
Enhancing Colistin Activity against Colistin-Resistant Escherichia coli through Combination with Alginate Nanoparticles and Small Molecules

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Combination of Colistin with Alg NPs Has Increased Antibacterial Activity
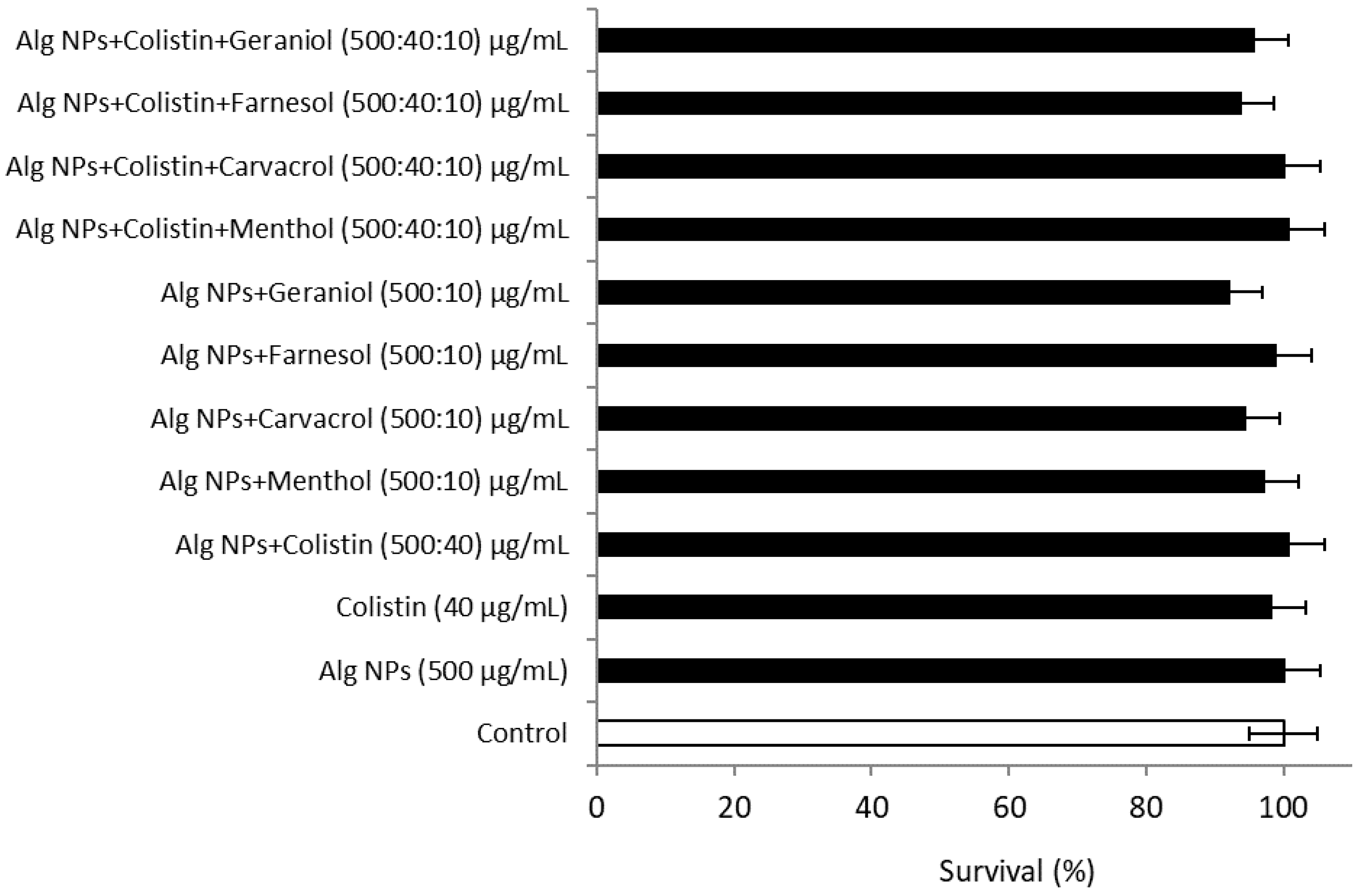
2.2. Combination of Colistin with Alg NPs and Components of Essential Oils
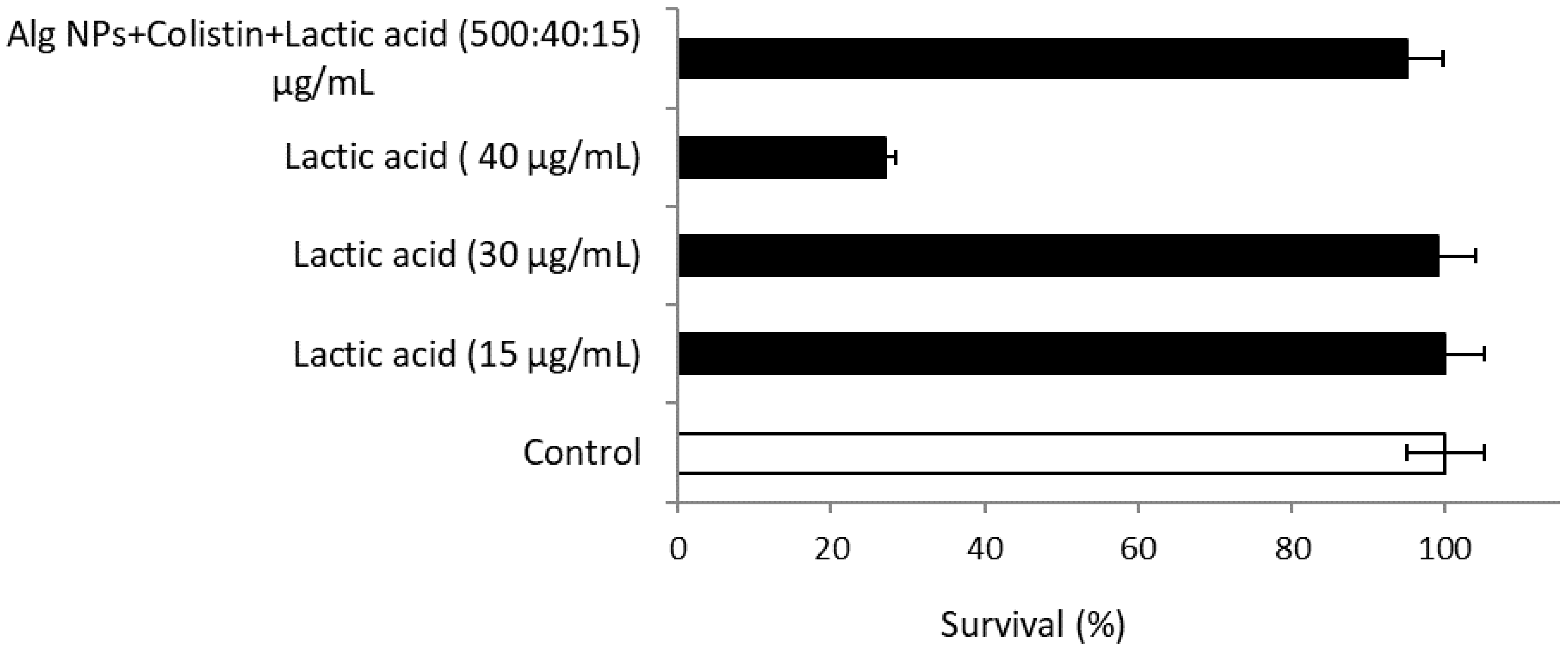
2.3. Combination of Colistin with Alg NPs and Lactic Acid
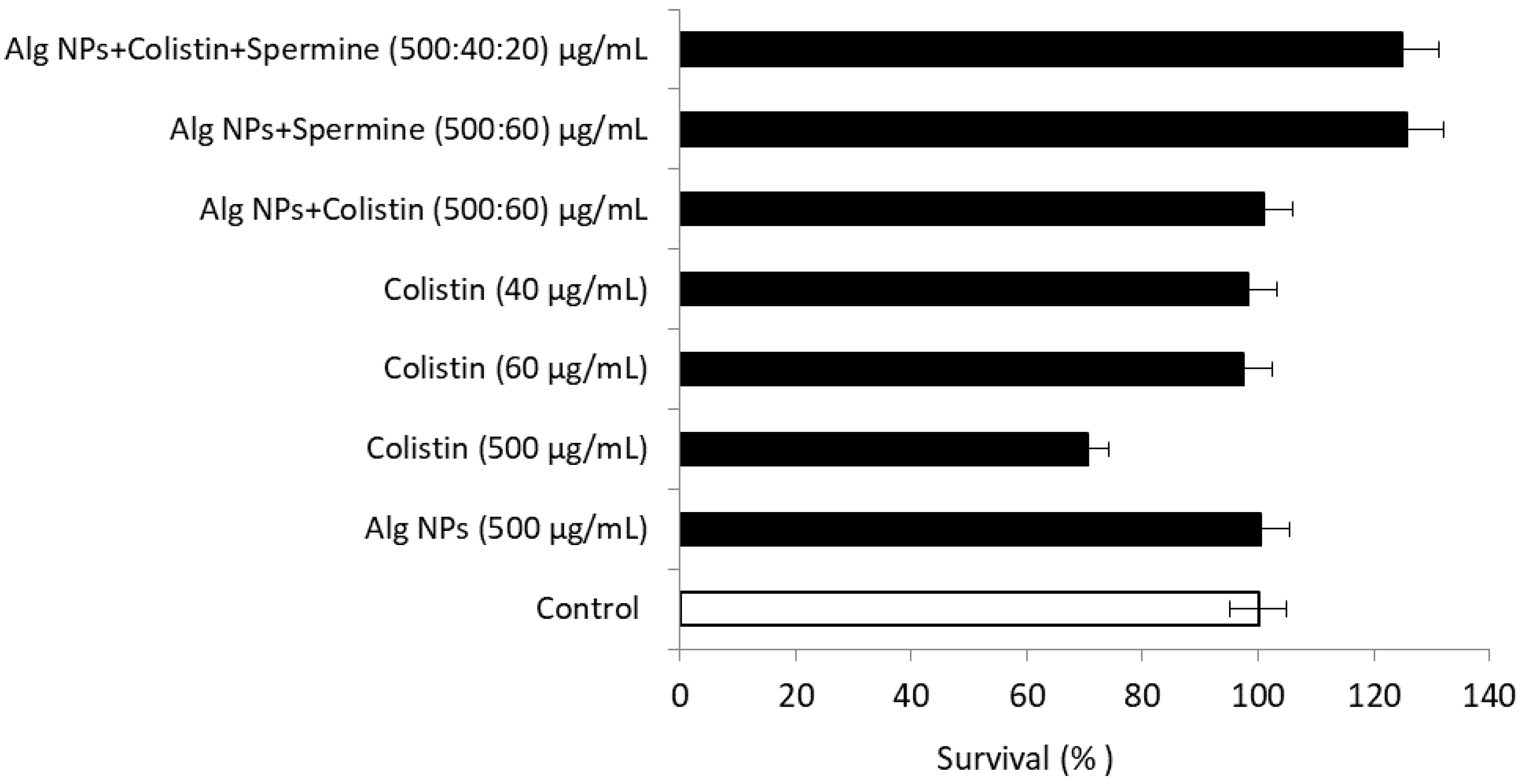
2.4. Combination of Colistin with Alg NPs and Polyamines (Spermine, Spermidine, Piperazine)
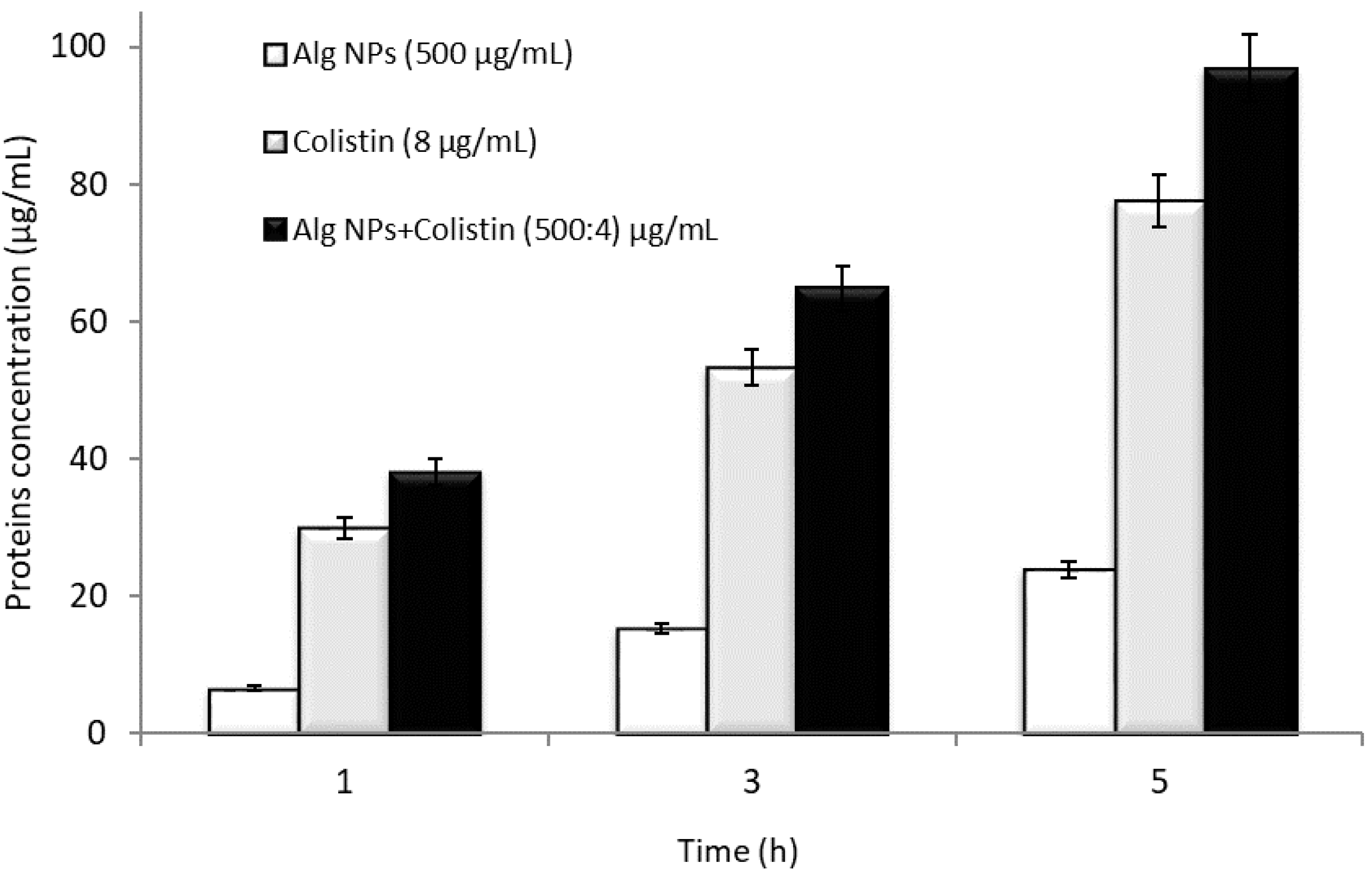
2.5. Mode of Action of the Alg NPs+Colistin Formulation
2.6. Determination of the Bacteriostatic or Bactericidal Effect of Colistin in the Different Formulations
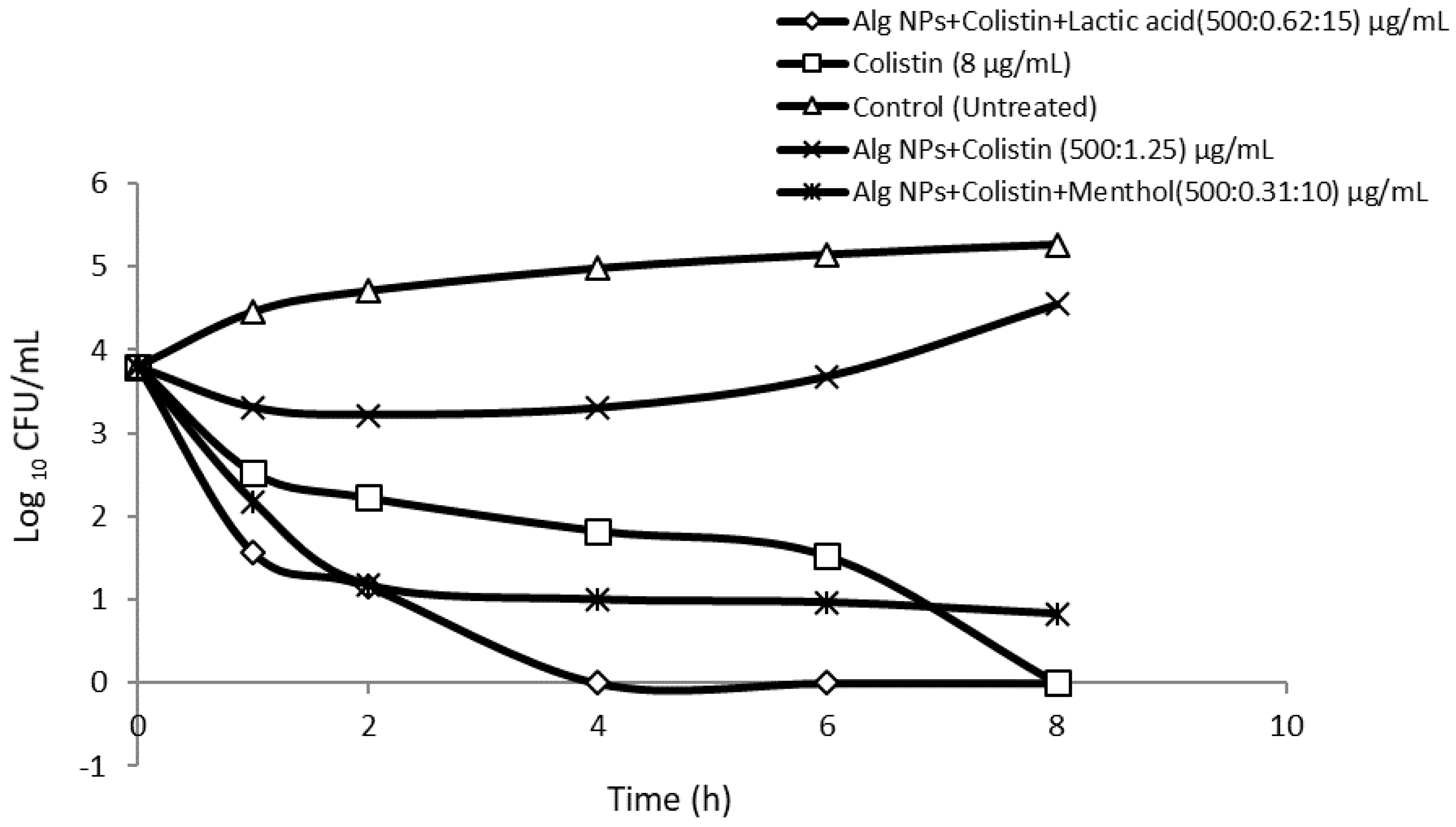
2.7. Killing Curve Assays
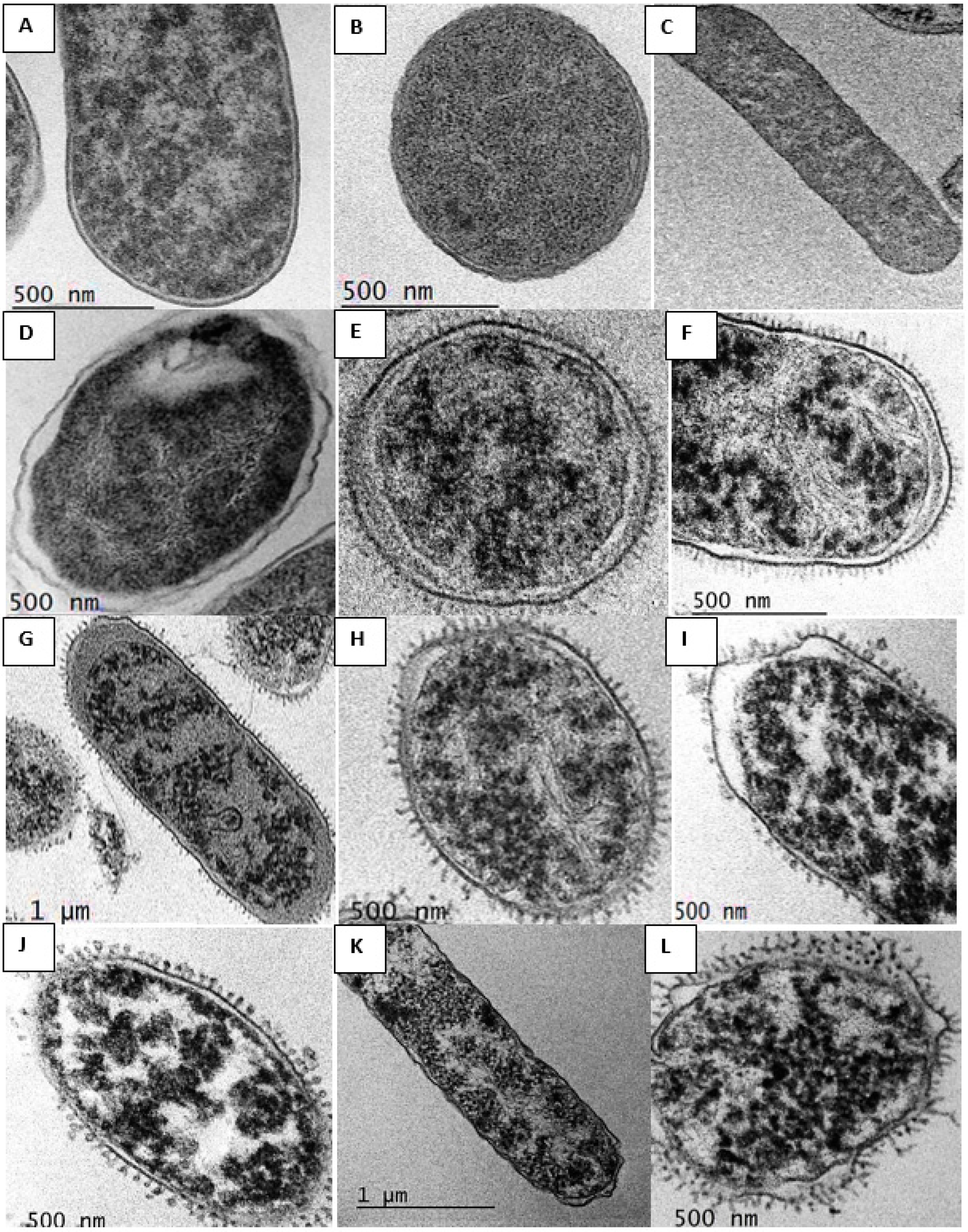
2.8. Transmission Electron Microscopy (TEM) Studies
2.9. Stability of Alg NPs-Colistin-Small Molecules in Conditions Mimicking the Passage through the Gastrointestinal Tract
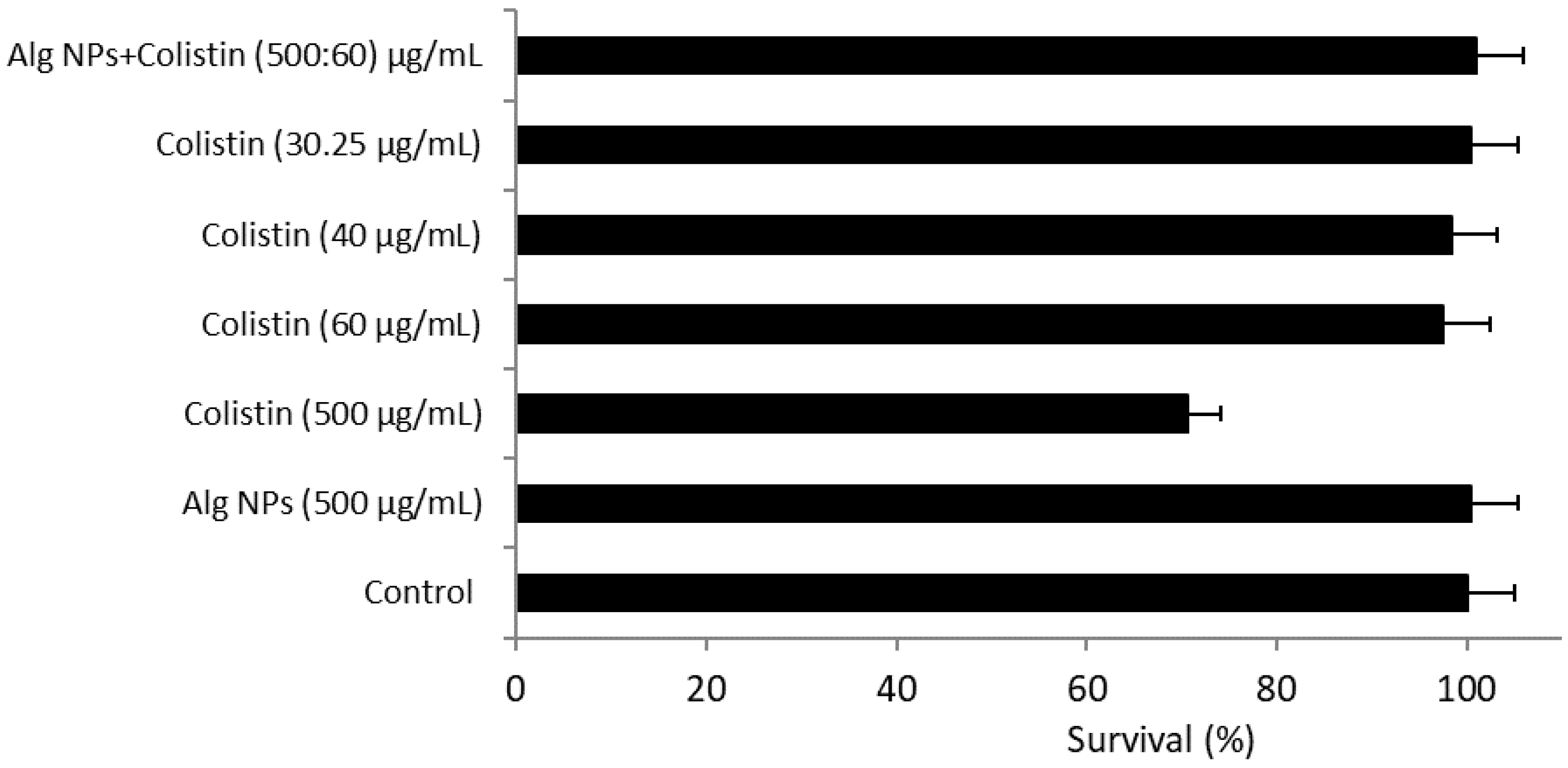
2.10. Cytotoxicity Assays
2.10.1. Alg NPs+Colistin Formulations
2.10.2. Alg NPs+Colistin+Components of Essential Oil Formulations
2.10.3. Alg NPs+Colistin+Lactic Acid
2.10.4. Alg NPs+Colistin+Spermine Formulation
3. Experimental Section
3.1. Escherichia coli (E. coli) Strains Used in This Study
3.2. Colistin Calibration Curve
3.3. Formation of Alginate Nanoparticles (Alg NPs)
3.4. Preparation of Colistin-Loaded Alg NPs and Quantification
3.5. Preparation of Alg NPs Loaded with Colistin and Small Molecules (Components of Essential Oils, Polyamines, Organic Acids) Formulations
- -
- Alg NPs+Colistin+Components of essential oil (components of essential oils = menthol, farnesol, geraniol or carvacrol) (Sigma-Aldrich, St. Quentin Fallavier, France) formulations at different concentrations of colistin (30, 40 or 50) µg/mL and fixed concentration of component of essential oils (10 μg/mL), chosen on the light of preliminary cytotoxicity assay (Figure S1).
- -
- Alg NPs+Colistin+Polyamines (spermine or piperazine) (Sigma-Aldrich, St. Quentin Fallavier, France) formulations: colistin = 40 µg/mL and polyamine = 20 μg/mL.
- -
- Alg NPs+Colistin+Spermidine (Sigma-Aldrich, St. Quentin Fallavier, France) formulations: colistin = 30, 20, or 10 µg/mL and spermidine = 30, 40, and 50 µg/mL.
- -
- Alg NPs+Colistin+Lactic acid (Sigma-Aldrich, St. Quentin Fallavier, France) formulation: colistin = 40 μg/mL and lactic acid = 15 μg/mL.
3.6. Determination of Minimal Inhibitory Concentration (MIC) and Minimal Bactericidal Concentrations (MBC)
3.7. Effect of Alg NPs and Alg NPs+Colistin on Leakage of Proteins on E. coli 184 (mcr-1) Strain
3.8. Killing Curves
3.9. Transmission Electron Microscopy (TEM)
3.10. Evaluation of Alg NPs-Colistin-Small Molecules Formulations Stability in the Conditions Mimicking the Passage through the Gastrointestinal Tract
3.11. Cytotoxicity Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Desselberger, U. Emerging and Re-Emerging Infectious Diseases. J. Infect. 2000, 40, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.I.; Steele, D.; Aguado, T. Ad Hoc ETEC Technical Expert Committee Analysis of Strategies to Successfully Vaccinate Infants in Developing Countries against Enterotoxigenic E. coli (ETEC) Disease. Vaccine 2007, 25, 2545–2566. [Google Scholar] [CrossRef] [PubMed]
- Witte, W. International dissemination of antibiotic resistant strains of bacterial pathogens. Infect. Genet Evol. 2004, 4, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; McArthur, J.V. Co-Selection of Antibiotic and Metal Resistance. Trends Microbiol. 2006, 14, 176–182. [Google Scholar] [CrossRef] [PubMed]
- De, S.; Robinson, D. Polymer Relationships during Preparation of Chitosan–Alginate and Poly-l-Lysine–Alginate Nanospheres. J. Control. Release 2003, 89, 101–112. [Google Scholar] [CrossRef]
- Florence, A.T. The Oral Absorption of Micro- and Nanoparticulates: Neither Exceptional nor Unusual. Pharm. Res. 1997, 14, 259–266. [Google Scholar] [CrossRef]
- Huh, A.J.; Kwon, Y.J. “Nanoantibiotics”: A New Paradigm for Treating Infectious Diseases Using Nanomaterials in the Antibiotics Resistant Era. J. Control. Release 2011, 156, 128–145. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Kasiakou, S.K. Colistin: The Revival of Polymyxins for the Management of Multidrug-Resistant Gram-Negative Bacterial Infections. Clin. Infect. Dis. 2005, 40, 1333–1341. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [Green Version]
- Bergen, P.J.; Landersdorfer, C.B.; Lee, H.J.; Li, J.; Nation, R.L. “Old” Antibiotics for Emerging Multidrug-Resistant Bacteria. Curr. Opin. Infect. Dis. 2012, 25, 626–633. [Google Scholar] [CrossRef] [Green Version]
- Drider, D.; Boukherroub, R.; Le Devendec, L.; Belguesmia, Y.; Hazime, N.; Mourand, G.; Paboeuf, F.; Kempf, I. Impact of Colistin and Colistin-Loaded on Alginate Nanoparticles on Pigs Infected with a Colistin-Resistant Enterotoxigenic Escherichia coli Strain. Vet. Microbiol. 2022, 266, 109359. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Valiakos, G.; Kapna, I. Colistin Resistant Mcr Genes Prevalence in Livestock Animals (Swine, Bovine, Poultry) from a Multinational Perspective. A Systematic Review. Vet. Sci. 2021, 8, 265. [Google Scholar] [CrossRef] [PubMed]
- Hussein, N.H.; Al-Kadmy, I.M.S.; Taha, B.M.; Hussein, J.D. Mobilized Colistin Resistance (Mcr) Genes from 1 to 10: A Comprehensive Review. Mol. Biol. Rep. 2021, 48, 2897–2907. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Abbas, M.; Rehman, M.U.; Wang, M.; Jia, R.; Chen, S.; Liu, M.; Zhu, D.; Zhao, X.; Gao, Q.; et al. Updates on the Global Dissemination of Colistin-Resistant Escherichia Coli: An Emerging Threat to Public Health. Sci. Total Environ. 2021, 799, 149280. [Google Scholar] [CrossRef]
- Tønnesen, H.H.; Karlsen, J. Alginate in Drug Delivery Systems. Drug Dev. Ind. Pharm. 2002, 28, 621–630. [Google Scholar] [CrossRef]
- Wee, S.; Gombotz, W.R. Protein Release from Alginate Matrices. Adv. Drug Deliv. Rev. 1998, 31, 267–285. [Google Scholar]
- Goh, C.H.; Heng, P.W.S.; Chan, L.W. Alginates as a Useful Natural Polymer for Microencapsulation and Therapeutic Applications. Carbohydr. Polym. 2012, 88, 1–12. [Google Scholar] [CrossRef]
- Shilpa, A.; Agrawal, S.S.; Ray, A.R. Controlled Delivery of Drugs from Alginate Matrix. J. Macromol. Sci. 2003, 43, 187–221. [Google Scholar] [CrossRef]
- Yang, J.-S.; Xie, Y.-J.; He, W. Research Progress on Chemical Modification of Alginate: A Review. Carbohydr. Polym. 2011, 84, 33–39. [Google Scholar] [CrossRef]
- Pawar, S.N.; Edgar, K.J. Alginate Derivatization: A Review of Chemistry, Properties and Applications. Biomaterials 2012, 33, 3279–3305. [Google Scholar] [CrossRef] [PubMed]
- Motelica, L.; Ficai, D.; Oprea, O.; Ficai, A.; Trusca, R.-D.; Andronescu, E.; Holban, A.M. Biodegradable Alginate Films with ZnO Nanoparticles and Citronella Essential Oil—A Novel Antimicrobial Structure. Pharmaceutics 2021, 13, 1020. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.; Paralikar, P.; Jogee, P.; Agarkar, G.; Ingle, A.P.; Derita, M.; Zacchino, S. Synergistic antimicrobial potential of essential oils in combination with nanoparticles: Emerging trends and future perspectives. Int. J. Pharm. 2017, 519, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.; Mallya, R.; Suvarna, V.; Asif Khan, T.; Momin, M.; Omri, A. Nanoparticles—Attractive Carriers of Antimicrobial Essential Oils. Antibiotics 2022, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- CLSI M100Ed30. Performance Standards for Antimicrobial Susceptibility Testing, 30th Edition. Available online: https://clsi.org/standards/products/microbiology/documents/m100/ (accessed on 30 April 2020).
- CA-SFM CASFM/EUCAST 2019. Société Française de Microbiologie 2019. Available online: https://www.sfm-microbiologie.org/wp-content/uploads/2019/02/CASFM2019_V1.0.pdf (accessed on 30 April 2020).
- Moffatt, J.H.; Harper, M.; Boyce, J.D. Mechanisms of Polymyxin Resistance. Adv. Exp. Med. Biol. 2019, 1145, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Stefaniuk, E.M.; Tyski, S. Colistin Resistance in Enterobacterales Strains—A Current View. Pol. J. Microbiol. 2019, 68, 417–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carattoli, A.; Villa, L.; Feudi, C.; Curcio, L.; Orsini, S.; Luppi, A.; Pezzotti, G.; Magistrali, C.F. Novel Plasmid-Mediated Colistin Resistance Mcr-4 Gene in Salmonella and Escherichia coli, Italy 2013, Spain and Belgium, 2015 to 2016. Euro Surveill. 2017, 22, 30589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Atya, A.K.; Abriouel, H.; Kempf, I.; Jouy, E.; Auclair, E.; Vachée, A.; Drider, D. Effects of Colistin and Bacteriocins Combinations on the In Vitro Growth of Escherichia coli Strains from Swine Origin. Probiotics Antimicrob. Proteins 2016, 8, 183–190. [Google Scholar] [CrossRef]
- Khan, S.; Tøndervik, A.; Sletta, H.; Klinkenberg, G.; Emanuel, C.; Onsøyen, E.; Myrvold, R.; Howe, R.A.; Walsh, T.R.; Hill, K.E.; et al. Overcoming Drug Resistance with Alginate Oligosaccharides Able to Potentiate the Action of Selected Antibiotics. Antimicrob. Agents Chemother. 2012, 56, 5134–5141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, M.F.; Powell, L.C.; Jack, A.A.; Powell, K.; Beck, K.; Florance, H.; Forton, J.; Rye, P.D.; Dessen, A.; Hill, K.E.; et al. A Low-Molecular-Weight Alginate Oligosaccharide Disrupts Pseudomonal Microcolony Formation and Enhances Antibiotic Effectiveness. Antimicrob. Agents Chemother. 2017, 61, e00762-17. [Google Scholar] [CrossRef] [Green Version]
- Winska, K.; Maczka, W.; Łyczko, J.; Grabarczyk, M.; Czubaszek, A.; Szumny, A. Essential Oils as Antimicrobial Agents—Myth or Real Alternative? Molecules 2019, 24, 2130. [Google Scholar] [CrossRef] [Green Version]
- Trombetta, D.; Castelli, F.; Sarpietro, M.G.; Venuti, V.; Cristani, M.; Daniele, C.; Saija, A.; Mazzanti, G.; Bisignano, G. Mechanisms of Antibacterial Action of Three Monoterpenes. Antimicrob. Agents Chemother. 2005, 49, 2474–2478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.-H.; Shin, S.-W. Activity of Essential Oil from Mentha Piperita against Some Antibiotic-Resistant Streptococcus pneumoniae Strains and Its Combination Effects with Antibiotics. Nat. Prod. Sci. 2007, 13, 164–168. [Google Scholar]
- Ettayebi, K.; El Yamani, J.; Rossi-Hassani, B. Synergistic Effects of Nisin and Thymol on Antimicrobial Activities in Listeria monocytogenes and Bacillus Subtilis. FEMS Microbiol. Lett. 2000, 183, 191–195. [Google Scholar] [CrossRef]
- Costa, A.F.; Silva, L.d.C.; Amaral, A.C. Farnesol: An Approach on Biofilms and Nanotechnology. Med. Mycol. 2021, 59, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Cristani, M.; D’Arrigo, M.; Mandalari, G.; Castelli, F.; Sarpietro, M.G.; Micieli, D.; Venuti, V.; Bisignano, G.; Saija, A.; Trombetta, D. Interaction of Four Monoterpenes Contained in Essential Oils with Model Membranes: Implications for Their Antibacterial Activity. J. Agric. Food Chem. 2007, 55, 6300–6308. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhou, F.; Ji, B.-P.; Pei, R.-S.; Xu, N. The Antibacterial Mechanism of Carvacrol and Thymol against Escherichia coli. Lett. Appl. Microbiol. 2008, 47, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kong, L.; Liu, C.; Ma, H. Synergistic Effect of Eugenol with Colistin against Clinical Isolated Colistin-Resistant Escherichia coli Strains. Antimicrob. Resist. Infect. Control 2018, 7, 17. [Google Scholar] [CrossRef]
- Lorenzi, V.; Muselli, A.; Bernardini, A.F.; Berti, L.; Pagès, J.-M.; Amaral, L.; Bolla, J.-M. Geraniol Restores Antibiotic Activities against Multidrug-Resistant Isolates from Gram-Negative Species. Antimicrob. Agents Chemother. 2009, 53, 2209–2211. [Google Scholar] [CrossRef] [Green Version]
- Jabra-Rizk, M.A.; Meiller, T.F.; James, C.E.; Shirtliff, M.E. Effect of Farnesol on Staphylococcus aureus Biofilm Formation and Antimicrobial Susceptibility. Antimicrob. Agents Chemother. 2006, 50, 1463–1469. [Google Scholar] [CrossRef] [Green Version]
- Koo, H.; Rosalen, P.L.; Cury, J.A.; Park, Y.K.; Bowen, W.H. Effects of Compounds Found in Propolis on Streptococcus mutans Growth and on Glucosyltransferase Activity. Antimicrob. Agents Chemother. 2002, 46, 1302–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brehm-Stecher, B.F.; Johnson, E.A. Sensitization of Staphylococcus aureus and Escherichia coli to Antibiotics by the Sesquiterpenoids Nerolidol, Farnesol, Bisabolol, and Apritone. Antimicrob. Agents Chemother. 2003, 47, 3357–3360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, H.; Holo, H. Synergistic Antimicrobial Activity Between the Broad Spectrum Bacteriocin Garvicin KS and Nisin, Farnesol and Polymyxin B Against Gram-Positive and Gram-Negative Bacteria. Curr. Microbiol. 2018, 75, 272–277. [Google Scholar] [CrossRef] [Green Version]
- Kostoulias, X.; Murray, G.L.; Cerqueira, G.M.; Kong, J.B.; Bantun, F.; Mylonakis, E.; Khoo, C.A.; Peleg, A.Y. Impact of a Cross-Kingdom Signaling Molecule of Candida albicans on Acinetobacter baumannii Physiology. Antimicrob. Agents Chemother. 2016, 60, 161–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellano, P.; Peña, N.; Ibarreche, M.P.; Carduza, F.; Soteras, T.; Vignolo, G. Antilisterial Efficacy of Lactobacillus Bacteriocins and Organic Acids on Frankfurters. Impact on Sensory Characteristics. J. Food Sci. Technol. 2018, 55, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Lajhar, S.A.; Brownlie, J.; Barlow, R. Survival Capabilities of Escherichia coli O26 Isolated from Cattle and Clinical Sources in Australia to Disinfectants, Acids and Antimicrobials. BMC Microbiol. 2017, 17, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mani-López, E.; García, H.S.; López-Malo, A. Organic Acids as Antimicrobials to Control Salmonella in Meat and Poultry Products. Food Res. Int. 2012, 45, 713–721. [Google Scholar] [CrossRef]
- Van Immerseel, F.; Russell, J.B.; Flythe, M.D.; Gantois, I.; Timbermont, L.; Pasmans, F.; Haesebrouck, F.; Ducatelle, R. The Use of Organic Acids to Combat Salmonella in Poultry: A Mechanistic Explanation of the Efficacy. Avian Pathol. 2006, 35, 182–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashket, E.R. Bioenergetics of Lactic Acid Bacteria: Cytoplasmic pH and Osmotolerance. FEMS Microbiol. Lett. 1987, 46, 233–244. [Google Scholar] [CrossRef]
- Kwon, D.-H.; Lu, C.-D. Polyamine Effects on Antibiotic Susceptibility in Bacteria. Antimicrob. Agents Chemother. 2007, 51, 2070–2077. [Google Scholar] [CrossRef] [Green Version]
- Dela Vega, A.L.; Delcour, A.H. Polyamines Decrease Escherichia coli Outer Membrane Permeability. J. Bacteriol. 1996, 178, 3715–3721. [Google Scholar] [CrossRef] [Green Version]
- Samartzidou, H.; Delcour, A.H. Excretion of Endogenous Cadaverine Leads to a Decrease in Porin-Mediated Outer Membrane Permeability. J. Bacteriol. 1999, 181, 791–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magoulas, G.E.; Kostopoulou, O.N.; Garnelis, T.; Athanassopoulos, C.M.; Kournoutou, G.G.; Leotsinidis, M.; Dinos, G.P.; Papaioannou, D.; Kalpaxis, D.L. Synthesis and Antimicrobial Activity of Chloramphenicol-Polyamine Conjugates. Bioorg. Med. Chem. 2015, 23, 3163–3174. [Google Scholar] [CrossRef]
- Blanchet, M.; Borselli, D.; Brunel, J.M. Polyamine Derivatives: A Revival of an Old Neglected Scaffold to Fight Resistant Gram-Negative Bacteria? Future Med. Chem. 2016, 8, 963–973. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Chopra, I. Leakage of Periplasmic Proteins from Escherichia coli Mediated by Polymyxin B Nonapeptide. Antimicrob. Agents Chemother. 1986, 29, 781–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mogana, R.; Adhikari, A.; Tzar, M.N.; Ramliza, R.; Wiart, C. Antibacterial Activities of the Extracts, Fractions and Isolated Compounds from Canarium patentinervium Miq. against Bacterial Clinical Isolates. BMC Complement Med. Ther. 2020, 20, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, Y.F.; Abou-Shleib, H.M.; Khalil, A.M.; El-Guink, N.M.; El-Nakeeb, M.A. Membrane Permeabilization of Colistin toward Pan-Drug Resistant Gram-Negative Isolates. Braz. J. Microbiol. 2016, 47, 381–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayaud, L.; Carricajo, A.; Zhiri, A.; Aubert, G. Comparison of Bacteriostatic and Bactericidal Activity of 13 Essential Oils against Strains with Varying Sensitivity to Antibiotics. Lett. Appl. Microbiol. 2008, 47, 167–173. [Google Scholar] [CrossRef]
- Rhouma, M.; Beaudry, F.; Thériault, W.; Bergeron, N.; Laurent-Lewandowski, S.; Fairbrother, J.M.; Letellier, A. Gastric Stability and Oral Bioavailability of Colistin Sulfate in Pigs Challenged or Not with Escherichia coli O149: F4 (K88). Res. Vet. Sci. 2015, 102, 173–181. [Google Scholar] [CrossRef]
- Li, J.; Nation, R.L.; Milne, R.W.; Turnidge, J.D.; Coulthard, K. Evaluation of Colistin as an Agent against Multi-Resistant Gram-Negative Bacteria. Int. J. Antimicrob. Agents 2005, 25, 11–25. [Google Scholar] [CrossRef]
- Frasca, D.; Dahyot-fizelier, C.; Mimoz, O. La Colistine En Réanimation Colistin in the Intensive Care Unit. Réanimation 2008, 17, 251–258. [Google Scholar] [CrossRef]
- Naghmouchi, K.; Baah, J.; Hober, D.; Jouy, E.; Rubrecht, C.; Sané, F.; Drider, D. Synergistic Effect between Colistin and Bacteriocins in Controlling Gram-Negative Pathogens and Their Potential to Reduce Antibiotic Toxicity in Mammalian Epithelial Cells. Antimicrob. Agents Chemother. 2013, 57, 2719–2725. [Google Scholar] [CrossRef] [Green Version]
- Severino, P.; Silveira, E.F.; Loureiro, K.; Chaud, M.V.; Antonini, D.; Lancellotti, M.; Sarmento, V.H.; da Silva, C.F.; Santana, M.H.A.; Souto, E.B. Antimicrobial Activity of Polymyxin-Loaded Solid Lipid Nanoparticles (PLX-SLN): Characterization of Physicochemical Properties and In Vitro Efficacy. Eur. J. Pharm. Sci. 2017, 106, 177–184. [Google Scholar] [CrossRef]
- Biswaro, L.S.; da Costa Sousa, M.G.; Rezende, T.M.B.; Dias, S.C.; Franco, O.L. Antimicrobial Peptides and Nanotechnology, Recent Advances and Challenges. Front. Microbiol. 2018, 9, 855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoff Brait, D.R.; Mattos Vaz, M.S.; da Silva Arrigo, J.; Borges de Carvalho, L.N.; Souza de Araújo, F.H.; Vani, J.M.; da Silva Mota, J.; Cardoso, C.A.L.; Oliveira, R.J.; Negrão, F.J.; et al. Toxicological Analysis and Anti-Inflammatory Effects of Essential Oil from Piper Vicosanum Leaves. Regul. Toxicol. Pharmacol. 2015, 73, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Miller-Fleming, L.; Olin-Sandoval, V.; Campbell, K.; Ralser, M. Remaining Mysteries of Molecular Biology: The Role of Polyamines in the Cell. J. Mol. Biol. 2015, 427, 3389–3406. [Google Scholar] [CrossRef] [PubMed]
- Til, H.P.; Falke, H.E.; Prinsen, M.K.; Willems, M.I. Acute and Subacute Toxicity of Tyramine, Spermidine, Spermine, Putrescine and Cadaverine in Rats. Food Chem. Toxicol. 1997, 35, 337–348. [Google Scholar] [CrossRef]
- Zgheib, H.; Belguesmia, Y.; Boukherroub, R.; Drider, D. Alginate Nanoparticles Enhance Anti-Clostridium perfringens Activity of the Leaderless Two-Peptide Enterocin DD14 and Affect Expression of Some Virulence Factors. Probiotics Antimicrob. Prot. 2021, 13, 1213–1227. [Google Scholar] [CrossRef] [PubMed]
- Belguesmia, Y.; Hazime, N.; Kempf, I.; Boukherroub, R.; Drider, D. New Bacteriocins from Lacticaseibacillus paracasei CNCM I-5369 Adsorbed on Alginate Nanoparticles Are Very Active against Escherichia coli. Int. J. Mol. Sci. 2020, 21, E8654. [Google Scholar] [CrossRef] [PubMed]
- Andrews, J.M. Determination of Minimum Inhibitory Concentrations. J. Antimicrob. Chemother. 2001, 48, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Thornsberry, C. NCCLS Standards for Antimicrobial Susceptibility Tests. Lab. Med. 1983, 14, 549–553. [Google Scholar] [CrossRef]
- Grubb, J.A.; Dehority, B.A. Variation in Colony Counts of Total Viable Anaerobic Rumen Bacteria as Influenced by Media and Cultural Methods. Appl. Environ. Microbiol. 1976, 31, 262–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.-Q.; Xu, J.; Yin, Z.-Q.; Jia, R.-Y.; Lu, Y.; Yang, F.; Du, Y.-H.; Zou, P.; Lv, C.; Hu, T.-X.; et al. Isolation and Identification of the Antibacterial Active Compound from Petroleum Ether Extract of Neem Oil. Fitoterapia 2010, 81, 747–750. [Google Scholar] [CrossRef] [PubMed]
- Belguesmia, Y.; Madi, A.; Sperandio, D.; Merieau, A.; Feuilloley, M.; Prévost, H.; Drider, D.; Connil, N. Growing Insights into the Safety of Bacteriocins: The Case of Enterocin S37. Res. Microbiol. 2011, 162, 159–163. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Colistin (MIC [µg/mL]) | Alg NPs+Colistin (MIC [µg/mL]) |
|---|---|---|
| E. coli 184 | 8 | 1 |
| E. coli 289 | 8 | 4 |
| E. coli E4A4v | 16 | 4 |
| E. coli E5A16v | 16 | 4 |
| E. coli ATTC8739 | 2 | 1 |
| E. coli CIP 7624 | 1 | 1 |
| E. coli SBS363 | 2 | 1 |
| E. coli E4A4wt | 2 | 2 |
| E. coli E5A16wt | 2 | 2 |
| E. coli Top 10 | 1 | 0.5 |
| Formulations (µg/mL) | MIC (µg/mL) |
|---|---|
| Alg NPs+Menthol (500:10) | - |
| Alg NPs+Carvacrol (500:10) | - |
| Alg NPs+Farnesol (500:10) | - |
| Alg NPs+Geraniol (500:10) | - |
| Alg NPs+Colistin+Menthol (500:40:10) | 0.31 |
| Alg NPs+Colistin+Carvacrol (500:40:10) | ≤0.62 |
| Alg NPs+Colistin+Farnesol (500:40:10) | ≤1.25 |
| Alg NPs+Colistin+Geraniol (500:40:10) | ≤1.25 |
| Formulation (µg/mL) | MIC (µg/mL) |
|---|---|
| Lactic acid (15) | - |
| Alg NPs+Colistin (500:40) | 1.25 |
| Alg NPs+Colistin+Lactic acid (500:40:15) | ≤0.62 |
| Formulation (µg/mL) | MIC (µg/mL) |
|---|---|
| Colistin | 8 |
| ACS Alg NPs+Colistin+Spermine (500:60:20) | 5 |
| ACSd: Alg NPs+Colistin+Spermidine (500:60:20) | 2.5 |
| ACP: Alg NPs+Colistin+Piperazine (500:60:20) | 10 |
| ACSd1: Alg NPs+Colistin+Spermidine (500:30:30) | 3.75 |
| ACSd2: Alg NPs+Colistin+Spermidine (500:20:40) | 5 |
| ACSd3: Alg NPs+Colistin+Spermidine (500:10:50) | 5 |
| Formulation | MIC (µg/mL) | MBC (µg/mL) | MBC/MIC |
|---|---|---|---|
| Colistin | 8 | 8 | 1 |
| Alg NPs+Colistin (500:40) | 1.25 | 5 | 4 |
| Alg NPs+Colistin+Menthol (500:40:10) | 0.31 | 0.62 | 2 |
| Alg NPs+Colistin+Lactic acid (500:40:15) | ≤0.62 | 1.25 | 2 |
| Formulation | Untreated | Incubation at pH 3 for 30 min and then at pH 6 for 2 h | Incubation at pH 3 for 30 min with Pepsin * and then at pH 6 for 2 h | Incubation at pH 3 for 30 min with Pepsin * and then at pH 6 for 2 h with Trypsin+Chymotrypsin * |
|---|---|---|---|---|
| Colistin (60 µg/mL) | 8 | 8 | 8 | ≥32 |
| Alg NPs+Colistin (500:40) µg/mL) | 1.25 | 1.25 | 5 | 5 |
| Alg NPs+Colistin+Lactic acid (500:40:10 µg/mL) | 0.62 | 1.25 | 2.5 | 2.5 |
| Alg NPs+Colistin+Menthol (500:40:10) µg/mL | 0.31 | 1.25 | 1.25 | 2.5 |
| Strains | Origin | Characteristics |
|---|---|---|
| E. coli 184 | Résapath network *, | Colistin R, (mcr-1 gene) |
| E. coli 289 | ANSES ** collection, | Colistin R, (mcr-4 gene) |
| E. coli E4A4wt | ANSES collection | Colistin S |
| E. coli E5A16wt | ANSES collection | Colistin S |
| E. coli ATTC8739 | ATCC collection | Colistin S, reference strain |
| E. coli CIP 7624 | Pasteur Institute collection (Paris, France) | Colistin S, reference strain |
| E. coli SBS363 | Pasteur Institute collection (Paris, France) | Strain with truncated LPS |
| E. coli E4A4v | ANSES collection | In vitro resistant variant to colistin |
| E. coli E5A16v | ANSES collection | In vitro resistant variant to colistin |
| E. coli Top 10 | Invitrogen ® | Modified strain (LPS-) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hazime, N.; Belguesmia, Y.; Kempf, I.; Barras, A.; Drider, D.; Boukherroub, R. Enhancing Colistin Activity against Colistin-Resistant Escherichia coli through Combination with Alginate Nanoparticles and Small Molecules. Pharmaceuticals 2022, 15, 682. https://doi.org/10.3390/ph15060682
Hazime N, Belguesmia Y, Kempf I, Barras A, Drider D, Boukherroub R. Enhancing Colistin Activity against Colistin-Resistant Escherichia coli through Combination with Alginate Nanoparticles and Small Molecules. Pharmaceuticals. 2022; 15(6):682. https://doi.org/10.3390/ph15060682
Chicago/Turabian StyleHazime, Noura, Yanath Belguesmia, Isabelle Kempf, Alexandre Barras, Djamel Drider, and Rabah Boukherroub. 2022. "Enhancing Colistin Activity against Colistin-Resistant Escherichia coli through Combination with Alginate Nanoparticles and Small Molecules" Pharmaceuticals 15, no. 6: 682. https://doi.org/10.3390/ph15060682
APA StyleHazime, N., Belguesmia, Y., Kempf, I., Barras, A., Drider, D., & Boukherroub, R. (2022). Enhancing Colistin Activity against Colistin-Resistant Escherichia coli through Combination with Alginate Nanoparticles and Small Molecules. Pharmaceuticals, 15(6), 682. https://doi.org/10.3390/ph15060682