M. Challenges in Chagas Disease Drug Development. Molecules 2020, 25, 2799. [Google Scholar] [CrossRef]
- Villalta, F.; Rachakonda, G. Advances in preclinical approaches to Chagas disease drug discovery. Expert Opin. Drug Discov. 2019, 14, 1161–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Huertas, P.; Cardona-Castro, N. Advances in the treatment of Chagas disease: Promising new drugs, plants and targets. Biomed. Pharmacother. 2021, 142, 112020. [Google Scholar] [CrossRef] [PubMed]
- Torrico, F.; Gascón, J.; Barreira, F.; Blum, B.; Almeida, I.C.; Alonso-Vega, C.; Barboza, T.; Bilbe, G.; Correia, E.; Garcia, W.; et al. New regimens of benznidazole monotherapy and in combination with fosravuconazole for treatment of Chagas disease (BENDITA): A phase 2, double-blind, randomised trial. Lancet Infect. Dis. 2021, 21, 1129–1140. [Google Scholar] [CrossRef]
- Don, R.; Ioset, J.-R. Screening strategies to identify new chemical diversity for drug development to treat kinetoplastid infections. Parasitology 2014, 141, 140–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsuno, K.; Burrows, J.N.; Duncan, K.; Van Huijsduijnen, R.H.; Kaneko, T.; Kita, K.; Mowbray, C.E.; Schmatz, D.; Warner, P.; Slingsby, B.T. Hit and lead criteria in drug discovery for infectious diseases of the developing world. Nat. Rev. Drug Discov. 2015, 14, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Martins, S.C.; Lazarin-Bidóia, D.; Desoti, V.C.; Falzirolli, H.; da Silva, C.C.; Ueda-Nakamura, T.; de O. Silva, S.; Nakamura, C.V. 1,3,4-Thiadiazole derivatives of R-(+)-limonene benzaldehyde-thiosemicarbazones cause death in Trypanosoma cruzi through oxidative stress. Microbes Infect. 2016, 18, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Tasdemir, D.; Kaiser, M.; Demirci, B.; Demirci, F.; Hüsnü Can Baser, K. Antiprotozoal Activity of Turkish Origanum onites Essential Oil and Its Components. Molecules 2019, 24, 4421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngahang Kamte, S.L.; Ranjbarian, F.; Cianfaglione, K.; Sut, S.; Dall’Acqua, S.; Bruno, M.; Afshar, F.H.; Iannarelli, R.; Benelli, G.; Cappellacci, L.; et al. Identification of highly effective antitrypanosomal compounds in essential oils from the Apiaceae family. Ecotoxicol. Environ. Saf. 2018, 156, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Kwofie, K.D.; Tung, N.H.; Suzuki-Ohashi, M.; Amoa-Bosompem, M.; Adegle, R.; Sakyiamah, M.M.; Ayertey, F.; Owusu, K.B.A.; Tuffour, I.; Atchoglo, P.; et al. Antitrypanosomal activities and mechanisms of action of novel tetracyclic iridoids from Morinda lucida Benth. Antimicrob. Agents Chemother. 2016, 60, 3283–3290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sut, S.; Dall’Acqua, S.; Baldan, V.; Ngahang Kamte, S.L.; Ranjbarian, F.; Biapa Nya, P.C.; Vittori, S.; Benelli, G.; Maggi, F.; Cappellacci, L.; et al. Identification of tagitinin C from Tithonia diversifolia as antitrypanosomal compound using bioactivity-guided fractionation. Fitoterapia 2018, 124, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimani, N.M.; Matasyoh, J.C.; Kaiser, M.; Brun, R.; Schmidt, T.J. Sesquiterpene lactones from Vernonia cinerascens Sch. Bip. and their in vitro antitrypanosomal activity. Molecules 2018, 23, 248. [Google Scholar] [CrossRef] [Green Version]
- Kimani, N.M.; Matasyoh, J.C.; Kaiser, M.; Brun, R.; Schmidt, T.J. Anti-trypanosomatid elemanolide sesquiterpene lactones from Vernonia lasiopus O. Hoffm. Molecules 2017, 22, 597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimani, N.M.; Matasyoh, J.C.; Kaiser, M.; Brun, R.; Schmidt, T.J. Antiprotozoal Sesquiterpene Lactones and Other Constituents from Tarchonanthus camphoratus and Schkuhria pinnata. J. Nat. Prod. 2018, 81, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Skaf, J.; Hamarsheh, O.; Berninger, M.; Balasubramanian, S.; Oelschlaeger, T.A.; Holzgrabe, U. Improving anti-trypanosomal activity of alkamides isolated from Achillea fragrantissima. Fitoterapia 2018, 125, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Laurella, L.C.; Cerny, N.; Bivona, A.E.; Alberti, A.S.; Giberti, G.; Malchiodi, E.L.; Martino, V.S.; Catalan, C.A.; Alonso, M.R.; Cazorla, S.I.; et al. Assessment of sesquiterpene lactones isolated from Mikania plants species for their potential efficacy against Trypanosoma cruzi and Leishmania sp. PLoS Negl. Trop. Dis. 2017, 11, e0005929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrelli, R.; Ranjbarian, F.; Dall’Acqua, S.; Papa, F.; Iannarelli, R.; Ngahang Kamte, S.L.; Vittori, S.; Benelli, G.; Maggi, F.; Hofer, A.; et al. An overlooked horticultural crop, Smyrnium olusatrum, as a potential source of compounds effective against African trypanosomiasis. Parasitol. Int. 2017, 66, 146–151. [Google Scholar] [CrossRef] [PubMed]
- De Mieri, M.; Monteleone, G.; Ismajili, I.; Kaiser, M.; Hamburger, M. Antiprotozoal Activity-Based Profiling of a Dichloromethane Extract from Anthemis nobilis Flowers. J. Nat. Prod. 2017, 80, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Lima, T.C.; Souza, R.D.J.; De Moraes, M.H.; Steindel, M.; Biavatti, M.W. A new furanoheliangolide sesquiterpene lactone from Calea pinnatifida (R. Br.) Less. (Asteraceae) and evaluation of its trypanocidal and leishmanicidal activities. J. Braz. Chem. Soc. 2017, 28, 367–375. [Google Scholar] [CrossRef]
- Ulloa, J.L.; Spina, R.; Casasco, A.; Petray, P.B.; Martino, V.; Sosa, M.A.; Frank, F.M.; Muschietti, L.V. Germacranolide-type sesquiterpene lactones from Smallanthus sonchifolius with promising activity against Leishmania mexicana and Trypanosoma cruzi. Parasites Vectors 2017, 10, 567. [Google Scholar] [CrossRef] [PubMed]
- Nyongbela, K.D.; Ntie-Kang, F.; Hoye, T.R.; Efange, S.M.N. Antiparasitic Sesquiterpenes from the Cameroonian Spice Scleria striatinux and Preliminary In Vitro and In Silico DMPK Assessment. Nat. Products Bioprospect. 2017, 7, 235–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elso, O.G.; Bivona, A.E.; Alberti, A.S.; Cerny, N.; Fabian, L.; Morales, C.; Catalán, C.A.N.; Malchiodi, E.L.; Cazorla, S.I.; Sülsen, V.P. Trypanocidal activity of four sesquiterpene lactones isolated from Asteraceae species. Molecules 2020, 25, 2014. [Google Scholar] [CrossRef]
- Gonçalves-Santos, E.; Vilas-Boas, D.F.; Diniz, L.F.; Veloso, M.P.; Mazzeti, A.L.; Rodrigues, M.R.; Oliveira, C.M.; Fernandes, V.H.C.; Novaes, R.D.; Chagas-Paula, D.A.; et al. Sesquiterpene lactone potentiates the immunomodulatory, antiparasitic and cardioprotective effects on anti-Trypanosoma cruzi specific chemotherapy. Int. Immunopharmacol. 2019, 77, 105961. [Google Scholar] [CrossRef] [PubMed]
- Galkina, A.; Krause, N.; Lenz, M.; Daniliuc, C.G.; Kaiser, M.; Schmidt, T.J. Antitrypanosomal Activity of Sesquiterpene Lactones from Helianthus tuberosus L. Including a new furanoheliangolide with an unusual structure. Molecules 2019, 24, 1068. [Google Scholar] [CrossRef] [Green Version]
- Sosa, A.; Salamanca Capusiri, E.; Amaya, S.; Bardón, A.; Giménez-Turba, A.; Vera, N.; Borkosky, S. Trypanocidal activity of South American Vernonieae (Asteraceae) extracts and its sesquiterpene lactones. Nat. Prod. Res. 2021, 35, 5224–5228. [Google Scholar] [CrossRef]
- Elso, O.G.; Clavin, M.; Hernandez, N.; Sgarlata, T.; Bach, H.; Catalan, C.A.N.; Aguilera, E.; Alvarez, G.; Sülsen, V.P. Antiprotozoal Compounds from Urolepis hecatantha (Asteraceae). Evid. -Based Complement. Altern. Med. 2021, 2021, 6622894. [Google Scholar] [CrossRef] [PubMed]
- Milagre, M.M.; Branquinho, R.T.; Gonçalves, M.F.; de Assis, G.; de Oliveira, M.T.; Reis, L.; Saúde-Guimarães, D.A.; de Lana, M. Activity of the sesquiterpene lactone goyazensolide against Trypanosoma cruzi in vitro and in vivo. Parasitology 2020, 147, 108–119. [Google Scholar] [CrossRef]
- Murakami, C.; Cabral, R.S.A.; Gomes, K.S.; Costa-Silva, T.A.; Amaral, M.; Romanelli, M.; Tempone, A.G.; Lago, J.H.G.; da Bolzani, V.S.; Moreno, P.R.H.; et al. Hedyosulide, a novel trypanosomicidal sesterterpene lactone from Hedyosmum brasiliense Mart. ex Miq. Phytochem. Lett. 2019, 33, 6–11. [Google Scholar] [CrossRef]
- Zeouk, I.; Sifaoui, I.; López-Arencibia, A.; Reyes-Batlle, M.; Bethencourt-Estrella, C.J.; Bazzocchi, I.L.; Bekhti, K.; Lorenzo-Morales, J.; Jiménez, I.A.; Piñero, J.E. Sesquiterpenoids and flavonoids from Inula viscosa induce programmed cell death in kinetoplastids. Biomed. Pharmacother. 2020, 130, 110518. [Google Scholar] [CrossRef] [PubMed]
- Londero, V.S.; Costa-Silva, T.A.; Tempone, A.G.; Namiyama, G.M.; Thevenard, F.; Antar, G.M.; Baitello, J.B.; Lago, J.H.G. Anti-Trypanosoma cruzi activity of costic acid isolated from Nectandra barbellata (Lauraceae) is associated with alterations in plasma membrane electric and mitochondrial membrane potentials. Bioorg. Chem. 2020, 95, 103510. [Google Scholar] [CrossRef]
- Mofidi Tabatabaei, S.; Nejad Ebrahimi, S.; Salehi, P.; Sonboli, A.; Tabefam, M.; Kaiser, M.; Hamburger, M.; Moridi Farimani, M. Antiprotozoal Germacranolide Sesquiterpene Lactones from Tanacetum sonbolii. Planta Med. 2019, 85, 424–430. [Google Scholar] [CrossRef] [Green Version]
- Turner, D.N.; Just, J.; Dasari, R.; Smith, J.A.; Bissember, A.C.; Kornienko, A.; Rogelj, S. Activity of natural and synthetic polygodial derivatives against Trypanosoma cruzi amastigotes, trypomastigotes and epimastigotes. Nat. Prod. Res. 2021, 35, 792–795. [Google Scholar] [CrossRef] [PubMed]
- Bombaça, A.C.S.; Von Dossow, D.; Barbosa, J.M.C.; Paz, C.; Burgos, V.; Menna-Barreto, R.F.S. Trypanocidal activity of natural sesquiterpenoids involves mitochondrial dysfunction, ROS production and autophagic phenotype in trypanosomacruzi. Molecules 2018, 23, 2800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, G.E.G.; Morais, T.R.; Gomes, K.d.S.; Costa-Silva, T.A.; Tempone, A.G.; Lago, J.H.G.; Caseli, L. Antitrypanosomal activity of epi-polygodial from Drimys brasiliensis and its effects in cellular membrane models at the air-water interface. Bioorg. Chem. 2019, 84, 186–191. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, A.L.; Amaral, M.; Hasegawa, F.R.; Lago, J.H.G.; Tempone, A.G.; Sartorelli, P. (-)-T-Cadinol—A Sesquiterpene Isolated From Casearia sylvestris (Salicaceae)—Displayed In Vitro Activity and Causes Hyperpolarization of the Membrane Potential of Trypanosoma cruzi. Front. Pharmacol. 2021, 12, 734127. [Google Scholar] [CrossRef] [PubMed]
- Zaki, A.A.; Ashour, A.A.; Qiu, L. New sesquiterpene glycoside ester with antiprotozoal activity from the flowers of Calendula officinalis L. Nat. Prod. Res. 2021, 35, 5250–5254. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, M.; Parai, D.; Dhar, P.; Roy, M.; Barik, R.; Chattopadhyay, S.; Mukherjee, S.K. Andrographolide induces oxidative stress-dependent cell death in unicellular protozoan parasite Trypanosoma brucei. Acta Trop. 2017, 176, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Ueno, A.K.; Barcellos, A.F.; Costa-Silva, T.A.; Mesquita, J.T.; Ferreira, D.D.; Tempone, A.G.; Romoff, P.; Antar, G.M.; Lago, J.H.G. Antitrypanosomal activity and evaluation of the mechanism of action of diterpenes from aerial parts of Baccharis retusa (Asteraceae). Fitoterapia 2018, 125, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Rocha, A.C.F.S.; Morais, G.O.; da Silva, M.M.; Kovatch, P.Y.; Ferreira, D.S.; Esperandim, V.R.; Pagotti, M.C.; Magalhães, L.G.; Heleno, V.C.G. In vitro anti-trypanosomal potential of kaurane and pimarane semi-synthetic derivatives. Nat. Prod. Res. 2020, 36, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Kuźma; Kaiser, M.; Wysokińska, H. The production and antiprotozoal activity of abietane diterpenes in Salvia austriaca hairy roots grown in shake flasks and bioreactor. Prep. Biochem. Biotechnol. 2017, 47, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, M.S.; Da Costa, F.B.; Brun, R.; Kaiser, M.; Schmidt, T.J. ent-pimarane and ent-kaurane diterpenes from Aldama discolor (Asteraceae) and their antiprotozoal activity. Molecules 2016, 21, 1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirttimaa, M.; Nasereddin, A.; Kopelyanskiy, D.; Kaiser, M.; Yli-Kauhaluoma, J.; Oksman-Caldentey, K.M.; Brun, R.; Jaffe, C.L.; Moreira, V.M.; Alakurtti, S. Abietane-Type Diterpenoid Amides with Highly Potent and Selective Activity against Leishmania donovani and Trypanosoma cruzi. J. Nat. Prod. 2016, 79, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Farimani, M.M.; Khodaei, B.; Moradi, H.; Aliabadi, A.; Ebrahimi, S.N.; De Mieri, M.; Kaiser, M.; Hamburger, M. Phytochemical Study of Salvia leriifolia Roots: Rearranged Abietane Diterpenoids with Antiprotozoal Activity. J. Nat. Prod. 2018, 81, 1384–1390. [Google Scholar] [CrossRef] [PubMed]
- Zadali, R.; Nejad Ebrahimi, S.; Tofighi, Z.; Es-haghi, A.; Hamburger, M.; Kaiser, M.; D’ Ambola, M.; De Tommasi, N.; Hadjiakhoondi, A. Antiprotozoal activity of diterpenoids isolated from Zhumeria majdae- absolute configuration by circular dichroism. DARU J. Pharm. Sci. 2020, 28, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Tabefam, M.; Farimani, M.M.; Danton, O.; Ramseyer, J.; Kaiser, M.; Ebrahimi, S.N.; Salehi, P.; Batooli, H.; Potterat, O.; Hamburger, M. Antiprotozoal Diterpenes from Perovskia abrotanoides. Planta Med. 2018, 84, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Nvau, J.B.; Alenezi, S.; Ungogo, M.A.; Alfayez, I.A.M.; Natto, M.J.; Gray, A.I.; Ferro, V.A.; Watson, D.G.; de Koning, H.P.; Igoli, J.O. Antiparasitic and Cytotoxic Activity of Bokkosin, A Novel Diterpene-Substituted Chromanyl Benzoquinone From Calliandra portoricensis. Front. Chem. 2020, 8, 574103. [Google Scholar] [CrossRef] [PubMed]
- Anyam, J.V.; Daikwo, P.E.; Ungogo, M.A.; Nweze, N.E.; Igoli, N.P.; Gray, A.I.; De Koning, H.P.; Igoli, J.O. Two New Antiprotozoal Diterpenes From the Roots of Acacia nilotica. Front. Chem. 2021, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Catteau, L.; Schioppa, L.; Beaufay, C.; Girardi, C.; Hérent, M.F.; Frédérich, M.; Quetin-Leclercq, J. Antiprotozoal activities of Triterpenic Acids and Ester Derivatives Isolated from the Leaves of Vitellaria paradoxa. Planta Med. 2021, 87, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Labib, R.; Ebada, S.; Youssef, F.; Ashour, M.; Ross, S. Ursolic acid, a natural pentacylcic triterpene from Ochrosia elliptica and its role in the management of certain neglected tropical diseases. Pharmacogn. Mag. 2016, 12, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Osman, A.G.; Ali, Z.; Fantoukh, O.; Raman, V.; Kamdem, R.S.T.; Khan, I. Glycosides of ursane-type triterpenoid, benzophenone, and iridoid from Vangueria agrestis (Fadogia agrestis) and their anti-infective activities. Nat. Prod. Res. 2020, 34, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Meira, C.S.; Barbosa-Filho, J.M.; Lanfredi-Rangel, A.; Guimarães, E.T.; Moreira, D.R.M.; Soares, M.B.P. Antiparasitic evaluation of betulinic acid derivatives reveals effective and selective anti-Trypanosoma cruzi inhibitors. Exp. Parasitol. 2016, 166, 108–115. [Google Scholar] [CrossRef]
- Sousa, P.L.; da Silva Souza, R.O.; Tessarolo, L.D.; de Menezes, R.R.P.P.B.; Sampaio, T.L.; Canuto, J.A.; Martins, A.M.C. Betulinic acid induces cell death by necrosis in Trypanosoma cruzi. Acta Trop. 2017, 174, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Steverding, D.; Sidjui, L.S.; Ferreira, É.R.; Ngameni, B.; Folefoc, G.N.; Mahiou-Leddet, V.; Ollivier, E.; Stephenson, G.R.; Storr, T.E.; Tyler, K.M. Trypanocidal and leishmanicidal activity of six limonoids. J. Nat. Med. 2020, 74, 606–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carothers, S.; Nyamwihura, R.; Collins, J.; Zhang, H.; Park, H.; Setzer, W.N.; Ogungbe, I.V. Bauerenol acetate, the pentacyclic triterpenoid from Tabernaemontana longipes, is an antitrypanosomal agent. Molecules 2018, 23, 355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabefam, M.; Farimani, M.M.; Danton, O.; Ramseyer, J.; Nejad Ebrahimi, S.; Neuburger, M.; Kaiser, M.; Salehi, P.; Potterat, O.; Hamburger, M. Antiprotozoal Isoprenoids from Salvia hydrangea. J. Nat. Prod. 2018, 81, 2682–2691. [Google Scholar] [CrossRef] [PubMed]
- Muganza, D.M.; Fruth, B.; Nzunzu, J.L.; Tuenter, E.; Foubert, K.; Cos, P.; Maes, L.; Kanyanga, R.C.; Exarchou, V.; Apers, S.; et al. In vitro antiprotozoal activity and cytotoxicity of extracts and isolated constituents from Greenwayodendron suaveolens. J. Ethnopharmacol. 2016, 193, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Szabó, L.U.; Kaiser, M.; Mäser, P.; Schmidt, T.J. Antiprotozoal nor-triterpene alkaloids from Buxus sempervirens L. Antibiotics 2021, 10, 696. [Google Scholar] [CrossRef]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug repurposing: Progress, challenges and recommendations. Nat. Rev. Drug Discov. 2018, 18, 41–58. [Google Scholar] [CrossRef]
- Babaei, G.; Aliarab, A.; Abroon, S.; Rasmi, Y.; Aziz, S.G.G. Application of sesquiterpene lactone: A new promising way for cancer therapy based on anticancer activity. Biomed. Pharmacother. 2018, 106, 239–246. [Google Scholar] [CrossRef]
- Moujir, L.; Callies, O.; Sousa, P.M.C.; Sharopov, F.; Seca, A.M.L. Applications of sesquiterpene lactones: A review of some potential success cases. Appl. Sci. 2020, 10, 3001. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
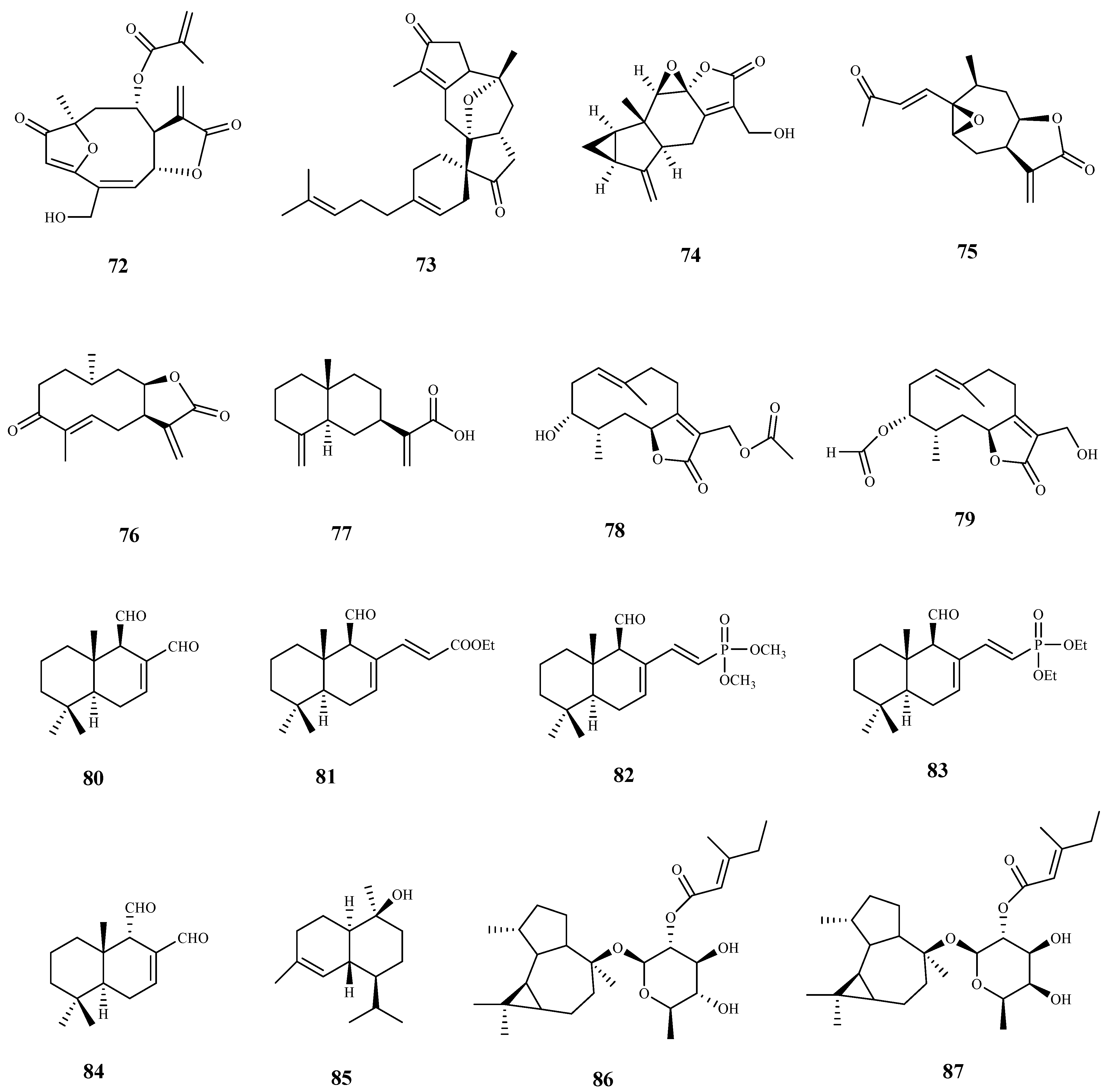{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound a | Plant | Parasite (Form) b | IC50 (µM) | SI | Refs. |
|---|---|---|---|---|---|
| Monoterpenes and Iridoids | |||||
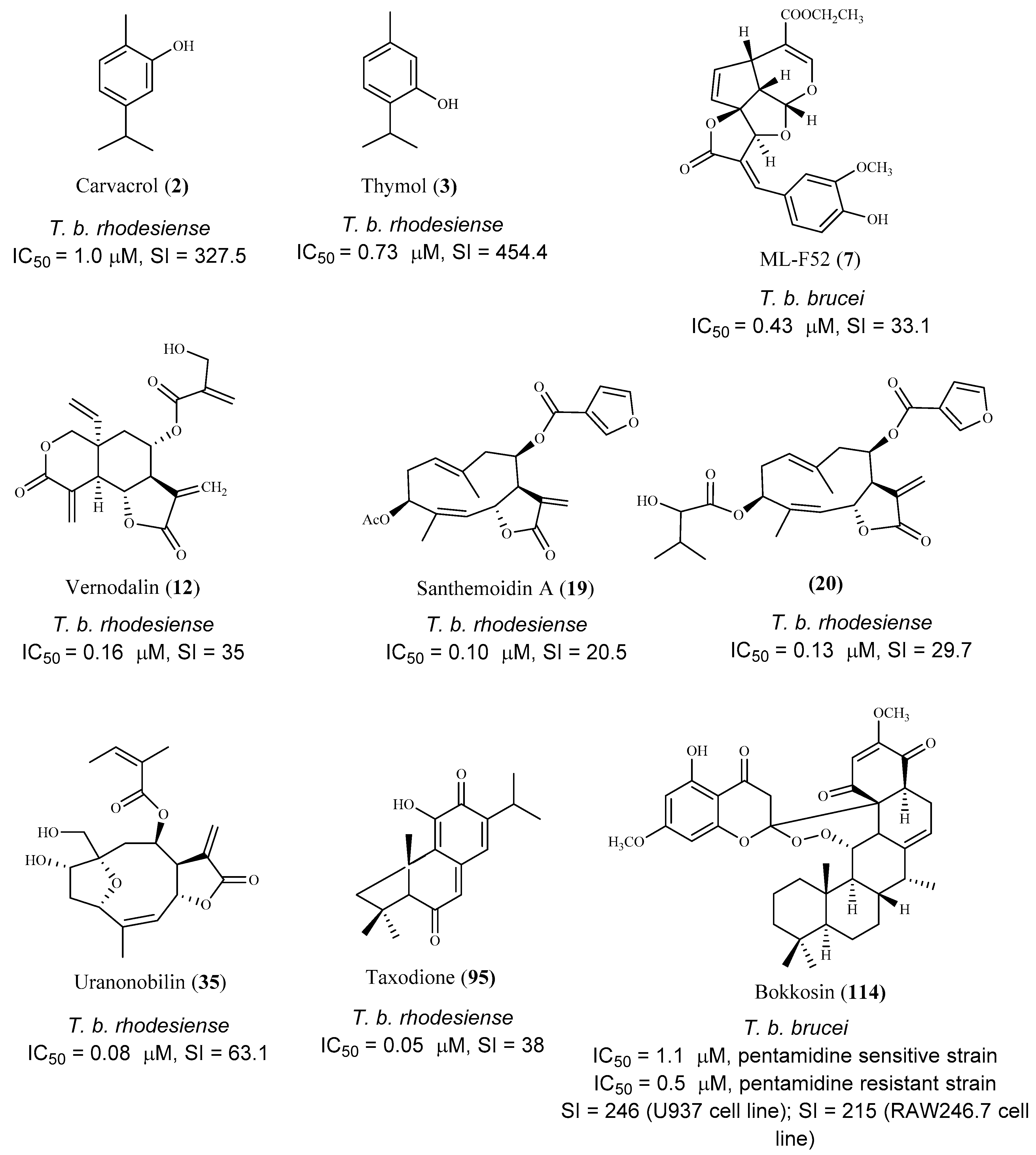
| carvacrol (2) | Origanum onites L. | T. b. rhodesiense (tryp.) | 1.0 | 327.5 | [54] |
| thymol (3) | Origanum onites L. | T. b. rhodesiense (tryp.) | 0.73 | 454.4 | [54] |
| terpinolene (4) | Crithmum maritimum L. | T. brucei (tryp.) | 0.26 | 180.0 | [55] |
| α-pinene (5) | Crithmum maritimum L. | T. brucei (tryp.) | 7.4 | >100 | [55] |
| β-ocimene (6) | Crithmum maritimum L. | T. brucei (tryp.) | 8.0 | >91 | [55] |
| ML-F52 (7) | Morinda lucida Benth. | T. b. brucei (tryp.) | 0.43 | 33.1 | [56] |
| molucidin (8) | Morinda lucida Benth. | T. b. brucei (tryp.) | 1.27 | 3.7 | [56] |
| ML-2-3 (9) | Morinda lucida Benth. | T. b. brucei (tryp.) | 3.75 | >13.3 | [56] |
| Sesquiterpenes | |||||
| tagitinin C (10) | Tithonia diversifolia (Hemsl) A. Grey | T. brucei (tryp.) | 0.012 | 3.0 | [57] |
| tagitinin A (11) | Tithonia diversifolia (Hemsl) A. Grey | T. brucei (tryp.) | 0.97 | 1.3 | [57] |
| vernodalin (12) | Vernonia cinerascens Sch.Bip. | T. b. rhodesiense (tryp.) | 0.16 | 35.0 | [58,59] |
| 11β, 13-dihydrovernolide (13) | Vernonia cinerascens Sch.Bip. | T. b. rhodesiense (tryp.) | 0.5 | 13.0 | [58,59] |
| 11β, 13-dihydrovernodaline (14) | Vernonia cinerascens Sch.Bip. | T. b. rhodesiense (tryp.) | 1.1 | 4.2 | [58,59] |
| vernocinerascolide (15) | Vernonia cinerascens Sch.Bip. | T. b. rhodesiense (tryp.) | 4.8 | 27.0 | [58,59] |
| 11β, 13-dihydrohydroxtvernolide (16) | Vernonia cinerascens Sch.Bip. | T. b. rhodesiense (tryp.) | 5.0 | 4.3 | [58,59] |
| parthenolide (17) | Tarchonanthus camphoratus L. | T. b. rhodesiense (tryp.) | 0.39 | 18.6 | [60] |
| 3-oxo-1,2-dehydrocostic acid (18) | Tarchonanthus camphoratus L. | T. b. rhodesiense (tryp.) | 2.8 | 6.2 | [60] |
| santhemoidin A (19) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 0.10 | 20.5 | [60] |
| 3β-(2″-hydroxyisovaleroyloxy)-8β-(3-furoyloxy)costunolide (20) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 0.13 | 29.7 | [60] |
| 2″-dehydroeucannabinolidesemiacetal (21) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 0.35 | 11.5 | [60] |
| 3-desacetyl-3-isovaleroyleu-cannabinolide (22) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 0.52 | 13.0 | [60] |
| 3-oxo-4β,15-dihydroliqustrin-[4′,5′-dihydroxytigloyloxy] (23) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 0.60 | 19.2 | [60] |
| schkuhrin II (24) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 0.82 | 13.4 | [60] |
| 1(10)-epoxy-3β-hydroxy-8β-[5′-hydroxy-4′-(2″- hydroxyisovaleroyloxy)tigloyloxy]costunolide (25) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 0.91 | 15.8 | [60] |
| eucannabinolide (26) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 0.92 | 15.8 | [60] |
| 2′(3′)-Z-eucannabinolide (27) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 1.7 | 31.1 | [60] |
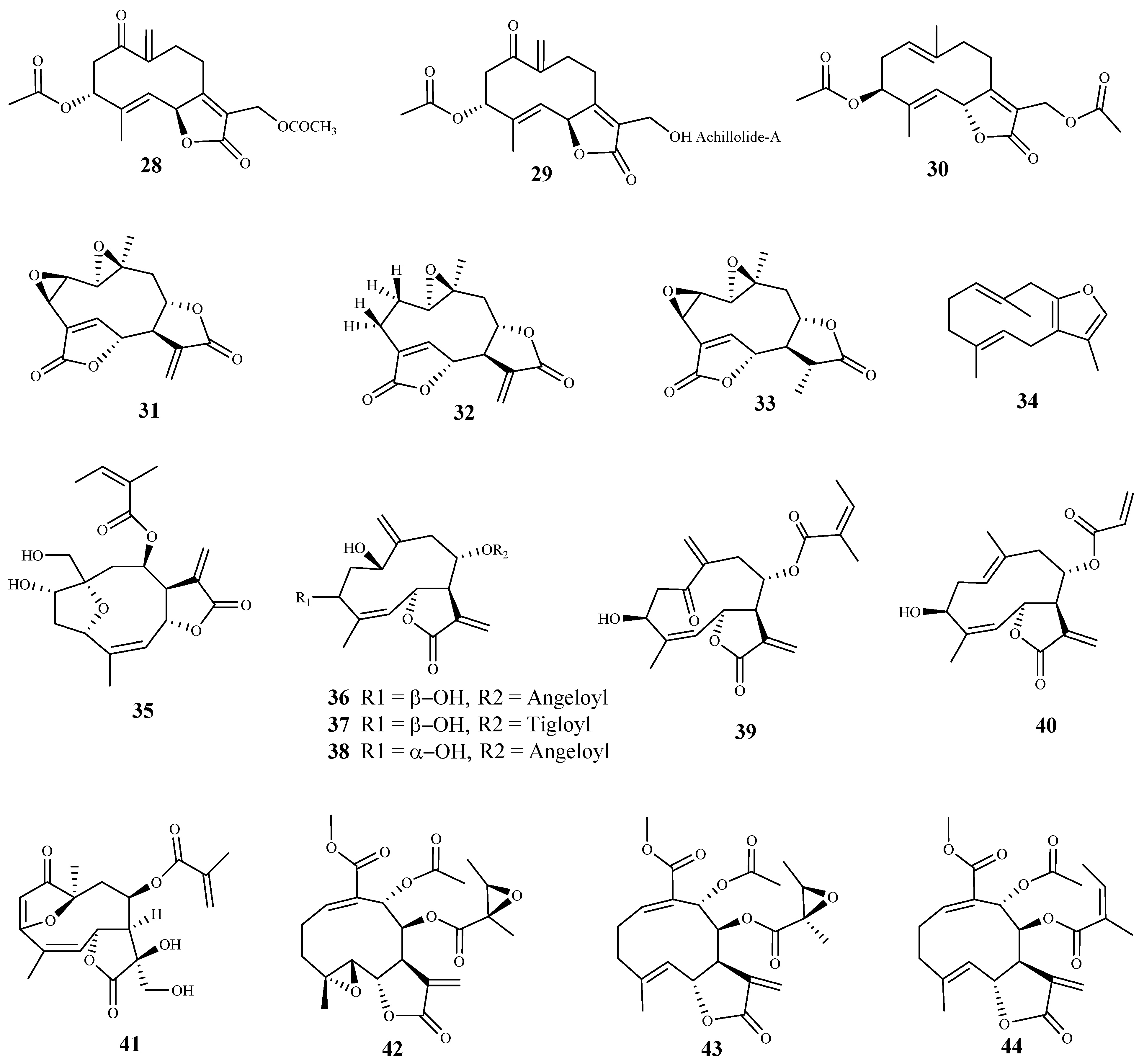
| 28 | Achillea fragrantissima (Forssk.) Sch.Bip. | T. b. brucei (tryp.) | 3.03 | n.d | [61] |
| 29 | Achillea fragrantissima (Forssk.) Sch.Bip. | T. b. brucei (tryp.) | 10.97 | n.d | [61] |
| 30 | Achillea fragrantissima (Forssk.) Sch.Bip. | T. b. brucei (tryp.) | 10.97 | n.d | [61] |
| isofuranodiene (34) | Smyrnium olusatrum L. | T. brucei (tryp.) | 3.0 | 30.0 | [63] |
| furanonobilin (35) | Anthemis nobilis L. | T. b. rhodesiense (tryp.) | 0.08 | 63.1 | [64] |
| hydroxyisonobilin (36) | Anthemis nobilis L. | T. b. rhodesiense (tryp.) | 0.61 | 8.3 | [64] |
| 8-tigloylhydroxyisonobilin (37) | Anthemis nobilis L. | T. b. rhodesiense (tryp.) | 0.36 | 14.1 | [64] |
| 3-epi-hydroxyisonobilin (38) | Anthemis nobilis L. | T. b. rhodesiense (tryp.) | 0.88 | 8.3 | [64] |
| nobilinon A (39) | Anthemis nobilis L. | T. b. rhodesiense (tryp.) | 0.4 | 3.8 | [64] |
| 45 | Scleria striatinux De Wild. | T. b. rhodesiense (tryp.) | 0.002 | 8.3 | [67] |
| 46 | Scleria striatinux De Wild. | T. b. rhodesiense (tryp.) | 0.025 | 3.4 | [67] |
| vernolepin (47) | Vernonia lasiopus (O.Hoffm.) H.Rob. | T. b. rhodesiense (tryp.) | 0.185 | 14.5 | [59] |
| vernodalol (48) | Vernonia lasiopus (O.Hoffm.) H.Rob. | T. b. rhodesiense (tryp.) | 0.26 | 14.4 | [59] |
| vernomenin (49) | Vernonia lasiopus (O.Hoffm.) H.Rob. | T. b. rhodesiense (tryp.) | 0.51 | 4.5 | [59] |
| 8-desacylvernodalol (50) | Vernonia lasiopus (O.Hoffm.) H.Rob. | T. b. rhodesiense (tryp.) | 2.53 | 13.7 | [59] |
| 4,15-iso-atriplicolide methacrylate (56) | Helianthus tuberosus L. | T. b. rhodesiense (tryp.) | 0.077 | 6.7 | [70] |
| 4,15-iso-atriplicolide isobutryrate (57) | Helianthus tuberosus L. | T. b. rhodesiense (tryp.) | 0.26 | 3.38 | [70] |
| heliantuberolide-8-O-tiglate is (58) | Helianthus tuberosus L. | T. b. rhodesiense (tryp.) | 0.92 | 4.24 | [70] |
| 78 | Tanacetum sonbolii Mozaff. | T. b. rhodesiense (tryp.) | 5.1 | 3.9 | [77] |
| 79 | Tanacetum sonbolii Mozaff. | T. b. rhodesiense (tryp.) | 10.2 | 4.0 | [77] |
| Diterpenes | |||||
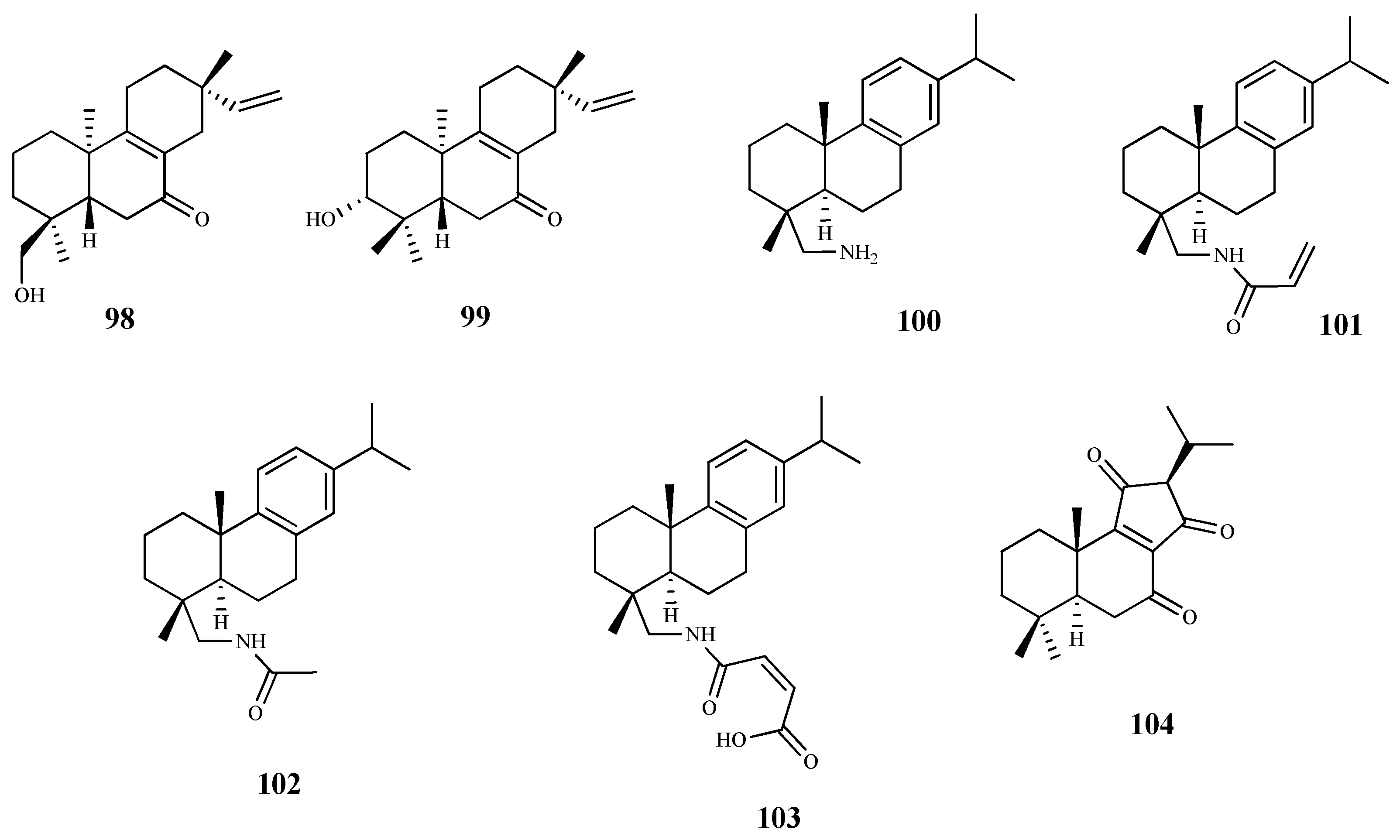
| andrographolide (88) | Andrographis paniculate (Burm. F.) Wall. Ex Nees | T. brucei procyclic trypomastigotes | 8.3 | 8.5 | [83] |
| taxodione (95) | Salvia austriaca Jacq. | T. b. rhodesiense (tryp.) | 0.05 | 38.0 | [86] |
| 7-(20-oxohexyl)-taxodione (96) | Salvia austriaca Jacq. | T. b. rhodesiense (tryp.) | 0.62 | 5.0 | [86] |
| taxodone (97) | Salvia austriaca Jacq. | T. b. rhodesiense (tryp.) | 1.67 | 2.4 | [86] |
| ent-7-oxo-pimara-8,15-diene-18-ol (98) | Aldama discolors (Baker) E.E.Schill. & Panero | T. b. rhodesiense (tryp.) | 24.3 | 2.0 | [87] |
| leriifolione (104) | Salvia leriifolia Benth. | T. b. rhodesiense (tryp.) | 1.0 | 2.6 | [89] |
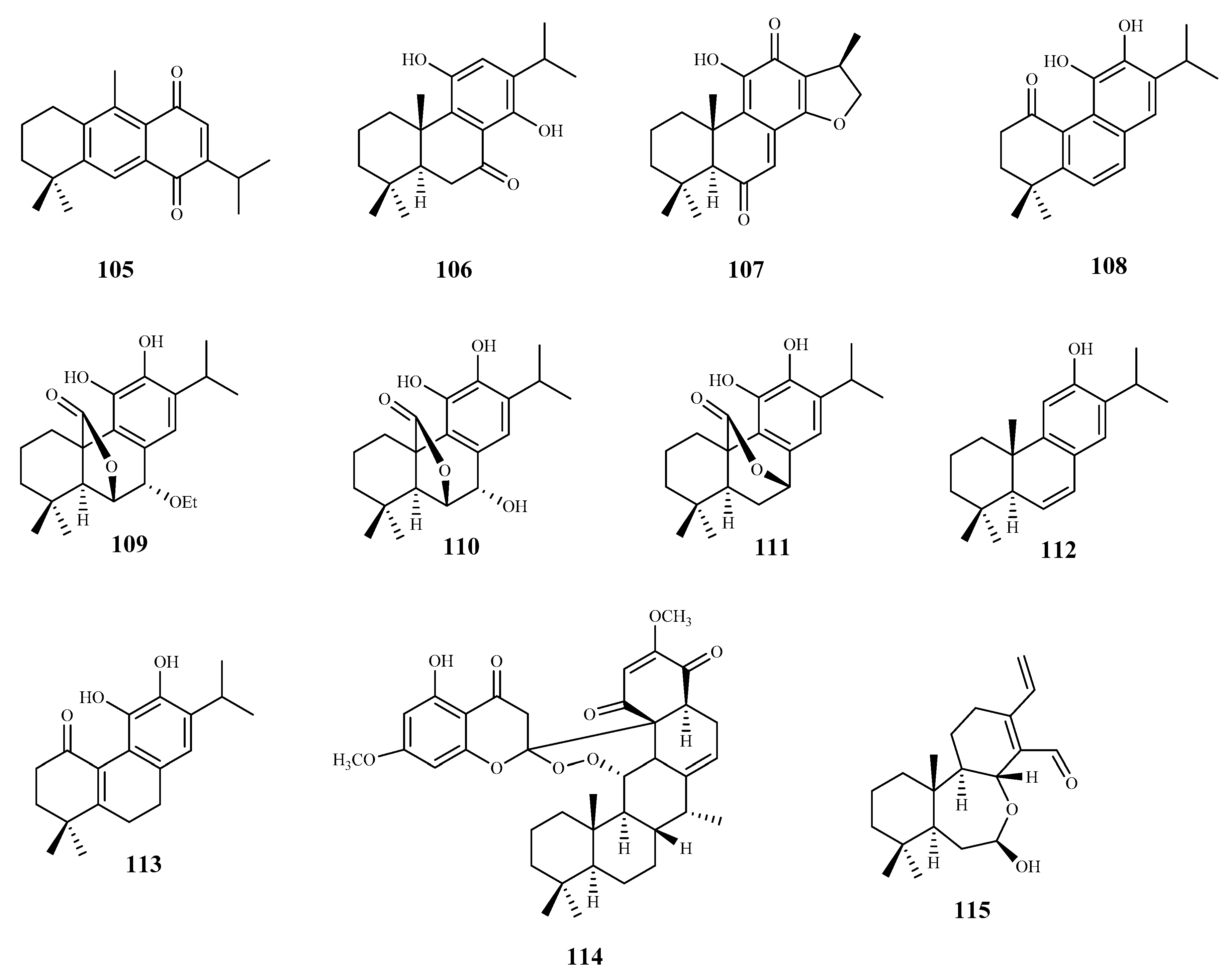
| 12, 16-dideoxy aegyptinone B (105) | Zhumeria majdae Rech. F | T. b. rhodesiense (tryp.) | 3.6 | 1.7 | [90] |
| 11,14-dihydroxy-8, 11,13- abietatrien-7-one (106) | Zhumeria majdae Rech. F | T. b. rhodesiense (tryp.) | 1.8 | 21.9 | [90] |
| lanugon Q (107) | Zhumeria majdae Rech. F | T. b. rhodesiense (tryp.) | 0.1 | 15.4 | [90] |
| miltiodiol (108) | Perovskia abrotanoides Kar. | T. b. rhodesiense (tryp.) | 0.5 | 10.5 | [91] |
| 7α-ethoxyrosmanol (109) | Perovskia abrotanoides Kar. | T. b. rhodesiense (tryp.) (trypomastigote) | 0.8 | 14.9 | [91] |
| rosmanol (110) | Perovskia abrotanoides Kar. | T. b. rhodesiense (tryp.) | 3.8 | 1.5 | [91] |
| carnosol (111) | Perovskia abrotanoides Kar. | T. b. rhodesiense (tryp.) | 5.4 | 2.4 | [91] |
| Δ9-dehydro-ferruginol (112) | Perovskia abrotanoides Kar. | T. b. rhodesiense (tryp.) | 7.2 | 12.3 | [91] |
| 11,12-dihydroxy-20-norabieta-5(10),8,11,13-tetraen-1-one (113) | Perovskia abrotanoides Kar. | T. b. rhodesiense (tryp.) | 12.0 | 12.5 | [91] |
| bokkosin (114) | Calliandra portoricensis Hassk. | T. b. brucei ((ryp.) | 1.1 | 246 | [92] |
| 8-oxacassa-13,15-dien-7-ol-17-al (115) | Acacia nilotica L. | T. b. brucei (tryp.) | 1.4 | 21.1 | [93] |
| Triterpenes | |||||
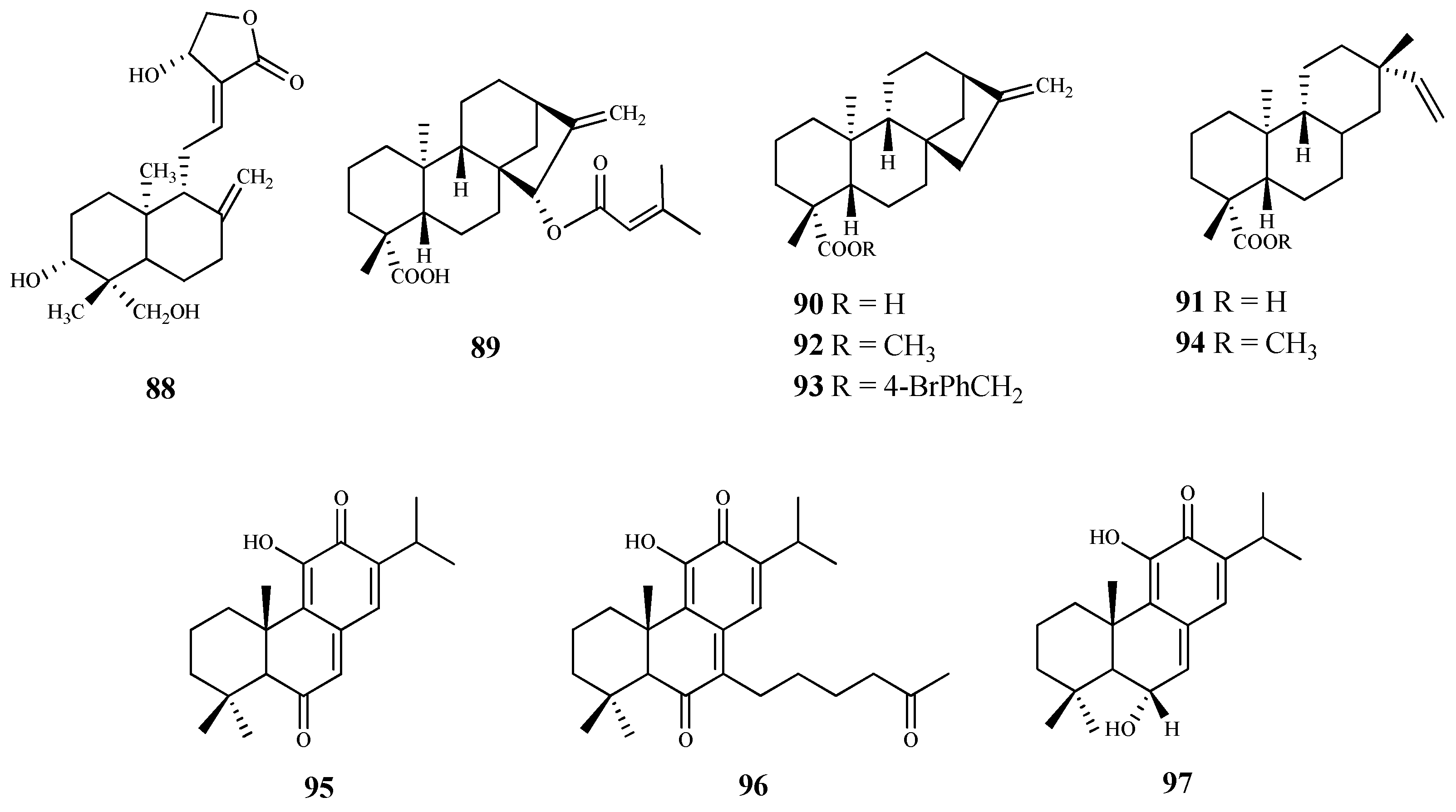
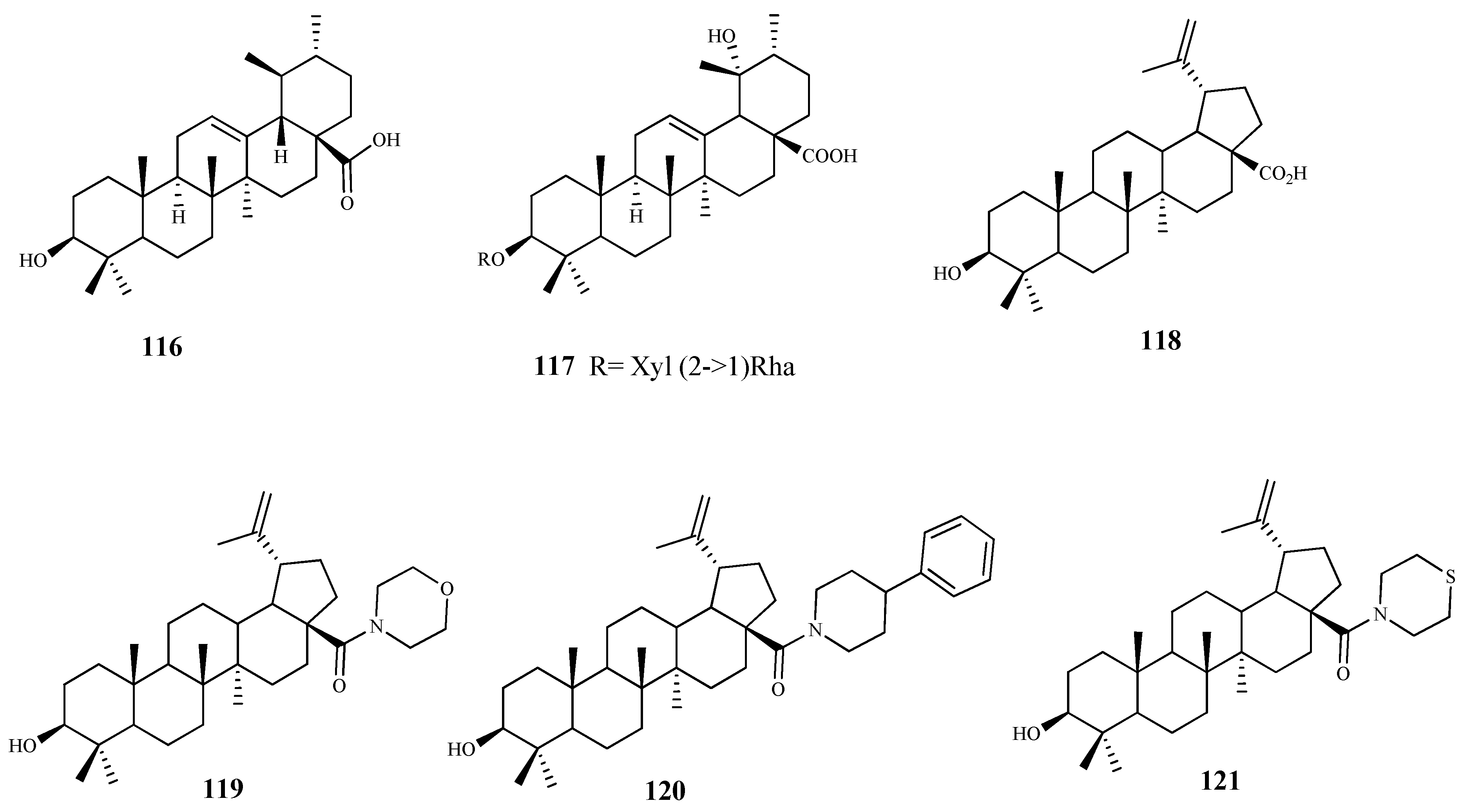
| ursolic acid (116) | Vitellaria paradoxa C. F. Gaertn | T. brucei (tryp.) | 2.4 | 4.6 | [94] |
| 3-O-[α-L-rhamnopyranosyl-(1→2)-β-D- xylopyranosyl]pomolic acid (117) | Vangueria agrestis (Schweinf. ex Hiern) Lantz | T. brucei (tryp.) | 11.1 | n.d. | [96] |
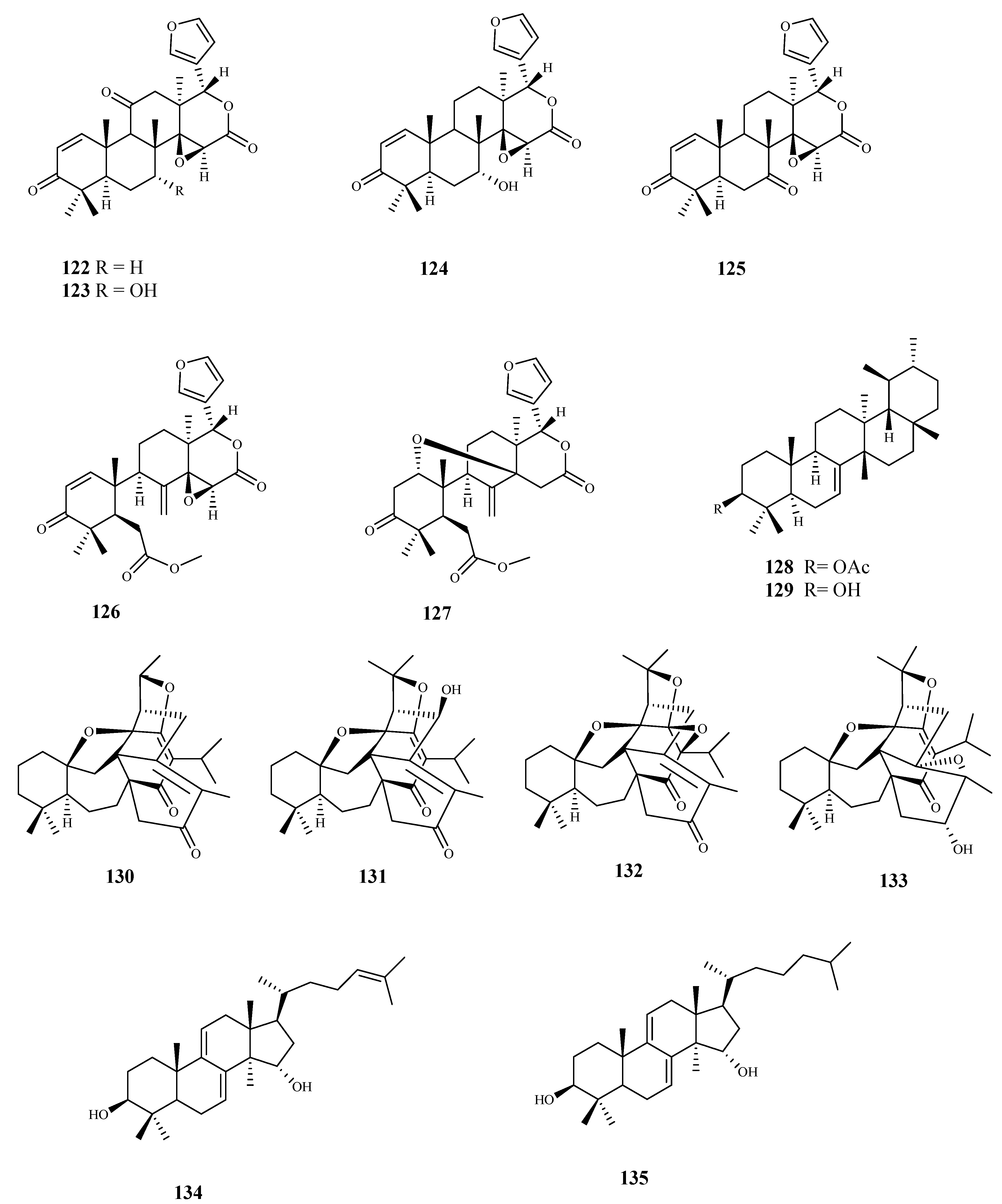
| kotschyienone A (122) | Pseudocedrela kotschyi (Schweinf.) Harms | T. brucei (tryp.) | 2.5 | 12.6 | [99] |
| 7-deacetyl-7-oxogedunin (125) | Pseudocedrela kotschyi (Schweinf.) Harms | T. brucei (tryp.) | 3.18 | >31.4 | [99] |
| baurenol acetate (128) | Tabernaemontana longipes Donn.Sm. | T. brucei (tryp.) | 3.1 | >25.8 | [100] |
| baurenol (129) | Tabernaemontana longipes Donn.Sm. | T. brucei (tryp.) | 2.7 | >29.6 | [100] |
| polycarpol (134) | Greenwayodendron suaveolens (Engl. & Diels) Verdc. | T. b. brucei (tryp.) | 8.1 | 1.0 | [102] |
| dihydropolycarpol (135) | Greenwayodendron suaveolens (Engl. & Diels) Verdc. | T. b. brucei (tryp.) | 8.1 | 2.4 | [102] |
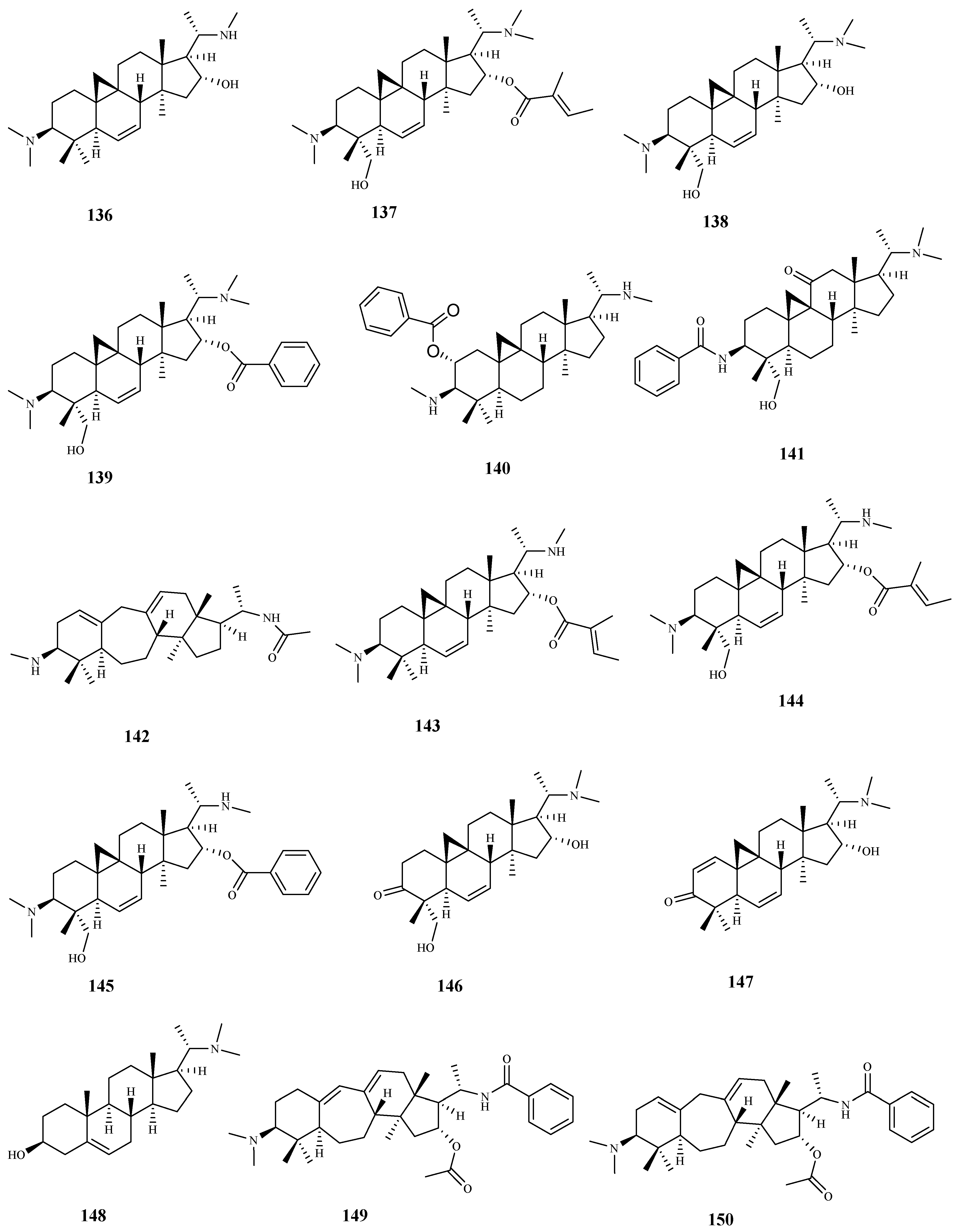
| cyclovirobuxeine-B (136) | Buxus sempervirens L. | T. b. rhodesiense (tryp.) | 1.5 | 25.0 | [103] |
| cyclomicrophylline-A (138) | Buxus sempervirens L. | T. b. rhodesiense (tryp.) | 2.3 | 42.0 | [103] |
| N-benzoyl-O-acetyl-cycloxo-buxoline-F (141) | Buxus sempervirens L. | T. b. rhodesiense (tryp.) | 2.4 | 30.0 | [103] |
| N20-acetylbuxadine- G (142) | Buxus sempervirens L. | T. b. rhodesiense (tryp.) | 1.3 | 33.0 | [103] |
| O-benzoyl-cycloprotobuxoline-D (146) | Buxus sempervirens L. | T. b. rhodesiense (tryp.) | 1.1 | 12.0 | [103] |
| Compound a | Plant | Parasite Form | IC50 (µM) | SI | Refs. |
|---|---|---|---|---|---|
| Monoterpenes and Iridoids | |||||
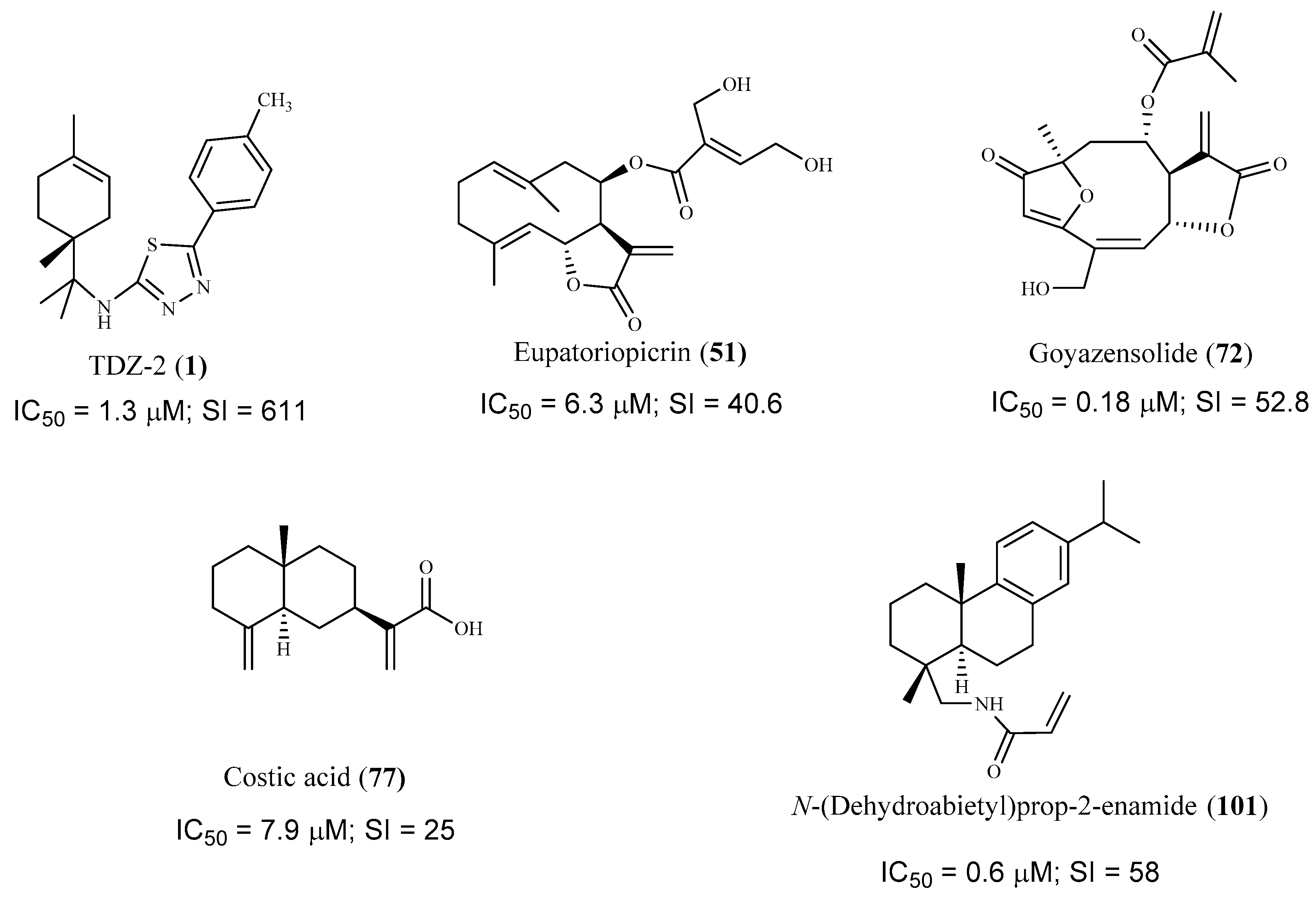
| N-{1-methyl-1-[(1R)-4-methylcyclohex-3-en-1- yl]ethyl}-5-(4-methylphenyl)-1,3,4-thiadiazol-2-amine (1) | n.d | Amastigote | 1.3 | 611.2 | [53] |
| Sesquiterpenes | |||||
| mikanolide (31) | Mikania variifolia Hieron. and Mikania micrantha Kunth | Epimastigote Trypomastigote Amastigote | 2.41 7.24 15.5 | 31.9 10.6 4.3 | [62] |
| deoxymikanolide (32) | Mikania variifolia Hieron. and Mikania micrantha Kunth | Epimastigote Trypomastigote Amastigote | 0.29 5.43 22.8 | 992.5 54.0 12.5 | [62] |
| dihydromikanolide (33) | Mikania micrantha Kunth | Epimastigote Trypomastigote Amastigote | 8.55 1.03 29.1 | 5.2 49.0 1.5 | [62] |
| nobilinon A (39) | Anthemis nobilis L. | Intracellular amastigote | 2.8 | 0.5 | [64] |
| nobilinon B (40) | Anthemis nobilis L. | Intracellular amastigote | 4.2 | 6.1 | [64] |
| 11,13-dihydroxy-calaxin (41) | Calea pinnatifida (R. Br.) Less. | Amastigote | 8.30 | 1.88 | [65] |
| enhydrin (42) | Smallanthus sonchifolius (Poepp.) H. Rob. | Epimastigote | 0.78 | n.d. | [66] |
| uvedalin (43) | Smallanthus sonchifolius (Poepp.) H. Rob. | Epimastigote | 0.79 | n.d. | [66] |
| polymatin (44) | Smallanthus sonchifolius (Poepp.) H. Rob. | Epimastigote | 1.38 | n.d. | [66] |
| 45 | Scleria striatinux De Wild. | Amastigote | 0.025 | 0.74 | [67] |
| 46 | Scleria striatinux De Wild. | Amastigote | 0.085 | 1.0 | [67] |
| eupatoriopicrin (51) | Astearaceae species | Trypomastigote bloodstream form Intracellular amastigote | 19.9 6.3 | 12.9 40.6 | [68] |
| estafietin (52) | Astearaceae species | Trypomastigote bloodstream form | 117 | 6.8 | [68] |
| eupahakonenin B (53) | Astearaceae species | Trypomastigote bloodstream form Intracellular amastigote | 33.0 89.3 | 10.4 3.8 | [68] |
| minimolide (54) | Astearaceae species | Trypomastigote bloodstream form Intracellular amastigote | 21.0 25.1 | 12.8 10.7 | [68] |
| tagitinin C (55) | Tithonia diversifolia (Hemsl.) A. Gray | Epimastigote | 1.15 | 5.69 | |
| 4,15-iso-atriplicolide methacrylate (56) | Helianthus tuberosus L. | Trypomastigote | 1.6 | 0.32 | [73] |
| 4,15-iso-atriplicolide isobutryrate (57) | Helianthus tuberosus L. | Trypomastigote | 3.1 | 0.28 | [73] |
| heliantuberolide-8-O-tiglate (58) | Helianthus tuberosus L. | Trypomastigote | 5.7 | 0.68 | [73] |
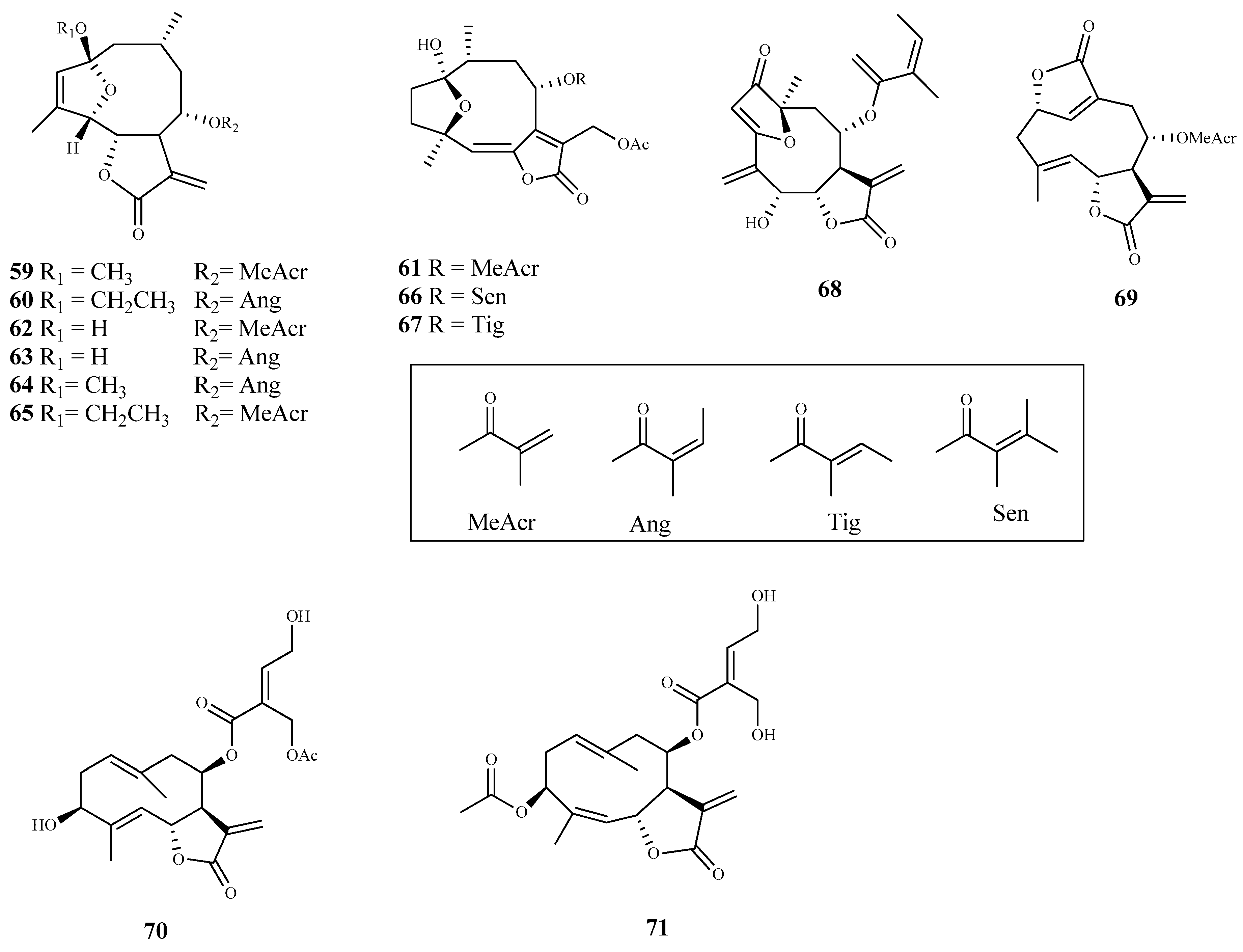
| (2-methoxy-2,5-epoxy-8-metha- cryloxygermacra-3Z,11(13)-dien-6,12-olide (59) | Vernonanthura nebularum (Cabrera) H. Rob. | Epimastigote | 1.5 | > 14 | [71] |
| (2-ethoxy-2, 5-epoxy-8-angeloxygermacra-3Z,11(13)-dien-6,12-olide (60) | Vernonanthura nebularum (Cabrera) H. Rob. | Epimastigote | 2.1 | > 14 | [71] |
| 8a-methacryloxyhirsutinolide 13-O-acetate (61) | Vernonanthura nebularum (Cabrera) H. Rob. | Epimastigote | 2.0 | > 14 | [71] |
| 62 | Vernonanthura nebularum (Cabrera) H. Rob. | Epimastigote | 3.7 | 14.3 | [71] |
| 66 | Vernonanthura nebularum (Cabrera) H. Rob. | Epimastigote | 10.7 | 9.0 | [71] |
| 67 | Vernonanthura nebularum (Cabrera) H. Rob. | Epimastigote | 8.1 | 13.9 | [71] |
| 68 | Centratherum puctatum ssp. Punctatum Cass. | Epimastigote | 6.8 | 1.6 | [71] |
| 69 | Elephantopus mollis Kunth | Epimastigote | 4.7 | 11.5 | [71] |
| eucannabinolide (70) | Urolepis hecatantha (DC.) R.King & H.Rob. | Epimastigote | 10 | 1.5 | [72] |
| santhemoidin C (71) | Urolepis hecatantha (DC.) R.King & H.Rob. | Epimastigote | 18 | 0.83 | [72] |
| goyazensolide (72) | Lychnophora passerina (Mart ex DC) Gardn. | Intracellular amastigote | 0.181/24 h 0.020/48 h | 52.82/24h 915.0/48h | [73] |
| hedyosulide (73) | Hedyosmum brasiliense Mart. ex Miq. | Trypomastigote Intracellular amastigote | 28.1 21.6 | >7 >9 | [74] |
| 8-epi-xanthatin-1β,5β-epoxide (75) | Inula viscosa (L.) Greuter | Epimastigote | 4.99 | 3.67 | [75] |
| inuloxin A (76) | Inula viscosa (L.) Greuter | Epimastigote | 15.52 | 3.38 | [75] |
| costic acid (77) | Nectandra barbellata Coe-Teix. | Intracellular amastigote | 7.9 | >25 | [76] |
| polygodial (80) | Polygonum hydropiper L. | Epimastigote Trypomastigote Amastigote | 51.0 68.2 34.4 | n.d | [79] |
| Polygodial derivative (81) | n.d | Epimastigote Trypomastigote Amastigote | 13.0 8.4 9.9 | n.d | [79] |
| Polygodial derivative (82) | n.d | Epimastigote Trypomastigote Amastigote | 12.3 6.4 6.7 | n.d | [79] |
| Polygodial derivative (83) | n.d | Epimastigote Trypomastigote Amastigote | 7.2 6.9 8.3 | n.d | [79] |
| epi-polygodial (84) | Drimys brasiliensis Miers | Trypomastigote | 5.01 | >40 | [80] |
| (-)-T-cadinol (85) | Casearia sylvestris Sw. | Trypomastigote Amastigote | 18.2 15.8 | >15 | [81] |
| Diterpenes | |||||
| ent-15β-senecioyl-oxy-kaur-16-en-19-oic acid (89) | Baccharis retusa DC. | Trypomastigote | 3.8 | 50.0 | [84] |
| taxodione (95) | Salvia austriaca Jacq. | Amastigote | 7.11 | 0.27 | [86] |
| 7-(20-oxohexyl)-taxodione (96) | Salvia austriaca Jacq. | Amastigote | 7.76 | 0.4 | [86] |
| Taxodone (97) | Salvia austriaca Jacq. | Amastigote | 7.63 | 0.5 | [86] |
| ent-7-oxo-pimara-8,15-diene-18-ol (98) | Aldama discolors (Baker) E.E.Schill. & Panero | Amastigote | 15.4 | 3.0 | [87] |
| ent-2S,4S-2-19-epoxy-pimara-8(3),15-diene-7β-ol (98) | Aldama discolors (Baker) E.E.Schill. & Panero | Amastigote | 19.4 | 4.0 | [87] |
| dehydroabietylamine derivative(103) | n.d | Amastigote | 0.6 | 58.0 | [88] |
| Leriifolione (104) | Salvia leriifolia Benth. | Amastigote | 2.6 | 0.6 | [89] |
| Triterpenes | |||||
| Betulinic acid (118) | n.d | trypomastigote | 19.5 | 18.8 | [97] |
| Betulinic acid derivative (119) | n.d | trypomastigote | 1.8 | 17.3 | [97] |
| Betulinic acid derivative (120) | n.d | trypomastigote | 5.0 | 10.7 | [97] |
| Betulinic acid derivative (121) | n.d | trypomastigote | 5.4 | 5.3 | [97] |
| Perovskone C (130) | Salvia hydrangea DC. ex Benth. | Amastigote | 3.5 | 10.7 | [101] |
| perovskone D (131) | Salvia hydrangea DC. ex Benth. | Amastigote | 3.8 | 3.6 | [101] |
| perovskone E (132) | Salvia hydrangea DC. ex Benth. | Amastigote | 11.5 | 6.3 | [101] |
| perovskone F (133) | Salvia hydrangea DC. ex Benth. | Amastigote | 19.8 | 2.4 | [101] |
| polycarpol (134) | Greenwayodendron suaveolens (Engl. & Diels) Verdc. | trypomastigote | 1.4 | 2.0 | [102] |
| dihydropolycarpol (135) | Greenwayodendron suaveolens (Engl. & Diels) Verdc. | trypomastigote | 2.4 | 8.1 | [102] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durão, R.; Ramalhete, C.; Madureira, A.M.; Mendes, E.; Duarte, N. Plant Terpenoids as Hit Compounds against Trypanosomiasis. Pharmaceuticals 2022, 15, 340. https://doi.org/10.3390/ph15030340
Durão R, Ramalhete C, Madureira AM, Mendes E, Duarte N. Plant Terpenoids as Hit Compounds against Trypanosomiasis. Pharmaceuticals. 2022; 15(3):340. https://doi.org/10.3390/ph15030340
Chicago/Turabian StyleDurão, Raquel, Cátia Ramalhete, Ana Margarida Madureira, Eduarda Mendes, and Noélia Duarte. 2022. "Plant Terpenoids as Hit Compounds against Trypanosomiasis" Pharmaceuticals 15, no. 3: 340. https://doi.org/10.3390/ph15030340
APA StyleDurão, R., Ramalhete, C., Madureira, A. M., Mendes, E., & Duarte, N. (2022). Plant Terpenoids as Hit Compounds against Trypanosomiasis. Pharmaceuticals, 15(3), 340. https://doi.org/10.3390/ph15030340