
Assessing the Immunomodulatory Effect of Size on the Uptake and Immunogenicity of Influenza- and Hepatitis B Subunit Vaccines In Vitro

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results
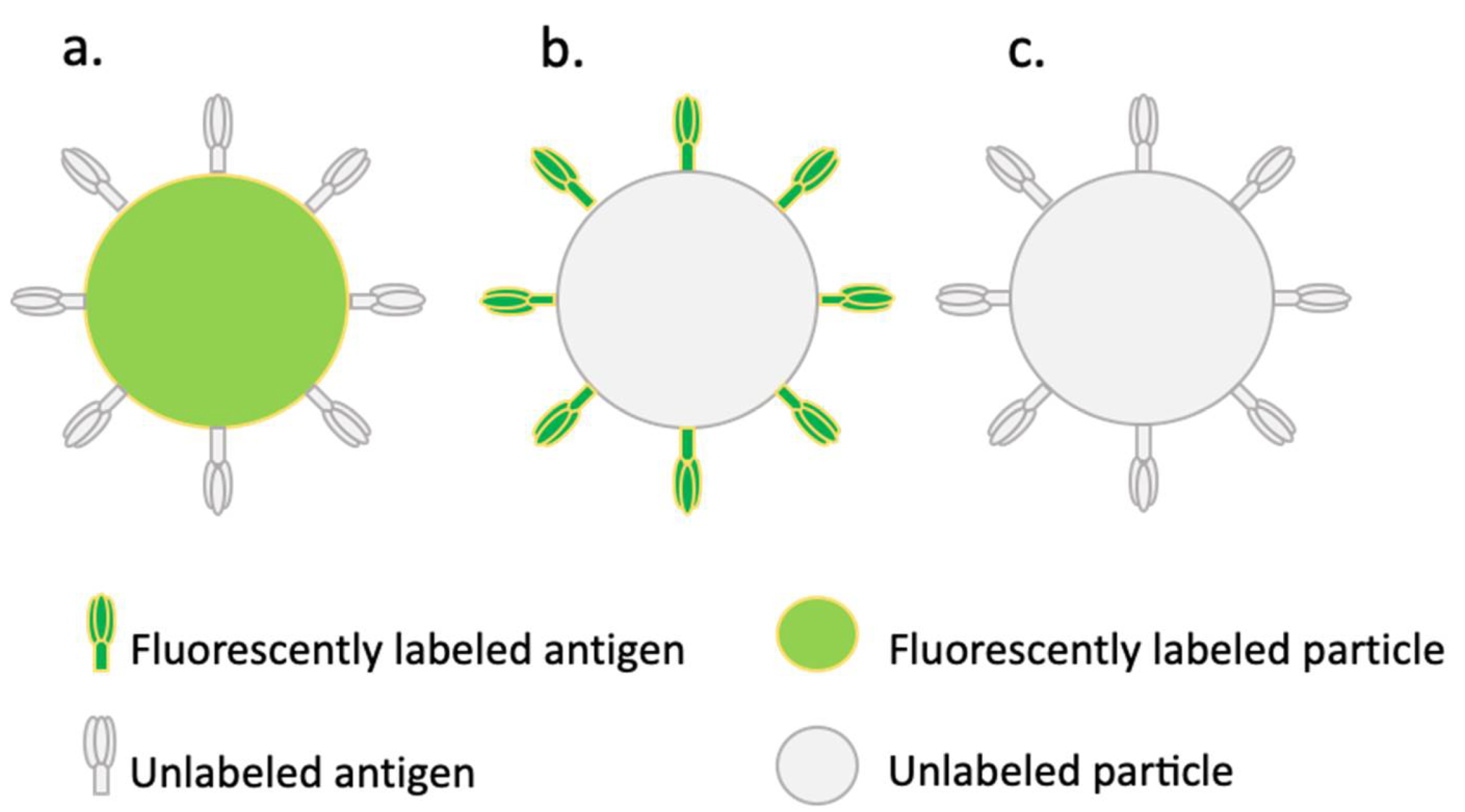
2.1. Conjugates and Conjugation Efficiencies
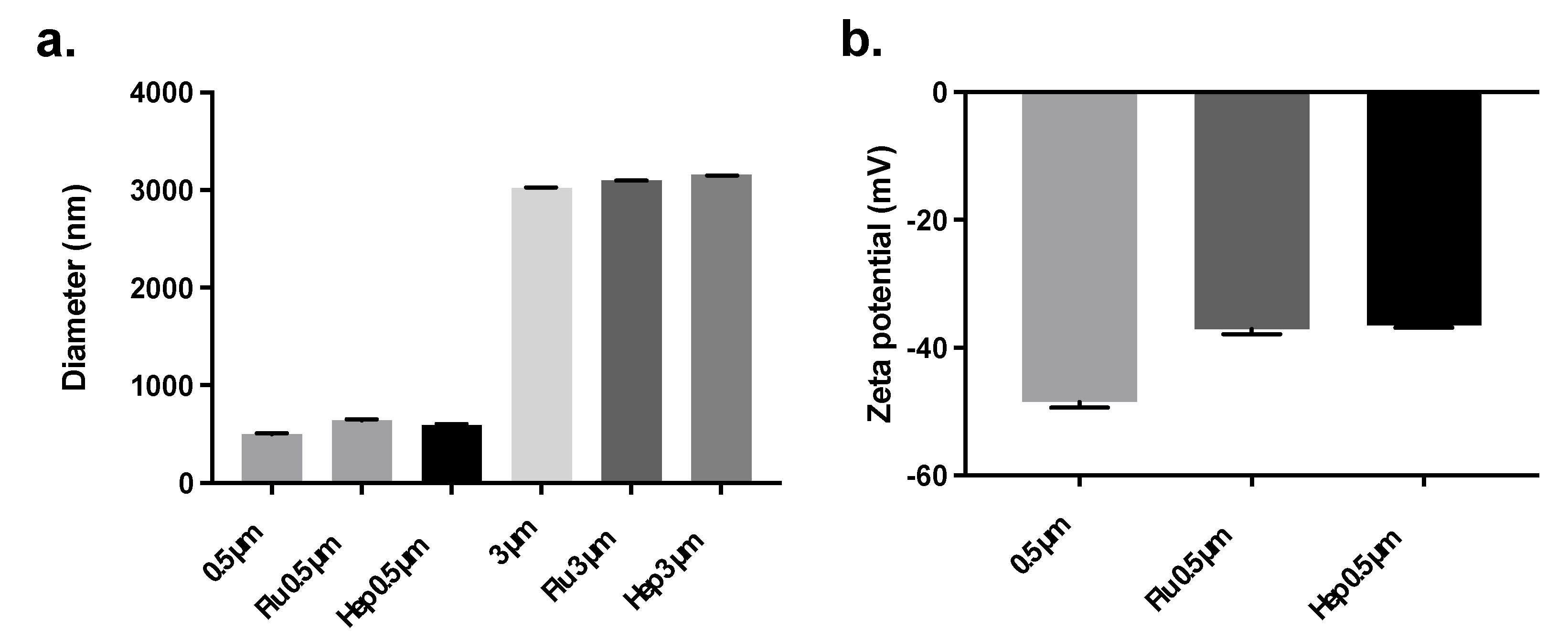
2.2. Conjugate Particle Size and Zeta Potential
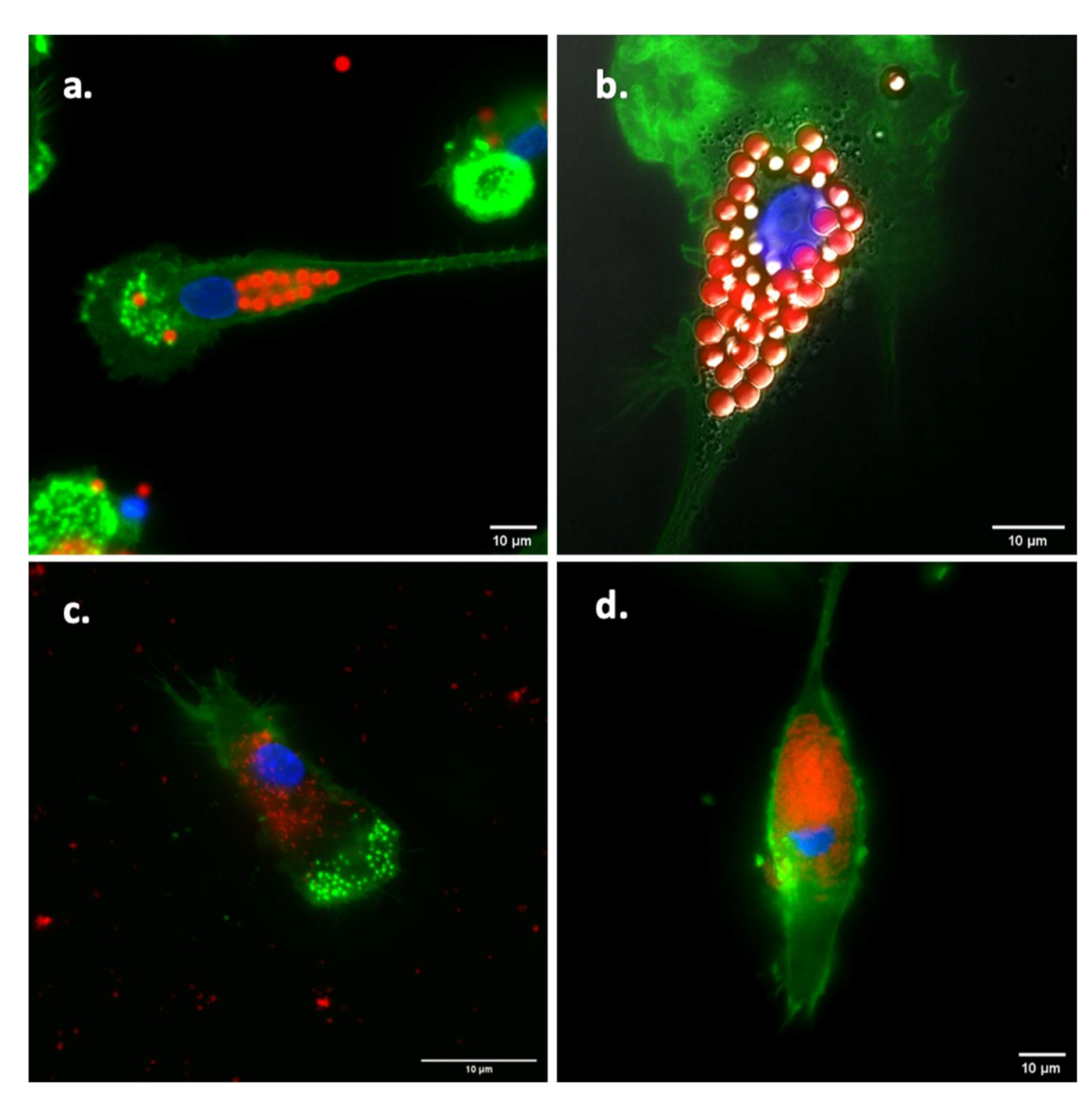
2.3. Visualization of Antigen Uptake
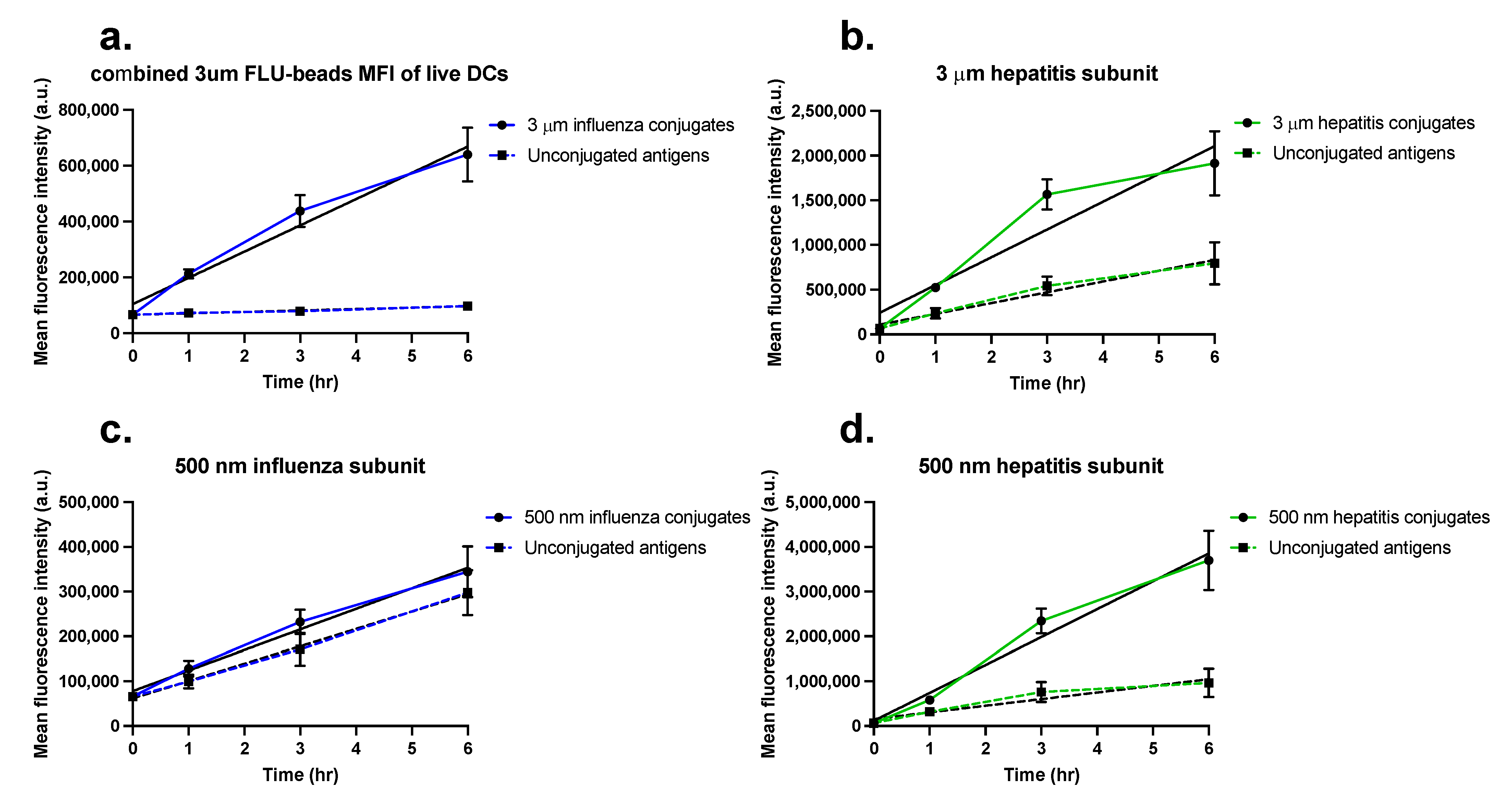
2.4. Quantification of Antigen Uptake in Mo-DCs
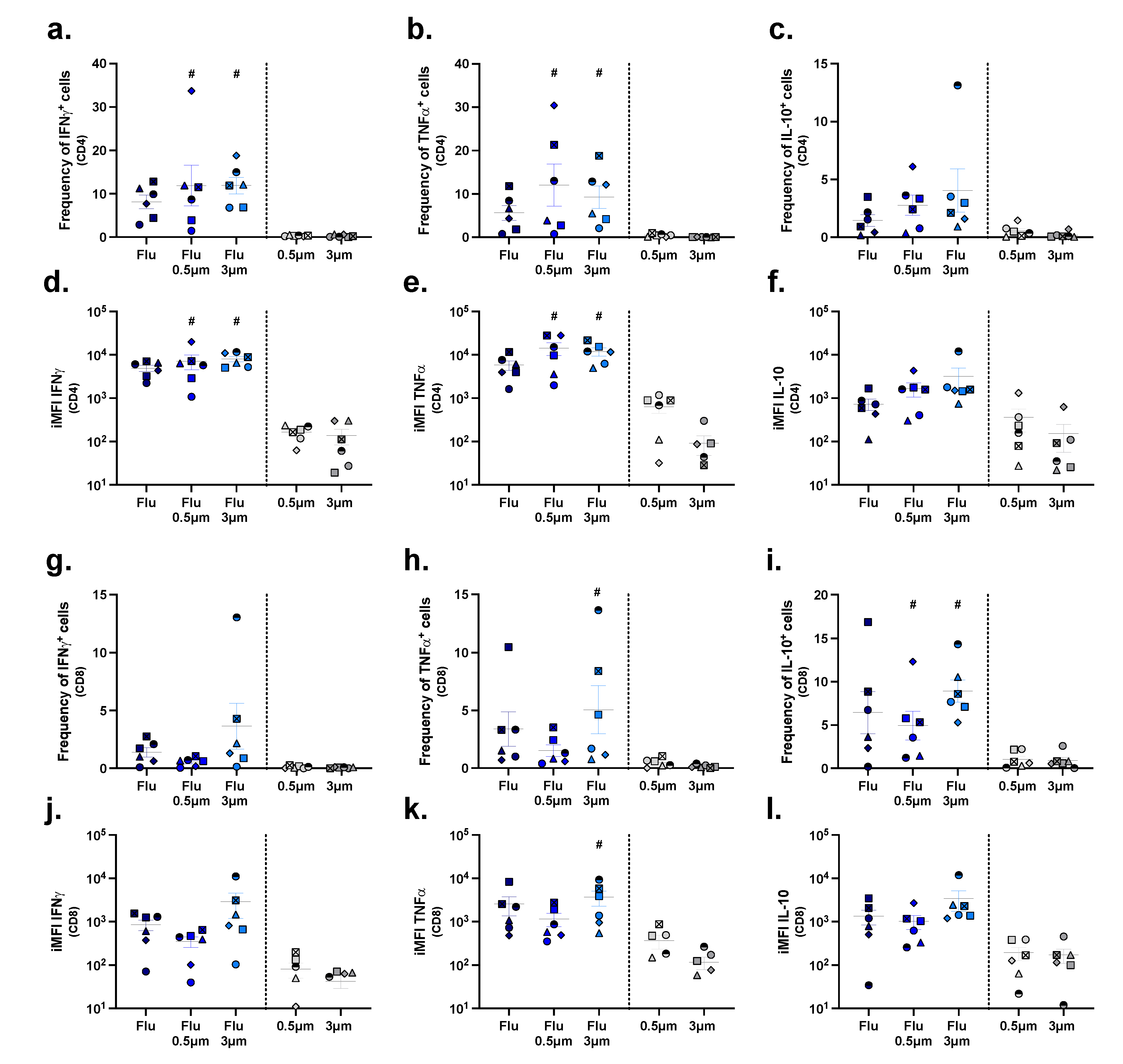
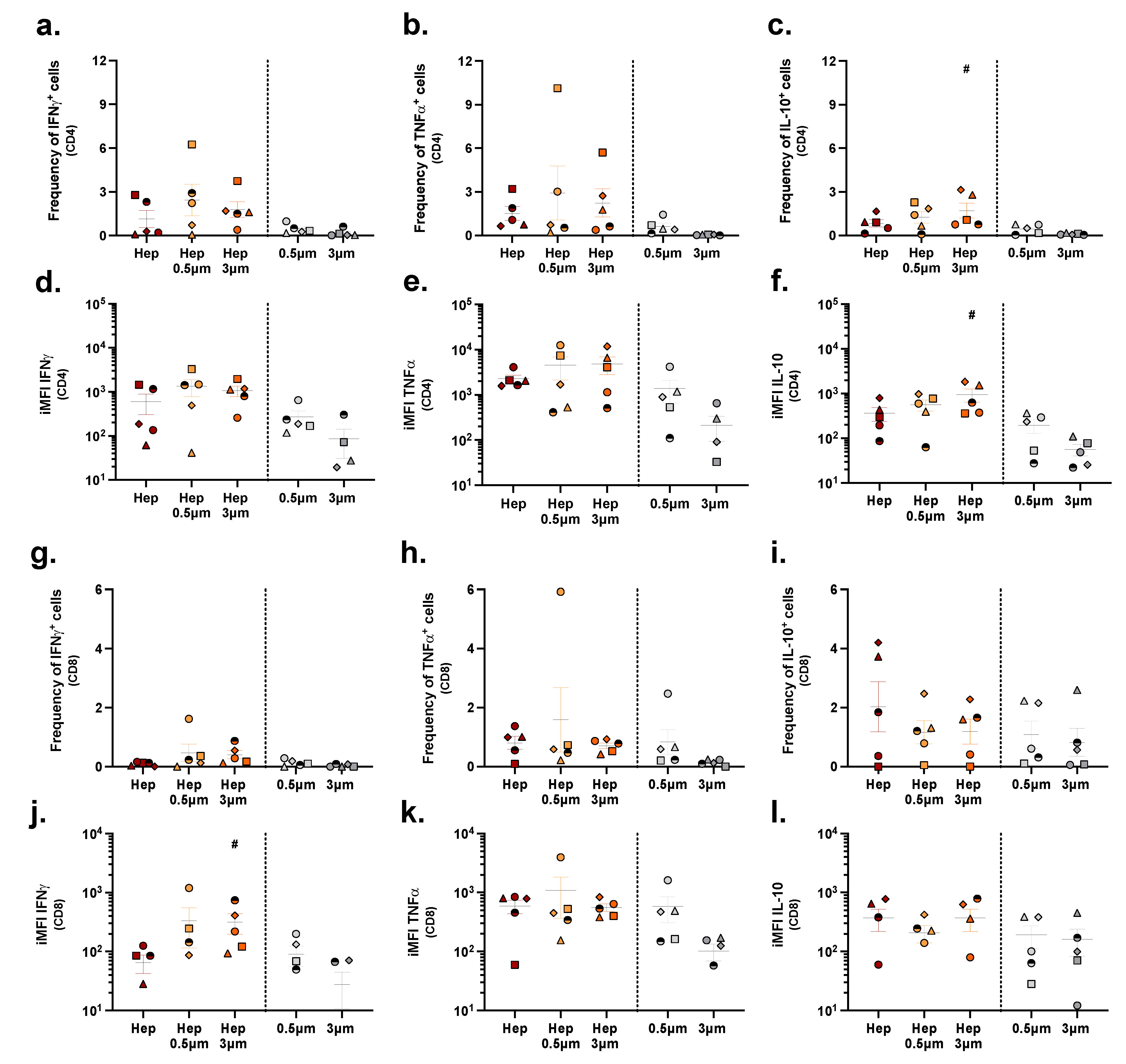
2.5. Assessing T Cell Stimulation in a Population of PBMCs
3. Discussion
4. Materials and Methods
4.1. Influenza Subunit Production
4.2. Conjugation of Antigens to 3 µm and 500 nm Polystyrene Particles
4.2.1. Fluorescent Particles for In Vitro DC Stimulation and Uptake Imaging
4.2.2. Non-Fluorescent Particles for In Vitro PBMC Stimulation
4.2.3. Conjugation of FITC-Labeled Antigens to Non-Fluorescent Particles for In Vitro Uptake Kinetics in Monocyte-Derived Dendritic Cells
4.3. Determination of Size and Zeta Potential of the Conjugates
4.4. Visualization of Antigen Uptake by Monocyte-Derived Dendritic Cells
4.5. Quantification of Antigen Uptake in Monocyte-Derived Dendritic Cells
4.6. Evaluation of the Antigen-Induced T Cell Response
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geeraedts, F.; Goutagny, N.; Hornung, V.; Severa, M.; de Haan, A.; Pool, J.; Wilschut, J.; Fitzgerald, K.A.; Huckriede, A. Superior Immunogenicity of Inactivated Whole Virus H5N1 Influenza Vaccine Is Primarily Controlled by Toll-like Receptor Signalling. PLoS Pathog. 2008, 4, e1000138. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.F.; Jennings, G.T. Vaccine Delivery: A Matter of Size, Geometry, Kinetics and Molecular Patterns. Nat. Rev. Immunol. 2010, 10, 787–796. [Google Scholar] [CrossRef]
- De Temmerman, M.-L.; Rejman, J.; Demeester, J.; Irvine, D.J.; Gander, B.; De Smedt, S.C. Particulate Vaccines: On the Quest for Optimal Delivery and Immune Response. Drug Discov. Today 2011, 16, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Vartak, A.; Sucheck, S. Recent Advances in Subunit Vaccine Carriers. Vaccines 2016, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benne, N.; van Duijn, J.; Kuiper, J.; Jiskoot, W.; Slütter, B. Orchestrating Immune Responses: How Size, Shape and Rigidity Affect the Immunogenicity of Particulate Vaccines. J. Control. Release 2016, 234, 124–134. [Google Scholar] [CrossRef]
- Zhao, L.; Seth, A.; Wibowo, N.; Zhao, C.-X.; Mitter, N.; Yu, C.; Middelberg, A.P.J. Nanoparticle Vaccines. Vaccine 2014, 32, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Tonnis, W.F.; Huckriede, A.L.W.; Hinrichs, W.L.J.; Frijlink, H.W. Pulmonary Administration of Subunit Vaccines; Springer: New York, NY, USA, 2015; pp. 307–329. [Google Scholar]
- Jung, T.; Kamm, W.; Breitenbach, A.; Hungerer, K.D.; Hundt, E.; Kissel, T. Tetanus Toxoid Loaded Nanoparticles from Sulfobutylated Poly(Vinyl Alcohol)-Graft-Poly(Lactide-Co-Glycolide): Evaluation of Antibody Response after Oral and Nasal Application in Mice. Pharm. Res. 2001, 18, 352–360. [Google Scholar] [CrossRef]
- Gutierro, I.; Hernández, R..; Igartua, M.; Gascón, A..; Pedraz, J. Size Dependent Immune Response after Subcutaneous, Oral and Intranasal Administration of BSA Loaded Nanospheres. Vaccine 2002, 21, 67–77. [Google Scholar] [CrossRef]
- Fifis, T.; Gamvrellis, A.; Crimeen-Irwin, B.; Pietersz, G.A.; Li, J.; Mottram, P.L.; McKenzie, I.F.C.; Plebanski, M. Size-Dependent Immunogenicity: Therapeutic and Protective Properties of Nano-Vaccines against Tumors. J. Immunol. 2004, 173, 3148–3154. [Google Scholar] [CrossRef] [Green Version]
- Katare, Y.K.; Muthukumaran, T.; Panda, A.K. Influence of Particle Size, Antigen Load, Dose and Additional Adjuvant on the Immune Response from Antigen Loaded PLA Microparticles. Int. J. Pharm. 2005, 301, 149–160. [Google Scholar] [CrossRef]
- Kanchan, V.; Panda, A.K. Interactions of Antigen-Loaded Polylactide Particles with Macrophages and Their Correlation with the Immune Response. Biomaterials 2007, 28, 5344–5357. [Google Scholar] [CrossRef] [PubMed]
- Wendorf, J.; Chesko, J.; Kazzaz, J.; Ugozzoli, M.; Vajdy, M.; O’Hagan, D.; Singh, M. A Comparison of Anionic Nanoparticles and Microparticles as Vaccine Delivery Systems. Hum. Vaccin. 2008, 4, 44–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blank, F.; Stumbles, P.A.; Seydoux, E.; Holt, P.G.; Fink, A.; Rothen-Rutishauser, B.; Strickland, D.H.; von Garnier, C. Size-Dependent Uptake of Particles by Pulmonary Antigen-Presenting Cell Populations and Trafficking to Regional Lymph Nodes. Am. J. Respir. Cell Mol. Biol. 2013, 49, 67–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seydoux, E.; Rothen-Ruthishauser, B.; Nita, I.; Gazdhar, A.; Petri-Fink, A.; Blank, F.; von Garnier, C.; Balog, S.; Stumbles, P. Size-Dependent Accumulation of Particles in Lysosomes Modulates Dendritic Cell Function through Impaired Antigen Degradation. Int. J. Nanomed. 2014, 9, 3885. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, G.; BenMohamed, L. Of Mice and Not Humans: How Reliable Are Animal Models for Evaluation of Herpes CD8+-T Cell-Epitopes-Based Immunotherapeutic Vaccine Candidates? Vaccine 2011, 29, 5824–5836. [Google Scholar] [CrossRef] [Green Version]
- Barré-Sinoussi, F.; Montagutelli, X. Animal Models Are Essential to Biological Research: Issues and Perspectives. Futur. Sci. OA 2015, 1, FSO63. [Google Scholar] [CrossRef] [Green Version]
- Van der Worp, B.H.; Howells, D.W.; Sena, E.S.; Porritt, M.J.; Rewell, S.; O’Collins, V.; Macleod, M.R. Can Animal Models of Disease Reliably Inform Human Studies? PLoS Med. 2010, 7, e1000245. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Chen, S.; Eisenbarth, S.C. Dendritic Cell Regulation of T Helper Cells. Annu. Rev. Immunol. 2021, 39, 759–790. [Google Scholar] [CrossRef]
- Thiele, L.; Rothen-Rutishauser, B.; Jilek, S.; Wunderli-Allenspach, H.; Merkle, H.P.; Walter, E. Evaluation of Particle Uptake in Human Blood Monocyte-Derived Cells in Vitro. Does Phagocytosis Activity of Dendritic Cells Measure up with Macrophages? J. Control. Release 2001, 76, 59–71. [Google Scholar] [CrossRef]
- Cruz, L.J.; Tacken, P.J.; Fokkink, R.; Joosten, B.; Stuart, M.C.; Albericio, F.; Torensma, R.; Figdor, C.G. Targeted PLGA Nano- but Not Microparticles Specifically Deliver Antigen to Human Dendritic Cells via DC-SIGN in Vitro. J. Control. Release 2010, 144, 118–126. [Google Scholar] [CrossRef]
- Foged, C.; Brodin, B.; Frokjaer, S.; Sundblad, A. Particle Size and Surface Charge Affect Particle Uptake by Human Dendritic Cells in an in Vitro Model. Int. J. Pharm. 2005, 298, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Kadu, P.J.; Kushare, S.S.; Thacker, D.D.; Gattani, S.G. Enhancement of Oral Bioavailability of Atorvastatin Calcium by Self-Emulsifying Drug Delivery Systems (SEDDS). Pharm. Dev. Technol. 2011, 16, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Calle, G.; Born, P.A.; Koutsoumpli, G.; Gonzalez-Rodriguez, M.I.; Hinrichs, W.L.J.; Huckriede, A.L.W. A PBMC-Based System to Assess Human T Cell Responses to Influenza Vaccine Candidates In Vitro. Vaccines 2019, 7, 181. [Google Scholar] [CrossRef] [Green Version]
- Metz, D.P.; Farber, D.L.; Taylor, T.; Bottomly, K. Differential Role of CTLA-4 in Regulation of Resting Memory Versus Naive CD4 T Cell Activation. J. Immunol. 1998, 161, 5855–5861. [Google Scholar] [PubMed]
- Slütter, B.; Jiskoot, W. Sizing the Optimal Dimensions of a Vaccine Delivery System: A Particulate Matter. Expert Opin. Drug Deliv. 2016, 13, 167–170. [Google Scholar] [CrossRef]
- Champion, J.A.; Walker, A.; Mitragotri, S. Role of Particle Size in Phagocytosis of Polymeric Microspheres. Pharm. Res. 2008, 25, 1815–1821. [Google Scholar] [CrossRef] [Green Version]
- Thiele, L.; Merkle, H.P.; Walter, E. Phagocytosis and Phagosomal Fate of Surface-Modified Microparticles in Dendritic Cells and Macrophages. Pharm. Res. 2003, 20, 221–228. [Google Scholar] [CrossRef]
- Salatin, S.; Yari Khosroushahi, A. Overviews on the Cellular Uptake Mechanism of Polysaccharide Colloidal Nanoparticles. J. Cell. Mol. Med. 2017, 21, 1668–1686. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Anselmo, A.C.; Banerjee, A.; Zakrewsky, M.; Mitragotri, S. Shape and Size-Dependent Immune Response to Antigen-Carrying Nanoparticles. J. Control. Release 2015, 220, 141–148. [Google Scholar] [CrossRef]
- Peterson, G.L. A Simplification of the Protein Assay Method of Lowry et Al. Which Is More Generally Applicable. Anal. Biochem. 1977, 83, 346–356. [Google Scholar] [CrossRef]
- Khramtsov, P.; Kalashnikova, T.; Bochkova, M.; Kropaneva, M.; Timganova, V.; Zamorina, S.; Rayev, M. Measuring the Concentration of Protein Nanoparticles Synthesized by Desolvation Method: Comparison of Bradford Assay, BCA Assay, Hydrolysis/UV Spectroscopy and Gravimetric Analysis. Int. J. Pharm. 2021, 599, 120422. [Google Scholar] [CrossRef] [PubMed]
- Yap, W.T.; Song, W.K.; Chauhan, N.; Scalise, P.N.; Agarwal, R.; Miller, S.D.; Shea, L.D. Quantification of Particle-Conjugated or Particle-Encapsulated Peptides on Interfering Reagent Backgrounds. Biotechniques 2014, 57, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Shukla, A.K.; Bagga, E.; Kumari, S.; Bajpai, R.P.; Bharadwaj, L.M. 1-Ethyl-3-(3-Dimethylaminopropyl) Carbodiimide Interference with Lowry Method. Anal. Biochem. 2005, 336, 132–134. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapia-Calle, G.; Stoel, M.; Huckriede, A. Distinctive Responses in an In Vitro Human Dendritic Cell-Based System upon Stimulation with Different Influenza Vaccine Formulations. Vaccines 2017, 5, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleiveland, C.R. Peripheral Blood Mononuclear Cells. In The Impact of Food Bioactives on Health; Springer International Publishing: Cham, Switzerland, 2015; pp. 161–167. ISBN 9783319161044. [Google Scholar]
- Salvati, A.; Nelissen, I.; Haase, A.; Åberg, C.; Moya, S.; Jacobs, A.; Alnasser, F.; Bewersdorff, T.; Deville, S.; Luch, A.; et al. Quantitative Measurement of Nanoparticle Uptake by Flow Cytometry Illustrated by an Interlaboratory Comparison of the Uptake of Labelled Polystyrene Nanoparticles. NanoImpact 2018, 9, 42–50. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unlabeled 500 nm Particles | Unlabeled 3 µm Particles | Fluorescently Labeled 500 nm Particles | Fluorescently Labeled 3 µm Particles | |
|---|---|---|---|---|
| Unlabeled Influenza antigen | 1.25 × 10−9 µg/µm2 | 6.90 × 10−9 µg/µm2 | 1.29 × 10−9 µg/µm2 | 4.16 × 10−9 µg/µm2 |
| Unlabeled HBsAg | 0.926 × 10−9 µg/µm2 | 4.69 × 10−9 µg/µm2 | - | - |
| Fluorescently Labeled influenza antigen | 0.410 × 10−9 µg/µm2 | 0.196 × 10−9 µg/µm2 | - | - |
| Fluorescently Labeled HBsAg | 1.31 × 10−9 µg/µm2 | 6.91 × 10−9 µg/µm2 | - | - |
| Conjugate Type | Slope of Uptake | Fold-Change in Slope Compared to Free Antigen |
|---|---|---|
| 3 µm-influenza antigen | 94,027 | 18.4 |
| 500 nm-influenza antigen | 45,899 | 1.2 |
| 3 µm-HBsAg | 310,806 | 2.6 |
| 500 nm-HBsAg | 622,956 | 4.2 |
| Conjugate Type | Number of Particlesper Well | Amount of Corresponding Antigen per Well |
|---|---|---|
| 3 µm-influenza antigen | 20 µg | 6.4 ng |
| 500 nm-influenza antigen | 20 µg | 93.8 ng |
| 3 µm-HBsAg | 20 µg | 248.6 ng |
| 500 nm-HBsAg | 20 µg | 298.7 ng |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heida, R.; Born, P.A.; Tapia-Calle, G.; Frijlink, H.W.; Salvati, A.; Huckriede, A.L.W.; Hinrichs, W.L.J. Assessing the Immunomodulatory Effect of Size on the Uptake and Immunogenicity of Influenza- and Hepatitis B Subunit Vaccines In Vitro. Pharmaceuticals 2022, 15, 887. https://doi.org/10.3390/ph15070887
Heida R, Born PA, Tapia-Calle G, Frijlink HW, Salvati A, Huckriede ALW, Hinrichs WLJ. Assessing the Immunomodulatory Effect of Size on the Uptake and Immunogenicity of Influenza- and Hepatitis B Subunit Vaccines In Vitro. Pharmaceuticals. 2022; 15(7):887. https://doi.org/10.3390/ph15070887
Chicago/Turabian StyleHeida, Rick, Philip A. Born, Gabriela Tapia-Calle, Henderik W. Frijlink, Anna Salvati, Anke L. W. Huckriede, and Wouter L. J. Hinrichs. 2022. "Assessing the Immunomodulatory Effect of Size on the Uptake and Immunogenicity of Influenza- and Hepatitis B Subunit Vaccines In Vitro" Pharmaceuticals 15, no. 7: 887. https://doi.org/10.3390/ph15070887
APA StyleHeida, R., Born, P. A., Tapia-Calle, G., Frijlink, H. W., Salvati, A., Huckriede, A. L. W., & Hinrichs, W. L. J. (2022). Assessing the Immunomodulatory Effect of Size on the Uptake and Immunogenicity of Influenza- and Hepatitis B Subunit Vaccines In Vitro. Pharmaceuticals, 15(7), 887. https://doi.org/10.3390/ph15070887