
Nitrative Stress and Auditory Dysfunction



Abstract
:1. Introduction
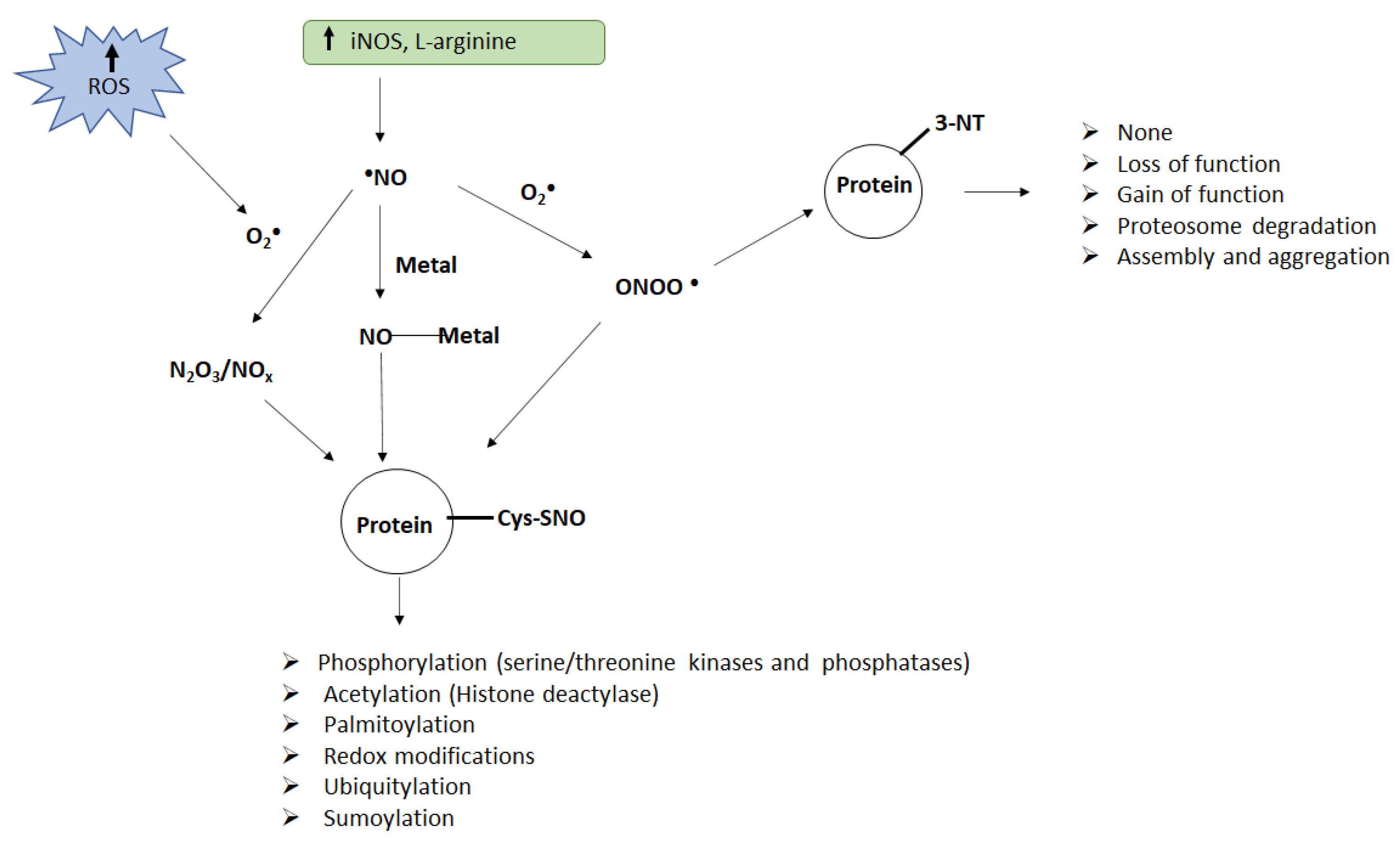
2. Nitrative Stress-Induced Post-Translational Modifications
2.1. Pathophysiological Consequences of Nitrative Stress
2.2. Nitrative Stress and Otopathology
2.3. Assessment of Nitrative Stress
2.4. Targeting Nitrative Stress for Otoprotection
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shahani, N.; Sawa, A. Protein S-nitrosylation: Role for nitric oxide signaling in neuronal death. Biochim. Biophys. Acta 2012, 1820, 736–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alderton, W.K.; Cooper, C.E.; Knowles, R.G. Nitric oxide synthases: Structure, function and inhibition. Biochem. J. 2001, 357, 593–615. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.T.; Pae, H.O.; Choi, B.M.; Billiar, T.R.; Kim, Y.M. Nitric oxide as a bioregulator of apoptosis. Biochem. Biophys. Res. Commun. 2001, 282, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.M.; Pae, H.O.; Jang, S.I.; Kim, Y.M.; Chung, H.T. Nitric oxide as a pro-apoptotic as well as anti-apoptotic modulator. J. Biochem. Mol. Biol. 2002, 35, 116–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.M.; Kim, P.K.; Kwon, Y.G.; Bai, S.K.; Nam, W.D.; Kim, Y.M. Regulation of apoptosis by nitrosative stress. J. Biochem. Mol. Biol. 2002, 35, 127–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razavi, H.M.; Hamilton, J.A.; Feng, Q. Modulation of apoptosis by nitric oxide: Implications in myocardial ischemia and heart failure. Pharmacol. Ther. 2005, 106, 147–162. [Google Scholar] [CrossRef]
- Martinez, M.C.; Andriantsitohaina, R. Reactive nitrogen species: Molecular mechanisms and potential significance in health and disease. Antioxid. Redox Signal. 2009, 11, 669–702. [Google Scholar] [CrossRef] [PubMed]
- Quijano, C.; Hernandez-Saavedra, D.; Castro, L.; McCord, J.M.; Freeman, B.A.; Radi, R. Reaction of peroxynitrite with Mn-superoxide dismutase. Role of the metal center in decomposition kinetics and nitration. J. Biol. Chem. 2001, 276, 11631–11638. [Google Scholar] [CrossRef] [Green Version]
- Radi, R. Nitric oxide, oxidants, and protein tyrosine nitration. Proc. Natl. Acad. Sci. USA 2004, 101, 4003–4008. [Google Scholar] [CrossRef] [Green Version]
- Donnini, S.; Monti, M.; Roncone, R.; Morbidelli, L.; Rocchigiani, M.; Oliviero, S.; Casella, L.; Giachetti, A.; Schulz, R.; Ziche, M. Peroxynitrite inactivates human-tissue inhibitor of metalloproteinase-4. FEBS Lett. 2008, 582, 1135–1140. [Google Scholar] [CrossRef] [Green Version]
- Yakovlev, V.A.; Bayden, A.S.; Graves, P.R.; Kellogg, G.E.; Mikkelsen, R.B. Nitration of the tumor suppressor protein p53 at tyrosine 327 promotes p53 oligomerization and activation. Biochemistry 2010, 49, 5331–5339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubbo, H.; Radi, R. Protein and lipid nitration: Role in redox signaling and injury. Biochim. Biophys. Acta 2008, 1780, 1318–1324. [Google Scholar] [CrossRef] [PubMed]
- Lanone, S.; Manivet, P.; Callebert, J.; Launay, J.M.; Payen, D.; Aubier, M.; Boczkowski, J.; Mebazaa, A. Inducible nitric oxide synthase (NOS2) expressed in septic patients is nitrated on selected tyrosine residues: Implications for enzymic activity. Biochem. J. 2002, 366, 399–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohinata, Y.; Miller, J.M.; Schacht, J. Protection from noise-induced lipid peroxidation and hair cell loss in the cochlea. Brain Res. 2003, 966, 265–273. [Google Scholar] [CrossRef]
- Foster, M.W. S-nitrosylation in health and disease. Trends Mol. Med. 2003, 9, 160–168. [Google Scholar] [CrossRef]
- Kovacs, L.; Lindermayr, T. Nitric oxide-based protein modification: Formation and site-specificity of protein S-nitrosylation. Front. Plant Sci. 2013, 4, 137. [Google Scholar] [CrossRef] [Green Version]
- Hess, D.T.; Matsumoto, A.; Kim, S.; Marshall, H.E.; Stamler, J.S. Protein S-nitrosylation: Purview and parameters. Nat. Rev. Mol. Cell Biol. 2005, 6, 150–166. [Google Scholar] [CrossRef]
- Nakamura, T.; Lipton, S.A. Emerging roles of S-nitrosylation in protein misfolding and neurodegenerative diseases. Antioxid. Redox Signal. 2008, 10, 87–101. [Google Scholar] [CrossRef]
- Fukuyama, N.; Takizawa, S.; Ishida, H.; Hoshiai, K.; Shinohara, Y.; Nakazawa, H. Peroxynitrite formation in focal cerebral ischemia-reperfusion in rats occurs predominantly in the peri-infarct region. J. Cereb. Blood Flow Metab. 1998, 18, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Ozdemir, Y.; Bolay, H.; Saribas, O.; Dalkara, T. Role of Endothelial Nitric Oxide Generation and Peroxynitrite Formation in Reperfusion Injury After Focal Cerebral Ischemia. Stroke 2000, 31, 1974–1980. [Google Scholar] [CrossRef]
- Park, J.S.; Jou, I.; Park, S.M. Attenuation of noise-induced hearing loss using methylene blue. Cell Death Dis. 2014, 5, e1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, W.; Shi, X.; Nuttall, A. Noise-induced nitrotyrosine increase and outer hair cell death in guinea pig cochlea. Chin. Med. J. 2013, 126, 2923–2927. [Google Scholar] [PubMed]
- Yuan, H.; Wang, X.; Hill, K.; Chen, J.; Lemasters, J.; Yang, S.; Sha, S. Autophagy Attenuates Noise-Induced Hearing Loss by Reducing Oxidative Stress. Antioxid. Redox Signal. 2015, 22, 1308–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, J.M.; Daikhin, E.; Yudkoff, M.; Raman, C.S.; Ischiropoulos, H. Factors determining the selectivity of protein tyrosine nitration. Arch. Biochem. Biophys. 1999, 371, 169–178. [Google Scholar] [CrossRef] [PubMed]
- MacMillan-Crow, L.A.; Crow, J.P.; Kerby, J.D.; Beckman, J.S.; Thompson, J.A. Nitration and inactivation of manganese superoxide dismutase in chronic rejection of human renal allografts. Proc. Natl. Acad. Sci. USA 1996, 93, 11853–11858. [Google Scholar] [CrossRef] [Green Version]
- Cassina, A.M.; Hodara, R.; Souza, J.M.; Thomson, L.; Castro, H.; Ischiropoulos, H.; Freeman, B.; Radi, R. Cytochrome C nitration by peroxynitrite. J. Biol. Chem. 2000, 275, 21409–21415. [Google Scholar] [CrossRef] [Green Version]
- Savvides, S.N.; Scheiwein, M.; Bohme, C.C.; Artee, G.E.; Karplus, A.; Becker, K.; Heiner, R.S. Crystal structure of the antioxidant enzyme glutathione reductase inactivated by peroxynitrite. J. Biol. Chem. 2002, 277, 2779–2784. [Google Scholar] [CrossRef] [Green Version]
- Zou, M.; Martin, C.; Ullrich, V. Tyrosine nitration as a mechanism of selective inactivation of prostacyclin synthase by peroxynitrite. Biol. Chem. 1997, 378, 707–713. [Google Scholar] [CrossRef]
- Blanchard-Fillion, B.; Servy, C.; Ducrocq, C. 1-Nitrosomelatonin is a spontaneous NO-releasing compound. Free Radic. Res. 2001, 35, 857–866. [Google Scholar] [CrossRef]
- Vadseth, C.; Souza, J.M.; Thomson, L.; Seagraves, A.; Nagaswami, C.; Scheiner, T.; Torbet, J.; Vilaire, G.; Bennett, J.S.; Murciano, J.; et al. Pro-thrombotic state induced by post-translational modification of fibrinogen by reactive nitrogen species. J. Biol. Chem. 2004, 279, 8820–8826. [Google Scholar] [CrossRef] [Green Version]
- Hodara, R.; Norris, E.H.; Giasson, B.I.; Mishizen-Eberz, A.J.; Lynch, D.R.; Lee, V.; Ischiropoulos, H. Functional consequences of alpha-synuclein tyrosine nitration: Diminished binding to lipid vesicles and increased fibril formation. J. Biol. Chem. 2004, 279, 47746–47753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gole, M.D.; Souza, J.M.; Choi, I.; Hertkorn, C.; Malcolm, S.; FoustIII, R.F.; Finkel, B.; Lanken, P.N.; Ischiropoulos, H. Plasma proteins modified by tyrosine nitration in acute respiratory distress syndrome. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 278, L961–L967. [Google Scholar] [CrossRef] [PubMed]
- Forrester, M.T.; Benhar, M.; Stamler, J.S. Nitrosative stress in the ER: A new role for S-nitrosylation in neurodegenerative diseases. ACS Chem. Biol. 2006, 1, 355–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benhar, M.; Stamler, J.S. A central role for S-nitrosylation in apoptosis. Nat. Cell Biol. 2005, 7, 645–646. [Google Scholar] [CrossRef]
- Ozawa, K.; Komatsubara, A.T.; Nishimura, Y.; Sawada, T.; Kawafune, H.; Tsumoto, H.; Tsuji, Y.; Zhao, J.; Kyotani, Y.; Tanaka, T.; et al. S-nitrosylation regulates mitochondrial quality control via activation of parkin. Sci. Rep. 2013, 3, 2202. [Google Scholar] [CrossRef] [Green Version]
- Singel, D.S.; Stamler, J.S. Chemical physiology of blood flow regulation by red blood cells: The role of nitric oxide and S-nitrosohemoglobin. Annu. Rev. Physiol. 2005, 67, 99–145. [Google Scholar] [CrossRef]
- Jamesdaniel, S.; Manohar, S.; Hinduja, S. Is S-nitrosylation of cochlear proteins a critical factor in cisplatin-induced ototoxicity? Antioxid. Redox Signal. 2012, 17, 929–933. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, D.; Jiang, H.; Schacht, J.; Miller, J.M. Delayed production of free radicals following noise exposure. Brain Res. 2004, 1019, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, D.; Jiang, H.Y.; LePrell, C.G.; Schacht, J.; Miller, J.M. Post-exposure treatment attenuates noise-induced hearing loss. Neuroscience 2005, 134, 633–642. [Google Scholar] [CrossRef]
- Xu, F.; Cheng, Y.; Yan, W. Up-regulation of autophagy and apoptosis of cochlear hair cells in mouse models for deafness. Arch. Med. Sci. 2018, 17, 535–541. [Google Scholar] [CrossRef]
- Yamasoba, T.; Someya, S.; Yamada, C.; Weindruch, R.; Prolla, T.A.; Tanokura, M. Role of mitochondrial dysfunction and mitochondrial DNA mutations in age-related hearing loss. Hear. Res. 2007, 226, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Ruan, R.S. Possible roles of nitric oxide in the physiology and pathophysiology of the mammalian cochlea. Ann. N. Y. Acad. Sci. 2002, 962, 260–274. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, R.; Yamaguchi, T.; Tanaka, H.; Ogita, K. Mechanism underlying the protective effect of tempol and Nω-nitro-L-arginine methyl ester on acoustic injury: Possible involvement of c-JunNterminal kinase pathway and connexin26 in the cochlear spiral ligament. J. Pharmacol. Sci. 2010, 114, 50–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Ren, T.; Nuttall, A.L. The electrochemical and fluorescence detection of nitric oxide in the cochlea and its increase following loud sound. Hear. Res. 2002, 164, 49–58. [Google Scholar] [CrossRef]
- Honkura, Y.; Matsuo, H.; Murakami, S.; Sakiyama, M.; Mizutari, K.; Shiotani, A.; Yamamoto, M.; Morita, I.; Shinomiya, N.; Kawase, T.; et al. NRF2 Is a Key Target for Prevention of Noise-Induced Hearing Loss by Reducing Oxidative Damage of Cochlea. Sci. Rep. 2016, 6, 19329. [Google Scholar] [CrossRef] [Green Version]
- Alvarado, J.; Fuentes-Santamaría, V.; Gabaldón-Ull, M.; Jareño-Flores, T.; Miller, J.; Juiz, J.M. Noise-Induced “Toughening” Effect in Wistar Rats: Enhanced Auditory Brainstem Responses Are Related to Calretinin and Nitric Oxide Synthase Upregulation. Front. Neuroanat. 2015, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Jamesdaniel, S.; Coling, D.; Hinduja, S.; Ding, D.; Li, J.; Cassidy, L.; Seige, G.M.; Qu, J.; Salvi, R. Cisplatin-induced ototoxicity is mediated by nitroxidative modification of cochlear proteins characterized by nitration of Lmo4. J. Biol. Chem. 2012, 287, 18674–18686. [Google Scholar] [CrossRef] [Green Version]
- Jamesdaniel, S.; Rathinam, R.; Neumann, W. Targeting nitrative stress for attenuating cisplatin-induced downregulation of cochlear LIM domain only 4 and ototoxicity. Redox Biol. 2015, 10, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Jamesdaniel, S. Downstream Targets of Lmo4 Are Modulated by Cisplatin in the Inner Ear of Wistar Rats 2014. PLoS ONE 2014, 9, e115263. [Google Scholar] [CrossRef] [Green Version]
- Heinrich, U.-R.; Selivanova, O.; Feltens, R.; Brieger, J.; Mann, W. Endothelial nitric oxide synthase up-regulation in the guinea pig organ of Corti after acute noise trauma. Brain Res. 2005, 1047, 85–96. [Google Scholar] [CrossRef]
- Jia, Z.; He, Q.; Shun, C.; Li, F. Tauroursodeoxycholic acid attenuates gentamicin-induced cochlear hair cell death in vitro. Toxicol. Lett. 2018, 294, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Rathinam, R.; Ghosh, S.; Neumann, W.L.; Jamesdaniel, S. Cisplatin-induced apoptosis in auditory, renal, and neuronal cells is associated with nitration and downregulation of LMO4. Cell Death Discov. 2015, 1, 15052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahab, M.; Rosati, R.; Meyers, D.; Sheilds, J.; Crofts, E.; Baker, T.R.; Jamesdaniel, S. Cisplatin-induced hair cell loss in zebrafish neuromasts is accompanied by protein nitration and Lmo4 degradation. Toxicol. Appl. Pharmacol. 2021, 410, 115342. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Talaska, A.E.; Schacht, J.; Sha, S.H. Oxidative imbalance in the aging inner ear. Neurobiol. Aging 2007, 28, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishiyama, G.; Wester, J.; Lopez, I.A.; Beltran-Parrazal, L.; Ishiyama, A. Oxidative Stress in the Blood Labyrinthine Barrier in the Macula Utricle of Meniere’s Disease Patients. Front. Physiol. 2018, 9, 1068. [Google Scholar] [CrossRef] [PubMed]
- Vlajkovic, S.M.; Lee, K.; Wong, A.H.; Guo, C.; Gupta, R.; Housley, G.; Thorne, P. Adenosine aminecongener mitigates noise-induced cochlear injury. Purinergic Signal. 2010, 6, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Rosati, R.; Shahab, M.; Neumann, W.; Jamesdaniel, S. Inhibition of protein nitration prevents cisplatin-induced inactivation of STAT3 and promotes anti-apoptotic signaling in organ of Corti cells. Exp. Cell Res. 2019, 381, 105–111. [Google Scholar] [CrossRef]
- Diao, M.; Gao, W.; Sun, J. Nitric oxide synthase inhibitor reduces noise-induced cochlear damage in guinea pigs. Acta Otolaryngol. 2007, 127, 1162–1167. [Google Scholar] [CrossRef]
- Wu, G.; Lee, T.D.; Moore, R.E.; Rao, N.A. Photoreceptor mitochondrial tyrosine nitration in experimental uveitis. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2271–2281. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.H.; Li, Y.N.; Qi, J.S.; Jia, X.X. Peroxynitrite-induced protein nitration is responsible for renal mitochondrial damage in diabetic rat. J. Endocrinol. Investig. 2010, 33, 140–146. [Google Scholar] [CrossRef]
- Greenacre, S.A.; Ischiropoulos, H. Tyrosine nitration: Localization, quantification, consequences for protein function and signal transduction. Free Radic. Res. 2001, 34, 541–581. [Google Scholar] [CrossRef] [PubMed]
- Pourbakht, A.; Yamasoba, T. Ebselen attenuates cochlear damage caused by acoustic trauma. Hear. Res. 2005, 181, 100–108. [Google Scholar] [CrossRef]
- Lynch, E.; Gua, R.; Piercea, C.; Kil, J. Combined oral delivery of ebselen and allopurinol reduces multiple cisplatin toxicities in rat breast and ovarian cancer models while enhancing anti-tumor activity. Anticancer Drugs 2005, 16, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Kil, J.; Lobarinas, E.; Spankovich, C.; Griffiths, S.; Antonelli, P.; Lynch, E.; Prell, C. Safety and efficacy of ebselen for the prevention of noise-induced hearing loss: A randomized, double-blind, placebo-controlled, phase 2 trial. Lancet 2017, 390, 969–979. [Google Scholar] [CrossRef]
- Wu, S.B.; Wu, Y.T.; Wu, T.P.; Wei, Y.H. Role of AMPK-mediated adaptive responses in human cells with mitochondrial dysfunction to oxidative stress. Biochem. Biophys. Acta 2020, 1840, 1331–1344. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, C.; Martin-Sanz, E.; Cuadrado, E.; Granizo, J.J.; Sanz-Fernandez, R. Protective effect of polyphenols on presbycusis via oxidative/nitrosative stress suppression in rats. Exp. Gerontol. 2016, 83, 31–36. [Google Scholar] [CrossRef]
- Xiong, M.; Lai, H.; He, Q.; Wang, J. Astragaloside IV attenuates impulse noise-induced trauma in guinea pig. Acta Otolaryngol. 2011, 131, 809–816. [Google Scholar] [CrossRef]
- Yang, D.H.; Xie, J.; Liu, K.; Peng, Z.; Guo, J.Y.; Yu, S.K.; Wang, G.P.; Gong, S.S. The histone deacetylase inhibitor sodium butyrate protects against noise-induced hearing loss in Guinea pigs. Neurosci. Lett. 2017, 660, 140–146. [Google Scholar] [CrossRef]
- Lu, S.; Fan, Z.; Xu, W.; Han, Y.; Zhang, G.; Liu, W.; Bai, X.; Wang, X.; Xin, H.; Li, J.; et al. L-cysteine attenuates peroxynitrite-elicited cytotoxicity to spiral ganglion neurons: Possible relation to hearing loss. Neurol. Res. 2011, 33, 935–941. [Google Scholar] [CrossRef]
- Ciodaro, F.; Freni, F.; Alberti, G.; Forelli, M.; Gazia, F.; Bruno, R.; Sherdell, E.P.; Galletti, B.; Galletti, F. Application of Cervical Vestibular-Evoked Myogenic Potentials in Adults with Moderate to Profound Sensorineural Hearing Loss: A Preliminary Study. Int. Arch. Otorhinolaryngol. 2020, 24, e5–e10. [Google Scholar] [CrossRef] [Green Version]
- Toumpas, C.J.; Clark, J.; Harris, A.; Beswick, R.; Nourse, C.B. Congenital cytomegalovirus infection is a significant cause of moderate to profound sensorineural hearing loss in Queensland children. J. Paediatr. Child Health 2015, 51, 541–544. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Author/Year | Model (Animal/Cell Culture) | Nitrative Stress Molecule | Ototraumatic Agent | Region Affected in Inner Ear |
|---|---|---|---|---|
| Alvarado et al., 2015 | Wistar rats | nNOS | Noise | Spiral ganglion, spiral ligament, and cochlear nerves |
| Han et al., 2013 | Guinea pigs | 3-NT | Noise | OHCs, IHCs, spiral ganglion, and pillar cells |
| Jamesdaniel et al., 2012 | UBOC1 cell lines | 3-NT | Cisplatin | UBOC1 cells |
| Jamesdaniel, 2014 | Wistar rats | 3-NT | Cisplatin | Spiral ganglion, stria vascularis, and organ of Corti |
| Jamesdaniel et al., 2016 | UBOC1 cell lines | 3-NT | Cisplatin | UBOC1 cells |
| Jiang et al., 2007 | CBA/J mice | 3-NT, OONO− | Aging | Deiters’ cells, pillar cells, stria vascularis, spiral ganglion |
| Lu et al., 2011 | Rats | 3-NT | OONO- | Spiral ganglion neurons |
| Lynch et al., 2005 | Female rats F-344 | 3-NT | Cisplatin | OHCs |
| Nagashima et al., 2010 | Std-ddY mice | 3-NT | Noise | Spiral ligament |
| Pourbakht et al., 2005 | Guinea pigs | 3-NT | Noise | OHCs |
| Rathinam et al., 2015 | UBOC1, HK-2, and SH-SY5Y cell lines | 3-NT | Cisplatin | UBOC1, HK-2, and SH-SY5Y cells |
| Rosati et al., 2019 | UBOC1 cell lines | 3-NT | Cisplatin | UBOC1 cells |
| Shahab et al., 2020 | Zebrafish | 3-NT | Cisplatin | Hair cells in neuromast |
| Shi et al., 2002 | Guinea pigs | 3-NT | Noise | OHCs |
| Vlajkovic et al., 2010 | Wistar rats | 3-NT | Noise | Organ of Corti, spiral ganglion neurons, spiral ligament, spiral limbus, inner sulcus cells, inner phalangeal cells, pillar cells, Deiters’ cells, Hensen’s cells |
| Wu et al., 2020 | CBA/J mice and HEI-OC1 cells | 3-NT | Noise | OHCs |
| Xiong et al., 2011 | Guinea pigs | 3-NT and iNOS synthase | Noise | Stria vascularis, spiral ligament, organ of Corti |
| Yamasoba et al., 2007 | Guinea pigs | NO | Noise | Organ of Corti, afferent dendrites beneath IHCs |
| Yamashita et al., 2004 | Guinea pigs | 3-NT | Noise | Spiral ganglion, organ of Corti, lateral wall |
| Yamashita et al., 2005 | Guinea pigs | 3-NT | Noise | OHCs |
| Yang et al., 2017 | Guinea pigs | 3-NT | Noise | OHCs |
| Yuan et al., 2015 | CBA/J mice | 3-NT | Noise | IHCs and OHCs |
| Author/Year | Model (Animal/Cell Culture) | Inhibitors of Nitration | Biochemical Effect of Intervention | Outcome of Intervention |
|---|---|---|---|---|
| Diao et al., 2007 | Guinea pigs | NG-Nitro-L-arginine methyl ester (L-NAME) | Decreased NO production | L-NAME protected the cochlea after noise exposure |
| Jamesdaniel et al., 2012 | Wistar rats | Trolox | Decreased nitration of cochlear proteins | Attenuated cisplatin-induced OHC loss and hearing threshold shifts |
| Jamesdaniel, 2014 | Wistar rats | Trolox | Decreased nitration of cochlear proteins | Attenuated cisplatin-induced ototoxicity and nitration of cochlear proteins |
| Jamesdaniel et al., 2016 | UBOC1 cell lines | SRI110 | Decreased 3-NT | SRI110 inhibited cisplatin-induced cytotoxicity |
| Jia et al., 2018 | HEI-OC1 cells and explanted cochlear tissue | Tauroursodeoxycholic acid (TUDCA) | Decreased NO production | SRI110 inhibited cisplatin-induced cytotoxicity |
| Lu et al., 2011 | Rats | L-Cysteine | Decreased peroxynitrite | Decreased gentamicin-induced ototoxicity |
| Lyncha et al., 2005 | Fisher-344 rats | Ebselen | Decreased lipid peroxidation | Protected cochlea from cisplatin-induced OHC loss and hearing loss |
| Nagashima et al., 2010 | Std-ddY mice | Tempol and Nω-nitro-L-arginine methyl ester | Decreased 3-NT and 4-HNE | Protected noise exposed animals from hearing loss |
| Pourbakht et al., 2005 | Guinea pigs | Ebselen | Decreased peroxynitrite | Reduced the noise-induced permanent threshold shifts |
| Rosati et al., 2019 | UBOC1 cell lines | SRI110 | Decreased 3-NT | Prevented cisplatin-induced cytotoxicity |
| Sánchez-Rodríguez et al., 2016 | Sprague-Dawley Rats | Polyphenols | Decreased 3-NT and superoxide anions, increased glutathione peroxidase and SOD | Protected against age-related hearing loss |
| Wu et al., 2020 | CBA/J mice and HEI-OC1 cells | N-Acetyl cysteine (NAC) | Decreased 3-NT and superoxides | Prevented loss of OHCs induced by noise exposure |
| Vlajkovic et al., 2010 | Wistar rats | Adenosine amine congener (ADAC) | Decreased 3-NT | Attenuated hearing threshold shifts after noise exposure |
| Xiong et al., 2011 | Guinea pigs | Astragaloside IV | Inhibited iNOS | Protected the cochlea from noise-induced damage |
| Yamashita et al., 2005 | Guinea pigs | Salicylate and trolox | Decreased 3-NT and superoxides | Reduced ABR shifts post noise exposure |
| Yamasoba et al., 2005 | Guinea pigs | Ebselen | Decreased NO production | Reduced ABR shifts post noise exposure |
| Yang et al., 2017 | Guinea pigs | Sodium butyrate (SB) | Decreased 3-NT and HDAC1 | Attenuated noise-induced loss of OHC and hearing loss |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahab, M.; Jamesdaniel, S. Nitrative Stress and Auditory Dysfunction. Pharmaceuticals 2022, 15, 649. https://doi.org/10.3390/ph15060649
Shahab M, Jamesdaniel S. Nitrative Stress and Auditory Dysfunction. Pharmaceuticals. 2022; 15(6):649. https://doi.org/10.3390/ph15060649
Chicago/Turabian StyleShahab, Monazza, and Samson Jamesdaniel. 2022. "Nitrative Stress and Auditory Dysfunction" Pharmaceuticals 15, no. 6: 649. https://doi.org/10.3390/ph15060649
APA StyleShahab, M., & Jamesdaniel, S. (2022). Nitrative Stress and Auditory Dysfunction. Pharmaceuticals, 15(6), 649. https://doi.org/10.3390/ph15060649