
Therapeutic Targets in Allergic Conjunctivitis

Abstract
:1. Introduction
2. Conjunctiva
3. Mediators of Allergy
4. Pathological and Immune Mechanism
5. Current Treatment Approaches
5.1. Non-Pharmacologic Options
5.2. Antihistamines
5.3. Mast Cell Stabilizers
5.4. Combination Antihistamine/Mast Cell Stabilizers
5.5. Nonsteroidal Anti-Inflammatory Drugs
5.6. Corticosteroids
5.7. Immunomodulators

{kind=link}
| Drug Class | Mechanism | Current and Potential Drug Examples and Dosage |
|---|---|---|
| Antihistamines | Block histamine receptors, thereby, preventing histamine from interacting with histamine receptors [11]. | Levocabastine 0.05% ophthalmic suspension: one drop four times per day [99,100,101]. Emedastine 0.05% ophthalmic solution: one drop four times per day [99,100,101]. Bilastine 0.6% ophthalmic formulation: one drop per day [98]. |
| Mast cell stabilizers | Inhibit mast cell degranulation through prevention of calcium mobilization in the mast cell [102]. | Lodoxamide tromethamine 0.1% ophthalmic solution: one drop four times per day [95,100,103,104]. Pemirolast potassium 0.1% ophthalmic solution: one drop four times per day [95,100,103,104]. Nedocromil sodium 2% ophthalmic solution: one drop twice per day [95,100,103,104]. Sodium cromoglycate 4% ophthalmic solution: one drop four per day [95,100,103,104]. |
| Combination antihistamines and mast cell stabilizers | Histamine receptor antagonist and prevent mast cell degranulation [11,87]. | Olopatadine ophthalmic solution (0.1% Patanol, 0.2% Pataday, 0.7% Pazeo, Novartis, Basel, Switzerland): one to two drops per day [8,11,107]. Ketotifen 0.025% ophthalmic solution (Zaditor®, Novartis): one drop twice per day [8,11,107]. Bepotastine besilate 1.5% ophthalmic solution (Bepreve®, Bausch & Lomb, Laval, QC, Canada): one drop twice per day [8,11,107]. Epinastine 0.05% ophthalmic solution: one drop twice per day [8,11,107]. Alcaftadine 0.25% ophthalmic solution: one drop per day [8,11,107]. Azelastine 0.05% ophthalmic solution: one drop twice per day [8,11,107]. |
| NSAIDs | Blocks cyclooxygenase in the inflammatory pathway which inhibits prostaglandin release [10,90,123]. | Ketorolac tromethamine 0.5% ophthalmic suspension: one drop four times per day [1,3]. |
| Corticosteroids | Controls inflammation by forming a GC/GCR complex, which is transported to the nucleus where it binds to GREs to downregulate the generation of pro-inflammatory mediators [125,126]. | Loteprednol etabonate (0.2% Alrex®, 0.5% Lotemax® suspension, Bausch & Lomb): one drop four times per day [130,131,132]. |
| Immunomodulators | Blocks IL-2 production, thereby, suppressing IL-2 mediated proliferation of T lymphocytes [137]. | Cyclosporine A (0.05%) ophthalmic emulsion: one drop twice per day [138]. Tacrolimus ophthalmic solution or ointment: twice per day [139]. |
5.8. Immunotherapy
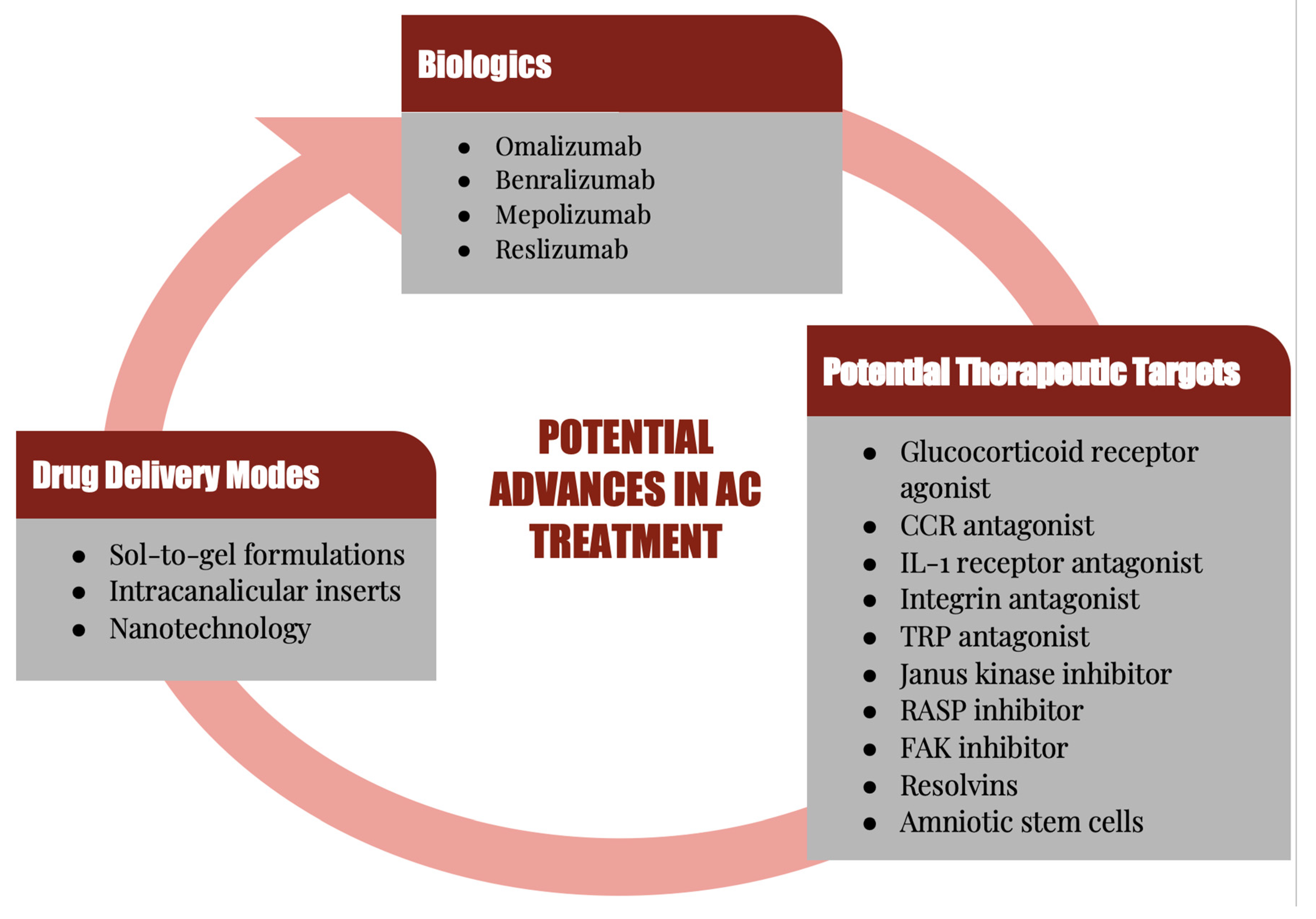
6. Potential Therapeutic Targets
6.1. Glucocorticoid Receptor Agonists
6.2. Receptor Antagonists
6.3. Transient Receptor Membrane Potential Antagonists
| Receptor Agonists | Mechanism | Outcome |
|---|---|---|
| Glucocorticoid receptor agonist | Trans-repression through interaction with activating protein 1 and nuclear factor-kappa B [3,143]. | Anti-inflammatory action with less potent side effects than steroids [143,150,151]. |
| Receptor Antagonists | Mechanism | Outcome |
| CCR3 antagonist | Prevents mast cell activation and suppresses recruitment of eosinophils [152,160]. | Suppression of clinical signs and symptoms in both early and late phase allergic responses [160]. |
| CCR2 antagonist | Reduces recruitment and subsequent accumulation of mast cell progenitors [40,74,162]. | Suppression of signs and symptoms in both early and late phase allergic responses [162]. |
| IL-1 receptor antagonist | Blocks binding of IL-1 to IL-1 receptor, causing the inhibition of IL-1-mediated inflammation [164,165]. | Reduces ocular surface inflammation [11,163]. |
| Integrin antagonist | Blocks ICAM-1 and LFA-1 interaction which inhibits Th2 cell mediated inflammation [166]. | Reduction in ocular surface inflammation, mainly in dry eye disease [11,166]. |
| Transient receptor membrane potential (TRP) antagonist | Inhibits Th2 activity in allergic disease [167,168,169]. | Clinical improvement in AC symptoms and reduced IgE levels [170]. |
6.4. Janus Kinase Inhibitors
6.5. Reactive Aldehyde Species Inhibitor
6.6. Focal Adhesion Kinase Inhibitors
6.7. Biologics
6.8. Resolvins
6.9. Amniotic Stem Cells
| Non-Receptor Antagonists | Mechanism | Outcome |
|---|---|---|
| Janus kinase inhibitor | Blocks cytokine signaling and inhibits IL-2 mediated proliferation of T cells [171,172,173]. | Significant improvement in the signs and symptoms of AC [176]. |
| Reactive aldehyde species (RASP) inhibitor | Blocks pro-inflammatory cytokines and reduces histamine levels [179,180,181,182]. | Prophylaxis and treatment of AC [183,184]. |
| Focal adhesion kinase (FAK) inhibitor | Inhibits tyrosine 397 phosphorylation, thereby, preventing cell growth, survival, and migration [60,185,188,189]. | Improvement in the signs and symptoms of allergic conjunctivitis [190]. |
| Other | Mechanism | Outcome |
| Omalizumab | Binds to Fc region of free IgE to prevent IgE binding to receptors on mast cells [191,192,193,194,195]. | Reduces redness, tearing, and inflammation [191,192]. |
| Resolvins | Endogenous component in the resolution of inflammation [201,202]. | Reducing signs and symptoms of AC [201,202]. |
| Amniotic stem cells | Inhibition of effector T cells, effector B cells, neutrophils, dendritic cells, and mast cells and the secretion of anti-inflammatory cells [203,210]. | Significant reduction in the signs and symptoms of AC [203]. |
7. Novel Delivery Methods
8. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chigbu, D.I.; Jain, P.; Khan, Z.K. Immune mechanisms, pathology, and management of allergic ocular diseases. In Advanced Concepts in Human Immunology: Prospects for Disease Control; Jain, P., Ndhlovu, L.C., Eds.; Springer Nature: Berlin, Germany, 2020; pp. 229–277. [Google Scholar]
- Chigbu, D.I. The pathophysiology of ocular allergy: A review. Cont. Lens. Anterior Eye 2009, 32, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Bielory, B.P.; O’Brien, T.P.; Bielory, L. Management of seasonal allergic conjunctivitis: Guide to therapy. Acta Ophthalmol. 2012, 90, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Rosario, N.; Bielory, L. Epidemiology of allergic conjunctivitis. Curr. Opin. Allergy Clin. Immunol. 2011, 11, 471–476. [Google Scholar] [CrossRef]
- Wong, A.; Barg, S.; Leung, A. Seasonal and perennial allergic conjunctivitis. Recent Pat. Inflamm. Allergy Drug Discov. 2009, 3, 118–127. [Google Scholar] [CrossRef]
- Wong, A.H.; Barg, S.S.; Leung, A.K. Seasonal and perennial allergic conjunctivitis. Recent Pat. Inflamm. Allergy Drug Discov. 2014, 8, 139–153. [Google Scholar] [CrossRef]
- Mimura, T.; Yamagami, S.; Kamei, Y.; Goto, M.; Matsubara, M. Specific IgE in tear fluid and features of allergic conjunctivitis. Curr. Eye Res. 2013, 38, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Chigbu, D.I.; Coyne, A.M. Update and clinical utility of alcaftadine ophthalmic solution 0.25% in the treatment of allergic conjunctivitis. Clin. Ophthalmol. 2015, 9, 1215–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elieh Ali Komi, D.; Rambasek, T.; Bielory, L. Clinical implications of mast cell involvement in allergic conjunctivitis. Allergy 2018, 73, 528–539. [Google Scholar] [CrossRef]
- La Rosa, M.; Lionetti, E.; Reibaldi, M.; Russo, A.; Longo, A.; Leonardi, S.; Tomarchio, S.; Avitabile, T.; Reibaldi, A. Allergic conjunctivitis: A comprehensive review of the literature. Ital. J. Pediatrics 2013, 39, 18. [Google Scholar] [CrossRef] [Green Version]
- Villegas, B.V.; Benitez-Del-Castillo, J.M. Current Knowledge in Allergic Conjunctivitis. Turk. J. Ophthalmol. 2021, 51, 45–54. [Google Scholar] [CrossRef]
- Rigoli, L.; Briuglia, S.; Caimmi, S.; Ferrau, V.; Gallizzi, R.; Leonardi, S.; La Rosa, M.; Salpietro, C. Gene-environment interaction in childhood asthma. Int. J. Immunopathol. Pharmacol. 2011, 24, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbee, R.A.; Kaltenborn, W.; Lebowitz, M.D.; Burrows, B. Longitudinal changes in allergen skin test reactivity in a community population sample. J. Allergy Clin. Immunol. 1987, 79, 16–24. [Google Scholar] [CrossRef]
- Maziak, W.; Behrens, T.; Brasky, T.M.; Duhme, H.; Rzehak, P.; Weiland, S.K.; Keil, U. Are asthma and allergies in children and adolescents increasing? Results from ISAAC phase I and phase III surveys in Munster, Germany. Allergy 2003, 58, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Verlato, G.; Corsico, A.; Villani, S.; Cerveri, I.; Migliore, E.; Accordini, S.; Carolei, A.; Piccioni, P.; Bugiani, M.; Lo Cascio, V.; et al. Is the prevalence of adult asthma and allergic rhinitis still increasing? Results of an Italian study. J. Allergy Clin. Immunol. 2003, 111, 1232–1238. [Google Scholar] [CrossRef] [PubMed]
- Pflugfelder, S.; Stern, M. Immunoregulation on the ocular surface: 2nd Cullen Symposium. Ocul. Surf. 2009, 7, 67–77. [Google Scholar] [CrossRef]
- Yi, X.; Wang, Y.; Yu, F.S. Corneal epithelial tight junctions and their response to lipopolysaccharide challenge. Investig. Ophthalmol. Vis. Sci. 2000, 41, 4093–4100. [Google Scholar]
- Kimura, K.; Teranishi, S.; Nishida, T. Interleukin-1beta-induced disruption of barrier function in cultured human corneal epithelial cells. Investig. Ophthalmol. Vis. Sci. 2009, 50, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Schneeberger, E.E.; Lynch, R.D. The tight junction: A multifunctional complex. Am. J. Physiol. Cell Physiol. 2004, 286, C1213–C1228. [Google Scholar] [CrossRef]
- Ohbayashi, M.; Manzouri, B.; Morohoshi, K.; Fukuda, K.; Ono, S.J. The Role of Histamine in Ocular Allergy. Adv. Exp. Med. Biol. 2010, 709, 43–52. [Google Scholar] [CrossRef]
- Forrester, J.V.; Xu, H.; Kuffova, L.; Dick, A.D.; McMenamin, P.G. Dendritic cell physiology and function in the eye. Immunol. Rev. 2010, 234, 282–304. [Google Scholar] [CrossRef]
- Irkeç, M.; Bozkurt, B. Epithelial cells in ocular allergy. Curr. Allergy Asthma Rep. 2003, 3, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Knop, E.; Knop, N. Influence of the eye-associated lymphoid tissue (EALT) on inflammatory ocular surface disease. Ocul. Surf. 2005, 3, S180–S186. [Google Scholar] [CrossRef]
- Knop, E.; Knop, N. Anatomy and immunology of the ocular surface. Chem. Immunol. Allergy 2007, 92, 36–49. [Google Scholar] [PubMed]
- Knop, E.; Knop, N. The role of eye-associated lymphoid tissue in corneal immune protection. J. Anat. 2005, 206, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Knop, N.; Knop, E. Conjunctiva-associated lymphoid tissue in the human eye. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1270–1279. [Google Scholar]
- McKenna, K.C.; Miguel, R.V. Adaptive immune system and the eye: T cell-mediated immunity. In Immunology, Inflammation and Diseases of the Eye; Dartt, D.A.D.R., D’Amore, P., Niederkorn, J.Y., Eds.; Academic Press: Boston, MA, USA, 2011; pp. 12–17. [Google Scholar]
- Calonge, M.; Enríquez-de-Salamanca, A. The role of the conjunctival epithelium in ocular allergy. Curr. Opin. Allergy Clin. Immunol. 2005, 5, 441–445. [Google Scholar] [CrossRef]
- Hingorani, M.; Calder, V.; Buckley, R.; Lightman, S. The role of conjunctival epithelial cells in chronic ocular allergic disease. Exp. Eye Res. 1998, 67, 491–500. [Google Scholar] [CrossRef]
- Kumagai, N.; Fukuda, K.; Fujitsu, Y.; Yamamoto, K.; Nishida, T. Role of structural cells of the cornea and conjunctiva in the pathogenesis of vernal keratoconjunctivitis. Prog. Retin. Eye Res. 2006, 25, 165–187. [Google Scholar] [CrossRef]
- Enriquez-de-Salamanca, A.; Castellanos, E.; Stern, M.E.; Fernandez, I.; Carreno, E.; Garcia-Vazquez, C.; Herreras, J.M.; Calonge, M. Tear cytokine and chemokine analysis and clinical correlations in evaporative-type dry eye disease. Mol. Vis. 2010, 16, 862–873. [Google Scholar]
- Asano-Kato, N.; Fukagawa, K.; Okada, N.; Kawakita, T.; Takano, Y.; Dogru, M.; Tsubota, K.; Fujishima, H. TGF-beta1, IL-1beta, and Th2 cytokines stimulate vascular endothelial growth factor production from conjunctival fibroblasts. Exp. Eye Res. 2005, 80, 555–560. [Google Scholar] [CrossRef]
- Fukuda, K.; Kumagai, N.; Fujitsu, Y.; Nishida, T. Fibroblasts as local immune modulators in ocular allergic disease. Allergol. Int. 2006, 55, 121–129. [Google Scholar] [CrossRef] [Green Version]
- Chigbu, D.I.; Labib, B.A. Immunopharmacology in Vernal Keratoconjunctivitis: Current and Future Perspectives. Pharmaceuticals 2021, 14, 658. [Google Scholar] [CrossRef] [PubMed]
- Elhers, W.H.; Donshik, P.C. Giant papillary conjunctivitis. Curr. Opin. Allergy Clin. Immunol. 2008, 8, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Kambe, N.; Hiramatsu, H.; Shimonaka, M.; Fujino, H.; Nishikomori, R.; Heike, T.; Ito, M.; Kobayashi, K.; Ueyama, Y.; Matsuyoshi, N.; et al. Development of both human connective tissue-type and mucosal-type mast cells in mice from hematopoietic stem cells with identical distribution pattern to human body. Blood 2004, 103, 860–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parham, P. IgE-mediated immunity and allergy. In The Immune System; Garland Science: New York, NY, USA, 2015; pp. 401–431. [Google Scholar]
- Brown, J.; Wilson, T.; Metcalfe, D. The mast cell and allergic diseases: Role in pathogenesis and implications for therapy. Clin. Exp. Allergy 2008, 38, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Wernersson, S.; Pejler, G. Mast cell secretory granules: Armed for battle. Nat. Rev. Immunol. 2014, 14, 478–494. [Google Scholar] [CrossRef] [PubMed]
- Gilfillan, A.M.; Austin, S.J.; Metcalfe, D.D. Mast cell biology: Introduction and overview. Adv. Exp. Med. Biol. 2011, 716, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, H.G.; Agrawal, Y.K.; Raval, H.G.; Manna, K.; Desai, P.R. Histamine H4 receptor: A novel therapeutic target for immune and allergic responses. Mini Rev. Med. Chem. 2010, 10, 1293–1308. [Google Scholar] [CrossRef]
- Saravanan, C.; Bharti, S.K.; Jaggi, S.; Singh, S.K. Histamine H(4) receptor: A novel target for inflammation therapy. Mini Rev. Med. Chem. 2011, 11, 143–158. [Google Scholar] [CrossRef]
- Criado, P.R.; Criado, R.F.; Maruta, C.W.; Machado Filho, C. Histamine, histamine receptors and antihistamines: New concepts. An. Bras. Dermatol. 2010, 85, 195–210. [Google Scholar] [CrossRef] [Green Version]
- Marson, C.M. Targeting the histamine H4 receptor. Chem. Rev. 2011, 111, 7121–7156. [Google Scholar] [CrossRef] [PubMed]
- Kmiecik, T.; Otocka-Kmiecik, A.; Gorska-Ciebiada, M.; Ciebiada, M. T lymphocytes as a target of histamine action. Arch. Med. Sci. AMS 2012, 8, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Thangam, E.B.; Jemima, E.A.; Singh, H.; Baig, M.S.; Khan, M.; Mathias, C.B.; Church, M.K.; Saluja, R. The Role of Histamine and Histamine Receptors in Mast Cell-Mediated Allergy and Inflammation: The Hunt for New Therapeutic Targets. Front. Immunol. 2018, 9, 1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galicia-Carreon, J.; Santacruz, C.; Hong, E.; Jimenez-Martinez, M.C. The ocular surface: From physiology to the ocular allergic diseases. Rev. Alerg. Mex. 2013, 60, 172–183. [Google Scholar] [PubMed]
- Hayashi, D.; Li, D.; Hayashi, C.; Shatos, M.; Hodges, R.R.; Dartt, D.A. Role of histamine and its receptor subtypes in stimulation of conjunctival goblet cell secretion. Investig. Ophthalmol. Vis. Sci. 2012, 53, 2993–3003. [Google Scholar] [CrossRef] [Green Version]
- Damaj, B.B.; Becerra, C.B.; Esber, H.J.; Wen, Y.; Maghazachi, A.A. Functional expression of H4 histamine receptor in human natural killer cells, monocytes, and dendritic cells. J. Immunol. 2007, 179, 7907–7915. [Google Scholar] [CrossRef] [Green Version]
- Jemima, E.A.; Prema, A.; Thangam, E.B. Functional characterization of histamine H4 receptor on human mast cells. Mol. Immunol. 2014, 62, 19–28. [Google Scholar] [CrossRef]
- Buckland, K.F.; Williams, T.J.; Conroy, D.M. Histamine induces cytoskeletal changes in human eosinophils via the H(4) receptor. Br. J. Pharmacol. 2003, 140, 1117–1127. [Google Scholar] [CrossRef] [Green Version]
- Leonardi, A. Role of histamine in allergic conjunctivitis. Acta Ophthalmol. Scand. Suppl. 2000, 78, 18–21. [Google Scholar] [CrossRef]
- Huang, C.C.; Kim, Y.S.; Olson, W.P.; Li, F.; Guo, C.; Luo, W.; Huang, A.J.W.; Liu, Q. A histamine-independent itch pathway is required for allergic ocular itch. J. Allergy Clin. Immunol. 2016, 137, 1267–1270.e1266. [Google Scholar] [CrossRef] [Green Version]
- Reinach, P.S.; Mergler, S.; Okada, Y.; Saika, S. Ocular transient receptor potential channel function in health and disease. BMC Ophthalmol. 2015, 15 (Suppl. S1), 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dustin, M.L.; Springer, T.A. Lymphocyte function-associated antigen-1 (LFA-1) interaction with intercellular adhesion molecule-1 (ICAM-1) is one of at least three mechanisms for lymphocyte adhesion to cultured endothelial cells. J. Cell Biol. 1988, 107, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Walling, B.L.; Kim, M. LFA-1 in T Cell Migration and Differentiation. Front. Immunol. 2018, 9, 952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, B.J.; McDowall, A.; Taylor, P.C.; Hogg, N.; Haskard, D.O.; Landis, R.C. Shedding of lymphocyte function-associated antigen-1 (LFA-1) in a human inflammatory response. Blood 2006, 107, 3593–3599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyun, Y.M.; Lefort, C.T.; Kim, M. Leukocyte integrins and their ligand interactions. Immunol. Res. 2009, 45, 195–208. [Google Scholar] [CrossRef] [Green Version]
- Marlin, S.D.; Springer, T.A. Purified intercellular adhesion molecule-1 (ICAM-1) is a ligand for lymphocyte function-associated antigen 1 (LFA-1). Cell 1987, 51, 813–819. [Google Scholar] [CrossRef]
- Kleinschmidt, E.G.; Schlaepfer, D.D. Focal adhesion kinase signaling in unexpected places. Curr. Opin. Cell Biol. 2017, 45, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Yokomizo, T. The role of leukotrienes in allergic diseases. Allergol. Int. 2015, 64, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Garceau, D.; Ford-Hutchinson, A. The role of leukotriene D4 as a mediator of allergic conjunctivitis in the guinea-pig. Eur. J. Pharmacol. 1987, 134, 285–292. [Google Scholar] [CrossRef]
- Gane, J.; Buckley, R. Leukotriene receptor antagonists in allergic eye disease: A systematic review and meta-analysis. J. Allergy Clin. Immunol. Pract. 2013, 1, 65–74. [Google Scholar] [CrossRef]
- Yamanishi, R.; Okada, N.; Shimizu, E.; Fujishima, H. Elevated levels of prostaglandin E2 in the tears of patients with severe allergic conjunctivitis and primary cultured conjunctival cells are suppressed by ketotifen and dexamethasone. BMJ Open Ophthalmol. 2021, 6, e000571. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Lee, S.H.; Kim, T.H. The Biology of Prostaglandins and Their Role as a Target for Allergic Airway Disease Therapy. Int. J. Mol. Sci. 2020, 21, 1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonardi, A.; Curnow, S.; Zhan, H.; Calder, V. Multiple cytokines in human tear specimens in seasonal and chronic allergic eye disease and in conjunctival fibroblast cultures. Clin. Exp. Allergy 2006, 36, 777–784. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, J.J.; Gadina, M.; Schreiber, R.D. Cytokine signaling in 2002: New surprises in the Jak/Stat pathway. Cell 2002, 109, S121–S131. [Google Scholar] [CrossRef] [Green Version]
- Seif, F.; Khoshmirsafa, M.; Aazami, H.; Mohsenzadegan, M.; Sedighi, G.; Bahar, M. The role of JAK-STAT signaling pathway and its regulators in the fate of T helper cells. Cell Commun. Signal. 2017, 15, 23. [Google Scholar] [CrossRef] [Green Version]
- O’Shea, J.J.; Murray, P.J. Cytokine signaling modules in inflammatory responses. Immunity 2008, 28, 477–487. [Google Scholar] [CrossRef] [Green Version]
- Leonard, W.J.; Lin, J.X. Cytokine receptor signaling pathways. J. Allergy Clin. Immunol. 2000, 105, 877–888. [Google Scholar] [CrossRef]
- Hughes, C.E.; Nibbs, R.J.B. A guide to chemokines and their receptors. FEBS J. 2018, 285, 2944–2971. [Google Scholar] [CrossRef]
- Dyer, D.P. Understanding the mechanisms that facilitate specificity, not redundancy, of chemokine-mediated leukocyte recruitment. Immunology 2020, 160, 336–344. [Google Scholar] [CrossRef]
- Collington, S.J.; Hallgren, J.; Pease, J.E.; Jones, T.G.; Rollins, B.J.; Westwick, J.; Austen, K.F.; Williams, T.J.; Gurish, M.F.; Weller, C.L. The role of the CCL2/CCR2 axis in mouse mast cell migration in vitro and in vivo. J. Immunol. 2010, 184, 6114–6123. [Google Scholar] [CrossRef] [PubMed]
- Alam, R.; Forsythe, P.A.; Stafford, S.; Lett-Brown, M.A.; Grant, J.A. Macrophage inflammatory protein-1 alpha activates basophils and mast cells. J. Exp. Med. 1992, 176, 781–786. [Google Scholar] [CrossRef]
- Erdinest, N.; London, N.; Solomon, A. Chemokines in allergic conjunctivitis. Curr. Opin. Allergy Clin. Immunol. 2020, 20, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.Z.; Chiu, B.C.; Stolberg, V.; Lukacs, N.W.; Kunkel, S.L.; Murphy, H.S.; Chensue, S.W. Eosinophil recruitment in type-2 hypersensitivity pulmonary granulomas: Source and contribution of monocyte chemotactic protein-3 (CCL7). Am. J. Pathol. 2002, 161, 257–266. [Google Scholar] [CrossRef]
- Kuo, C.H.; Collins, A.M.; Boettner, D.R.; Yang, Y.; Ono, S.J. Role of CCL7 in Type I Hypersensitivity Reactions in Murine Experimental Allergic Conjunctivitis. J. Immunol. 2017, 198, 645–656. [Google Scholar] [CrossRef] [Green Version]
- Leonardi, A.; De Dominicis, C.; Motterle, L. Immunopathogenesis of ocular allergy: A schematic approach to different clinical entities. Curr. Opin. Allergy Clin. Immunol. 2007, 7, 429–435. [Google Scholar] [CrossRef]
- Chigbu, D.I.; Minhas, B.K. Immunopathology of allergic conjunctivitis. Eur. Med. J. 2018, 3, 76–83. [Google Scholar]
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast Cell: A Multi-Functional Master Cell. Front. Immunol. 2015, 6, 620. [Google Scholar] [CrossRef] [Green Version]
- Hitomi, T.; Zhang, J.; Nicoletti, L.M.; Grodzki, A.C.; Jamur, M.C.; Oliver, C.; Siraganian, R.P. Phospholipase D1 regulates high-affinity IgE receptor-induced mast cell degranulation. Blood 2004, 104, 4122–4128. [Google Scholar] [CrossRef]
- Smuda, C.; Bryce, P.J. New developments in the use of histamine and histamine receptors. Curr. Allergy Asthma Rep. 2011, 11, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Amaral, M.M.; Davio, C.; Ceballos, A.; Salamone, G.; Canones, C.; Geffner, J.; Vermeulen, M. Histamine improves antigen uptake and cross-presentation by dendritic cells. J. Immunol. 2007, 179, 3425–3433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Hernandez, M.C.; Montero, J.; Rondon, C.; Benitez del Castillo, J.M.; Velazquez, E.; Herreras, J.M.; Fernandez-Parra, B.; Merayo-Lloves, J.; Del Cuvillo, A.; Vega, F.; et al. Consensus document on allergic conjunctivitis (DECA). J. Investig. Allergol. Clin. Immunol. 2015, 25, 94–106. [Google Scholar] [PubMed]
- Portnoy, J.; Kennedy, K.; Sublett, J.; Phipatanakul, W.; Matsui, E.; Barnes, C.; Grimes, C.; Miller, J.D.; Seltzer, J.M.; Williams, P.B.; et al. Environmental assessment and exposure control: A practice parameter--furry animals. Ann. Allergy Asthma Immunol. 2012, 108, 223.e1–223.e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupuis, P.; Prokopich, C.L.; Hynes, A.; Kim, H. A contemporary look at allergic conjunctivitis. Allergy Asthma Clin. Immunol. Off. J. Can. Soc. Allergy Clin. Immunol. 2020, 16, 5. [Google Scholar] [CrossRef]
- Owen, C.; Shah, A.; Henshaw, K.; Smeeth, L.; Sheikh, A. Topical treatments for seasonal allergic conjunctivitis: Systematic review and meta-analysis of efficacy and effectiveness. Br. J. Gen. Pract. 2004, 54, 451–456. [Google Scholar]
- Azari, A.A.; Barney, N.P. Conjunctivitis: A systematic review of diagnosis and treatment. JAMA 2013, 310, 1721–1729. [Google Scholar] [CrossRef]
- Fauquert, J.L.; Jedrzejczak-Czechowicz, M.; Rondon, C.; Calder, V.; Silva, D.; Kvenshagen, B.K.; Callebaut, I.; Allegri, P.; Santos, N.; Doan, S.; et al. Conjunctival allergen provocation test: Guidelines for daily practice. Allergy 2017, 72, 43–54. [Google Scholar] [CrossRef]
- Leonardi, A.; Bogacka, E.; Fauquert, J.L.; Kowalski, M.L.; Groblewska, A.; Jedrzejczak-Czechowicz, M.; Doan, S.; Marmouz, F.; Demoly, P.; Delgado, L. Ocular allergy: Recognizing and diagnosing hypersensitivity disorders of the ocular surface. Allergy 2012, 67, 1327–1337. [Google Scholar] [CrossRef]
- O’Brien, T.P. Allergic conjunctivitis: An update on diagnosis and management. Curr. Opin. Allergy Clin. Immunol. 2013, 13, 543–549. [Google Scholar] [CrossRef]
- Abelson, M.; Paradis, A.; George, M.; Smith, L.; Maguire, L.; Burns, R. Effects of Vasocon-A in the allergen challenge model of acute allergic conjunctivitis. Arch. Ophthalmol. 1990, 108, 520–524. [Google Scholar] [CrossRef]
- Leonardi, S.; Marchese, G.; Marseglia, G.; La Rosa, M. Montelukast in allergic diseases beyond asthma. Allergy Asthma Proc. 2007, 28, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Chigbu, D.I. The management of allergic eye diseases in primary eye care. Cont. Lens. Anterior Eye 2009, 32, 260–272. [Google Scholar] [CrossRef] [PubMed]
- Welch, D.; Ousler, G.W., 3rd; Nally, L.A.; Abelson, M.B.; Wilcox, K.A. Ocular drying associated with oral antihistamines (loratadine) in the normal population-an evaluation of exaggerated dose effect. Adv. Exp. Med. Biol. 2002, 506, 1051–1055. [Google Scholar] [CrossRef] [PubMed]
- Bielory, L.; Schoenberg, D. Emerging Therapeutics for Ocular Surface Disease. Curr. Allergy Asthma Rep. 2019, 19, 16. [Google Scholar] [CrossRef] [PubMed]
- Gomes, P.J.; Ciolino, J.B.; Arranz, P.; Hernandez, G.; Fernandez, N. Efficacy of once-daily ophthalmic bilastine for the treatment of allergic conjunctivitis: A dose-finding study. J. Investig. Allergol. Clin. Immunol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Noble, S.; McTavish, D. Levocabastine. An update of its pharmacology, clinical efficacy and tolerability in the topical treatment of allergic rhinitis and conjunctivitis. Drugs 1995, 50, 1032–1049. [Google Scholar] [CrossRef]
- Bielory, L. Ocular allergy overview. Immunol. Allergy Clin. N. Am. 2008, 28, 1–23. [Google Scholar] [CrossRef]
- D’Arienzo, P.; Leonardi, A.; Bensch, G. Randomized, double-masked, placebo-controlled comparison of the efficacy of emedastine difumarate 0.05% ophthalmic solution and ketotifen fumarate 0.025% ophthalmic solution in the human conjunctival allergen challenge model. Clin. Ther. 2002, 24, 409–416. [Google Scholar] [CrossRef]
- Finn, D.F.; Walsh, J.J. Twenty-first century mast cell stabilizers. Br. J. Pharmacol. 2013, 170, 23–37. [Google Scholar] [CrossRef] [Green Version]
- Bielory, L. Ocular allergy treatments. Clin. Rev. Allergy Immunol. 2001, 20, 201–213. [Google Scholar] [CrossRef]
- Bielory, L. Ocular allergy guidelines: A practical treatment algorithm. Drugs 2002, 62, 1611–1634. [Google Scholar] [CrossRef] [PubMed]
- Verin, P.; Dicker, I.; Mortemousque, B. Nedocromil sodium eye drops are more effective than sodium cromoglycate eye drops for the long-term management of vernal keratoconjunctivitis. Clin. Exp. Allergy 1999, 29, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Alexander, M.; Allegro, S.; Hicks, A. Efficacy and acceptability of nedocromil sodium 2% and olopatadine hydrochloride 0.1% in perennial allergic conjunctivitis. Adv. Ther. 2000, 17, 140–147. [Google Scholar] [CrossRef]
- Mishra, G.P.; Tamboli, V.; Jwala, J.; Mitra, A.K. Recent patents and emerging therapeutics in the treatment of allergic conjunctivitis. Recent Pat. Inflamm. Allergy Drug Discov. 2011, 5, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Woerly, G.; Loiseau, S.; Loyens, M.; Schoch, C.; Capron, M. Inhibitory effects of ketotifen on eotaxin-dependent activation of eosinophils: Consequences for allergic eye diseases. Allergy 2003, 58, 397–406. [Google Scholar] [CrossRef]
- Nabe, M.; Miyagawa, H.; Agrawal, D.K.; Sugiyama, H.; Townley, R.G. The effect of ketotifen on eosinophils as measured at LTC4 release and by chemotaxis. Allergy Proc. 1991, 12, 267–271. [Google Scholar] [CrossRef]
- Canonica, G.; Ciprandi, G.; Petzold, U.; Kolb, C.; Ellers-Lenz, B.; Hermann, R. Topical azelastine in perennial allergic conjunctivitis. Curr. Med. Res. Opin. 2003, 19, 321–329. [Google Scholar] [CrossRef]
- Bielory, L.; Lien, K.; Bigelsen, S. Efficacy and tolerability of newer antihistamines in the treatment of allergic conjunctivitis. Drugs 2005, 65, 215–228. [Google Scholar] [CrossRef]
- Leonardi, A.; Abelson, M. Double-masked, randomized, placebo-controlled clinical study of the mast cell-stabilizing effects of treatment with olopatadine in the conjunctival allergen challenge model in humans. Clin. Ther. 2003, 25, 2539–2552. [Google Scholar] [CrossRef]
- Berger, W.; Abelson, M.; Gomes, P.; Beck, M.; Kimura, S.; Westbrook, T.; Storms, W.; Galant, S. Effects of adjuvant therapy with 0.1% olopatadine hydrochloride ophthalmic solution on quality of life in patients with allergic rhinitis using systemic or nasal therapy. Ann. Allergy Asthma Immunol. 2005, 95, 361–371. [Google Scholar] [CrossRef]
- Abelson, M.; Turner, D. A randomized, double-blind, parallel-group comparison of olopatadine 0.1% ophthalmic solution versus placebo for controlling the signs and symptoms of seasonal allergic conjunctivitis and rhinoconjunctivitis. Clin. Ther. 2003, 25, 931–947. [Google Scholar] [CrossRef]
- Abelson, M.; Pratt, S.; Mussoline, J.; Townsend, D. One-visit, randomized, placebo-controlled, conjunctival allergen challenge study of scanning and imaging technology for objective quantification of eyelid swelling in the allergic reaction with contralateral use of olopatadine and artificial tears. Clin. Ther. 2003, 25, 2070–2084. [Google Scholar] [CrossRef]
- Kidd, M.; McKenzie, S.; Steven, I.; Cooper, C.; Lanz, R. Efficacy and safety of ketotifen eye drops in the treatment of seasonal allergic conjunctivitis. Br. J. Ophthalmol. 2003, 87, 1206–1211. [Google Scholar] [CrossRef]
- Avunduk, A.; Tekelioglu, Y.; Turk, A.; Akyol, N. Comparison of the effects of ketotifen fumarate 0.025% and olopatadine HCl 0.1% ophthalmic solutions in seasonal allergic conjunctivities: A 30-day, randomized, double-masked, artificial tear substitute-controlled trial. Clin. Ther. 2005, 27, 1392–1402. [Google Scholar] [CrossRef]
- Castillo, M.; Scott, N.W.; Mustafa, M.Z.; Mustafa, M.S.; Azuara-Blanco, A. Topical antihistamines and mast cell stabilisers for treating seasonal and perennial allergic conjunctivitis. Cochrane Database Syst. Rev. 2015, CD009566. [Google Scholar] [CrossRef] [Green Version]
- Kam, K.W.; Chen, L.J.; Wat, N.; Young, A.L. Topical Olopatadine in the Treatment of Allergic Conjunctivitis: A Systematic Review and Meta-analysis. Ocul. Immunol. Inflamm. 2017, 25, 663–677. [Google Scholar] [CrossRef]
- Abelson, M.B.; Torkildsen, G.L.; Williams, J.I.; Gow, J.A.; Gomes, P.J.; McNamara, T.R.; Bepotastine Besilate Ophthalmic Solutions Clinical Study Group. Time to onset and duration of action of the antihistamine bepotastine besilate ophthalmic solutions 1.0% and 1.5% in allergic conjunctivitis: A phase III, single-center, prospective, randomized, double-masked, placebo-controlled, conjunctival allergen challenge assessment in adults and children. Clin. Ther. 2009, 31, 1908–1921. [Google Scholar] [CrossRef]
- Macejko, T.T.; Bergmann, M.T.; Williams, J.I.; Gow, J.A.; Gomes, P.J.; McNamara, T.R.; Abelson, M.B.; Bepotastine Besilate Ophthalmic Solutions Clinical Study Group. Multicenter clinical evaluation of bepotastine besilate ophthalmic solutions 1.0% and 1.5% to treat allergic conjunctivitis. Am. J. Ophthalmol. 2010, 150, 122–127.e125. [Google Scholar] [CrossRef]
- McCabe, C.F.; McCabe, S.E. Comparative efficacy of bepotastine besilate 1.5% ophthalmic solution versus olopatadine hydrochloride 0.2% ophthalmic solution evaluated by patient preference. Clin. Ophthalmol. 2012, 6, 1731–1738. [Google Scholar] [CrossRef] [Green Version]
- Kari, O.; Saari, K.M. Updates in the treatment of ocular allergies. J. Asthma Allergy 2010, 3, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Gray, S.L.; Anderson, M.L.; Dublin, S.; Hanlon, J.T.; Hubbard, R.; Walker, R.; Yu, O.; Crane, P.K.; Larson, E.B. Cumulative use of strong anticholinergics and incident dementia: A prospective cohort study. JAMA Intern. Med. 2015, 175, 401–407. [Google Scholar] [CrossRef]
- Ehrchen, J.M.; Roth, J.; Barczyk-Kahlert, K. More than Suppression: Glucocorticoid Action on Monocytes and Macrophages. Front. Immunol. 2019, 10, 2028. [Google Scholar] [CrossRef] [Green Version]
- Comstock, T.L.; Decory, H.H. Advances in corticosteroid therapy for ocular inflammation: Loteprednol etabonate. Int. J. Inflamm. 2012, 2012, 789623. [Google Scholar] [CrossRef]
- Ohta, K.; Yamashita, N. Apoptosis of eosinophils and lymphocytes in allergic inflammation. J. Allergy Clin. Immunol. 1999, 104, 14–21. [Google Scholar] [CrossRef]
- Sautebin, L.; Carnuccio, R.; Ialenti, A.; Di Rosa, M. Lipocortin and vasocortin: Two species of anti-inflammatory proteins mimicking the effects of glucocorticoids. Pharmacol. Res. 1992, 25, 1–12. [Google Scholar] [CrossRef]
- Kitamura, Y.; Das, A.K.; Murata, Y.; Maeyama, K.; Dev, S.; Wakayama, Y.; Kalubi, B.; Takeda, N.; Fukui, H. Dexamethasone suppresses histamine synthesis by repressing both transcription and activity of HDC in allergic rats. Allergol. Int. 2006, 55, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Ilyas, H.; Slonim, C.; Braswell, G.; Favetta, J.; Schulman, M. Long-term safety of loteprednol etabonate 0.2% in the treatment of seasonal and perennial allergic conjunctivitis. Eye Contact Lens. 2004, 30, 10–13. [Google Scholar] [CrossRef]
- Shulman, D.; Lothringer, L.; Rubin, J.; Briggs, R.; Howes, J.; Novack, G.; Hart, K. A randomized, double-masked, placebo-controlled parallel study of loteprednol etabonate 0.2% in patients with seasonal allergic conjunctivitis. Ophthalmology 1999, 106, 362–369. [Google Scholar] [CrossRef]
- Dell, S.; Lowry, G.; Northcutt, J.; Howes, J.; Novack, G.; Hart, K. A randomized, double-masked, placebo-controlled parallel study of 0.2% loteprednol etabonate in patients with seasonal allergic conjunctivitis. J. Allergy Clin. Immunol. 1998, 102, 251–255. [Google Scholar] [CrossRef]
- Yaylali, V.; Demirlenk, I.; Tatlipinar, S.; Ozbay, D.; Esme, A.; Yildirim, C.; Ozden, S. Comparative study of 0.1% olopatadine hydrochloride and 0.5% ketorolac tromethamine in the treatment of seasonal allergic conjunctivitis. Acta Ophthalmol. Scand. 2003, 81, 378–382. [Google Scholar] [CrossRef]
- Anolik, R.; Pearlman, D.; Teper, A.; Gates, D. Mometasone furoate improves nasal and ocular symptoms of seasonal allergic rhinitis in adolescents. Allergy Asthma Proc. 2009, 30, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Andrews, C.P.; Martin, B.G.; Jacobs, R.L.; Mohar, D.E.; Diaz, J.D.; Amar, N.J.; Kaiser, H.B.; Vandewalker, M.L.; Bernstein, J.; Toler, W.T.; et al. Fluticasone furoate nasal spray is more effective than fexofenadine for nighttime symptoms of seasonal allergy. Allergy Asthma Proc. 2009, 30, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Bielory, L. Allergic conjunctivitis and the impact of allergic rhinitis. Curr. Allergy Asthma Rep. 2010, 10, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Singhal, D.; Sahay, P.; Maharana, P.K.; Raj, N.; Sharma, N.; Titiyal, J.S. Vernal Keratoconjunctivitis. Surv. Ophthalmol. 2019, 64, 289–311. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.M.; Yau, G.S.; Lee, J.W.; Wong, A.L.; Tam, V.T.; Yuen, C.Y. Retrospective review on the use of topical cyclosporin a 0.05% for paediatric allergic conjunctivitis in Hong Kong Chinese. Sci. World J. 2014, 2014, 396987. [Google Scholar] [CrossRef] [Green Version]
- Hazarika, A.K.; Singh, P.K. Efficacy of topical application of 0.03% tacrolimus eye ointment in the management of allergic conjunctivitis. J. Nat. Sci. Biol. Med. 2015, 6, S10–S12. [Google Scholar] [CrossRef] [Green Version]
- Broide, D.H. Immunomodulation of allergic disease. Annu. Rev. Med. 2009, 60, 279–291. [Google Scholar] [CrossRef] [Green Version]
- Walker, S.M.; Varney, V.A.; Gaga, M.; Jacobson, M.R.; Durham, S.R. Grass pollen immunotherapy: Efficacy and safety during a 4-year follow-up study. Allergy 1995, 50, 405–413. [Google Scholar] [CrossRef]
- Moller, C.; Juto, P.; Dreborg, S.; Bjorksten, B. Blood lymphocyte proliferation response to pollen extract as a monitor of immunotherapy. Allergy 1984, 39, 291–296. [Google Scholar] [CrossRef]
- Baiula, M.; Bedini, A.; Baldi, J.; Cavet, M.E.; Govoni, P.; Spampinato, S. Mapracorat, a selective glucocorticoid receptor agonist, causes apoptosis of eosinophils infiltrating the conjunctiva in late-phase experimental ocular allergy. Drug Des. Dev. Ther. 2014, 8, 745–757. [Google Scholar] [CrossRef]
- Baiula, M.; Spampinato, S. Mapracorat, a novel non-steroidal selective glucocorticoid receptor agonist for the treatment of allergic conjunctivitis. Inflamm. Allergy Drug Targets 2014, 13, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Baiula, M.; Sparta, A.; Bedini, A.; Carbonari, G.; Bucolo, C.; Ward, K.W.; Zhang, J.Z.; Govoni, P.; Spampinato, S. Eosinophil as a cellular target of the ocular anti-allergic action of mapracorat, a novel selective glucocorticoid receptor agonist. Mol. Vis. 2011, 17, 3208–3223. [Google Scholar] [PubMed]
- Cavet, M.E.; Volhejn, S.; Harrington, K.L.; Zhang, J.Z. Anti-allergic effects of mapracorat, a novel selective glucocorticoid receptor agonist, in human conjunctival fibroblasts and epithelial cells. Mol. Vis. 2013, 19, 1515–1525. [Google Scholar]
- Cavet, M.E.; Harrington, K.L.; Ward, K.W.; Zhang, J.Z. Mapracorat, a novel selective glucocorticoid receptor agonist, inhibits hyperosmolar-induced cytokine release and MAPK pathways in human corneal epithelial cells. Mol. Vis. 2010, 16, 1791–1800. [Google Scholar] [PubMed]
- Schacke, H.; Zollner, T.M.; Docke, W.D.; Rehwinkel, H.; Jaroch, S.; Skuballa, W.; Neuhaus, R.; May, E.; Zugel, U.; Asadullah, K. Characterization of ZK 245186, a novel, selective glucocorticoid receptor agonist for the topical treatment of inflammatory skin diseases. Br. J. Pharmacol. 2009, 158, 1088–1103. [Google Scholar] [CrossRef] [PubMed]
- Shafiee, A.; Bucolo, C.; Budzynski, E.; Ward, K.W.; Lopez, F.J. In vivo ocular efficacy profile of mapracorat, a novel selective glucocorticoid receptor agonist, in rabbit models of ocular disease. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1422–1430. [Google Scholar] [CrossRef] [Green Version]
- Pfeffer, B.A.; DeWitt, C.A.; Salvador-Silva, M.; Cavet, M.E.; Lopez, F.J.; Ward, K.W. Reduced myocilin expression in cultured monkey trabecular meshwork cells induced by a selective glucocorticoid receptor agonist: Comparison with steroids. Investig. Ophthalmol. Vis. Sci. 2010, 51, 437–446. [Google Scholar] [CrossRef]
- De Bosscher, K.; Haegeman, G.; Elewaut, D. Targeting inflammation using selective glucocorticoid receptor modulators. Curr. Opin. Pharmacol. 2010, 10, 497–504. [Google Scholar] [CrossRef]
- Komatsu, N.; Miyazaki, D.; Tominaga, T.; Kuo, C.; Namba, S.; Takeda, S.; Higashi, H.; Inoue, Y. Transcriptional analyses before and after suppression of immediate hypersensitivity reactions by CCR3 blockade in eyes with experimental allergic conjunctivitis. Investig. Ophthalmol. Vis. Sci. 2008, 49, 5307–5313. [Google Scholar] [CrossRef]
- Fulkerson, P.C.; Fischetti, C.A.; McBride, M.L.; Hassman, L.M.; Hogan, S.P.; Rothenberg, M.E. A central regulatory role for eosinophils and the eotaxin/CCR3 axis in chronic experimental allergic airway inflammation. Proc. Natl. Acad. Sci. USA 2006, 103, 16418–16423. [Google Scholar] [CrossRef] [Green Version]
- Ono, S. Molecular genetics of allergic diseases. Annu. Rev. Immunol. 2000, 18, 347–366. [Google Scholar] [CrossRef] [PubMed]
- Cook, E. Tear cytokines in acute and chronic ocular allergic inflammation. Curr. Opin. Allergy Clin. Immunol. 2004, 4, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Mimura, T.; Usui, T.; Mori, M.; Aixinjueluo, W.; Funatsu, H.; Noma, H.; Amano, S. Immunochromatographic assay for measurement of total IgE in tears, nasal mucus, and saliva of patients with allergic rhinoconjunctivitis. J. Asthma 2010, 47, 1153–1160. [Google Scholar] [CrossRef] [PubMed]
- Carreno, E.; Enriquez-de-Salamanca, A.; Teson, M.; Garcia-Vazquez, C.; Stern, M.E.; Whitcup, S.M.; Calonge, M. Cytokine and chemokine levels in tears from healthy subjects. Acta Ophthalmol. 2010, 88, e250–e258. [Google Scholar] [CrossRef] [PubMed]
- LaFrance, M.W.; Kehinde, L.E.; Fullard, R.J. Multiple cytokine analysis in human tears: An optimized procedure for cytometric bead-based assay. Curr. Eye Res. 2008, 33, 525–544. [Google Scholar] [CrossRef] [PubMed]
- Abu El-Asrar, A.; Struyf, S.; Al-Kharashi, S.; Missotten, L.; Van Damme, J.; Geboes, K. Chemokines in the limbal form of vernal keratoconjunctivitis. Br. J. Ophthalmol. 2000, 84, 1360–1366. [Google Scholar] [CrossRef] [Green Version]
- Eperon, S.; Berguiga, M.; Ballabeni, P.; Guex-Crosier, C.; Guex-Crosier, Y. Total IgE and eotaxin (CCL11) contents in tears of patients suffering from seasonal allergic conjunctivitis. Graefes Arch. Clin. Exp. Ophthalmol. 2014, 252, 1359–1367. [Google Scholar] [CrossRef] [Green Version]
- Haig, D.M.; Huntley, J.F.; MacKellar, A.; Newlands, G.F.; Inglis, L.; Sangha, R.; Cohen, D.; Hapel, A.; Galli, S.J.; Miller, H.R. Effects of stem cell factor (kit-ligand) and interleukin-3 on the growth and serine proteinase expression of rat bone-marrow-derived or serosal mast cells. Blood 1994, 83, 72–83. [Google Scholar] [CrossRef] [Green Version]
- Tominaga, T.; Miyazaki, D.; Sasaki, S.; Mihara, S.; Komatsu, N.; Yakura, K.; Inoue, Y. Blocking mast cell-mediated type I hypersensitivity in experimental allergic conjunctivitis by monocyte chemoattractant protein-1/CCR2. Investig. Ophthalmol. Vis. Sci. 2009, 50, 5181–5188. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, M.H.; Martel, J.R.; Sall, K.; Goldberg, D.F.; Abrams, M.; Rubin, J.; Sheppard, J.; Tauber, J.; Korenfeld, M.; Agahigian, J.; et al. Multicenter Study of a Novel Topical Interleukin-1 Receptor Inhibitor, Isunakinra, in Subjects With Moderate to Severe Dry Eye Disease. Eye Contact Lens. 2017, 43, 287–296. [Google Scholar] [CrossRef]
- Amparo, F.; Dastjerdi, M.H.; Okanobo, A.; Ferrari, G.; Smaga, L.; Hamrah, P.; Jurkunas, U.; Schaumberg, D.A.; Dana, R. Topical interleukin 1 receptor antagonist for treatment of dry eye disease: A randomized clinical trial. JAMA Ophthalmol. 2013, 131, 715–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keane-Myers, A.; Miyazaki, D.; Liu, G.; Dekaris, I.; Ono, S.; Dana, M. Prevention of allergic eye disease by treatment with IL-1 receptor antagonist. Investig. Ophthalmol. Vis. Sci. 1999, 40, 3041–3046. [Google Scholar]
- Tauber, J.; Karpecki, P.; Latkany, R.; Luchs, J.; Martel, J.; Sall, K.; Raychaudhuri, A.; Smith, V.; Semba, C.P.; Investigators, O. Lifitegrast Ophthalmic Solution 5.0% versus Placebo for Treatment of Dry Eye Disease: Results of the Randomized Phase III OPUS-2 Study. Ophthalmology 2015, 122, 2423–2431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cevikbas, F.; Wang, X.; Akiyama, T.; Kempkes, C.; Savinko, T.; Antal, A.; Kukova, G.; Buhl, T.; Ikoma, A.; Buddenkotte, J.; et al. A sensory neuron-expressed IL-31 receptor mediates T helper cell-dependent itch: Involvement of TRPV1 and TRPA1. J. Allergy Clin. Immunol. 2014, 133, 448–460. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Escalera, J.; Balakrishna, S.; Fan, L.; Caceres, A.I.; Robinson, E.; Sui, A.; McKay, M.C.; McAlexander, M.A.; Herrick, C.A.; et al. TRPA1 controls inflammation and pruritogen responses in allergic contact dermatitis. FASEB J. 2013, 27, 3549–3563. [Google Scholar] [CrossRef] [Green Version]
- Shim, W.S.; Tak, M.H.; Lee, M.H.; Kim, M.; Kim, M.; Koo, J.Y.; Lee, C.H.; Kim, M.; Oh, U. TRPV1 mediates histamine-induced itching via the activation of phospholipase A2 and 12-lipoxygenase. J. Neurosci. 2007, 27, 2331–2337. [Google Scholar] [CrossRef]
- Kwon, J.Y.; Lee, H.S.; Joo, C.K. TRPV1 Antagonist Suppresses Allergic Conjunctivitis in a Murine Model. Ocul. Immunol. Inflamm. 2018, 26, 440–448. [Google Scholar] [CrossRef]
- Ghoreschi, K.; Jesson, M.I.; Li, X.; Lee, J.L.; Ghosh, S.; Alsup, J.W.; Warner, J.D.; Tanaka, M.; Steward-Tharp, S.M.; Gadina, M.; et al. Modulation of innate and adaptive immune responses by tofacitinib (CP-690,550). J. Immunol. 2011, 186, 4234–4243. [Google Scholar] [CrossRef] [Green Version]
- Fragoulis, G.E.; McInnes, I.B.; Siebert, S. JAK-inhibitors. New players in the field of immune-mediated diseases, beyond rheumatoid arthritis. Rheumatology 2019, 58, i43–i54. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, D.M.; Kanno, Y.; Villarino, A.; Ward, M.; Gadina, M.; O’Shea, J.J. JAK inhibition as a therapeutic strategy for immune and inflammatory diseases. Nat. Rev. Drug Discov. 2017, 17, 78. [Google Scholar] [CrossRef] [Green Version]
- Berekmeri, A.; Mahmood, F.; Wittmann, M.; Helliwell, P. Tofacitinib for the treatment of psoriasis and psoriatic arthritis. Expert Rev. Clin. Immunol. 2018, 14, 719–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandborn, W.J.; Ghosh, S.; Panes, J.; Vranic, I.; Su, C.; Rousell, S.; Niezychowski, W.; Study, A.I. Tofacitinib, an oral Janus kinase inhibitor, in active ulcerative colitis. N. Engl. J. Med. 2012, 367, 616–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Liu, X.; Yu, J.; Li, Z.; Chen, Y.; Li, H.; Chen, X.; Su, W.; Liang, D. Tofacitinib suppresses mast cell degranulation and attenuates experimental allergic conjunctivitis. Int. Immunopharmacol. 2020, 86, 106737. [Google Scholar] [CrossRef] [PubMed]
- Mandell, K.J.; Clark, D.; Chu, D.S.; Foster, C.S.; Sheppard, J.; Brady, T.C. Randomized Phase 2 Trial of Reproxalap, a Novel Reactive Aldehyde Species Inhibitor, in Patients with Noninfectious Anterior Uveitis: Model for Corticosteroid Replacement. J. Ocul. Pharmacol. Ther. 2020, 36, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.; Sheppard, J.; Brady, T.C. A Randomized Double-Masked Phase 2a Trial to Evaluate Activity and Safety of Topical Ocular Reproxalap, a Novel RASP Inhibitor, in Dry Eye Disease. J. Ocul. Pharmacol. Ther. 2021, 37, 193–199. [Google Scholar] [CrossRef]
- Higdon, A.; Diers, A.R.; Oh, J.Y.; Landar, A.; Darley-Usmar, V.M. Cell signalling by reactive lipid species: New concepts and molecular mechanisms. Biochem. J. 2012, 442, 453–464. [Google Scholar] [CrossRef] [Green Version]
- Kalariya, N.M.; Ramana, K.V.; Srivastava, S.K.; van Kuijk, F.J. Carotenoid derived aldehydes-induced oxidative stress causes apoptotic cell death in human retinal pigment epithelial cells. Exp. Eye Res. 2008, 86, 70–80. [Google Scholar] [CrossRef] [Green Version]
- Kauppinen, A.; Niskanen, H.; Suuronen, T.; Kinnunen, K.; Salminen, A.; Kaarniranta, K. Oxidative stress activates NLRP3 inflammasomes in ARPE-19 cells—Implications for age-related macular degeneration (AMD). Immunol. Lett. 2012, 147, 29–33. [Google Scholar] [CrossRef]
- Sapkota, M.; DeVasure, J.M.; Kharbanda, K.K.; Wyatt, T.A. Malondialdehyde-acetaldehyde (MAA) adducted surfactant protein induced lung inflammation is mediated through scavenger receptor a (SR-A1). Respir. Res. 2017, 18, 36. [Google Scholar] [CrossRef] [Green Version]
- Clark, D.; Karpecki, P.; Salapatek, A.M.; Sheppard, J.D.; Brady, T.C. Reproxalap Improves Signs and Symptoms of Allergic Conjunctivitis in an Allergen Chamber: A Real-World Model of Allergen Exposure. Clin. Ophthalmol. 2022, 16, 15–23. [Google Scholar] [CrossRef]
- Clark, D.; Cavanagh, B.; Shields, A.L.; Karpecki, P.; Sheppard, J.; Brady, T.C. Clinically Relevant Activity of the Novel RASP Inhibitor Reproxalap in Allergic Conjunctivitis: The Phase 3 ALLEVIATE Trial. Am. J. Ophthalmol. 2021, 230, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K. FAK-Dependent Cell Motility and Cell Elongation. Cells 2020, 9, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.; Khoe, M.; Befekadu, M.; Chung, S.; Takata, Y.; Ilic, D.; Bryer-Ash, M. Focal adhesion kinase mediates cell survival via NF-kappaB and ERK signaling pathways. Am. J. Physiology. Cell Physiol. 2007, 292, C1339–C1352. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, Y.; Matsumoto, Y.; Funakoshi, M.; Yamamoto, D.; Hanks, S.K.; Kasahara, T. Anti-apoptotic role of focal adhesion kinase (FAK). Induction of inhibitor-of-apoptosis proteins and apoptosis suppression by the overexpression of FAK in a human leukemic cell line, HL-60. J. Biol. Chem. 2000, 275, 16309–16315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, J.L. Role of focal adhesion kinase in integrin signaling. Int. J. Biochem. Cell Biol. 1997, 29, 1085–1096. [Google Scholar] [CrossRef]
- Salazar, E.P.; Rozengurt, E. Src family kinases are required for integrin-mediated but not for G protein-coupled receptor stimulation of focal adhesion kinase autophosphorylation at Tyr-397. J. Biol. Chem. 2001, 276, 17788–17795. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.J.; Zhang, L.N.; Wang, H.N.; Xie, C.C.; Li, W.Y.; Gao, P.; Hu, W.Z.; Zhao, Z.F.; Ji, K. FAK inhibitor PF-431396 suppresses IgE-mediated mast cell activation and allergic inflammation in mice. Biochem. Pharmacol. 2021, 192, 114722. [Google Scholar] [CrossRef]
- Okubo, K.; Ogino, S.; Nagakura, T.; Ishikawa, T. Omalizumab is effective and safe in the treatment of Japanese cedar pollen-induced seasonal allergic rhinitis. Allergol. Int. 2006, 55, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Kirikkaya, E.; Degirmenci, P. Effects of omalizumab on allergic conjunctivitis. Int. Ophthalmol. 2022, 42, 167–175. [Google Scholar] [CrossRef]
- Taille, C.; Doan, S.; Neukirch, C.; Aubier, M. Omalizumab for severe atopic keratoconjunctivitis. BMJ Case Rep. 2010, 2010, bcr0420102919. [Google Scholar] [CrossRef]
- Heffler, E.; Picardi, G.; Liuzzo, M.T.; Pistorio, M.P.; Crimi, N. Omalizumab Treatment of Vernal Keratoconjunctivitis. JAMA Ophthalmol. 2016, 134, 461–463. [Google Scholar] [CrossRef] [PubMed]
- de Klerk, T.A.; Sharma, V.; Arkwright, P.D.; Biswas, S. Severe vernal keratoconjunctivitis successfully treated with subcutaneous omalizumab. J. AAPOS 2013, 17, 305–306. [Google Scholar] [CrossRef] [PubMed]
- Wollenberg, A.; Ariens, L.; Thurau, S.; van Luijk, C.; Seegraber, M.; de Bruin-Weller, M. Conjunctivitis occurring in atopic dermatitis patients treated with dupilumab-clinical characteristics and treatment. J. Allergy Clin. Immunol. Pract. 2018, 6, 1778–1780.e1. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Venkateswaran, N.; Wang, Z.; Galor, A. Ocular involvement in atopic disease: A review. Curr. Opin. Ophthalmol. 2018, 29, 576–581. [Google Scholar] [CrossRef]
- Bozkurt, M.K.; Tulek, B.; Bozkurt, B.; Akyurek, N.; Oz, M.; Kiyici, A. Comparison of the efficacy of prednisolone, montelukast, and omalizumab in an experimental allergic rhinitis model. Turk. J. Med. Sci. 2014, 44, 439–447. [Google Scholar] [CrossRef]
- van der Schaft, J.; Thijs, J.L.; de Bruin-Weller, M.S.; Balak, D.M.W. Dupilumab after the 2017 approval for the treatment of atopic dermatitis: What’s new and what’s next? Curr. Opin. Allergy Clin. Immunol. 2019, 19, 341–349. [Google Scholar] [CrossRef]
- Nahum, Y.; Mimouni, M.; Livny, E.; Bahar, I.; Hodak, E.; Leshem, Y.A. Dupilumab-induced ocular surface disease (DIOSD) in patients with atopic dermatitis: Clinical presentation, risk factors for development and outcomes of treatment with tacrolimus ointment. Br. J. Ophthalmol. 2020, 104, 776–779. [Google Scholar] [CrossRef]
- Cholkar, K.; Gilger, B.C.; Mitra, A.K. Topical delivery of aqueous micellar resolvin E1 analog (RX-10045). Int. J. Pharm. 2016, 498, 326–334. [Google Scholar] [CrossRef] [Green Version]
- Baiula, M.; Spampinato, S. Experimental Pharmacotherapy for Dry Eye Disease: A Review. J. Exp. Pharmacol. 2021, 13, 345–358. [Google Scholar] [CrossRef]
- Wu, B.; Gao, F.; Lin, J.; Lu, L.; Xu, H.; Xu, G.T. Conditioned Medium of Human Amniotic Epithelial Cells Alleviates Experimental Allergic Conjunctivitis Mainly by IL-1ra and IL-10. Front. Immunol. 2021, 12, 774601. [Google Scholar] [CrossRef]
- Trounson, A.; McDonald, C. Stem Cell Therapies in Clinical Trials: Progress and Challenges. Cell Stem Cell 2015, 17, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Zhang, J.; Tsang, K.S.; Yang, H.; Gao, W.Q. Therapeutic Potential of Human Amniotic Epithelial Cells on Injuries and Disorders in the Central Nervous System. Stem Cells Int. 2019, 2019, 5432301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohshima, M.; Yamahara, K.; Ishikane, S.; Harada, K.; Tsuda, H.; Otani, K.; Taguchi, A.; Miyazato, M.; Katsuragi, S.; Yoshimatsu, J.; et al. Systemic transplantation of allogenic fetal membrane-derived mesenchymal stem cells suppresses Th1 and Th17 T cell responses in experimental autoimmune myocarditis. J. Mol. Cell Cardiol. 2012, 53, 420–428. [Google Scholar] [CrossRef]
- Chen, M.; Su, W.; Lin, X.; Guo, Z.; Wang, J.; Zhang, Q.; Brand, D.; Ryffel, B.; Huang, J.; Liu, Z.; et al. Adoptive transfer of human gingiva-derived mesenchymal stem cells ameliorates collagen-induced arthritis via suppression of Th1 and Th17 cells and enhancement of regulatory T cell differentiation. Arthritis Rheum. 2013, 65, 1181–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Tian, Y.; Xie, L.; Liu, X.; Huang, Z.; Su, W. Mesenchymal stem cells in allergic diseases: Current status. Allergol. Int. 2020, 69, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Royce, S.G.; Tominaga, A.M.; Shen, M.; Patel, K.P.; Huuskes, B.M.; Lim, R.; Ricardo, S.D.; Samuel, C.S. Serelaxin improves the therapeutic efficacy of RXFP1-expressing human amnion epithelial cells in experimental allergic airway disease. Clin. Sci. 2016, 130, 2151–2165. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Cauchois, G.; Louvet, N.; Chen, Y.; Rahouadj, R.; Huselstein, C. Comparison of MSC properties in two different hydrogels. Impact of mechanical properties. Biomed. Mater. Eng. 2017, 28, S193–S200. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Song, L.; Chen, F.; Ma, X. Effects of combination of mizolastine and proteoglycan on chronic urticaria: A randomized controlled trial. Arch. Dermatol. Res. 2019, 311, 801–805. [Google Scholar] [CrossRef]
- El-Emam, G.A.; Girgis, G.N.S.; Hamed, M.F.; El-Azeem Soliman, O.A.; Abd El Gawad, A. Formulation and Pathohistological Study of Mizolastine-Solid Lipid Nanoparticles-Loaded Ocular Hydrogels. Int. J. Nanomed. 2021, 16, 7775–7799. [Google Scholar] [CrossRef]
- Gous, P.; Ropo, A. A comparative trial of the safety and efficacy of 0.1 percent pemirolast potassium ophthalmic solution dosed twice or four times a day in patients with seasonal allergic conjunctivitis. J. Ocul. Pharmacol. Ther. 2004, 20, 139–150. [Google Scholar] [CrossRef]
- Shulman, D. Two mast cell stabilizers, pemirolast potassium 0.1% and nedocromil sodium 2%, in the treatment of seasonal allergic conjunctivitis: A comparative study. Adv. Ther. 2003, 20, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Abelson, M.; Berdy, G.; Mundorf, T.; Amdahl, L.; Graves, A. Pemirolast potassium 0.1% ophthalmic solution is an effective treatment for allergic conjunctivitis: A pooled analysis of two prospective, randomized, double-masked, placebo-controlled, phase III studies. J. Ocul. Pharmacol. Ther. 2002, 18, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Minami, K.; Hossen, M.; Kamei, C. Increasing effect by simultaneous use of levocabastine and pemirolast on experimental allergic conjunctivitis in rats. Biol. Pharm. Bull. 2005, 28, 473–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ron, E.S.; Bromberg, L.E. Temperature-responsive gels and thermogelling polymer matrices for protein and peptide delivery. Adv. Drug Deliv. Rev. 1998, 31, 197–221. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Yang, Z. In vivo and in vitro Evaluation of in situ Gel Formulation of Pemirolast Potassium in Allergic Conjunctivitis. Drug Des. Dev. Ther. 2021, 15, 2099–2107. [Google Scholar] [CrossRef]
- Pall, B.; Gomes, P.; Yi, F.; Torkildsen, G. Management of Ocular Allergy Itch With an Antihistamine-Releasing Contact Lens. Cornea 2019, 38, 713–717. [Google Scholar] [CrossRef]
- Xue, Y.; Zhang, W.; Lei, Y.; Dang, M. Novel Polyvinyl Pyrrolidone-Loaded Olopatadine HCl-Laden Doughnut Contact Lens to Treat Allergic Conjunctivitis. J. Pharm. Sci. 2020, 109, 1714–1724. [Google Scholar] [CrossRef]
- Minami, T.; Ishida, W.; Kishimoto, T.; Nakajima, I.; Hino, S.; Arai, R.; Matsunaga, T.; Fukushima, A.; Yamagami, S. In vitro and in vivo performance of epinastine hydrochloride-releasing contact lenses. PLoS ONE 2019, 14, e0210362. [Google Scholar] [CrossRef]
- McLaurin, E.B.; Evans, D.; Repke, C.S.; Sato, M.A.; Gomes, P.J.; Reilly, E.; Blender, N.; Silva, F.Q.; Vantipalli, S.; Metzinger, J.L.; et al. Phase 3 Randomized Study of Efficacy and Safety of a Dexamethasone Intracanalicular Insert in Patients With Allergic Conjunctivitis. Am. J. Ophthalmol. 2021, 229, 288–300. [Google Scholar] [CrossRef]
- Liu, Y.C.; Ng, X.W.; Teo, E.P.W.; Ang, H.P.; Lwin, N.C.; Chan, N.S.W.; Venkatraman, S.S.; Wong, T.T.; Mehta, J.S. A Biodegradable, Sustained-Released, Tacrolimus Microfilm Drug Delivery System for the Management of Allergic Conjunctivitis in a Mouse Model. Investig. Ophthalmol. Vis. Sci. 2018, 59, 675–684. [Google Scholar] [CrossRef]
- Leonardi, A.; Doan, S.; Amrane, M.; Ismail, D.; Montero, J.; Nemeth, J.; Aragona, P.; Bremond-Gignac, D.; Group, V.S. A Randomized, Controlled Trial of Cyclosporine A Cationic Emulsion in Pediatric Vernal Keratoconjunctivitis: The VEKTIS Study. Ophthalmology 2019, 126, 671–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, V.; Jain, G.K.; Nirmal, J.; Kohli, K. Topical tacrolimus nanoemulsion, a promising therapeutic approach for uveitis. Med. Hypotheses 2013, 81, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.C.; Lin, M.T.; Ng, A.H.C.; Wong, T.T.; Mehta, J.S. Nanotechnology for the Treatment of Allergic Conjunctival Diseases. Pharmaceuticals 2020, 13, 351. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Labib, B.A.; Chigbu, D.I. Therapeutic Targets in Allergic Conjunctivitis. Pharmaceuticals 2022, 15, 547. https://doi.org/10.3390/ph15050547
Labib BA, Chigbu DI. Therapeutic Targets in Allergic Conjunctivitis. Pharmaceuticals. 2022; 15(5):547. https://doi.org/10.3390/ph15050547
Chicago/Turabian StyleLabib, Bisant A., and DeGaulle I. Chigbu. 2022. "Therapeutic Targets in Allergic Conjunctivitis" Pharmaceuticals 15, no. 5: 547. https://doi.org/10.3390/ph15050547
APA StyleLabib, B. A., & Chigbu, D. I. (2022). Therapeutic Targets in Allergic Conjunctivitis. Pharmaceuticals, 15(5), 547. https://doi.org/10.3390/ph15050547