
Quercetin-Rich Extracts from Onions (Allium cepa) Play Potent Cytotoxicity on Adrenocortical Carcinoma Cell Lines, and Quercetin Induces Important Anticancer Properties

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
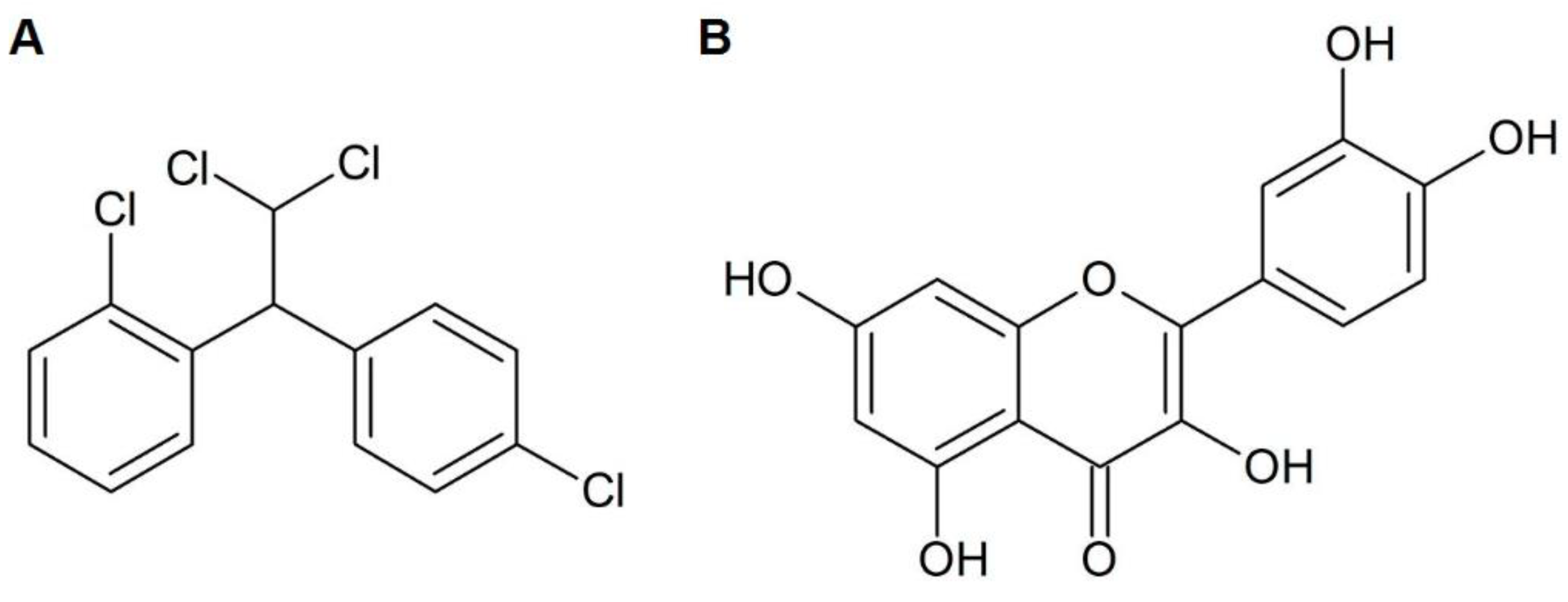
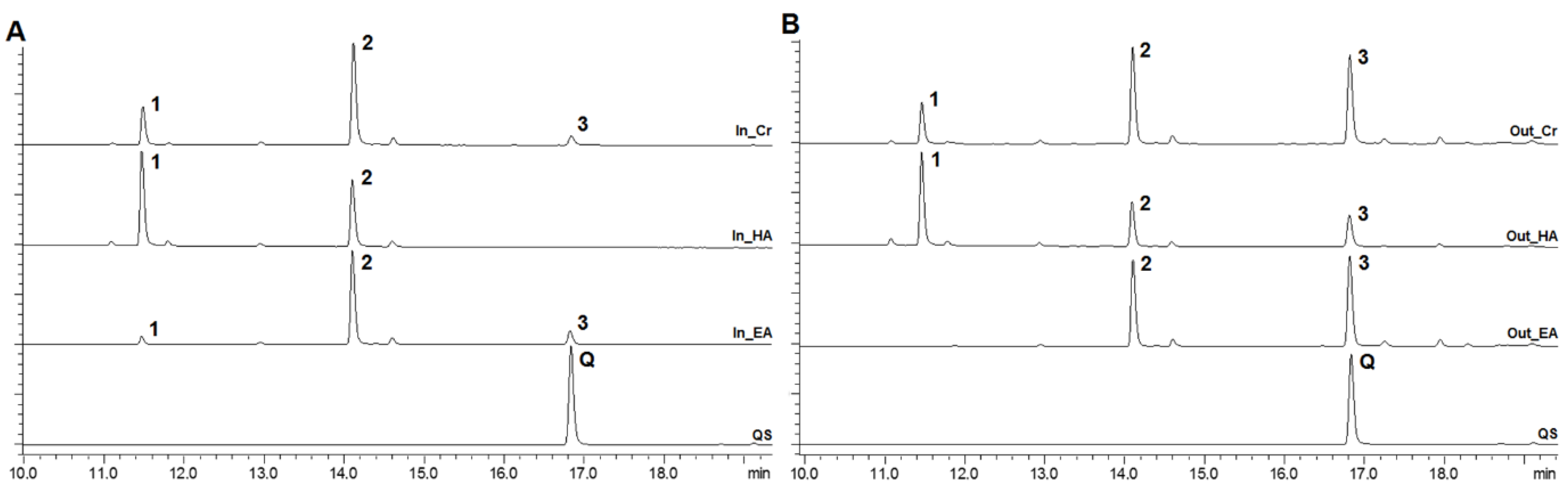
2.1. Composition Analysis of Allium cepa Extracts and Fractions
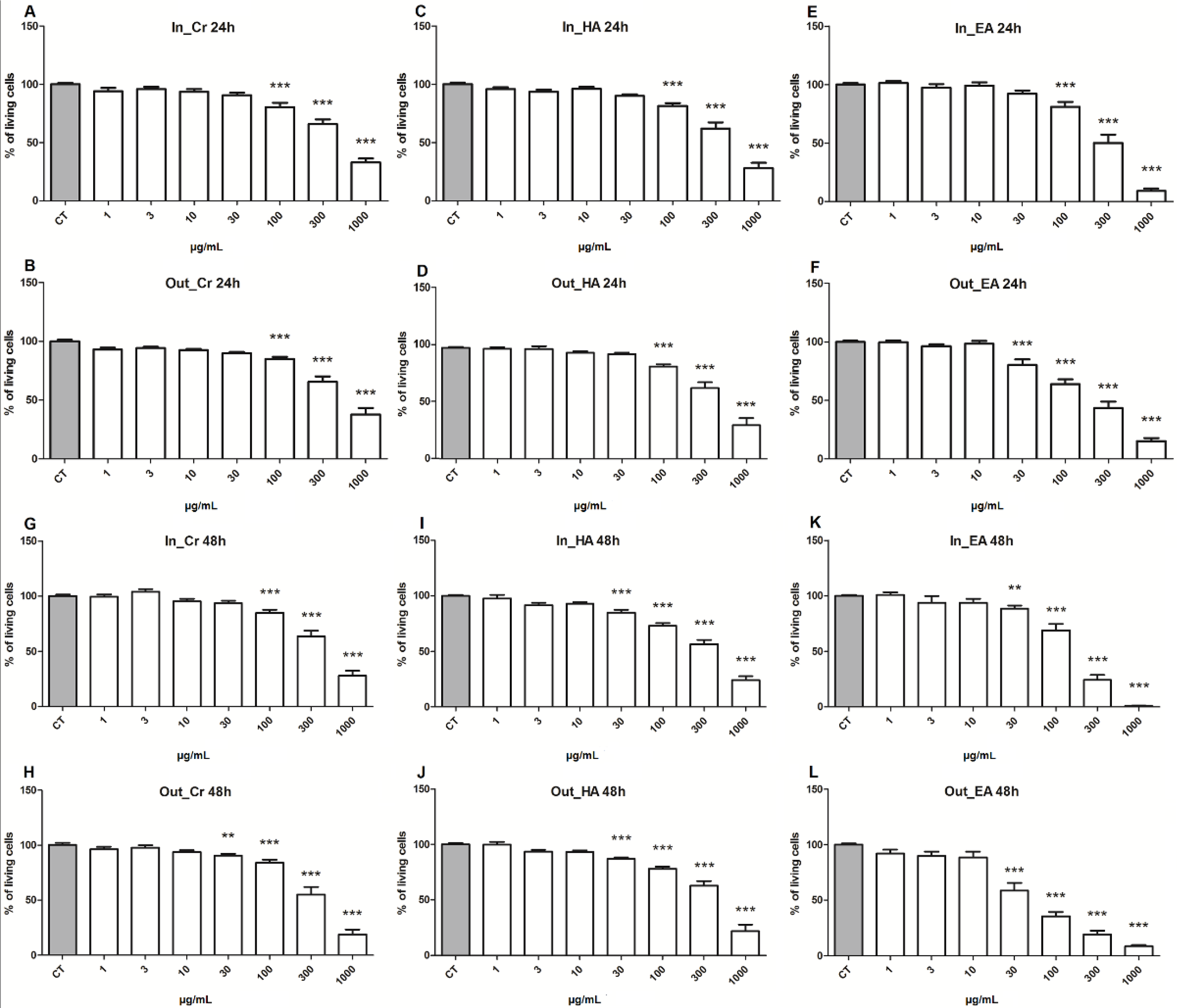
2.2. Cytotoxicity on Adrenocortical Carcinoma Cell Line
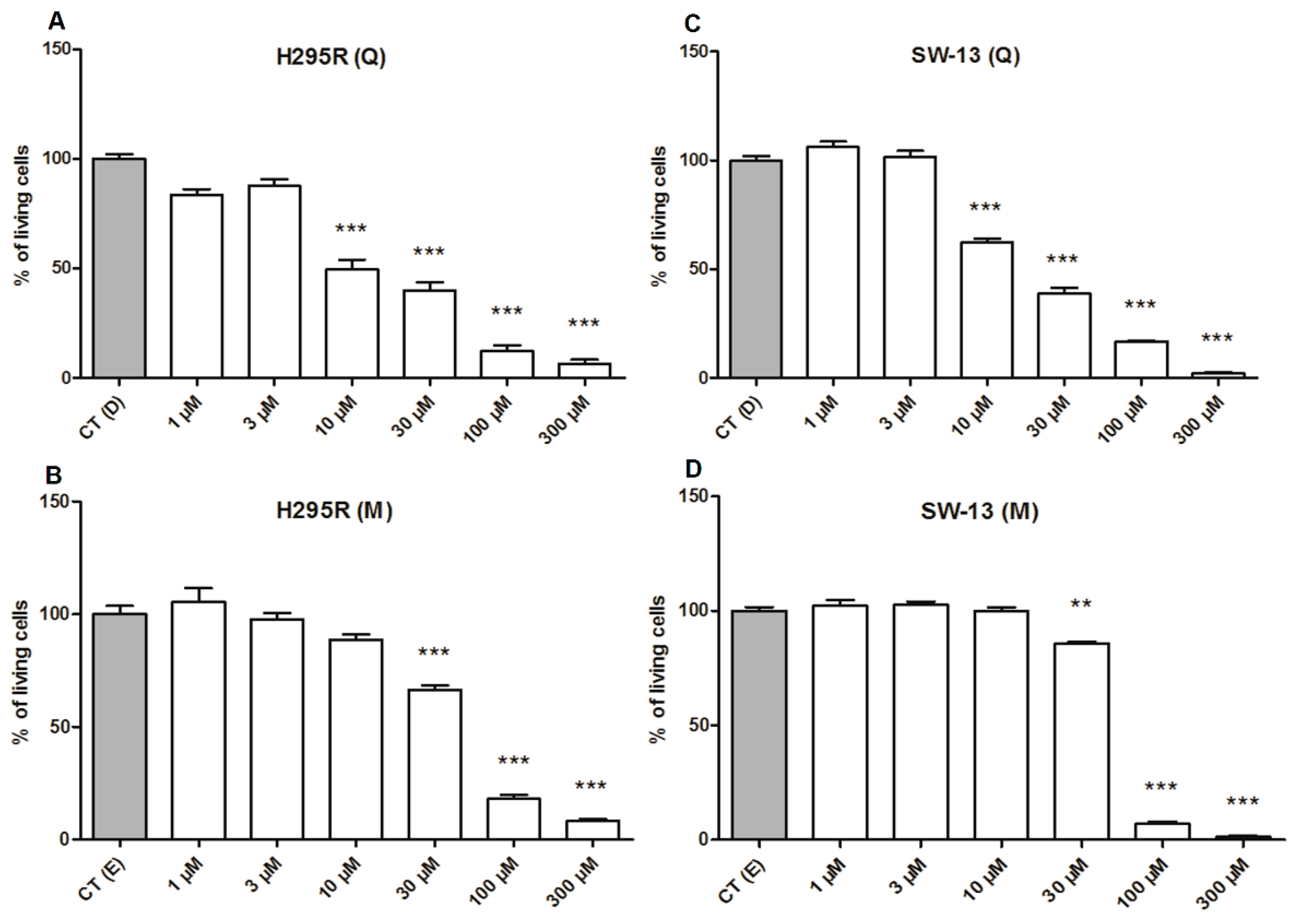
2.3. Quercetin Plays Higher Cytotoxicity Than Mitotane
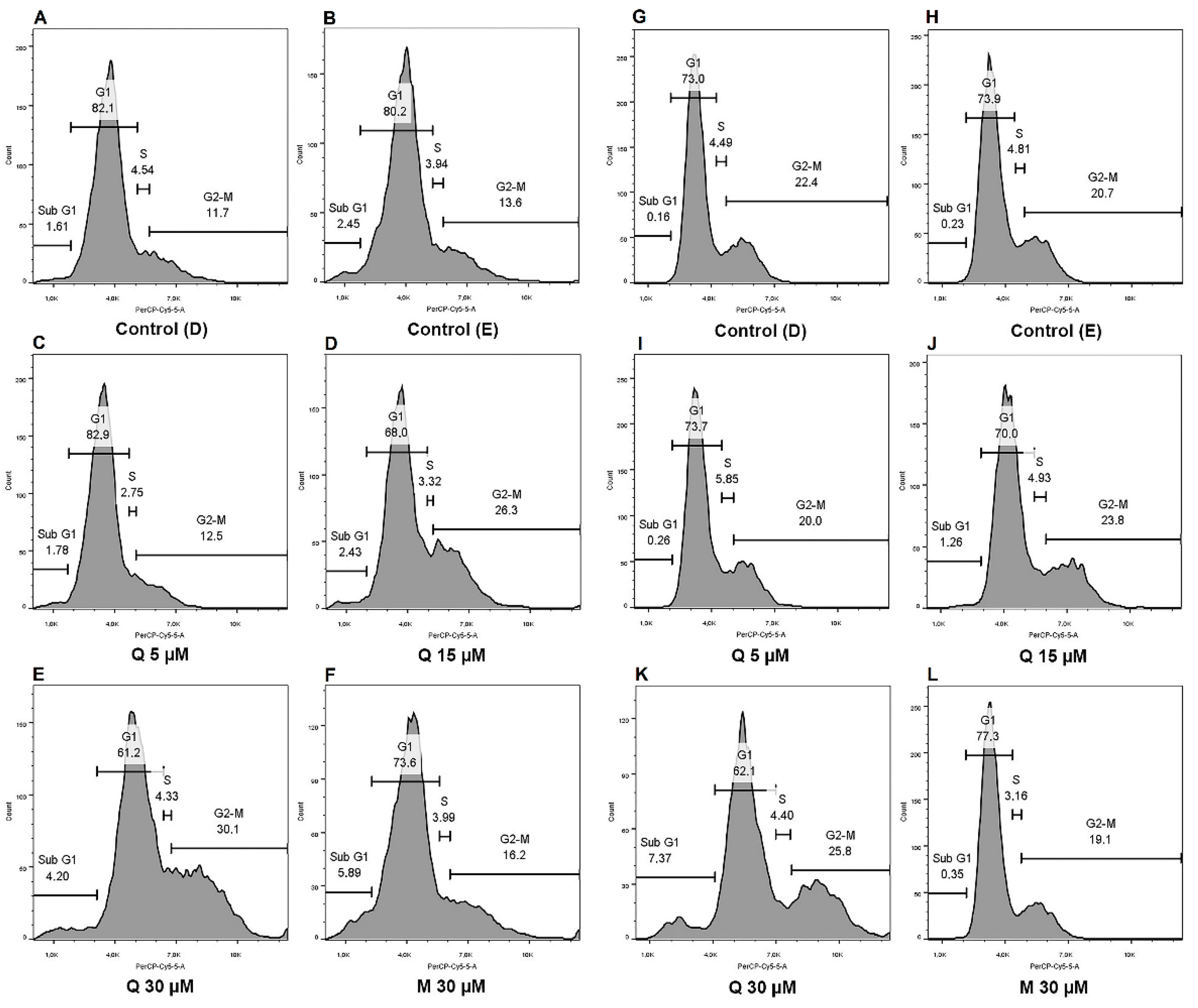
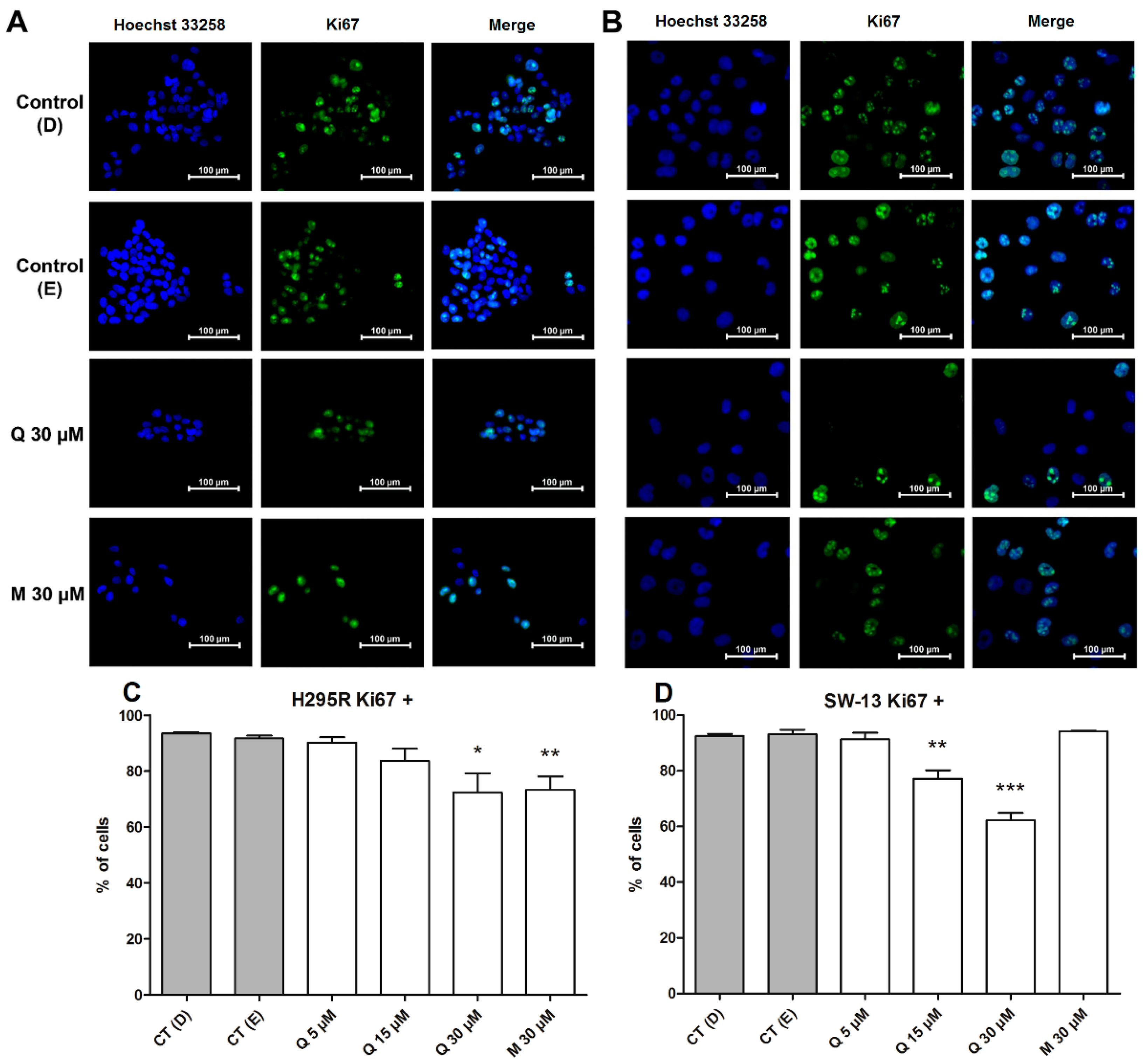
2.4. Cell Cycle Dynamics and Proliferation Rates Show Different Distributions between Cells and Treatments
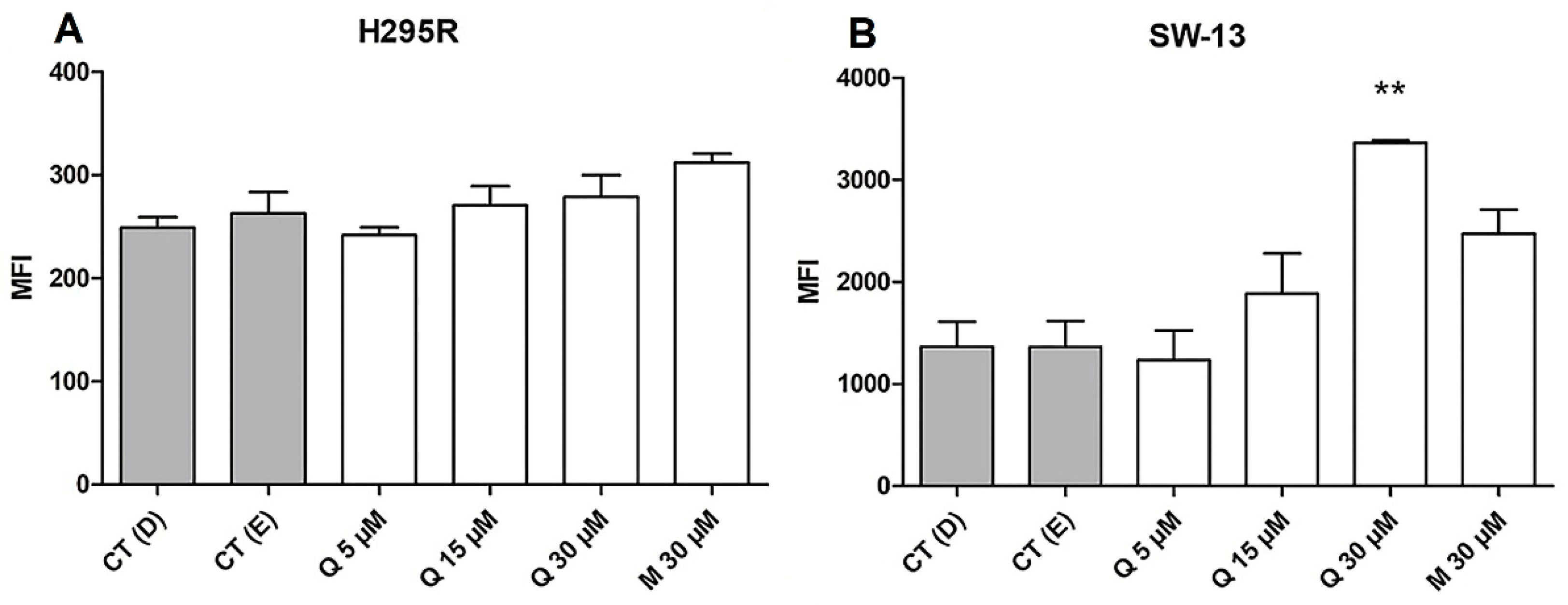
2.5. Quercetin Induced the Early and Late Apoptosis Rates and Increases Free Ca2+ Release on SW-13 Cells
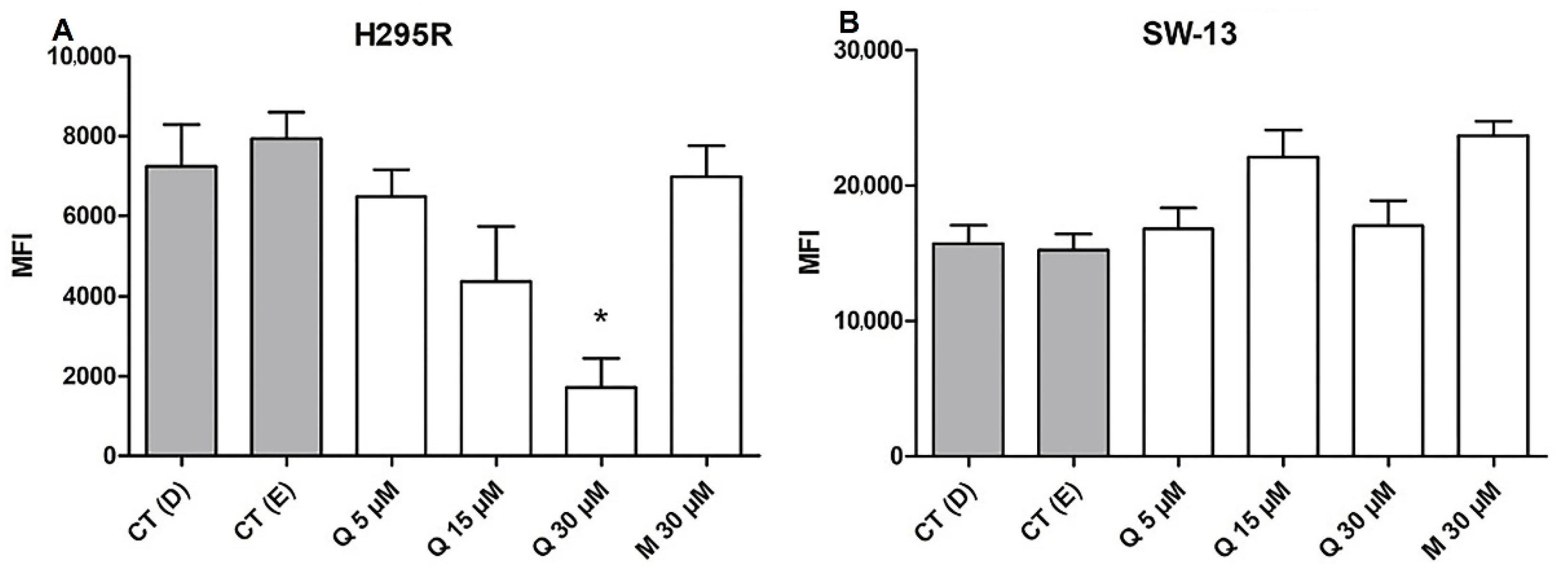
2.6. Quercetin Decreases ROS Generation in the H295R Cells
3. Materials and Methods
3.1. Cell Culture and Reagents
3.2. Extraction and Fractionation of A. cepa Components
3.3. Chromatographic Analysis
3.4. Cytotoxicity Assays
3.5. Cell Cycle and Proliferation
3.6. Apoptosis and Necrosis Rate
3.7. Intracellular Calcium Rate
3.8. Reactive Oxygen Species (ROS) Measurement
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Allolio, B.; Fassnacht, M. Clinical review: Adrenocortical carcinoma: Clinical update. J. Clin. Endocrinol. Metab. 2006, 91, 2027–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, T.E.J.; Gerber, V.K.Q.; Ibañez, H.C.; Melanda, V.S.; Parise, I.Z.S.; Watanabe, F.M.; Pianovski, M.A.D.; Fiori, C.M.C.M.; Fabro, A.L.M.R.; da Silva, D.B.; et al. Penetrance of the TP53 R337H Mutation and Pediatric Adrenocortical Carcinoma Incidence Associated with Environmental Influences in a 12-Year Observational Cohort in Southern Brazil. Cancers 2019, 11, 1804. [Google Scholar] [CrossRef] [Green Version]
- Corso, C.R.; Acco, A.; Bach, C.; Bonatto, S.J.R.; de Figueiredo, B.C.; de Souza, L.M. Pharmacological Profile and Effects of Mitotane in Adrenocortical Carcinoma. Br. J. Clin. Pharmacol. 2021, 87, 2698–2710. [Google Scholar] [CrossRef]
- Woiski, T.D.; Poncio, L.D.C.; De Moura, J.; Orsato, A.; Bezerra, A.G.; Minozzo, J.C.; De Figueiredo, B.C. Anti-HMC2RL1 Functionalized Gold Nanoparticles for Adrenocortical Tumor Cells Targeting and Imaging. J. Biomed. Nanotechnol. 2017, 13, 68–76. [Google Scholar] [CrossRef]
- Patra, S.; Pradhan, B.; Nayak, R.; Behera, C.; Panda, K.C.; Das, S.; Jena, M.; Bhutia, S.K. Apoptosis and Autophagy Modulating Dietary Phytochemicals in Cancer Therapeutics: Current Evidences and Future Perspectives. Phyther. Res. 2021, 35, 4194–4214. [Google Scholar] [CrossRef]
- Zhu, L.; Chen, L. Progress in Research on Paclitaxel and Tumor Immunotherapy. Cell. Mol. Biol. Lett. 2019, 24, 40. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.K.; Xu, J.K.; Tian, H.Y.; Wang, L.; Zhang, X.Q.; Xiao, X.Z.; Li, P.; Ye, W.C. Two New Vinblastine-Type N -Oxide Alkaloids from Catharanthus Roseus. Nat. Prod. Res. 2013, 27, 1911–1916. [Google Scholar] [CrossRef]
- Al-Mahayri, Z.N.; AlAhmad, M.M.; Ali, B.R. Current Opinion on the Pharmacogenomics of Paclitaxel-Induced Toxicity. Expert Opin. Drug Metab. Toxicol. 2021, 17, 785–801. [Google Scholar] [CrossRef]
- González-de-Peredo, A.V.; Vázquez-Espinosa, M.; Carrera, C.; Espada-Bellido, E.; Ferreiro-González, M.; Barbero, G.F.; Palma, M. Development of a Rapid UHPLC-PDA Method for the Simultaneous Quantification of Flavonol Contents in Onions (Allium cepa L.). Pharmaceuticals 2021, 14, 310. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Evolution of Secondary Metabolites from an Ecological and Molecular Phylogenetic Perspective. Phytochemistry 2003, 64, 3–19. [Google Scholar] [CrossRef]
- Srivastava, S.; Somasagara, R.R.; Hegde, M.; Nishana, M.; Tadi, S.K.; Srivastava, M.; Choudhary, B.; Raghavan, S.C. Quercetin, a Natural Flavonoid Interacts with DNA, Arrests Cell Cycle and Causes Tumor Regression by Activating Mitochondrial Pathway of Apoptosis. Sci. Rep. 2016, 6, 24049. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.H.; Lai, S.L.; Chen, W.S.; Hung, W.Y.; Chow, J.M.; Hsiao, M.; Lee, W.J.; Chien, M.H. Quercetin Suppresses the Metastatic Ability of Lung Cancer through Inhibiting Snail-Dependent Akt Activation and Snail-Independent ADAM9 Expression Pathways. Biochim. Biophys. Acta-Mol. Cell Res. 2017, 1864, 1746–1758. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.B.; Mir, H.; Kapur, N.; Gales, D.N.; Carriere, P.P.; Singh, S. Quercetin Inhibits Prostate Cancer by Attenuating Cell Survival and Inhibiting Anti-Apoptotic Pathways. World J. Surg. Oncol. 2018, 16, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Needs, P.W.; Lu, Y.; Kroon, P.A.; Ren, D.; Yang, X. Different Antitumor Effects of Quercetin, Quercetin-3′-Sulfate and Quercetin-3-Glucuronide in Human Breast Cancer MCF-7 Cells. Food Funct. 2018, 9, 1736–1746. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, M.; Kannan, A.; Stojković, D.S.; Glamočlija, J.; Calhelha, R.C.; Ferreira, I.C.F.R.; Sanglard, D.; Soković, M. Flavones, Flavonols, and Glycosylated Derivatives—Impact on Candida Albicans Growth and Virulence, Expression of Cdr1 and Erg11, Cytotoxicity. Pharmaceuticals 2021, 14, 27. [Google Scholar] [CrossRef]
- Grzelak-Błaszczyk, K.; Milala, J.; Kołodziejczyk, K.; Sójka, M.; Czarnecki, A.; Kosmala, M.; Klewicki, R.; Fotschki, B.; Jurgoński, A.; Juśkiewicz, J. Protocatechuic Acid and Quercetin Glucosides in Onions Attenuate Changes Induced by High Fat Diet in Rats. Food Funct. 2020, 11, 3585–3597. [Google Scholar] [CrossRef]
- Bouba, A.A.; Njintang, N.Y.; Nkouam, G.B.; Mang, Y.D.; El-Sayed Mehanni, A.H.; Scher, J.; Montet, D.; Mbofung, C.M. Desorption Isotherms, Net Isosteric Heat and the Effect of Temperature and Water Activity on the Antioxidant Activity of Two Varieties of Onion (Allium cepa L). Int. J. Food Sci. Technol. 2014, 49, 444–452. [Google Scholar] [CrossRef]
- Chakraborty, A.J.; Uddin, T.M.; Matin Zidan, B.M.R.; Mitra, S.; Das, R.; Nainu, F.; Dhama, K.; Roy, A.; Hossain, M.J.; Khusro, A.; et al. Allium cepa: A Treasure of Bioactive Phytochemicals with Prospective Health Benefits. Evid.-Based Compl. Altern. Med. 2022, 2022, 4586318. [Google Scholar] [CrossRef]
- Fredotovíc, Ž.; Šprung, M.; Soldo, B.; Ljubenkov, I.; Budić-Leto, I.; Bilušić, T.; Cikeš-Čulić, V.; Puizina, J. Chemical Composition and Biological Activity of Allium cepa L. and Allium × Cornutum (Clementi Ex Visiani 1842) Methanolic Extracts. Molecules 2017, 22, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cozzolino, R.; Malorni, L.; Martignetti, A.; Picariello, G.; Siano, F.; Forte, G.; De Giulio, B. Comparative Analysis of Volatile Profiles and Phenolic Compounds of Four Southern Italian Onion (Allium cepa L.) Landraces. J. Food Compos. Anal. 2021, 101, 103990. [Google Scholar] [CrossRef]
- Marrelli, M.; Russo, C.; Statti, G.; Argentieri, M.P.; Meleleo, D.; Mallamaci, R.; Avato, P.; Conforti, F. Phytochemical and Biological Characterization of Dry Outer Scales Extract from Tropea Red Onion (Allium cepa L. Var. Tropea)–A Promising Inhibitor of Pancreatic Lipase. Phytomedicine Plus 2022, 2, 100235. [Google Scholar] [CrossRef]
- Kwak, J.H.; Seo, J.M.; Kim, N.H.; Arasu, M.V.; Kim, S.; Yoon, M.K.; Kim, S.J. Variation of Quercetin Glycoside Derivatives in Three Onion (Allium cepa L. ) Varieties. Saudi J. Biol. Sci. 2017, 24, 1387–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gattuso, G.; Barreca, D.; Gargiulli, C.; Leuzzi, U.; Caristi, C. Flavonoid Composition of Citrus Juices. Molecules 2007, 12, 1641–1673. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Wu, B.; Tang, L.; Liu, Z.; Hu, M. Identification of the Position of Mono- O-Glucuronide of Flavones and Flavonols by Analyzing Shift in Online UV Spectrum (Λmax) Generated from an Online Diode Array Detector. J. Agric. Food Chem. 2010, 58, 9384–9395. [Google Scholar] [CrossRef] [Green Version]
- Moolenaar, A.J.; Van Slooten, H.; Van Seters, A.P.; Smenks, D. Blood Levels of o,p′-DDD Following Administration in Various Vehicles After a Single Dose and During Long-term Treatment. Cancer Chemother. Pharm. 1981, 7, 51–54. [Google Scholar] [CrossRef]
- Baudin, E.; Pellegriti, G.; Bonnay, M.; Penfornis, A.; Laplanche, A.; Vassal, G.; Schlumberger, M. Impact of Monitoring Plasma 1,1-Dichlorodiphenildichloroethane (o, P′DDD) Levels on the Treatment of Patients with Adrenocortical Carcinoma. Cancer 2001, 92, 1385–1392. [Google Scholar] [CrossRef]
- Ferry, D.R.; Smith, A.; Malkhandi, J.; Fyfe, D.W.; DeTakats, P.G.; Anderson, D.; Baker, J.; Kerr, D.J. Phase I Clinical Trial of the Flavonoid Quercetin: Pharmacokinetics and Evidence for in Vivo Tyrosine Kinase Inhibition. Clin. Cancer Res. 1996, 2, 659–668. [Google Scholar]
- Reed, S.I. Control of the G1/S transition. Cancer Surv. 1997, 29, 7–23. [Google Scholar]
- Lee, T.J.; Kim, O.H.; Kim, Y.H.; Lim, J.H.; Kim, S.; Park, J.W.; Kwon, T.K. Quercetin Arrests G2/M Phase and Induces Caspase-Dependent Cell Death in U937 Cells. Cancer Lett. 2006, 240, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Sobecki, M.; Mrouj, K.; Colinge, J.; Gerbe, F.; Jay, P.; Krasinska, L.; Dulic, V.; Fisher, D. Cell-Cycle Regulation Accounts for Variability in Ki-67 Expression Levels. Cancer Res. 2017, 77, 2722–2734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daker, M.; Ahmad, M.; Khoo, A.S.B. Quercetin-Induced Inhibition and Synergistic Activity with Cisplatin—A Chemotherapeutic Strategy for Nasopharyngeal Carcinoma Cells. Cancer Cell Int. 2012, 12, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingham, M.; Schwartz, G.K. Cell-Cycle Therapeutics Come of Age. J. Clin. Oncol. 2017, 35, 2949–2959. [Google Scholar] [CrossRef]
- Williams, G.H.; Stoeber, K. The Cell Cycle and Cancer. J. Pathol. 2012, 226, 352–364. [Google Scholar] [CrossRef]
- Choi, E.J.; Bae, S.M.; Ahn, W.S. Antiproliferative Effects of Quercetin through Cell Cycle Arrest and Apoptosis in Human Breast Cancer MDA-MB-453 Cells. Arch. Pharmacal Res. 2008, 31, 1281–1285. [Google Scholar] [CrossRef]
- Hisaka, T.; Sakai, H.; Sato, T.; Goto, Y.; Nomura, Y.; Fukutomi, S.; Fujita, F.; Mizobe, T.; Nakashima, O.; Tanigawa, M.; et al. Quercetin Suppresses Proliferation of Liver Cancer Cell Lines in Vitro. Anticancer Res. 2020, 40, 4695–4700. [Google Scholar] [CrossRef]
- Soll, F.; Ternent, C.; Berry, I.M.; Kumari, D.; Moore, T.C. Quercetin Inhibits Proliferation and Induces Apoptosis of B16 Melanoma Cells in Vitro. Assay Drug Dev. Technol. 2020, 18, 261–268. [Google Scholar] [CrossRef]
- Lee, W.J.; Hsiao, M.; Chang, J.L.; Yang, S.F.; Tseng, T.H.; Cheng, C.W.; Chow, J.M.; Lin, K.H.; Lin, Y.W.; Liu, C.C.; et al. Quercetin Induces Mitochondrial-Derived Apoptosis via Reactive Oxygen Species-Mediated ERK Activation in HL-60 Leukemia Cells and Xenograft. Arch. Toxicol. 2015, 89, 1103–1117. [Google Scholar] [CrossRef]
- Chan, L.Y.; Yim, E.K.F.; Choo, A.B.H. Normalized Median Fluorescence: An Alternative Flow Cytometry Analysis Method for Tracking Human Embryonic Stem Cell States During Differentiation. Tissue Eng. Part C-Meth. 2012, 19, 156–165. [Google Scholar] [CrossRef]
- Mattson, M.P.; Chan, S.L. Calcium Orchestrates Apoptosis. Nat. Cell Biol. 2003, 5, 1041–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salama, Y.A.; El-karef, A.; El Gayyar, A.M.; Abdel-Rahman, N. Beyond Its Antioxidant Properties: Quercetin Targets Multiple Signalling Pathways in Hepatocellular Carcinoma in Rats. Life Sci. 2019, 236, 116933. [Google Scholar] [CrossRef] [PubMed]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of Oxidative Stress as an Anticancer Strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Huang, K.; Gao, C.; Lau, Q.C.; Pan, H.; Xie, K.; Li, J.; Liu, R.; Zhang, T.; Xie, N.; et al. Proteomics Identification of ITGB3 as a Key Regulator in Reactive Oxygen Species-Induced Migration and Invasion of Colorectal Cancer Cells. Mol. Cell. Proteom. 2011, 10, M110.005397. [Google Scholar] [CrossRef] [Green Version]
- Gazdar, A.F.; Oie, H.K.; Shackleton, C.H.; Chen, T.R.; Triche, T.J.; Myers, C.E.; Chrousos, G.P.; Brennan, M.F.; Stein, C.A.; Rocca, R.V.L. Establishment and Characterization of a Human Adrenocortical Carcinoma Cell Line That Expresses Multiple Pathways of Steroid Biosynthesis. Cancer Res. 1990, 50, 5488–5496. [Google Scholar]
- Feoktistova, M.; Geserick, P.; Leverkus, M. Crystal Violet Assay for Determining Viability of Cultured Cells. Cold Spring Harb. Protoc. 2016, 4, 343–346. [Google Scholar] [CrossRef]
- Bailey, S.; Macardle, P.J. A Flow Cytometric Comparison of Indo-1 to Fluo-3 and Fura Red Excited with Low Power Lasers for Detecting Ca2+ Flux. J. Immunol. Meth. 2006, 311, 220–225. [Google Scholar] [CrossRef]
- Drehmer, D.L.; de Aguiar, A.M.; Brandt, A.P.; Petiz, L.; Cadena, S.M.S.C.; Rebelatto, C.K.; Brofman, P.R.S.; Filipak Neto, F.; Dallagiovanna, B.; Abud, A.P.R. Metabolic Switches during the First Steps of Adipogenic Stem Cells Differentiation. Stem Cell Res. 2016, 17, 413–421. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracts | Allium cepa Wet-Weight (g) | Allium cepa Dry-Weight (g) | Total Fraction Weight (g) | Extraction Yeld (%) (*) | Quercetin (µg/mg) |
|---|---|---|---|---|---|
| In_Cr | 178.3 | 34.4 | 3.9 | 11.3 | 0.241 ± 0.002 |
| In_Et | 3.4 | 9.9 | - | ||
| In_EA | 0.5 | 1.45 | 6.16 ± 0.013 | ||
| Out_CR | 159.3 | 36.8 | 1.3 | 3.5 | 7.815 ± 0.136 |
| Out_Et | 1.1 | 2.9 | 1.415 ± 0.011 | ||
| Out_EA | 0.2 | 0.5 | 108.85 ± 1.130 |
| G0/Sub G1 | G1 | S | G2/M | ||
|---|---|---|---|---|---|
| H295R | CT (D) | 1.71 ± 0.44 | 80.23 ± 2.27 | 4.95 ± 0.82 | 12.93 ± 1.04 |
| CT(E) | 1.92 ± 0.57 | 79.93 ± 1.27 | 3.52 ± 0.39 | 14.63 ± 1.08 | |
| Q 5 µM | 1.60 ± 0.64 | 78.47 ± 5.62 | 4.03 ± 0.73 | 15.93 ± 4.57 | |
| Q 15 µM | 2.02 ± 0.57 | 71.97 ± 4.16 | 3.95 ± 0.86 | 21.83 ± 4.12 | |
| Q 30 µM | 3.44 ± 0.45 | 65.40 ± 2.19 (**) | 5.43 ± 0.79 | 25.53 ± 2.78 (*) | |
| M 30 µM | 3.59 ± 1.17 | 74.57 ± 2.16 | 4.21 ± 0.28 | 17.30 ± 2.29 | |
| SW-13 | CT (D) | 0.26 ± 0.06 | 72.93 ± 0.34 | 4.80 ± 0.22 | 22.13 ± 0.59 |
| CT (E) | 0.28 ± 0.02 | 74.37 ± 1.57 | 4.80 ± 0.50 | 20.37 ± 2.02 | |
| Q 5 µM | 0.24 ± 0.03 | 73.77 ± 0.23 | 5.13 ± 0.51 | 20.77 ± 0.41 | |
| Q 15 µM | 1.78 ± 0.26 (**) | 69.73 ± 0.26 (**) | 3.60 ± 0.75 | 24.90 ± 0.56 | |
| Q 30 µM | 7.52 ± 0.65 (***) | 64.93 ± 1.41 (**) | 4.89 ± 0.90 | 22.50 ± 1.66 | |
| M 30 µM | 0.43 ± 0.08 | 77.33 ± 1.01 | 4.23 ± 0.64 | 18.03 ± 1.48 |
| Necrosis | Late Apoptosis | Early Apoptosis | Live Cells | ||
|---|---|---|---|---|---|
| H295R | CT (D) | 0.27 ± 0.21 | 1.15 ± 0.85 | 2.83 ± 0.71 | 95.77 ± 0.99 |
| CT(E) | 0.28 ± 0.23 | 1.22 ± 0.77 | 2.56 ± 1.45 | 95.93 ± 1.39 | |
| Q 5 µM | 0.26 ± 0.17 | 0.72 ± 0.38 | 1.93 ± 1.01 | 97.07 ± 0.75 | |
| Q 15 µM | 0.31 ± 0.25 | 1.89 ± 1.31 | 4.63 ± 2.59 | 93.17 ± 2.41 | |
| Q 30 µM | 0.39 ± 0.36 | 3.13 ± 2.39 | 5.71 ± 1.82 | 90.77 ± 3.01 | |
| M 30 µM | 0.31 ± 0.27 | 1.64 ± 1.19 | 6.58 ± 2.72 | 91.43 ± 2.62 | |
| SW-13 | CT (D) | 0.62 ± 0.12 | 0.64 ± 0.11 | 2.54 ± 0.66 | 96.16 ± 0.69 |
| CT (E) | 0.61 ± 0.11 | 0.60 ± 0.17 | 3.51 ± 0.88 | 95.26 ± 0.64 | |
| Q 5 µM | 0.59 ± 0.11 | 0.67 ± 0.11 | 3.63 ± 0.76 | 95.06 ± 0.73 | |
| Q 15 µM | 1.32 ± 1.02 | 2.46 ± 0.22 (**) | 44.30 ± 5.20 (**) | 64.01 ± 11.44 (*) | |
| Q 30 µM | 4.63 ± 4.28 | 15.06 ± 3.75 (*) | 71.80 ± 4,80 (***) | 26.46 ± 10.46 (**) | |
| M 30 µM | 0.89 ± 0.20 | 1.43 ± 0.04 | 5.71 ± 1.57 | 91.93 ± 1.50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veiga, A.A.; Irioda, A.C.; Mogharbel, B.F.; Bonatto, S.J.R.; Souza, L.M. Quercetin-Rich Extracts from Onions (Allium cepa) Play Potent Cytotoxicity on Adrenocortical Carcinoma Cell Lines, and Quercetin Induces Important Anticancer Properties. Pharmaceuticals 2022, 15, 754. https://doi.org/10.3390/ph15060754
Veiga AA, Irioda AC, Mogharbel BF, Bonatto SJR, Souza LM. Quercetin-Rich Extracts from Onions (Allium cepa) Play Potent Cytotoxicity on Adrenocortical Carcinoma Cell Lines, and Quercetin Induces Important Anticancer Properties. Pharmaceuticals. 2022; 15(6):754. https://doi.org/10.3390/ph15060754
Chicago/Turabian StyleVeiga, Alan A., Ana Carolina Irioda, Bassam F. Mogharbel, Sandro J. R. Bonatto, and Lauro M. Souza. 2022. "Quercetin-Rich Extracts from Onions (Allium cepa) Play Potent Cytotoxicity on Adrenocortical Carcinoma Cell Lines, and Quercetin Induces Important Anticancer Properties" Pharmaceuticals 15, no. 6: 754. https://doi.org/10.3390/ph15060754
APA StyleVeiga, A. A., Irioda, A. C., Mogharbel, B. F., Bonatto, S. J. R., & Souza, L. M. (2022). Quercetin-Rich Extracts from Onions (Allium cepa) Play Potent Cytotoxicity on Adrenocortical Carcinoma Cell Lines, and Quercetin Induces Important Anticancer Properties. Pharmaceuticals, 15(6), 754. https://doi.org/10.3390/ph15060754